|
LAS MOJARRAS DE LA
LAGUNA DE TÉRMINOS: TAXONOMÍA, BIOLOGÍA ECOLOGÍA Y
DINÁMICA TRÓFICA. (PISCES: GERREIDAE)
Trabajo presentado
parcialmente en el VII Congreso Nacional de Zoología, 4-10 de diciembre
de 1983. Recibido el 15 de mayo de 1984 y aceptado para su publicación
el 20 de octubre de 1984.
ARTURO AGUIRRE-LEÓN
ALEJANDRO YÁÑEZ-ARANCIBIA
Universidad Nacional Autónoma de México, Instituto
de Ciencias del Mar y Limnología, Laboratorio de Ictiología y
Ecología Estuarina Contribución 440 del Instituto de Ciencias del
Mar y Limnología, UNAM. Proyecto UNAM-CONACYT,
PCMABEU-005322.
Las mojarras de la Familia Gerreidae
constituyen un recurso abundante con importancia comercial y amplia
distribución en el medio ambiente lagunar-estuarino de latitudes
tropicales y subtropicales del mundo y, por lo tanto, muy significativo en
lagunas costeras y estuarios de México. Existen siete especies de esta
familia en la Laguna de Términos i. e. Gerres cinereus,
Eucinostomus gula, Eucinostomus argenteus, Eucinostomus melanopterus, Diapterus
rhombeus, Diapterus auratus y Eugerres plumieri. De estas especies,
Eucinostomus gula comprende el 83% y el 48% de las capturas
totales en peso y número respectivamente de todas las mojarras. Todas
las especies presentan patrones de distribución y abundancia propios,
correspondientes a los subsistemas ecológicos del litoral interno de la
Isla del Carmen con áreas protegidas deRhizophora mangle y
Thalassia testudinum, en el subsistema oligohalino de la cuenca
central de la laguna y en los subsistemas fluvio-lagunares asociados cón
ambientes de arrecifes deCrassostrea virginica y
vegetación sumergida de origen dulceacuícola. Por su diversidad,
distribución y abundancia, son componentes típicos y juegan un
papel muy importante en la estructura y función de las ictiocomunidades
del área. Las especies mejor representadas en abundancia numérica
son en orden de importancia: Eucinostomus gula, Diapterus rhombeus,
Eucinostomus argenteus y Eugerres plumieri. Además, las dos
primeras son típicas por biomasa y frecuencia. La introducción
general a la biología y ecología de las poblaciones de mojarras
se inicia con una síntesis taxonómica. Se elabora una diagnosis
para la familia, así como una clave ilustrada para determinar
taxonómicamente géneros y especies, proponiendo caracteres
diagnósticos sobre la base de una revisión de rangos
merístícos, morfométricos y coloración. Los
patrones ecológicos que controlan a las poblaciones en edad, talla y
madurez gonádica, son en mayor medida, la época del año,
la localidad dentro del sistema (habitats) y la disponibilidad de alimento,
presentándose diversos patrones de frecuencia y utilización de la
laguna. Tres especies se comportan como visitantes ocasionales: Gerres
cinereus, Eucinostomus melanpterus y Diapterus auratus, penetran a la
laguna para protegerse y/o alimentarse. Como componentes cíclicos o
estacionales se encuentran a Eucinostomus argenteus, Diapterus
rhombeus y Eugerres plumieri que utilizan la laguna como área
de crianza y alimentación. Eucinostomus gula es una
especie residente permanente en el área con registros de ictioplancton
en primavera y verano. Las poblaciones están constituidas principalmente
por juveniles en fases tempranas de maduración gonádica, con poca
representatividad de individuos adultos y maduros a lo largo del año. En
general, para todas las especies existe mayor proporción de machos que
de hembras. Presentan patrones estacionales de migración hacia la Sonda
de Campeche que obedecen a estrategias reproductivas. La mayoría de las
especies, alcanzan la madurez sexual después de los 120 mm de longitud
total. El espectro trófico de las especies es muy amplio y diverso,
considerándoseles consumidores de primer orden, incorporando a su dieta
principalmente pequeños invertebrados bentónicos e importantes
cantidades de destritus. Presentan variaciones particulares en sus dietas
principal, secundaria y ocasional de acuerdo a la edad, la época
climática la localidad y la disponibilidad de alimento en la laguna. Los
invertebrados mas comunes la localidad en su contenido estomacal son:
foraminíferos, nemátodos, poliquetos, oligoquetos,
ostrácodos, copépodos, cumáceos, tanaidáceos,
anfípodos, isópodos, pequeños decápodos,
pelecípodos, gasterópodos y sinpuncúlidos. La importancia
biológica, ecológica y económica de Eucinostomus
gula, Diapterus rhombeus y Eugerres plumieri determina que sean
estudiadas de manera más amplia.
SUMMARY
Mojarras (Gerreidae)
make up an abundant resource that is comercially important and widely
distributed in the lagoon-estuarine environment throughout the worl's tropical
and subtropical latitudes. Therefore they are very well-represented in Mexico's
coastal lagoons and estuaries. There are seven species oh this family in
Terminos Lagoon i.e. Gerres cinereus, Eucinostomus gula, Eucinostomus
argenteus, Eucinostomus melanopterus, Diapterus auratus and Eugerres
plumieri. Of these species, Eucinostomus
gulacomprises 83 % and 48 % of mojarra catch totals by weight and
number, respectively. All the species have their own distribution and abundance
patterns. These correspond to ecological subsystems in the internal littoral of
Carmen Island that has protected areas of Rhizophora mangle and
Thalassia testudinum, in the oligohaline subsystem found in the
laggon's central basin, and the fluvial-lagoon subsystem associated
withCrassostrea virginica reef environments and sumerged
vegetation of freshwater origen. Because of their diversity, distribution, and
abundance they are typical components and play an important role in t he
structure and function fish community. The best represented species by
numerical abundance are, in order of importance: Eucinostomus gula,
Diapterus rhombeus, Eucinostomus argenteus and Eugerres plumieri. In
addition, the first two are typical for biomass and frequency. The general
introduction, the first two are typical for biomass and frequency. The general
introduction to mojarra population biology and ecology begins with a taxonomic
synthesis. It elaborates a diagnosis for family as well as an illustrated key
to determine taxonomically, genus and species, proposing diagnostic
characteristics based on a review of meristic, morphometric, and coloration
ranges. The ecological patters that control the populations age, size, and
gonad maturity are, for the most part, the season for the year, the locality
within the system (habitat), and the availability of food. There are various
frequency and use patterns within the lagoon. Three species show the behavior
of occasional visitors: Gerres cinereus, Eucinostomus melanopterus and
Diapterus auratus enter the lagoon for protection and/or to feed. The
cyclical or season al components of the system are: Eucinostomus
argenteus, Diapterus rhombeus and Eugerres plumieri that use the
lagoon as a growing arid feeding area. Eucinostomus gula is
a permanent resident species in the area that appears in the ichthyoplankton
during the summer and the spring. The populations are made up mainly of
juveniles in the early stages of gonad maturity. Adult and mature fish are
poorly represented throughout the year. Generally, all the species have a
greater proportion of males than females. They show seasonal migration patterns
toward Campeche Sound that follow reproductive strategies. The majority of the
species achieve sexual maturity after reaching a total length of 120 mm. The
trophic spectrum is wide and diverse. We consider the first order consumer
species as those that have a diet consisting mainly of small benthonic
invertebrates and large quantities of detritus. They show certain variations in
their main, secondary and occasional diet according to their age, the climatic
conditions, the locality, and the availability on food in the lagoon. The
invertebrates most commoly found in stomach content are: foraminifers,
nematodes, polichaetes, oligochaetes, isopods, small decapods, pelicipods,
gasteropods and sipunculids. The biological, ecological, and economic
importance of Eucinostomus gula, Diapterus rhombeus and Eugerres
plumieri emphasizes the need for more studies of them.
Las investigaciones orientadas hacia el conocimiento de la ecología de la zona costera en los litorales de México han alcanzado actualmente un nivel de prioridad, debido a que constituyen una importante área de producción de energía, alimentos y recursos no renovables, entre otros. En el Sur del Golfo de México, estudios recientes sobre la ecología de sistemas y evaluación de sus recursos han definido a la región como prioritaria en investigaciones ecológicas debido a: 1) la gran diversidad de especies biológicas y de hábitats, 2) los recursos pesqueros, 3) las interacciones ecológicas entre la Laguna de Términos y su Plataforma Continental adyacente, 4) la expansión del desarrollo industrial costero de la región (puerto, petróleo y pesca principalmente), 5) por no presentarse todavía niveles críticos de contaminación, 6) la importante información científica que se incrementa para la región, 7) la probabilidad de establecer en el área santuarios de flora y fauna y 8) por tratarse de un ecosistema costero tropical natural de alta complejidad, lo cual permite definir a la región como un área "tipo" a nivel mundial (Yáñez-Arancibia y Day, 1982, Yáñez-Arancibia y Sánchez-Gil, 1983, y Yáñez-Arancibia et al.,1983 a). La comprensión ecológica de este ecosistema requiere necesariamente del conocimiento taxonómico preciso de las especies de peces, así como de su ecología y biología. La obtención de esta información ha sido fundamental para establecer que ciertas familias de peces dominantes entre ellas la familia Gerreidae- es un grupo de peces diverso, abundante y ampliamente distribuido a lo largo del año en la Laguna de Términos. Esto incide directamente en la trascendencia que las mojarras pueden desarrollar en el ecosistema por su papel en la transformación de energía, la conducción de ella, el intercambio con ecosistemas vecinos, el almacenamiento de la misma, la regulación y su probable función en el recielamiento de nutrientes, como ha sido primariamente planteado como hipótesis por Yáñez-Arancibia y Nugent (1977) y más tarde probado para diversas especies por Deegan y Thompson (1985). Estudios específicos sobre la familia Gerreidae en la Laguna de Términos muestran resultados preliminares sobre la importancia de las mojarras como recurso alimenticio (Aguirre León et al., 1982). Actualmente, dentro del renglón comercial de peces, las mojarras ocupan un lugar importante en el consumo local y nacional, por lo cual, el análisis de la distribución de frecuencia de tallas de las especies y la aplicación de los modelos de las regresiones lineales en la relación peso-longitud, establecerán un aspecto de la estructura poblacional de las especies, lo cual deberá cotejarse con la proporción de sexos y el estado de desarrollo de las glándulas sexuales, lo que permite precisar las tallas mínimas de madurez gonádica. El muestreo cuantitativo de las poblaciones, proporciona información estimativa de la abundancia de los recursos, el patrón de utilización del área y los factores que controlan y regulan su distribución. Para estimar la productividad biológica del necton es fundamental conocer el comportamiento de las comunidades; como también conocer los hábitos alimenticios de las especies y, dentro de la estructura trófica, de qué manera la materia orgánica se encuentra disponible para los consumidores. La estructura trófica indica un aspecto del flujo de la energía, muestra las relaciones entre predadorpresa y productor-consumidor, e indica las relaciones ecológicas de los organismos, con lo que se interpreta mejor la dinámica general estuarina y permite sugerir recomendaciones para la administración adecuada de los recursos pesqueros. El análisis e interpretación de la biología y ecología de las especies involucradas, permitirá establecer cuantitativamente las características de alimentación, relaciones tróficas, crecimiento, madurez, patrones de distribución, relaciones ecológicas y su papel en la estructura y función de las comunidades de peces de la Laguna de Términos. Asimismo, modelos futuros de evaluación, manejo y protección de estos recursos alimenticios del Sur del Golfo de México, podrán utilizar estos datos para fundamentar toma de decisiones. OBJETIVOSLos objetivos principales se puntualizan a continuación: 1. Elaborar diagnosis taxonómicas para familia y especies, estructurando una clave artificial ilustrada para su identificación. 2.Caracterizar los patrones de distribución, abundancia y frecuencia de las especies, de acuerdo a las épocas climáticas definidas para el área y a las características de los hábitats de la localidad. 3.Determinación de un modelo de regresión lineal de las especies para analizar la relación peso-longitud, considerando además la distribución de frecuencias de tallas en cielos anuales. 4. Analizar aspectos de madurez gonádica y distribución de fases sexuales para conocer probables áreas de crianza y reproducción así como también las tallas mínimas de madurez. 5. Determinar el espectro trófico de las especies sobre la base de su alimentación y hábitos alimenticios y discutir cuantitativamente su ecología trofodinámica. 6. Establecer el comportamiento de las especies y el patrón de utilización de la laguna como áreas de crianza, asi como establecer preliminarmente el papel ecológico de las especies en la estructura y función de las ictiocomunidades del área. 7. Analizar, interpretar e integrar esta información para establecer un marco de referencia que permita reorientar investigaciones futuras, sugerir recomendaciones, y contribuir a la evaluación, manejo y protección de estos recursos alimenticios del Sur del Golfo de México. ANTECEDENTESLos estudios ictiológicos realizados en la Laguna de Términos y áreas adyacentes han analizado la complejidad ambiental, estructural y funcional de las especies y poblaciones de peces a nivel regional (Toral y Reséndez, 1973; Carbajal, 1975; BravoNúñez y Yáñez-Arancibia, 1979; Yáñez-Arancibia y Amezcua Linares, 1979; Amezcua Linares y Yáñez-Arancibia, 1980; Reséndez, 1981; Sánchez-Gilet al., 1981; Lara Domínguez et al., 1981; Vargas Maldonado et al., 1981; Mallard Colmenero et al., 1982; Aguirre León et al., 1982; Chavance et al., 1982a y 1982b; Yáñez-Arancibia et al., 1980, 1981, 1982b, 1982c, 1982d, 1983a, 1985a, 1985b, 1985c y 1985d; Yáñez-Arancibia y Lara Domínguez, 1983; Yáñez-Arancibia y Sánchez-Gil, 1983 y Alvarez Guillén et al., 1984). Sobre la familia Gerreidae, los estudios ecológicos específicos realizados a la fecha en las costas de México siguen siendo escasos, aun cuando existe un apreciable grado de avance (CastroAguirre, 1978; Yáñez-Arancibia, 1978 y Aguirre León et al.,1982). Por otra parte la Secretaría de Pesca, a través de la Dirección de Informática y Estadística han proporciondo datos de importantes capturas comerciales de mojarras en las costas de Campeche y Tabasco. Sin embargo, dentro de estas capturas están consideradas varias especies de gerreidos, así como también lo están otras especies de varias familias que se les llama también mojarras. En Campeche, se han registrado siete especies de mojarras en la Laguna de Términos, i.e., Gerres cinereus, Eucinostomus gula, Eucinostomus argenteus, Eucinostomus melanopterus, Diapterus rhombeus, Diapterus auratus y Eugerres plumieri. Eucinostomus gula es la especie mejor representada en número, biomasa y distribución en la Laguna. Eugerres plumieri es a su vez un recurso económico en el área (Aguirre León et al., 1982). En el Golfo de México se encuentran además otras especies de gérreidos, i. e. Ulaema lefroyi(Goode), Eugerres brasilianus (Cuvier) y Eugerres mexicanus (Steindachner). Debe destacarse que todas estas especies han sido consideradas con frecuencia componentes importantes de las comunidades ictiofaunísticas por su distribución y abundancia en las zonas costeras del Golfo y Mar Caribe, así como también las especies analizadas en este estudio (Springer y Woodburn, 1960; Springer y Mc Erlean, 1962; Cervigón, 1966; Randall, 1967; Beebe y Tee-Van, 1970; Walls, 1975; Hoese y Moore, 1977; CastroAguirre, 1978 y Darnell et al., 1983). A su vez se considera que los siguientes trabajos son referencias comparativas importantes sobre antecedentes biológicos y ecológicos de las mojarras. Waldinger (1968) estudia la relación entre los parámetros ambientales y las capturas de Eucinostomus gula, Eucinostomus argenteus y Eugerres plumierien el Canal de Buttonwood, en el Parque Nacional Everglade, Florida. Austin (1971), analiza el cielo biológico de Diapterus rhombeus en aguas costeras de Puerto Rico. Aspectos parciales sobre la alimentación y hábitos alimenticios de Gerres cinereus, Eucinostomus gula, Eucinostomus argenteus y Eugerres plumieri han sido estudiados en lagunas costeras y estuarios del norte del Golfo de México por Odum y Heald (1972), Carr y Adams (1973), Brook (1977). A su vez Randall (1967) estudió los mismos aspectos en Gerres cinereus y Eucinostomus gula en el Mar Caribe. Charles (1975) estudia la biología de Eucinostomus gula en la Bahía de Biscayne, Florida. Yáñez-Arancibia (1978) en un trabajo amplio de las lagunas costeras del Pacífico Central de México, estudia aGerres cinereus, Eucinostomus currani Zahuranec, Eucinostomus entomelas Zahuranec, Eucinostomus dowii (Gill), Diapterus peruviansus (Cuvier y Valenciennes), Eugerres lineatus (Humboldt y Valenciennes) y Eugerres axillaris (Günther), desde el punto de vista taxonómico, generalidades biológicas y ecología trofodinámica. De igual manera, en aguas costeras del IndoPacífico Prabhakaro Rao (1968) ha estudiado a Gerres oblongus Cuvier, Gerres setifer (Hamilton) y Gerres abbreviatus Cuvier. Asimismo, Cyrus y Blaber(1982a, 1982b, y 1983) estudian a Gerres filamentosus Cuvier, Gerres oyena, Gerres poieti Cuvier, Gerres acinaces Bleecker, Gerres rappi (Barnard) y Gerres oblongus Cuvier. Rubio (1975) estudia la biología de Eugerres plumieri en la Ciénega de Santa Marta en Colombia. Rodríguez Castellanos (1983), analiza aspectos parciales de la biología de los gerreidos de la Ciénega de la Virgen en Colombia. González Sansón y Rodríguez Viñas (1983) estudian la alimentación deEugerres brasilianus y Gerres cinereus en lagunas costeras de Tunas de Zaza en Cuba. Otro tipo de estudios en relación a gerreidos es el realizado por Baez Hidalgo y Guevara Carrió (1983) quienes analizan relaciones morfométricas y merísticas deEugerres brasilanus y las comparan con las deEugerres plumierien Lagunas de Cuba. Fundamentalmente los avances de las investigaciones en la Laguna de Términos y la Sonda de Campeche están estableciendo, que los procesos físicos controlan la ecología de la laguna y consecuentemente los procesos biológicos de los recursos pesqueros. La información sistematizada que se está obteniendo permitirá: 1) detectar los cambios que se producen en forma natural como también los cambios que pueden provocar la contaminación, 2) evaluar el potencial de los recursos y, 3) aportar información para predecir y tomar decisiones de los usos y recursos de la zona costera en la región (Yáñez-Arancibia et al.,1982d). ÁREA DE ESTUDIOLa Laguna de Términos se localiza en el Sur del Golfo de México frente a la Sonda de Campeche (Fig. l). Es un área somera y amplia, con un promedio de 3.5 m de profundidad y 2,500 km² de superficie. El clima es tropical húmedo con un rango de precipitación anual de 1,100 a 2,000 mm. El rango de mareas es de 0.5 a 0.7 m. Dos bocas conectan a la Laguna con el mar y existe un fuerte flujo de agua hacia el oeste causado por los vientos predominantes del este. Por este patrón de circulación se presenta un gradiente semipermanente de salinidad, turbidez, niveles de nutrientes, tipos de sedimentos, asociaciones de foraminíferos y macrobentos y migración de peces y camarones. Existen tres épocas climáticas en el área: la época de lluvias de junio a septiembre, la época de nortes de octubre a noviembre, y la época de secas de febrero a mayo. La parte sur de la laguna recibe más del 50 % del aporte de agua dulce. Existe gran diversidad de habitat estuarinos, incluyendo pantanos de manglar salobres y de baja salinidad, pastos marinos. pastos de pantano, áreas de alta sedimentación y arrecifes de ostión, y la cuenca central oligohalina. La sahnidad es elevada al norte y noreste de la laguna durante la época de secas. Poblaciones bénticas definidas se restringen a los diferentes hábitat en la laguna y están controladas por la salinidad, aporte fluvial, turbidez y tipo de sedimento. Amplios detalles de esta síntesis pueden encontrarse en los trabajos siguientes: Yáñez-Arancibia y Day (1982) y Yáñez-Arancibia et al. (1983 a).  Fig. 1. Localización de la Laguna de Términos, Campeche. Se señalan los rasgos geográficos y toponímicos principales y la distribución de los diferentes hábitat ecológicos del sistema. Se destaca la dirección del flujo neto de las aguas y la zona de pastos marinos que indican áreas de mayor influencia marina. Se muestran los cuatro Sistemas Fluvio-Lagunares principales asociados a la laguna. Se señalan las estaciones de colecta de 1976 a 1979 y de 1980 a 1981. Ver anexo de abreviaturas en Metodología. MÉTODOS
ACTIVIDADES DE CAMPOLas capturas de peces se efectuaron desde el mes de julio de 1976 a abril de 1981 de acuerdo al siguiente esquema: Capturas de julio a diciembre de 1976. Capturas de enero, marzo, mayo, agosto, septiembre y octubre de 1977. Capturas de enero, febrero, abril, junio, agosto y octubre de 1978. Capturas en marzo de 1979. Capturas mensuales de febrero de 1980 a abril de 1981. La distribución de las estaciones de colecta, caracterizó áreas contrastantes de la laguna, desde ambientes dulceacuícolas y de baja salinidad y ambientes marinos. De 1976 a 1979 las colectas realizadas caracterizaron sólo áreas del litoral interno de la Isla del Carmen y de los sistemas fluvio-lagunares. De 1980 a 1981, una red de 18 estaciones distribuidas en toda la laguna, proporcionaron información de la Cuenca Central, además de información complementaria de las áreas anteriormente señaladas (Ver Fig. 1). Todas las capturas fueron diurnas en profundidades no mayores de 4 m. Se utilizó una lancha de fibra de vidrio de 7 m de eslora con motor fuera de borda de 50 HP, empleando una red de prueba camaronera (chango) de 10 m de largo y 9 m de boca (abertura de trabajo de 5 m, que equivale al 60 % de la longitud horizontal de la boca), puertas de 0.8 por 0.5 m y luz de malla de 314 de pulgada. Los arrastres fueron de 6 a 20 min de duración. El material colectado se fijó en formol al 10 %. A los ejemplares capturados se les realizó una incisión ventral para la fijación del contenido estomacal y gónadas. En cada estación se midió la temperatura, salinidad, profundidad y transparencia. Se realizaron observaciones de tipo de substrato, vegetación sumergida y circundante, macrofauna béntica y datos climáticos. MATERIAL DE ESTUDIODe los 22 cruceros realizados (173 colectas) se obtuvieron un total de 4,741 ejemplares correspondientes a 7 especies, distribuidas como se muestra en la tabla 1.  TABLA 1 RELACIÓN DEL NÚMERO DE EJEMPLARES CAPTURADOS, POR ESPECIE, ÉPOCA CLIMÁTICA Y LOCALIDAD ACTIVIDADES DE LABORATORIOLos peces fueron lavados, separados y envasados en frascos de vidrio con alcohol metílico al 70%. Los ejemplares se identificaron usando el criterio de Aguirre León et al. (1982). Las mediciones de longitud se hicieron con un ictiómetro de 50 cm y las de pesos sobre material fijado en una balanza Ohaus Triple Beam de 0. 1 grs. de precisión y 2,6 10 grs. de capacidad. RELACIÓN PESO-LONGITUDEl análisis de la relación peso-longitud mediante el método de regresión lineal, considera la relación entre el peso en gramos y la longitud total en milímetros. Se usó la ecuación: P = aL b(1) donde: P = peso en gramos, L = longitud en milímetros, a y b = constantes a determinar. en la que se calcula el coeficiente de correlación R. La transformación Logarítmica de la ecuación (1) da una función lineal del tipo: log P = log a + b log L (2) Para ajustar las rectas de regresión lineal se utilizó una rutina del paquete BASIS (Burroughs Advance Statical Inquiry System) llamada "Multiple Linear Regression Analysis" con instrucciones para el cálculo de logaritmos, con la finalidad de establecer un modelo lineal. DETERMINACIÓN DE SEXOS Y ANÁLISIS GONÁDICOLa diferenciación de los sexos y la fase de maduración gonádica se hizo siguiendo el criterio de Nikolsky (1963). RELACIONES TRÓFICASSe analizaron un total de 280 estómagos de todas las especies, distribuidos como se muestra en la Tabla 2. El examen de los estómagos y el estado del pez se hizo siguiendo el criterio de Laevastu (1971). El grado de llenado de los estómagos fue determinado de acuerdo a si éste se encontraba: lleno, medio lleno, casi vacío y vacío. Para el análisis de las fases de digestión del contenido estomacal se siguieron los criterios de Carranza (1969) y Yáñez-Arancibia (1975). De acuerdo al tamaño de los estómagos, tipo y tamaño de alimento y el grado de digestión, es necesario combinar diferentes métodos para obtener información más completa sobre la preferencia alimenticia de las especies. Se utilizaron los métodos volumétricos, numérico, de frecuencia, el índice de importancia relativa de Pinkas et al.(1971), así como el índice de importancia relativa de Yáñez-Arancibia et al. (1976). La descripción detallada de estos métodos se encuentra en el trabajo de Lara-Domínguez et al. (1981). El índice de importancia relativa (Pinkas et al., 1971) consiste en las relaciones obtenidas por los métodos volumétrico, numérico y de frecuencia, ya que por sí solos pueden aportar muy poco en la verdadera evaluación del contenido estomacal y las relaciones tróficas de la especie. Este método es útil para interpretar la importancia relativa de algún alimento en específico, constituido por elementos de tamaño homogéneo. Sin embargo, puede introducir errores, ya que pueden existir numerosos organismos pequeños que opaquen la importancia relativa de otros de gran tamaño; asimismo la velocidad digestiva distorciona las medidas volumétricas y en lo que se refiere a la frecuencia, ésta es sensible a los errores de muestreo. El cálculo del índice (IRI) se realiza por la suma del porcentaje numérico y el porcentaje volumétrico, multiplicados por el valor porcentual de la frecuencia, quedando la siguiente expresión: IRI = F (N + V) (3) donde: IRI representa el índice de importancia relativa y F representa el porcentaje de frecuencia, N el porcentaje numérico y V el porcentaje volumétrico. El índice de importancia relativa de Yáñez-Arancibia et al. (1976) permite la cuantificación de la importancia relativa de determinado grupo trófico dentro de la alimentación de cada especie. Relaciona la frecuencia y el volumen del alimento siendo los parámetros más importantes en la alimentación de los peces. La relación matemática es la siguiente: IIR =F.V/ 100 (4) donde: IIR representa el índice de importancia relativa, F el porcentaje de la frecuencia y V el porcentaje volumétrico. Esta expresión es porcentual presentando un rango natural de 0 a 100. Se descarta el parámetro numérico por otorgar la misma importancia a organismos pequeños y grandes, y en término de ecología energética el error de interpretación puede ser muy grande. La combinación del índice de importancia relativa con la frecuencia y el volumen en una gráfica (Fig. 2), permite la representación del espectro trófico el cual queda delimitado por el porcentaje volumétrico y el porcentaje de frecuencia y evaluados por el índice de importancia relativa en relación a tres cuadrantes: 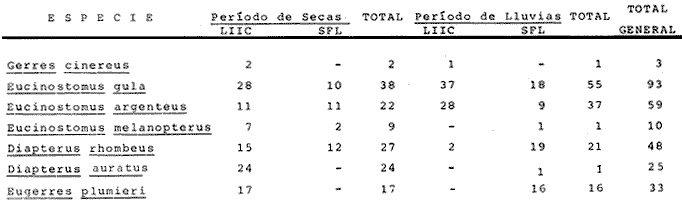 TABLA 2 RELACIÓN DEL NÚMERO DE ESTÓMAGOS ANALIZADOS, POR ESPECIE, ÉPOCA CLIMÁTICA Y LOCALIDAD 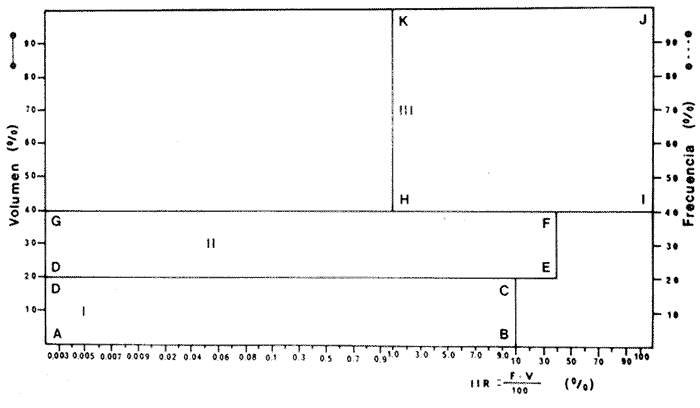 Fig. 2. Cuadro de relación volumen, frecuencia e índice de importancia relativa, para representar y evaluar espectros tróficos cuantitativos. Diagrama Trófico Combinado. (Tomado de Yáñez-Arancibia et al., 1976.) Cuadrante I. (ABCD)Zona de grupos tróficos ocasionales o circunstanciales. Está definido por el rango combinado de frecuencia y volumen de 0 a 20 % que representa grupos tróficos de importancia relativa baja; y para el índice de importancia relativa se define el rango evaluativo de 0 a 10 % que son grupos tróficos de importancia relativa baja. Cuadrante II (DEFG)Zona que define los grupos tróficos secundarios, presentando un rango combinado de volumen y frecuencia de 20 a 40 % que representan grupos tróficos de importancia secundaria y un rango evaluativo del índice de 10 a 40 % siendo grupos de importancia relativa secundaria. Cuadrante III (HIJK)Zona de grupos preferenciales o principales siendo determinados por un rango combinado de volumen y frecuencia de 40 a 100 % que define grupos tróficos de importancia alta, y el rango evaluativo del índice de 40 a 100 % que representa los grupos de importancia relativa alta. ABREVIATURA UTLILIZADAS EN TABLAS, FIGURAS Y TEXTOAn = Anfípodos Ao = Anomuros BPR = Boca de Puerto Real CA = Bajos del Cayo Ce = Cafalocordados CHB = Sistema Fluvio-Lagunar Chumpán-Balchacah CO = Copépodos CP = Sistema Fluvio-Lagunar Candelaria-Panlau CUC = Cuenca Central Cu = Cumáceos Es = Espículas ESP = Estero Pargo Esc = Escamas Fo = Foraminíferos Ga = Gasterépodos Hi = Huevos de Invertebrado HP = Huevos de Peces In = Insectos Is = Isópodos LIIC = Litoral Interno de la Isla del Carmen Li = Larvas de Insecto Mi = Misidáceos Min = Materia Inorgánica MNG = Puntilla Manigua MOND = Materia Orgánica no Determinable Ne = Nemátodos Ol = Oligoquetos Os = Ostrácodos PA = Sistema Fluvio-Lagunar Pom-Atasta Pe = Pelecípodos PE = Sistema Fluvio-Lagunar Palizada-del Este PG = Ensenada Punta Gorda Pi = Pignogónidos PJ = Punta San Julián Po = Poliquetos RA = Restos de Anélidos RC = Restos de Crustáceos Res = Esponjas Rp= Restos de peces RV = Restos Vegetales Si = Sipuncúlidos SFL = Sistemas Fluvio-Lagunares St = Estomatópodos Ta = Tanaidáceos RESULTADOS
FAMILIA GERREIDAE CUVIER (= GERRIDAE)
"Mojarras"Cuerpo elongado, elíptico o romboidal con escamas ctenoides. Línea lateral contínua, hasta la base de la caudal. Rostro puntiagudo; boca extremadamente protáctil. Surco premaxilar evidente en la parte superior de la cabeza, su pefil inferior cóncavo y casi toda escamada. Maxilar sin hueso suplementario. Dientes villiformes, sin incisivos ni morales, sin dientes sobre el vomer y palatinos. Pseudobranquias ocultas, membranas branquiales separadas, libres del itsmo; 6 radios branquiostegos, branquiespinas cortas, Nostrilos dobles. Dorsal simple, continua o hendida, 9 espinas y 10 radios. Anal simple, 2 o 3 espinas y 7 a 10 radios. Vaina escamosa en dorsal y anal. Pectorales puntiagudas; pélvicas 1-5, con procesos axilares; su origen posterior a las pectorales. Caudal ahorquillada. Cuerpo generalmente plateado. Algunas especies con barras verticales o longitudinales en los costados. Hocico obscuro. Aletas pares e impares con puntuaciones incoloras, obscuras o amarillas (CastroAguirre, 1978; Yáñez-Arancibia, 1978, Aguirre Leon et al.(1982).  CLAVE ARTIFICIAL ILUSTRADA PARA DETERMINAR ESPECIES DE LA FAMILIA GERREIDAE PRESENTES EN LA LAGUNA DE TÉRMINOSEstando este trabajo en prensa, fue colectada la especie Eugerres mexicanus(Steindachner, 1863) en sistemas fluvio-lagunares asociados a la Laguna de Términos. Se diferencia de otras del géneroEugerres de esta clave por presentar el cuerpo poco profundo y más alargado, bandas laterales de los costados pálidas, segunda espina dorsal muy fuerte y alta, segunda espina anal corta, escamas del cuerpo pequeñas, 43 a 45 escamas por debajo de la línea lateral.
 1. Cuerpo alargado o elíptico, moderadamente profundo; el maxilar llega a el margen anterior de la puplila, propérculo entero. De 6 a 8 branquiespinas bajo el primer arco. .........................2 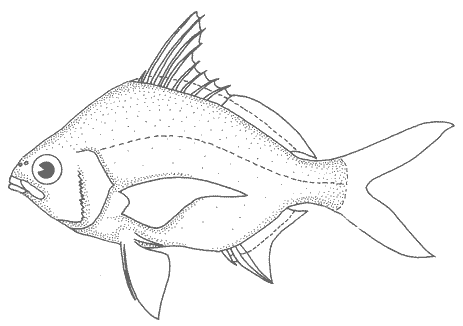 1'. Cuerpo corto profundo y romboidal; el maxilar sobrepasa el margen anterior de la pupila; preopérculo aserrado. De 10 a 18 branquiespinas bajo el primer arco..............5 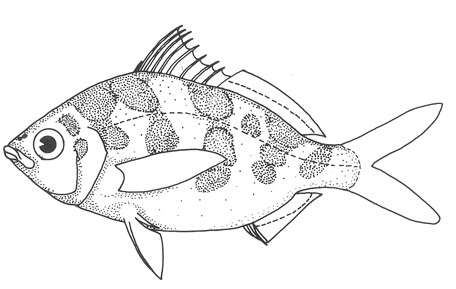 2. Sin barras verticales definidas sobre los costados. Los juveniles pueden presentar algunas barras obscuras de forma y número variable ......3 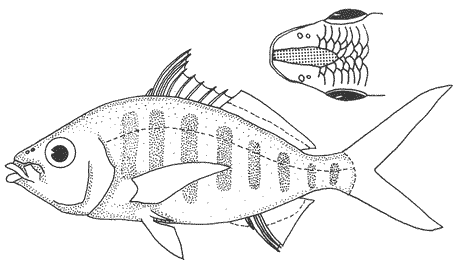 2'. Con 7 u 8 barras verticales azuladas siempre presentes en juveniles y adultos. Surco premaxilar ancho, oval y libre de escamas, segunda anal muy fuerte. ............................ Gerres cinereus (Walbaum). 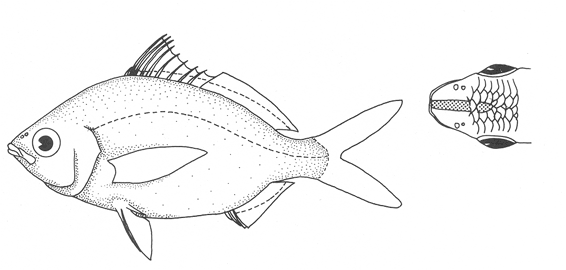 3. Cuerpo elíptico; profundidad menos de 2.6 veces en la LP; surco premaxilar ancho y escamado en su parte posterior...................................Eucinostomus gula (Cuvier). 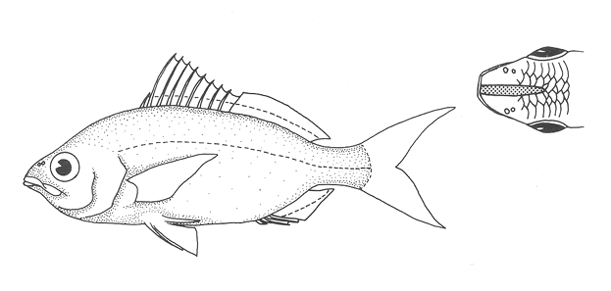 3' Cuerpo alargado; profundidad más de 2.6 veces en la LP; surco premaxilar estrecho y libre de escamas ...........4  4. Siete branquiespinas en la rama inferior del primer arco; dorsal espinosa con una porción obscura en su extremo distal. Ojos muy grandes, 2.0 a 3.4 en la cabeza ........................Eucinostomus argenteus Baird y Girard. 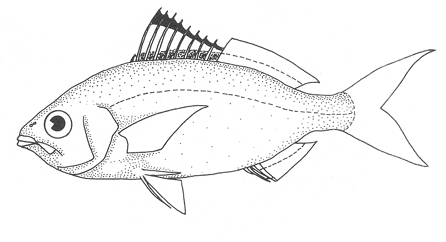 4 Ocho branquiespinas en la rama inferior del primer arco; dorsal espinosa tribandeada con una franja negra bien definida en su extremo distal. Ojos moderadamente grandes, 2.5 a 3.3 en la longitud cefálica........................Eucinostomus melanopterus (Bleeker)  5.Preorbital aserrado (entero en juveniles de menos de 90 mm de LT).Con 12 estrías longitudinales obscuras sobre los costados que siguen las hileras de las escamas, no muy evidentes en ejemplares juveniles. Segunda espina dorsal fuerte y más alta que la tercera. Anal II-III, 7/9.................................... Eugerres plumieri (Cuvier)  5'.Preorbital entero. Sin estrías longitudinales. Segunda espina dorsal fuerte, igual o más corta que la tercera ........................ 6  6. Anal III, 8-9. De 37 a 42 escamas por debajo de la línea lateral; 9 a 11 branquiespinas bajo el primer arco............................................... Diapterus auratus Ranzaní 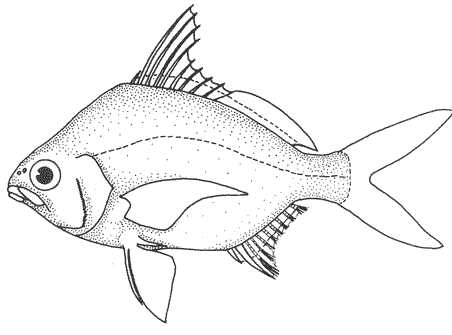 6'. Anal II, 8-10. De 30 a 40 escamas por debajo de la línea lateral. De 14 a 16 branquiespinas bajo el primer arco......................................Diapterus rhombeus (Cuvier) DIAGNOSIS DE LAS ESPECIES
Género Gerres Cuvier, 1824
Gerres cinereus (Walbaum, 1972)N.v.: "Mojarra blanca", "Mojarra plateada" Mugil cinereus Walbaum, Artedi General Piscium, 1792: 228 (Bahamas). Diagnosis. Cuerpo oblongo, 2.1 a 2.7 en la LP. Cabeza a 3.3 en la LP; perfil anterior convexo; maxilar llegando al margen anterior de la pupila. Surco premaxilar desnudo en forma de U. Preopérculo y preorbital enteros. Branquiespinas pequeñas y delgadas, 6 a 8 en la rama inferior del primer arco. D IX, 10: A 111, 7, la segunda espina fuerte. Pectorales más grandes que la cabeza, sobrepasando la abertura anal. Pélvicas pequeñas, no sobrepasando la abertura anal. De 39 a 47 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales claras. Dorsal y caudal obscuras. Pélvicas y anal amarillas; pélvicas con puntuaciones. Costados con barras verticales azules presentes en todas las edades. Género Eucinostomus Baird y Girard, 1854
Eucinostomus gula (Cuvier, 1830)N.v.: "Mojarrita", "Mojarrita de Ley" Gerres gula Cuvier, In: Cuvier y Valenciennes, Histoire Naturalle des Poissons, 1830, 6: 464 (Martinica). Diagnosis. Cuerpo elíptico, profundidad 2.0 a 2.5 en la LP. Cabeza 2.4 a 3.3 en la LP; perfil anterior suavemente convexo; maxilar sobrepasando ligeramente el margen anterior de la pupila; surco premaxilar ancho y atravesado por escamas; formándose una pequeña foseta circular. Preopérculo y preorbital enteros. Branquiespinas pequeñas y delgadas, 7 en la rama inferior del primer arco. D IX,10; A 111, 7-8, la segunda espina fuerte. Pectorales más grandes que la cabeza, sobrepasando la abertura anal. Pélvicas pequeñas, no sobrepasando la abertura anal. De 40 a 47 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales, pélvicas y caudal claras; caudal con puntuaciones. Dorsal y anal obscuras, margen distal de la dorsal espinosa negro. Eucinostomus argenteus Baird y Girard, 1854N.v.: "Mojarrita plateada", "Mojarra charrita" Eucinostomus argenteus Baird y Girard.In: Baird y Girard, Ann. Rep. Stnithsonian Inst., 1854: 335 (Beseley Point, New jersey). Diagnosis. Cuerpo alargado, profunidad 2.7 a 3.3 en la LP. Cabeza 2.6 a 3.5 en la LP: perfil anterior recto; maxilar llegando al margen anterior de la órbita; surco premaxilar estrecho y desnudo. Preopérculo y preorbital enteros. Branquiespinas pequeñas y delgadas, 7 en la rama inferior del primer arco. D IX,10; A 111, 7, la segunda espina fuerte. Pectorales iguales o más pequeñas que la cabeza, sin sobrepasar la abertura anal. Pélvicas más cortas que la cabeza. De 43 a 48 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales, pélvicas y caudal claras; caudal con puntuaciones. Dorsal obscura. Eucinostomus melanopterus (Bleeker, 1853)N.v.: "Mojarrita bandera", "Mojarra trompetera" Gerres melanopetrusBleeker, Nat. Verh. Holl. Maatsch. Wetensh, 1863, 2 (18): 44, lám. 8, fig. 1 (Costa de Guinea) Diagnosis.Cuerpo alargado, profundidad 2.6 a 3.2 en la LP. Cabeza 2.5 a 3.2 en la LP; perfíl anterior poco convexo; maxilar sobrepasando ligeramente el margen anterior de la órbita; surco premaxilar grande, en línea y libre de escamas. Preopérculo y preorbital enteros. Branquiespinas pequeñas y delgadas, 8 en la rama inferior del primer arco. D IX, 10; A 111, 7, la segunda espina fuerte. Pectorales iguales o más pequeñas que la cabeza, sobrepasando la abertura anal. Pélvicas más cortas que la cabeza, pueden alcanzar la abertura anal. De 46 a 51 escamas por debajo de la línea-lateral. Cuerpo plateado, claro ventralmente. Pectorales, pélvicas y anal claras. Caudal obscura. Dorsal claramente tribandeada. Género Diapterus Ranzani, 1840
Diapterus rhombeus (Cuvier, 1829)N.v.: "Mojarra pinta", "Mojarra" Gerres rhombeus Cuvier, Le Regne Animal, 1829, 2, Ed. 2: 189 (Martinica, Santo Domingo). Diagnosis. Cuerpo corto, profundo y romboidal, profundidad 1.7 a 2.0 en la LP. Cabeza 2.6 a 3.2 en la LP; perfil anterior convexo; maxilar llegando hasta el centro del ojo; surco premaxilar ancho y libre de escamas. Preopérculo aserrado, preorbital entero. Branquiespinas 12 a 18 (14-16) en la rama inferior del primer arco. D IX, 10; A 11, 9-10, segunda espina fuerte y más corta que la segunda dorsal. Pectorales y pélvicas poco más cortas que la cabeza, ambas sobrepasando la abertura anal. De 35 a 41 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales y caudal claras; caudal con puntuaciones. Dorsal algo obscura, con margen negro en la porción espinosa. Pélvicas y anal amarillas. Diapterus auratus Ranzani, 1842N.v.: "Mojarra blanca" Diapterus auratus Ranzani, 1842 (Costas de Brasil) Diagnosis.Cuerpo corto, profundo y romboidal, profundidad 1. 8 a 2.2 en la LP. Cabeza 2.6 a 3.2 en la LP; perfil anterior corto y convexo; maxilar sobrepasando el margen anterior de la pupila; surco premaxilar ancho y escamado. Preopérculo aserrado, preorbital entero. Branquiespinas 9 a 11 en la rama inferior del primer arco. D IX, 10; A 111, 8-9, la segunda espina fuerte. Pectorales algo más grandes que la cabeza, sobrepasando la abertura anal. De 37 a 42 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales y caudal claras; caudal con puntuaciones. Dorsal obscura con puntuaciones y margen distal negro. Pélvicas y anal amarillas con puntuaciones. Ampliamente utilizado en la literatura, es el nombre de D. olisthostomus. Ambas nomenclaturas se refieren a la misma especie (i. e., D. aratus = D. olisthostomus) pero recientemente se ha generalizado la utilización de D. auratus (Miller, 1976). Aguirre León et al. (1982) utilizan el nombre de D. olisthostomus en la Laguna de Términos y además numerosa literatura reciente utiliza este nombre (Cervigón, 1966; Guitart, 1977; Fisher, 1977; Castro-Aguirre, 1978 y Reséndez, 1981). Género Eugerres Jordan y Everman, 1927
Eugerres plumieri (Cuvier, 1830)Gerres plumieri Cuvier, In: Cuvier y Valenciennes, Histoire Naturalle des Poisson, 1830, 6: 452, lám. 167 (Puerto Rico, Antillas). Diagnosis Cuerpo corto, profundo y romboidal, profundidad 1.8 a 2.1 en la LP. Cabeza 2.5 a 3.0 en la LP; perfil anterior convexo encima de los nostrilos, recto encima de los ojos, muy convexo en la nuca: maxilar sobrepasando el margen anterior de la pupila; surco premaxilar ancho y desnudo. Preopérculo y preorbital aserrados (excepto en juveniles, el preorbital es entero). Branquiespinas 14 a 16 en la rama inferior del primer arco. D IX, 10, segunda espina fuerte, alta y ensanchada. A 11-111, 7-9, segunda espina fuerte. Pectorales y pélvicas iguales o más cortas que la cabeza, ambas sobrepasando la abertura anal. De 35 a 42 escamas por debajo de la línea lateral. Cuerpo plateado, claro ventralmente. Pectorales doradas. Dorsal obscura con puntuaciones. Caudal obscura y con puntuaciones. Alrededor de 12 estrias longitudinales obscuras sobre los costados. ASPECTOS BIOLÓGICOS Y ECOLÓGICOS DE LAS ESPECIES
Gerres cinereus (Walbaum, 1972)Es una especie poco estudiada en ambientes lagunares y estuarinos del Golfo de México y Mar Caribe a pesar de su amplia distribución en el Atlántico occidental. La mayoría de los trabajos que la reportan en aguas mexicanas tan sólo son registros de su presencia en aguas costeras y continentales, careciéndose de información sistemática sobre su biología y ecología. Es una especie rara en la Laguna de Términos por su baja frecuencia. Se considera como una especie visitante ocasional y consumidor de primer orden. En el área de estudio se ha colectado en las localidades Estero Pargo y Punta Gorda. Se obtuvieron 3 ejemplares de 89 a 159 mm de longitud total. Relación peso-longitud y frecuencia de tallas. No se efectuó el análisis de regresión debido a que sólo se colectaron 3 ejemplares de 89 a 159 mm en el área de estudio. Madurez.No fue posible determinar el sexo y los estadios de madurez de los ejemplares. Alimentación. El análisis de la alimentación de Gerres cinereusse basó en la observación del contenido estomacal de 3 ejemplares, 2 juveniles y 1 adulto. De las observaciones hechas se encontraron en los estómagos analizados los siguientes grupos: pelecípodos, MOND, gasteropodos, poliquetos, restos vegetales y ostrácodos, siendo los más abundantes la materia orgánica y los pelecípodos. Eucinostomus gula (Cuvier, 1830)Es una especie típica en ambientes lagunares y estuarinos del Golfo de México y Mar Caribe. Numerosos trabajos la citan dentro de sus listas ictiofaunísticas a lo largo de su amplia distribución en el Atlántico de América. Destaca como especie importante en ambientes estuarinos por su gran abundancia numérica y biomasa a lo largo de todo el año. Sin embargo, su presencia se restringe a tallas juveniles en esos ambientes. Publicaciones recientes en el Sur del Golfo de México resaltan el papel que juega esta especie en la estructura y función de las comunidades de peces de la Laguna de Términos (Aguirre León et al., 1982; Yáñez-Arancibia et al., 1983 y otras referencias por ellos citadas). Eucinostomus gula es la especie mejor representada en distribución y abundancia de todas las mojarras presentes en la Laguna de Términos. Constituyen un recurso económico potencial en el área. Es considerada como una especie residente permanente y consumidor de primer orden. Se ha colectado en toda la laguna, obteniéndose 2,349 ejemplares de 30 a 160 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación los valores calculados para las constantes a y b son dados en función de la época climática. Asimismo el análisis del rango de tallas se presenta comparativamente para las dos épocas climáticas y los dos subsistemas estudiados (Figs. 3 a 7). Se observa que predominan las tallas juveniles en las dos épocas climáticas reflejándose en las modas. Sin embargo las tallas más grandes están mejor representadas en el litoral interno de la Isla del Carmen en la época de secas. No obstante, se aprecia comparando los dos subsistemas que las tallas mayores están presentes en el litoral interno de la Isla del Carmen durante ambos períodos climáticos y las menores en los sistemas Fluvio-Lagunares. Madurez. La frecuencia de sexos y f ases sexuales en función de la época climática se muestra graficada en las figuras 8 y 9. Se observa que para los períodos climáticos definidos, predominan los estadios juveniles en fase I y II, presentándose pocos ejemplares en fase III en el litoral interno de la Isla del Carmen, durante la época de lluvias y otros más en fase IV en el mismo subsistema en la época de secas, por lo tanto no aparecen individuos en reproducción. Las fases I y II están representadas tanto por machos como por hembras, presentándose III y IV sólo en hembras. Se encontraron ejemplares indiferenciados en sexo y maduración en el litoral interno de la Isla del Carmen en ambas épocas climáticas. Del análisis de proporción de sexos macho: hembra se encontró que el total de la población tiene una proporción de 1. 2: 1. 0; siendo para la época de secas de 1.3: 1.0 y durante el período de lluvias 1.1: 1.0. Se aprecia claramente que existe una distribución similar en ambos sexos. La figura 10, muestra la relación de la madurez gonádica y la talla de los individuos así como la relación total del desarrollo sexual de la población independiente del sexo, lo cual es fundamental para determinar la longitud a la cual dicha población alcanza la madurez gonádica y es apta para reproducirse. Se observa que durante la época de lluvias se encuentran individuos en maduración desde los 102 mm de longitud total en fase III. La fase I está comprendida entre los 36 y 95 mm de longitud total y la fase II entre los 73 y los 117 mm. Por el contrario en la época de secas se encuentran individuos maduros en fase IV desde los 140 mm. Durante este período no se encontraron ejemplares en fase III y siguen predominando las fases I y II desde los 30 a 100 mm para la primera y entre los 76 y 115 mm para la segunda. Es importante destacar que los individuos maduros sólo se registraron en el litoral interno de la Isla del Carmen durante la época de secas, cuando la salinidad presenta valores elevados (mayores a 26%). 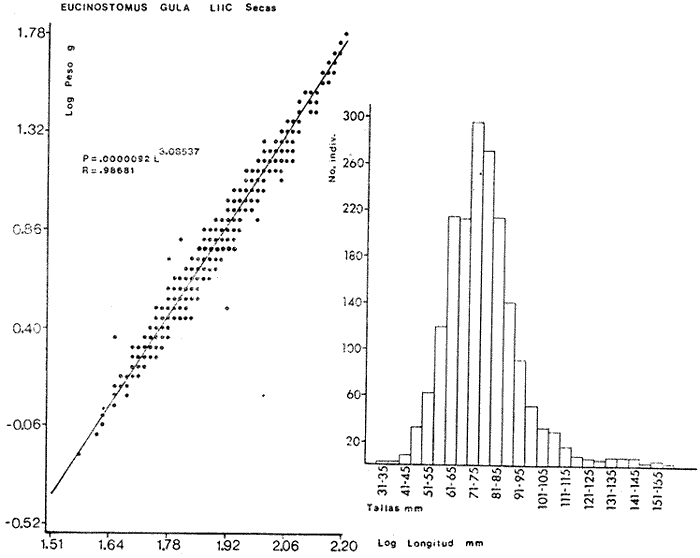 Fig. 3. Recta de regresión de la relación peso/longitud de Eucinostomus gula en el Litoral Interno de la Isla del Carmen durante la época de secas. Se señala la distribución de frecuencias de tallas. Alimentación. El estudio de las relaciones tróficos de Eucinostomus gula se basó en el análisis del contenido estomacal de ejemplares juveniles (menos de 100 mm) en función de las localidades estudiadas y las épocas climáticas. Litoral interno de la Isla del Carmen. Epoca de secas. El espectro trofico de E. gula (Fig. 1l), indica que se alimenta al menos de 16 grupos tróficos entre los que destacan como importantes: poliquetos con un 69.6% en volumen y 89.2% de frecuencia, MOND con un 9.8 % en volumen y 32. 1 % de frecuencia, oligoquetos con un 9.2 % en volumen y 17.8 % de frecuencia, tanaidáceos con un 5.6 % en volumen y 21.4 % de frecuencia y pequeños decápodos con un 2.4 % en volumen y 7.1 % de frecuencia. Numéricamente los grupos predominantes son: poliquetos con un 38.9 %, copépodos con un 12.5 % y nemátodos con 10. 2 %, otros como huevos de peces, tanaidáceos, anfipodos, restos de peces y oligoquetos se presentan en menor proporción. De acuerdo al análisis combinado del índice de importancia (IRI) destacan poliquetos, MOND, nemátodos, tanaidáceos, copépodos y oligoquetos con los valores más altos. Se puede establecer en función del índice IIR que el alimento preferencial lo constituyen los poliquetos con un 62 %, se considera secundario la materia orgánica (MOND) con un 3.1 % y los circunstanciales los 14 grupos restantes.  Fig. 4. Recta de regresión de la relación peso/longitud de Eucinostomus gula en el Litoral Interno de la Isla del Carmen durante la época de lluvias. Se señala la distribución de frecuencias de tallas. Sistemas Fluvio-Lagunares. Epoca de secas. El espectro trófico (Fig. 12), indica que se alimenta al menos de 12 grupos tróficos entre los que destacan como importantes: poliquetos con un 57.5 % en volumen y 70 % de frecuencia, MOND con un 21.7 % de volumen y 40 % de frecuencia, copepodos con un 12.9 % en volumen y 50 % de frecuencia, y ostrácodos con un 5.7 % en volumen y 50 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con un 84.4 %, poliquetos con 6.3 % y ostrácodos con 5.8 %. De acuerdo al análisis del índice de importancia relativa IRI destacan copépodos, poliquetos, MOND, ostrácodos y nemátodos con los valores más altos.  Fig. 5. Recta de regresión de la relación peso/longitud de Eucinostomus gula en los Sistemas Fluvio-Lagunares durante la época de secas. Se señala la distribución de frecuencia de tallas. En función del índice IIR se puede establecer que el alimento preferencial lo constituyen los poliquetos con un 40 %, se considera secundario la materia orgánica con un 8.6 % y circunstanciales los 10 grupos restantes. Litoral Interno de la Isla del Carmen. Epoca de lluvias. El espectro trófico. (Fig. 13), indica que se alimenta al menos de 19 grupos tróficos entre los cuales destacan como importantes: poliquetos con un 67 % en volumen y 81 % de frecuencia, MOND con un 11 % en volumen y 21.6 % de frecuencia, sipuncúlidos con un 8.2 % en volumen y 13.5 % de frecuencia, tanaidáceos con un 4.4 % en volumen y 10. 8 % de frecuencia, copépodos con un 2.8 % en volumen y 21.6 % de frecuencia y en menor proporción cefalocordardados. Numéricamente los copépodos se presentan en un 66.7 %, poliquetos con un 9.4 %, tanaidáceos en un 7.4 %, nemátodos con un 4. 0 % y gasterópodos y restos de peces en menor proporción. El índice IRI indica que los grupos más importantes son: poliquetos, copépodos, MOND, tanaidáceos, nemátodos y sipuncúlidos. En función del índice IIR del grupo preferencial son los poliquetos con un 54.2 % y los 18 restantes son circunstanciales, en este caso no aparece ningún grupo como secundario. 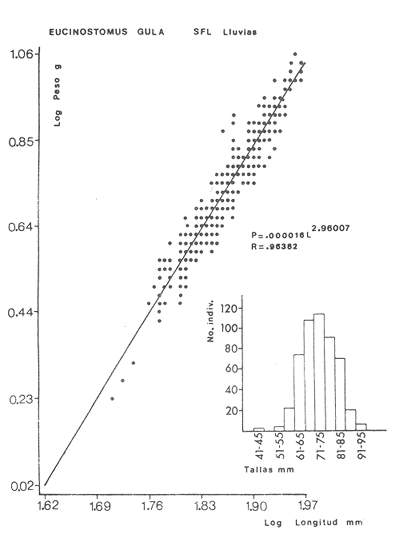 Fig. 6. Recta de regresión de la relación peso/longitud de Eucinostomus gula en los Sistemas Fluvio-Lagunares durante la época de lluvias. Se señala la distribución de frecuencia de tallas. 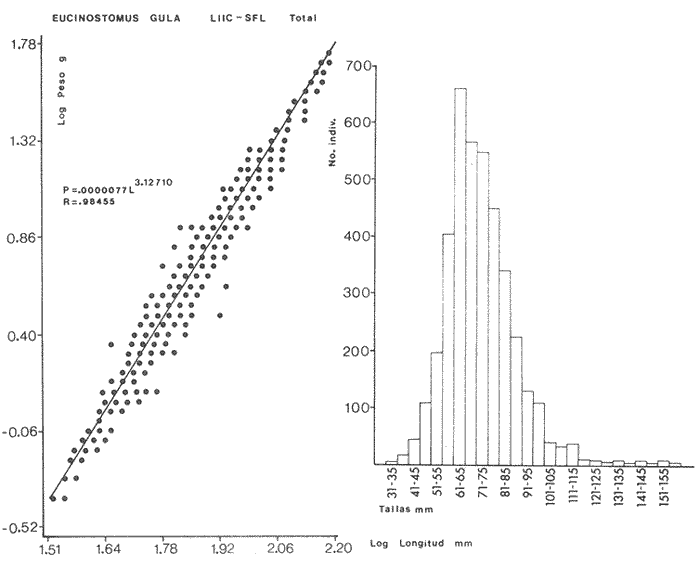 Fig. 7. Recta de regresión de la relación peso/longitud de Eucinostomus gula para las épocas de secas y lluvias en el Litoral de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Se señala la distribución de frecuencia de tallas. Sistemas Fluvio-Lagunares. Epoca de lluvias. El espectro trófico (Fig. 14), indica que la especie se alimenta al menos de 15 grupos tróficos. Los grupos que destacan son: poliquetos con 65.2% en volumen y 83.3% de frecuencia, MOND con un 14.5 % en volumen y 33.3 % de frecuencia y en menor proporción oligoquetos, y pequeños decápodos. Numéricamente los grupos predominantes son: copépodos con 34 %, ostrácodos con 18.5 %, poliquetos con 13.3 % y tanaidáceos con 12.9 %. De acuerdo al índice IRI, los poliquetos, tanaidáceos, copépodos, ostrácodos y MOND son los más importantes. En función del índice IIR el grupo preferencial lo constituyen los poliquetos, se considera como secundario la materia orgánica y los 13 restantes como grupos circunstanciales. Eucinostomus argenteus Baird y Girard, 1854Esta especie se considera también como típica de ambientes lagunares y estuarinos del Golfo de México y Mar Caribe. Su distribución es amplia en el Atlántico occidental y es citada por numerosos trabajos en sus listas ictiofaunísticas. Ha sido estudiada en el Norte del Golfo de México por Springer y Woodburn (1960) para las aguas de la Bahía de Tampa, Florida como especie ecológicamente importante. Su presencia en ambientes estuarinos es principalmente por tallas juveniles. Carece al igual que otras especies afines de estudios sobre su biología.  Fig. 8. Fases de desarrollo sexual y proporción de sexos de Eucinostomus gula durante la época de secas en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Eucinostomus argenteus es la tercera especie más abundante de las mojarras de la Laguna de Términos. Es considerada como visitante cíclico y consumidor de primer orden. En el área de estudio se ha colectado en las localidades PA, PE, CP, BPR, PJ, PG, ESP y CUC. Se obtuvieron 165 ejemplares de 20 a 145 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación, los valores calculados para las constantes a y b son dados en función de la época climátiea. Asimismo el análisis del rango de tallas se presenta comparativamente para las dos épocas climáticas y los dos subsistemas estudiados (Figs. 15 a 19). Se observa que pocos individuos representan a las tallas juveniles a preadultas en las dos épocas climáticas, sin embargo, predominan los juveniles en frecuencia. Las tallas más grandes están mejor representadas en el litoral interno de la Isla del Carmen en la época de secas. Comparando los dos subsistemas se aprecia que las tallas menores están presentes en el subsistema del litoral interno de la Isla pero en el período de lluvias. 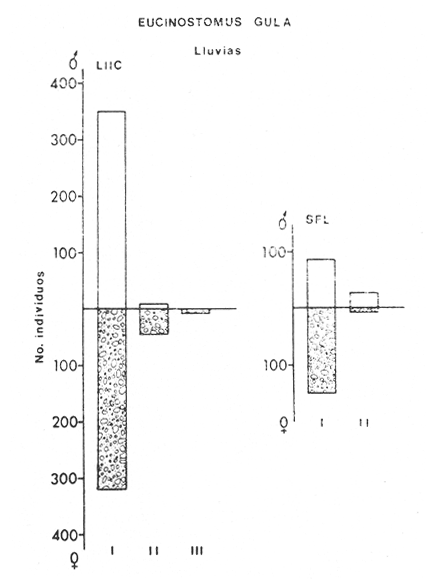 Fig. 9. Fases de desarrollo sexual y proporción de sexos de Eucinostomus gula durante la época de lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Madurez. La frecuencia de sexos y fases sexuales en función de la época climática se muestra graficado en las figuras 20 y 21. Se observa que para los períodos climáticos definidos predominan los estadios juveniles en fase I y II, presentándose pocos ejemplares en fase III en el litoral interno de la Isla del Carmen en la época de lluvias, por lo tanto no aparecen individuos en reproducción. Las fases I y II están representadas tanto por machos como por hembras, presentándose la fase III sólo en las hembras. Se encontraron ejemplares indiferenciados en sexo y maduración en el litoral interno de la Isla del Carmen en la época de lluvias. 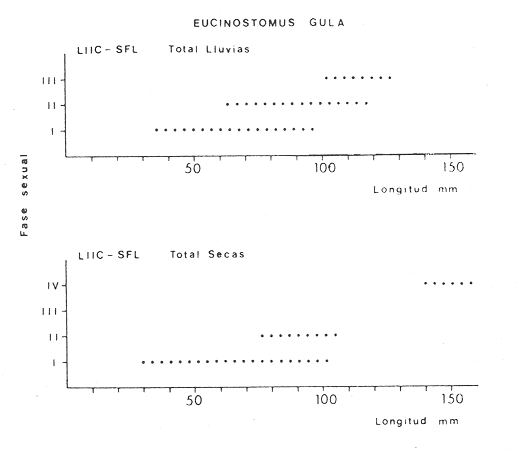 Fig. 10. Relación de la longitud total y las fases de desarrollo sexual de Eucinostomus gula independiente del sexo durante las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Del análisis de proporción de sexos macho: hembra se encontró que el total de la población guarda una relación de 1. 9: 1.0; siendo para la época de secas de 1.0: 1.3 y durante el período de lluvias de 4.2: 1.0. La figura 22, muestra la relación de la madurez gonádica y la talla de los individuos así como la relación total del desarrollo sexual de la población independiente del sexo, lo cual es fundamental para determinar la longitud a la cual dicha población alcanza la madurez gonádica y es apta para reproducirse. Se observa que durante la época de lluvias se encuentran individuos en maduración desde los 122 mm de longitud total en fase III. La fase I está comprendida entre los 30 y 88 mm y I fase II entre los 95 y 125 mm. En el período de secas se encuentran individuos en maduración desde los 130 mm en fase III y siguen predominando las fases I y II desde los 36 a 114 mm para la primera y entre los 66 a 137 mm para la segunda. Alimentación. El estudio de las relaciones tróficas de Eucinostomus argenteusse basó en el análisis del contenido estomacal de ejemplares juveniles (menos de 120 mm) en función de las localidades y la época climática. Litoral interno de la Isla del Carmen. Epoca desecas. El espectro deEucinostomus argenteus (Fig. 23), indica que se alimenta al menos de 15 grupos tróficos entre los que destacan como importantes: poliquetos con un 53.6 % en volumen y 63.6 % de frecuencia, MOND con un 39.5 % en volumen y 45.4 % de frecuencia y copépodos con un 2.9 % en volumen y 81.8 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con 43.6 %, ostrácodos con un 21.9%, huevos de invertebrados, nemátodos y restos de peces están presentes en menor proporción. De acuerdo al índice IRI destacan copepodos, poliquetos, MOND y ostrácodos con los valores más altos. En función del índice IIR se considera como alimento preferencial a los poliquetos, siendo el secundario la materia orgánica (MOND) y los 13 grupos restantes como circunstanciales. Sistemas Fluvio-Lagunares. Epoca desecas. El espectro trófico (Fig. 24), indica que se alimenta al menos de 12 grupos tróficos, destacando como importantes: MOND con un 55.5 % en volumen y 72.7 % de frecuencia, poliquetos con un 29.3 % en volumen y 90.9 % de frecuencia y tanaidáceos con 11.3% en volumen y 45.4% de frecuencia. Numéricamente los grupos predominantes son: huevos de invertebrados con 33.2 %, copépodos con 32.9.% y ostrácodos con 23.5 %. De acuerdo al índice IRI destacan: MOND, ostrácodos, copépodos, tanaidáceos, huevos de invertebrados y poliquetos con los valores más altos. En función del índice IIR se encuentra que el alimento preferencial es la materia orgánica con 41 %, como secundarios se considera a los tanaidáceos y poliquetos y los restantes como grupos circunstanciales.  Fig. 11. Espectro trófico de la población de Eucinostomus gula en el Litoral Interno de la Isla del Carmen durante la época de secas. Ver anexo de abreviaturas en Metodología. 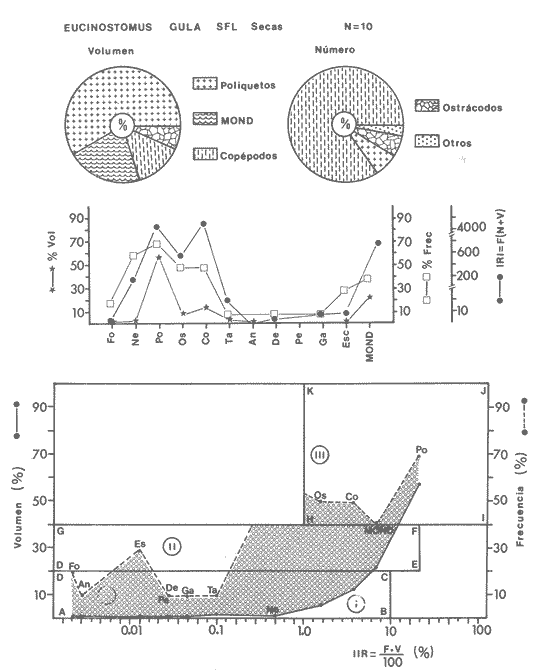 Fig. 12. Espectro trófico de la población de Eucinostomus gula en los Sistemas Fluvio-Lagunares durante la época de secas. Ver anexo de abreviaturas en Metodología. 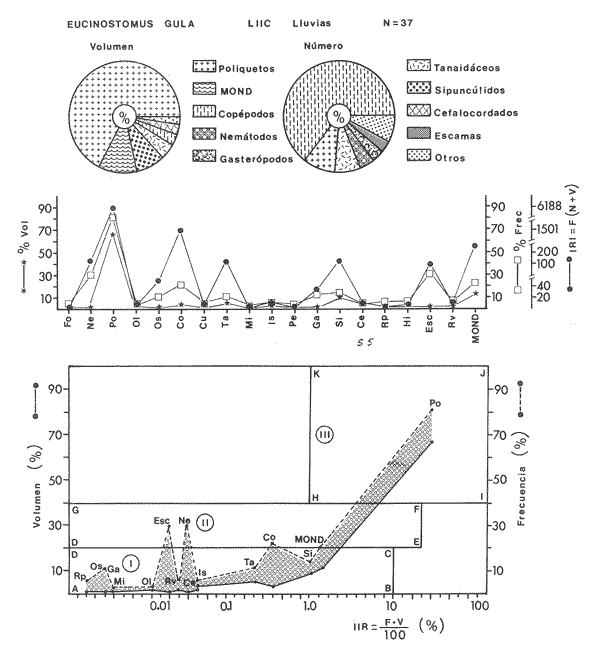 Fig. 13. Espectro trófico de la población de Eucinostomus gula en el Litoral de la Isla del Carmen durante la época de lluvias. Ver anexo de abreviaturas en Metodología. 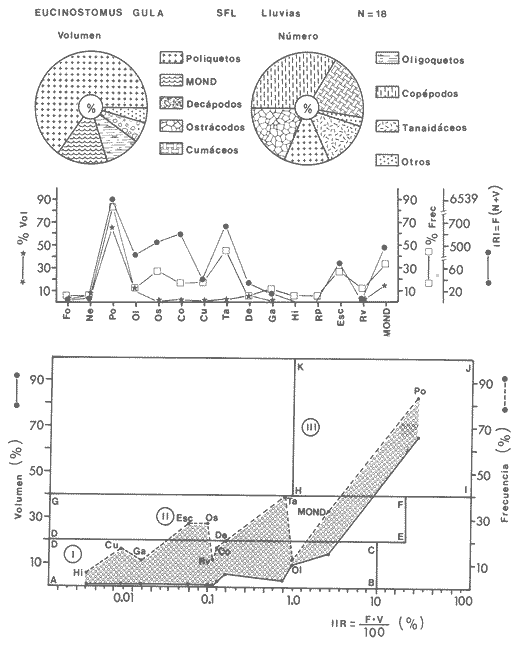 Fig. 14. Espectro trófico de la población de Eucinostomus gula en los Sistemas Fluvio-Lagunares durante la época de lluvias. Ver anexo de abreviaturas en Metodología.  Fig. 15. Recta de regresión de relación peso/longitud de Eucinostomus gula en el Litoral Interno de la Isla del Carmen durante la época de secas. Se señala la distribución de frecuencia de tallas. 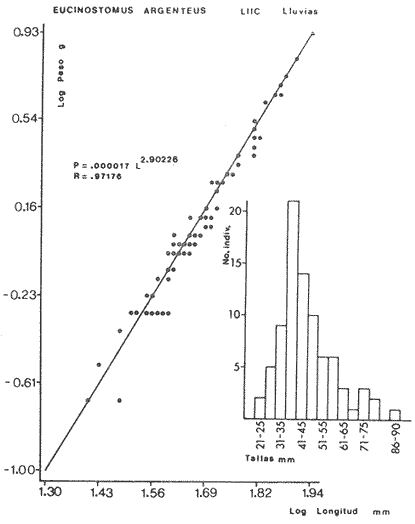 Fig. 16. Recta de regresión de la relación peso/longitud de Eucinostomus argenteus en el Litoral Interno de la Isla del Carmen durante la época de lluvias. Se señala la distribución de frecuencia de tallas. Litoral interno de la Isla del Carmen. Epoca de lluvias. El espectro trófico (Fig. 25), indica que se alimenta al menos 18 grupos tróficos entre los que destacan como importantes: poliquetos con un 42 % en volumen y 57.1 % de frecuencia, MOND con un 25.2 % en volumen y 39.2 % de frecuencia, tanaidáceos con un 23.5 % en volumen y 46.4 % de frecuencia y copépodos con un 6.5 % en volumen y 78.5 % de frecuencia. Numéricamente el grupo predominante lo constituye los copépodos con un 86.4 %. El índice IRI indica que los grupos más importantes son: poliquetos, copépodos, tanaidáceos y MOND. En función del índice IIR se considera como grupo preferencial a los poliquetos siendo secundarios los tanaidáceos y la materia organica (MOND), y los 15 grupos restantes como circunstanciales.  Fig. 17. Recta de regresión de la relación pesol/ongitud de Eucinostomus argenteus en los Sistemas Fluvio-Lagunares durante la época de secas. Se señala la distribución de frecuencia de tallas. Sistemas Fluvio-Lagunares. Epoca de lluvias. El espectro trófico (Fig. 26), indica que destacan los siguientes grupos importantes: tanaidáceos con un 71.6 % en volumen y 88.8 % de frecuencia, poliquetos con un 12.6 % en volumen y un 55.5 % de frecuencia y MOND con un 12 % en volumen y 44.4 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con un 35.8%, tanaidáceos con un 29.7%, ostrácodos con 15.2 % y poliquetos con 14.5 %. De acuerdo al índice IRI, los grupos más importantes son los tanaidáceos, los poliquetos y MOND con los valores más altos. Con relación al índice IIR, el grupo preferencial lo constituye los tanaidáceos con 63.5 %, se consideran como secundarios los poliquetos y a la materia orgánica (MOND), y los 8 restantes como grupos circunstanciales. 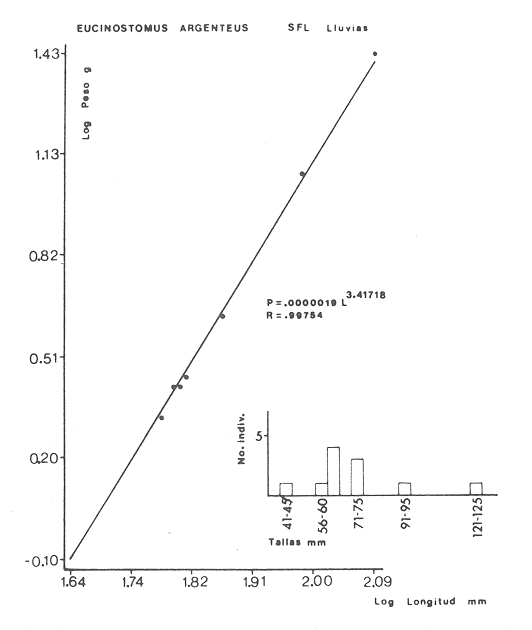 Fig. 18. Recta de regresión de la relación peso/longitud de Eucinostomus argenteus en los Sistemas Fluvio-Lagunares durante la época de lluvias. Se señala la distribución de frecuencia de tallas. Eucinostomus melanopterus (Bleeker, 1863)Es una especie poco estudiada en ambientes lagunares y estuarinos del Golfo de México y Mar Caribe. Tiene una amplia distribución en el Atlántico occidental. La mayoría de los trabajos que la reportan en aguas mexicanas tan sólo son registros de su presencia en aguas costeras y continentales, careciendo casi por completo de observaciones bioecológicas. Eucinostomus melanopterus es una especie rara en la Laguna de Términos por su baja frecuencia. Se considera como una especie visitante ocasional y consumidor de primer orden. En el área de estudio se ha colectado en las localidades PA, CP, PG, MNG, ESP y CUC. Se obtuvieron 13 ejemplares de 50 a 120 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación, los valores calculados para las constantes a y b sólo corresponden al litoral interno de la Isla del Carmen en la época de secas. Asimismo el análisis del rango de tallas se presenta para la época climática y el subsistema estudiado (Fig. 27). Se observa que no existe una distribución homogénea en las tallas, más bien, se presentan distribuidas al azar. Las tallas más grandes están mejor representadas en el litoral interno de la Isla del Carmen en la época de secas. Comparando los dos subsistemas se aprecia que las tallas menores están presentes en los sistemas fluvio-lagunares en el periodo de lluvias. Madurez. La frecuencia de sexos y fases sexuales en función de la época climática se muestra graficada en la figura 28. Se observa que para los períodos climáticos definidos, predominan los estadios juveniles en fase I y II en ambos subsistemas, por lo tanto no aparecen individuos en reproducción. Las fases I y II están representadas tanto por machos como por hembras. Del análisis de proporción de sexos macho: hembra se encontró que el total de la población guarda una proporción de 3.5: 1.0, siendo para la época de secas de 3.0: 1.0. Para el período de lluvias no se cuenta con datos. La fig. 29, muestra la relación de la madurez gonádica y la talla de los individuos, así como la relación total del desarrollo sexual de la población independiente del sexo, lo cual es fundamental para determinar la longitud a la cual dicha población alcanza la madurez gonádica y es apta para reproducirse. Los resultados para la época de secas muestran individuos sólo en fases I y II; la primera comprendida entre los 50 y 114 mm de longitud total y para la segunda entre 112 y 135 mm. Alimentación.El análisis de alimentación de Eucinostomus melanopterus se basó tan sólo en la observación del contenido estomacal de 10 ejemplares. Es una especie rara y sólo se han colectado pocos individuos. De las observaciones hechas se encontraron en los estomagos analizados los siguientes grupos tróficos: ostrácodos, tanaidáceos, poliquetos, copépodos, nemátodos, foraminíferos, cumáceos, materia orgánica, restos de decápodos, restos de vegetales y escamas, siendo los más abundantes la materia orgánica, los copépodos y los restos vegetales. 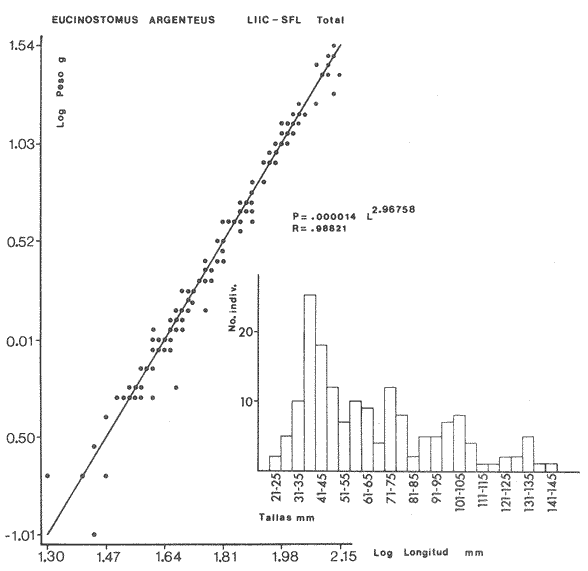 Fig. 19. Recta de regresión de la relación peso/longitud de Eucinostomus argentus para las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los sistemas Fluvio-Lagunares. Se señala la distribución de frecuencia de tallas. Diapterus rhombeus (Cuvier, 1829)Es una especie típica en ambientes lagunaresestuarinos del Golfo de México y Mar Caribe. Diversos trabajos la citan dentro de sus listas ictiofaunísticas a lo largo de su amplia distribución en el Atlántico occidental. Destaca como especie importante de ambientes estuarinos por su gran abundancia numérica y biomasa durante gran parte del año en el Golfo de México, sin embargo, se reporta que su presencia en esos ambientes es por tallas juveniles (Aguirre León et al., 1982 y Yáñez-Arancibiaet al. 1983). 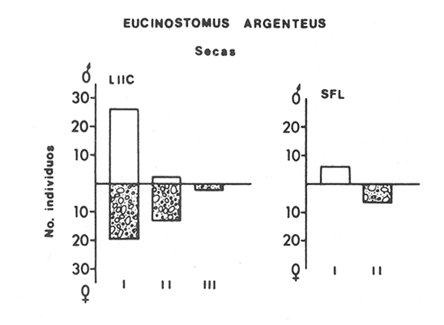 Fig. 20. Fases de desarrollo sexual y proporción de sexos de Eucinostomus argenteus durante la época de secas en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. 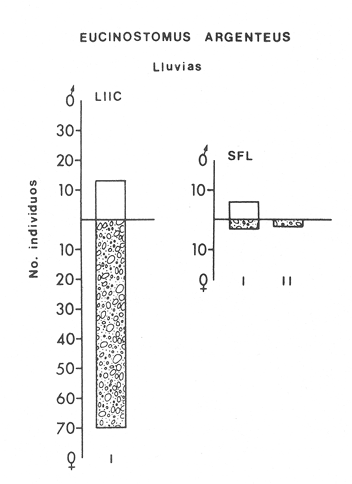 Fig. 21. Fases de desarrollo sexual y proporción de sexos de Eucinostomus argenteus durante la época de lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares.  Fig. 22. Relación de la longitud total y las fases de desarrollo sexual de Eucinostomus argenteus independiente del sexo durante las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. No obstante, es interesante citar que Cervigón (1966) ha reportado tallas adultas hasta 400 mm de longitud total en aguas estuarinas de Venezuela; siendo 115 mm la mayor talla en este estudio. Publicaciones recientes en el Sur del Golfo de México destacan el papel que juega esta especie en la estructura de las comunidades de peces de la Laguna de Términos (Amezcua Linares y YáñezArancibia, 1980; Yáñez-Arancibia et al., 1980; Vargas Maldonado et al. 1981; Yañez-Arancibia et al, 1982a; Aguirre Leónet al., 1982 y Yáñez-Arancibia y Lara Domínguez, 1983). Diapterus rhombeus es la segunda especie más abundante de las mojarras de la Laguna de Términos. Utiliza la laguna como área de crianza y alimentación y es consumidor de primer orden. Se ha colectado en toda la laguna obteniéndose 547 ejemplares de 40 a 115 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación, los valores calculados para las constantes a y b son dados en función de la época climática. Asimismo el análisis del rango de tallas se presenta comparativamente para las dos épocas climáticas y los dos subsistemas estudiados (Figs. 30 a 34). Se observa que predominan las tallas juveniles en los dos períodos climáticos, reflejándose ésto en las modas. Sin embargo las tallas más grandes están mejor representadas en el litoral interno de la Isla del Carmen en ambas épocas climáticas. Comparando los dos subsistemas se aprecia que las tallas menores se encontraron en los Sistemas Fluvio Lagunares en el período de lluvias. 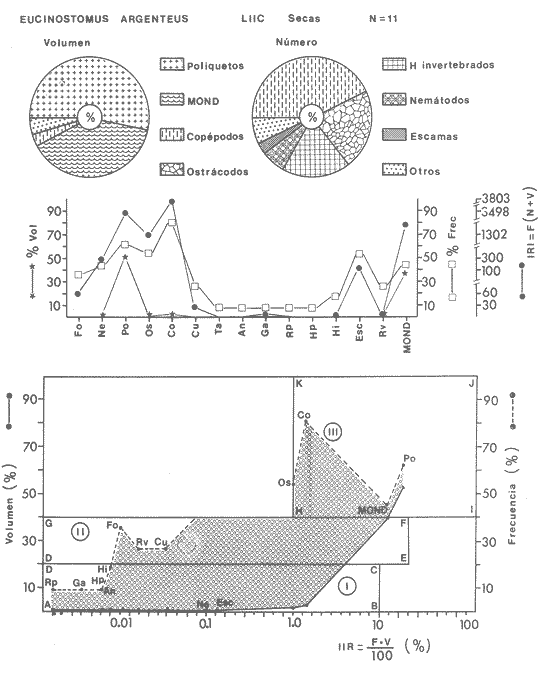 Fig. 23. Espectro trófico de la población de Eucinostomus argenteus en el Litoral Interno de la Isla del Carmen durante la época de secas. Ver anexo de abreviaturas en Metodología.  Fig. 24. Espectro trófico de la población de Eucinostomus argenteus en los Sistemas Fluvio-Lagunares durante la época de secas. Ver anexo de abreviaturas en Metodología.  Fig. 25. Espectro trófico de la población de Eucinostomus argenteus en el Litoral Interno de la Isla del Carmen durante la época de lluvias. Ver anexo de abreviaturas en Metodología. 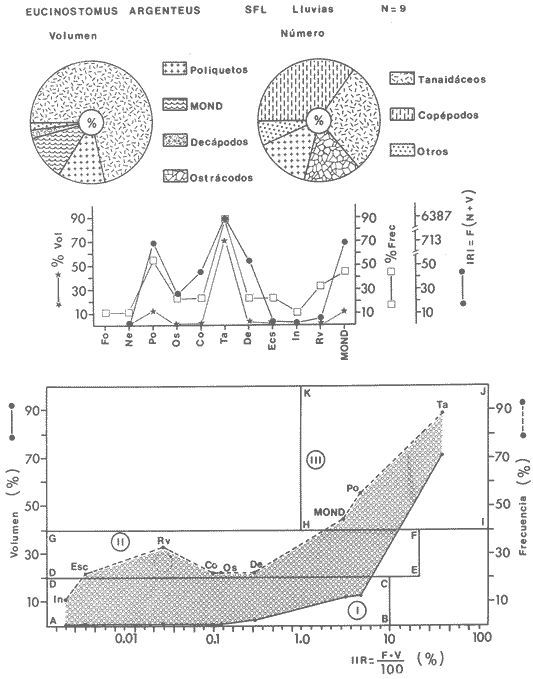 Fig. 26. Espectro trófico de la población de Eucinostomus argenteus en los Sistemas Fluvio-Lagunares durante la época de lluvias. Ver anexo de abreviaturas en Metodología, 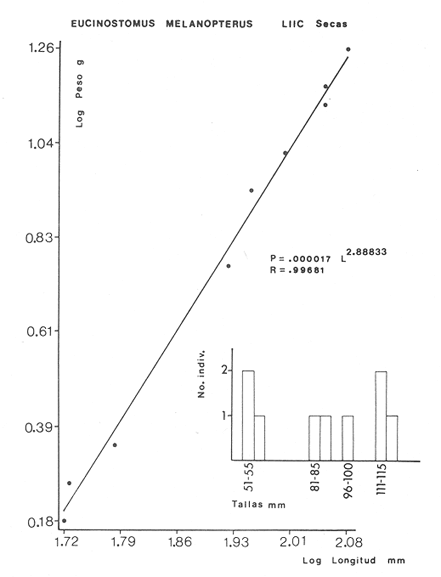 Fig. 27. Recta de regresión de la relación peso/longitud de Eucinostomus melanopterus en el Litoral Interno de la Isla del Carmen durante la época de secas. Se señala la distribución de frecuencia de tallas. 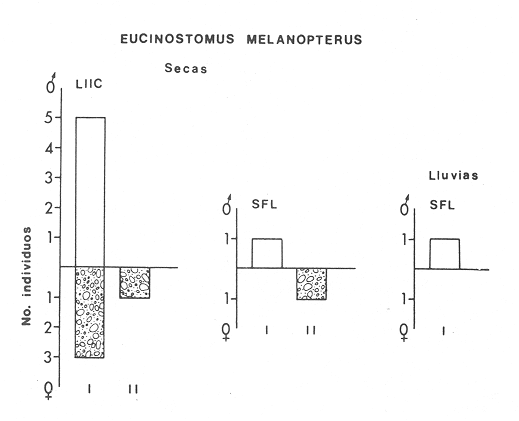 Fig. 28. Fases de desarrollo sexual y proporción de sexos de Eucinostomus melanopterus en las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares.  Fig. 29. Relación de la longitud total y las fases de desarrollo sexual de Eucinostomus melanopterus independiente del sexo durante la época de secas en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. 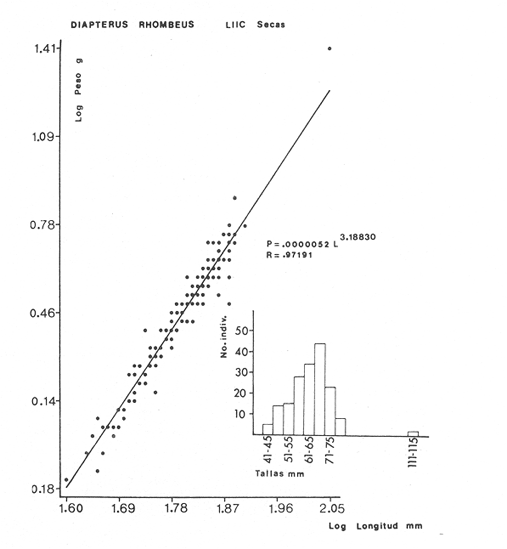 Fig. 30. Recta de regresión de la relación peso/longitud de Diapterus rhombeus en el litoral Interno de la Isla del Carmen durante la época de secas. Se señala la distribución de frecuencia de tallas.  Fig. 31. Recta de regresión de la relación peso/longitud de Diapterus rhombeus en el Litoral Interno de la Isla del Carmen durante la época de lluvias. Se señala la distribución de frecuencia de tallas. Madurez. La frecuencia de sexos y f ases sexuales en función de la época climática se muestra graficada en las figuras 35 y 36. Se observa que para los períodos climáticos definidos, predominan los estadíos juveniles en fase I y II, presentándose pocos ejemplares en fase III en el litoral interno de la Isla del Carmen en la época de lluvias y en los Sistemas Fluvio-Lagunares en la época de secas, por lo tanto, no aparecen individuos en reproducción. Las fases I y II están representadas tanto por machos como por hembras, presentándose la fase III sólo en hembras. 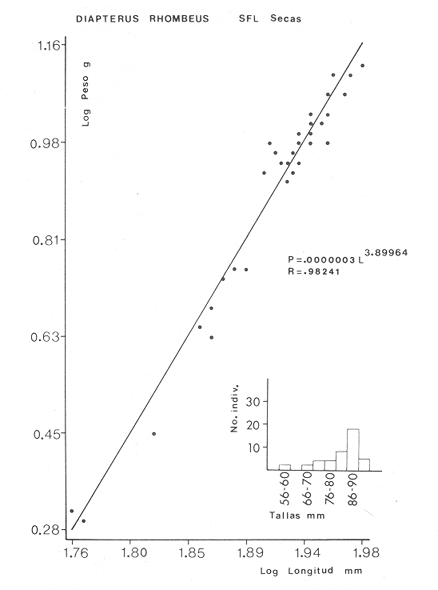 Fig. 32. Recta de regresión de la relación peso/longitud de Diapterus rhombeus en los Sistemas Fluvio-Lagunares durante la época de secas. Se señala la distribución de frecuencias de tallas. Del análisis de proporción de sexos macho: hembra se encontró que el total de la población guarda una proporción de 1.1: 1.0; siendo para la época de secas de 1.0: 1.3 y durante el período de lluvias de 1.4: 1.0. La figura 37 muestra la relación de la madurez gonádica y la talla de los individuos así como la relación total del desarrollo sexual de la población independiente del sexo, lo cual es fundamental para determinar la longitud a la cual dicha población alcanza la madurez gonádica y es apta para reproducirse. Se observa que durante la época de lluvias se encuentran individuos en maduración desde los 120 mm de longitud total en fase III. La fase I está comprendida entre los 40 y 85 mm. No se registraron individuos en fase II en este período.  Fig. 33. Recta de regresión de la relación peso/longitud de Diapterus rhombeus en los Sistemas Fluvio-Lagunares durante la época de lluvias. Se señala la distribución de frecuencia de tallas. Durante la época de secas se encontraron ejemplares maduros desde los 120 mm en fase III, apareciendo también las fases I y II, entre los 40 y 112 mm para la primera y desde los 87 mm para la segunda. Alimentación. El estudio de las relaciones tróficas de Diapterus rhombeus se basó en el análisis del contenido estomacal de ejemplares juveniles (menos de 120 m m) en función de las localidades y la época climática. Litoral interno de la Isla del Carmen. Epoca de secas. El espectro trófico de D. rhombeus (Fig. 38) indica que se alimenta al menos de 17 grupos tróficos entre los que destacan como importantes: MOND con un 66 % en volumen y 100 % de frecuencia, copépodos con un 19.7 % en volumen y 100 % de frecuencia, nemátodos con un 2.6 % en volumen y 100 % de frecuencia, restos vegetales con un 7.3 % en volumen y 80 % de frecuencia y foraminiferos con 1 % en volumen y 86.6 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con un 69.8 %, nemátodos con 11. 6 %, ostrácodos con 8.2 % y foraminíferos en menor proporción. De acuerdo al índice IRI destacan, copépodos, MOND, nemátodos, ostrácodos y foraminíferos con los valores más altos. En función del índice IIR se encuentra que la materia orgánica es el alimento preferencial con un 66 %, los copépodos se consideran como alimento secundario y los 15 grupos restantes como circunstanciales. 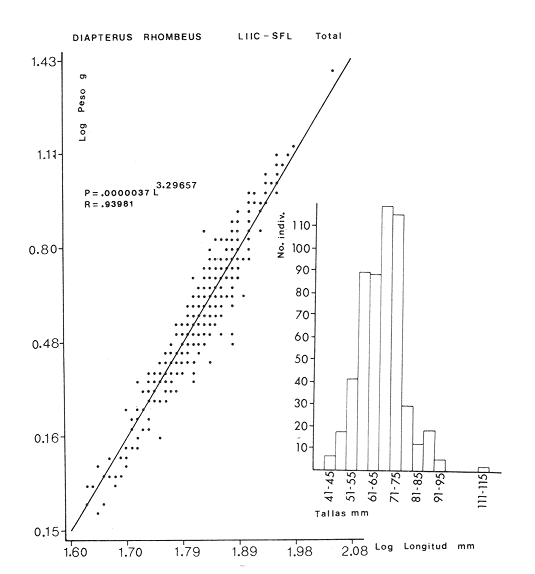 Fig. 34. Recta de regresión de la relación peso/longitud de Diapterus rhombeus para las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Se señala la distribución de frecuencia de tallas. Sistemas Fluvio-Lagunares. Epoca de secas. El espectro trófico (Fig. 39) indica que se alimenta al menos de 13 grupos tróficos entre los que destacan como importantes: poliquetos con un 48.4 % en volumen y 50 % de frecuencia, MONDcon un 38.3 % en volumen y 83.3 % de frecuencia, copépodos con un 5.1 % en volumen y 58.3 % de frecuencia y restos vegetales con un 4.7 % en volumen y 58.3 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con 35.5 %, huevos de invertebrados con 13.3 %, ostrácodos con 13.2 % y foraminíferos en menor proporción. De acuerdo al índice IRI destacan: copépodos, MOND, ostrácodos, nemátodos, huevos de invertebrados y foraminíferos con los valores más altos. 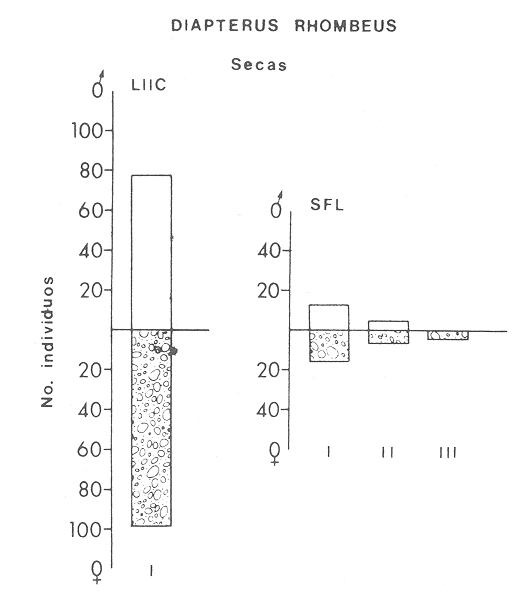 Fig. 35. Fases de desarrollo sexual y proporción de sexos de Diapterus rhombeus durante la época de secas en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Litoral interno de la Isla del Carmen. Epoca de lluvias. El espectro trófico en esta época (Fig. 40) indica que se alimenta al menos de 8 grupos tróficos entre los que destacan como importantes: MOND con un 75 % en volumen y 100 % de frecuencia, copépodos con 13 % en volumen y 100 % de frecuencia, foraminíferos con un 5.1 % en volumen y 100 % de frecuencia, otros grupos como nemátodos, ostrácodos y restos vegetales se presentan con menor proporción volumétrica, sin embargo se les encuentra con un 100 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con 59.8 %, foraminíferos con 22.4 %, nemátodos con 9.2 %, gasterópodos con 5.1 % y ostrácodos en menor proporción. De acuerdo al índice IRI destacan: MOND, copépodos, foraminíferos, nemátodos, ostrácodos y gasterópodos con los valores más altos. En función del índice IIR se encuentra que la materia orgánica es el alimento preferencial, se considera como grupo secundario a los copépodos y los 6 grupos restantes como circunstanciales.  Fig. 36. Fases de desarrollo sexual y proporción de sexos de Diapterus rhombeus durante la época de lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Sistemas Fluvio-Lagunares. Epoca de lluvias El espectro trófico (Fig. 41) indica que se alimenta al menos de 20 grupos tróficos entre los que destacan como importantes: MOND con un 68.8 % en volumen y 100 % de frecuencia, copépodos con un 13 % en volumen y 100 % de frecuencia, ostrácodos con un 5.9 % en volumen y 100 % de frecuencia, restos vegetales con un 1.9% en volumen y 78.9% de frecuencia, nemátodos con 1. 6 % en volumen y 89.4 % de frecuencia y cumáceos con un 3.4 % en volumen y 68.4 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con 57 %, ostrácodos con 19.6 %, nemátodos con 8.4 %, foraminíferos con 6.1 % y cumáceos con 3.5 %. De acuerdo al índice IRI destacan como importantes los copépodos, MOND, ostrácodos, nemátodos, foraminíferos con los valores más altos. En función del índice IIR se encuentra que la materia orgánica es el alimento preferencial con 68.8 % se considera como grupo secundario a los copépodos y los 9 grupos restantes se consideran como ocasionales.  Fig. 37. Relación de la longitud total y las fases de desarrollo sexual Diapterus rhombeus durante la época de secas y lluvias en el Litoral Interno de la Isla del Carmen y los Sistemas Fluvio-Lagunares. Diapterus auratus Ranzani. 1842Es una especie típica en ambientes lagunares y estuarinos del Golfo de México y Mar Caribe. Sin embargo, existen muy pocos estudios sobre la biología de esta especie. Diversos trabajos la citan dentro de sus listas ictiofaunísticas a lo largo de su amplia distribución en el Atlántico occidental. Destaca como especie importante de ambientes estuarinos por su gran abundancia numérica y biomasa a lo largo de todo el año, por lo que representa un recurso pesquero. Su presencia en esos ambientes está representada por tallas juveniles y adultas. Al igual que otras especies afines carece de estudios sistemáticos sobre su biología y ecología. D. auratus es una especie rara en la Laguna de Términos. Un sinónimo de esta especie (D. olisthostomus) ha sido reportada con anterioridad para la laguna por Reséndez (1981) y Aguirre León et al. (1982). Es una especie visitante ocasional y consumidor de primer orden. En el área de estudio se ha colectado en el Litoral interno de la Isla del Carmen y en el Sistema Fluvio-Lagunar Chumpan-Balchacah. Se obtuvieron 25 ejemplares de 30 a 105 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación, los valores calculados para las constantes a y b sólo corresponden al litoral interno de la Isla del Carmen en la época de secas. Asimismo el análisis del rango de tallas se presenta en la figura 42. Sólo aparecen tallas juveniles en el período de secas, representadas por pocos ejemplares. Madurez. La frecuencia de sexos y fases sexuales en función de la época climática se muestra graficado en la figura 43. Se observa en general que para las épocas climáticas definidas sólo se presentan estadios juveniles en fase 1 en el litoral interno de la Isla del Carmen, por lo tanto no aparecen individuos maduros. La fase I está representada tanto por machos como por hembras. Se encontraron individuos indiferenciados en sexo y maduración en el litoral interno de la Isla del Carmen en la época de secas. El análisis de proporción de sexos macho; hembra hecho para los ejemplares colectados en el litoral interno de la Isla del Carmen fue de 1.6: 1.0. Alimentación. El estudio de las relaciones tróficas de Diapterus auratus se basó en el análisis del contenido estomacal de ejemplares juveniles de menos de 70 mm, en función de la localidad y la época climática estudiada.  Fig. 38. Espectro trófico de la población de Diapterus rhombeus en el Litoral Interno de la Isla del Carmen durante la época de secas.Ver anexo de breviaturas en Metodología. 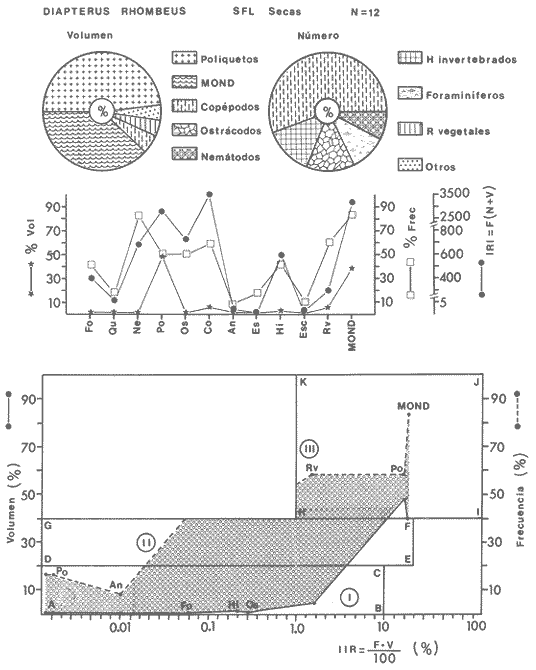 Fig. 39. Espectro trófico de la población de Diapterus rhombeus en los Sistemas Fluvio-Lagunares durante la época de secas. Ver anexo de abreviaturas en Metodología.  Fig. 40. Espectro trófico de la población de Diapterus rhombeus en el Litoral Interno de la Isla del Carmen durante la época de lluvias. Ver anexo de abreviaturas en Metodología.  Fig. 41. Espectro trófico de la población de Diapterus rhombeus en los Sistemas Fluvio-Lagunares durante la época de lluvias. Ver anexo de abreviaturas en Metodología.  Fig. 42. Recta de regresión de la relación peso/longitud de Diapterus auratus en el Litoral Interno de la Isla del Carmen durante la época de secas. Se señala la distribución de frecuencias de tallas. Litoral interno de la Isla del Carmen. Epoca de secas. El espectro trófico deD. auratus (Fig. 44) indica que se alimenta al menos de 11 grupos tróficos entre los que destacan como importantes: MOND con un 82.4 % en volumen y 95.8% en frecuencia, copépodos con un 7.3% en volumen y 95.8 % de frecuencia, restos vegetales con un 8.1 % en volumen y 45.8 % de frecuencia. Numéricamente los grupos predominantes son: copépodos con 80.7 %, ostrácodos con 8.6% y nemátodos con 7.6%. De acuerdo al índice IRI destacan, copépodos, MOND, nemátodos y ostrácodos. En función del índiceIIR se encuentra que la materia orgánica es el alimento principal con 78.9 %, y los 10 grupos restantes son circunstanciales.  Fig. 43. Fases de desarrollo sexual y proporción de sexos de Diapterus auratus durante la época de secas en el Litoral de la Isla del Carmen. Eugerres plumieri (Cuvier, 1830)Eugerres plumieri es una especie típica en ambientes lagunares y estuarinos del Golfo de México y Mar Caribe. Numerosos trabajos la citan dentro de sus listas ictiofaunísticas a lo largo de su amplia distribución en el Atlántico de América. Destaca como especie importante de ambientes estuarinos por su gran abundancia numérica y biomasa a lo largo de todo el año, por lo que es un recurso comercial en muchas lagunas costeras del Golfo. Su presencia en esos ambientes está representada por tallas juveniles y adultos. Carece al igual que otras especies afines, de estudios sobre su biología y ecología. 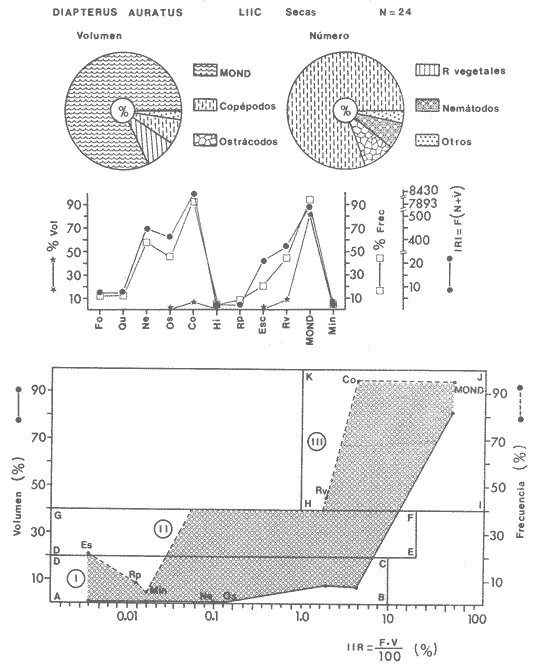 Fig. 44. Espectro trófico de la población de Diapterus auratus en el Litoral Interno de la Isla del Carmen durante la época de secas. Ver anexo de abreviaturas en Metodología.  Fig. 45. Recta de regresión de la relación peso/longittid de Eugerres plumieri en el Litoral Interno de la Isla del Carmen durante las épocas de secas. Se señala la distribución de frecuencias de tallas. E. plumieries la cuarta especie de mojarra más abundante en la Laguna de Términos. Constituye un recurso económico en el área. Utiliza la laguna como área de crianza y/o alimentación y es consumidor de primer orden. Se ha colectado en las localidades PE, CHB, PG, ESP y CUC. Se obtuvieron 59 ejemplares de 65 a 295 mm de longitud total. Relación peso-longitud y frecuencia de tallas. De acuerdo a la ecuación que define esta relación, los valores calculados para las constantes a y b son dados en función de la época climática. Asimismo el análisis del rango de tallas se presenta comparativamente para las dos épocas climáticas y los dos subsistemas estudiados (Figs. 45 a 48). Se observa que se presentan con más frecuencia tallas subadultas y adultas en ambas épocas climáticas reflejándose en las modas. Sin embargo, las tallas mas grandes están mejor representadas en el litoral interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares en la época de secas, encontrándose las mas pequeñas en el período de lluvias.  Fig. 46. Recta de regresión de la relación peso/longitud de Eugerres plumieri en el Litoral Interno de la Isla del Carmen durante la época de lluvias Se señala la distribución de frecuencias de tallas. Madurez. La frecuencia de sexos y fases sexuales en función de la época climática se muestra graficado en las figuras 49 y 50. Se observa que para los períodos climáticos definidos se presentan las fases I, II y III en la época de lluvias, encontrándose que para el período de secas aparecen además las fases IV, VI y VII, por lo tanto aparecen individuos en las fases reproductivas. Las fases maduras están representadas solo por hembras, encontrándose en los machos sólo fases I y II.  Fig. 47. Recta de regresión de la relación peso/longitud de Eugerres plumieri en los Sistemas Fluvio-Lagunares durante la época de lluvias. Se señala la distribución de frecuencias de tallas. Del análisis de proporción de sexos macho: hembra se encontró que el total de la población guarda una proporción de 1. 1: 1. 0, siendo para la época de secas de 1.5: 1.0 y para el período de lluvias de 1. 0: 1. 1. La figura 51 muestra la relación de la madurez gonádica y la talla de los individuos, así como la relación total del desarrollo sexual de la población independiente del sexo, lo cual es fundamental para determinar la longitud a la cual dicha población alcanza la madurez gonádica y es apta para reproducirse. Se aprecia que durante la época de lluvias se encuentran individuos en maduración desde los 184 mm de longitud total en fase III. La fase I está comprendida entre los 59 y 132 mm y la fase II entre los 130 y 180 mm. Durante el período de secas no se encuentran individuos en fase III, sin embargo, la fase IV se registra desde los 205 mm de longitud total. Asimismo las fases postreproductivas y en descanso están presentes desde los 260 mm. No obstante, siguen encontrándose fases I y II. Es importante destacar que los individuos maduros y en fases postreproductivas se registraron con mayor frecuencia en el litoral interno de la Isla del Carmen en el período de secas cuando la salinidad presenta valores elevados (mayores de 26%) Alimentación El estudio de las relaciones tróficas de Eugerres plumieri se basó en el análisis del contenido estomacal de ejemplares juveniles y adultos en función de las localidades estudiadas y la época climática. Litoral interno de la Isla del Carmen. Epoca de secas. El espectro trófico de E. plumieri(Fig. 52) indica que se alimenta al menos de 19 grupos tróficos entre los que destacan como importantes: MOND con un 65.7% en volumen y 100 % de frecuencia, ostrácodos con 16. 1 % en volumen y 82.3 % de frecuencia, restos vegetales con 11. 3 % en volumen y 82.3 % de frecuencia y copépodos con 2.6 % en volumen y 70.5 % de frecuencia. Numéricamente los grupos predominantes son ostrácodos con 66 %, copépodos con 18.6 % foraminíferos con 9.8 % y nemátodos con 3.3 %. De acuerdo al análisis combinado del índice de importancia relativa IRI destacan ostrácodos, MOND, copépodos, restos vegetales, foraminíferos y nemátodos con los valores más altos. En función del índice IIR se puede establecer que el alimento preferencial es la materia orgánica con 65.7 %, se considera alimento secundario a los copépodos, y los 17 grupos restantes como circunstanciales. Sistemas Fluvio-Lagunares. Epoca de lluvias. El espectro trófico deE. plumieri (Fig. 53) indica que se alimenta al menos de 18 grupos entre los que destacan como importantes: pelecípodos con 44.1 % en volumen y 68.7 de frecuencia, MOND con 31 % en volumen y 100 % de frecuencia, copépodos con 13.2 % en volumen y 75 % de frecuencia y ostrácodos con 6.8 % en volumen y 62.5 % de frecuencia. Numéricamente los grupos predominantes son copépodos con 87.9 %, huevos de invertebrados con 6.1 % y ostrácodos con 2.0 %. De acuerdo al análisis combinado del índice de importancia relativa IRI destacan copépodos, MOND, restos de pelecípodos y ostrácodos. En función del índice IIR se puede establecer que el alimento preferencial es la materia orgánica (MOND) y los pelecípodos, como secundario se considera a los copépodos y los grupos restantes como circunstanciales. 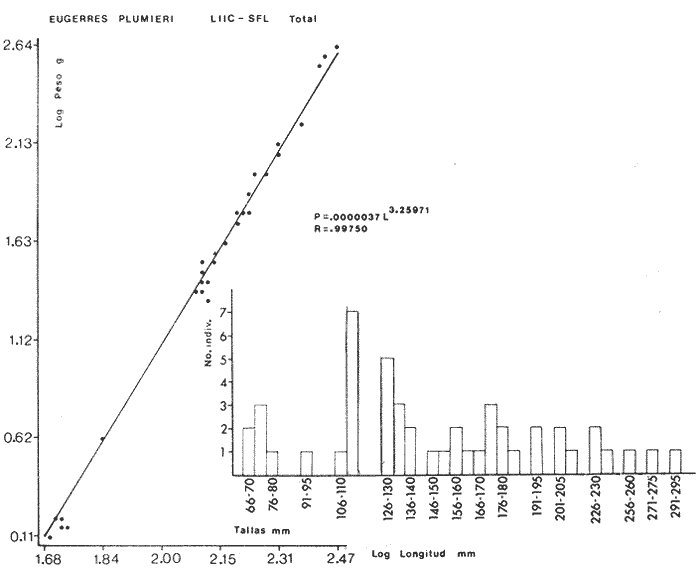 Fig. 48. Recta de regresión de la relación peso/longitud de Eugerres plumieri para las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. Se señala la distribución de frecuencias de tallas. DISCUSIÓN
RELACION PESO-LONGITUD Y FRECUENCIAS DE TALLASLa ecuación P = aLb describe las variaciones de peso y longitud a lo largo del cielo de vida de un pez. Esta expresión puede aplicarse para entender el comportamiento de estos parámetros en una población en función del tiempo. Además tiene aplicación en la evaluación de pesquerías debido al interés de conocer tasas de crecimiento, es decir, el aumento en peso y longitud por unidad de tiempo. El coeficiente exponencial b en la ecuación antes descrita representa la pendiente de la línea recta ajustada por el método de regresión. Biológicamente el exponente brepresenta un coeficiente alométrico cuyas variaciones están en función de los cambios proporcionales de peso y longitud de una especie en cuestión. Así, cuando el coeficiente btiene un valor de 3 describe un crecimiento ísometrico. Muchas especies parecen acercarse a este "ideal", sin embargo, el peso es afectado por factores tales como la época del año, edad del pez, variaciones en la alimentación, desove, sexo y madurez sexual, acumulación de ácidos grasos en la cavidad corporal entre otros. Otras especies presentan valores más grandes o más pequeños que 3, una condición descrita como crecimiento alométrico. Cuando el coeficiente b es mayor a 3, se dice que una especie presenta un crecimiento alométrico positivo, mientras que, valores deb menores a 3, representan un crecimiento alométrico negativo (Ricker, 1975). La constante a expresada en la ecuación de relación peso-longitud representa la corrección de la ordenada al origen de la regresión lineal.  Fig. 49. Fases de desarrollo sexual y proporción de sexos de Eugerres plumieri durante la época de secas en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares.  Fig. 50. Fases de desarrollo sexual y proporción de sexos de Eugerres plumieri durante la época de lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares.  Fig. 51. Relación de la longitud total y las fases de desarrollo sexual de Eugerres plumieri independiente del sexo durante las épocas de secas y lluvias en el Litoral Interno de la Isla del Carmen y en los Sistemas Fluvio-Lagunares. A partir del análisis de las frecuencias de tallas se puede obtener información de: la composición de las clases de tallas, los valores de los picos (curvas polimodales, gráficamente, y la época de reclutamiento (cuando este ocurre en una época precisa) de cada mes, entre otros.  Fig. 52. Espectro trófico de la población de Eugerres plumieri en el Litoral Interno de la Isla del Carmen durante la época de secas. Ver anexo de abreviaturas en Metodología. 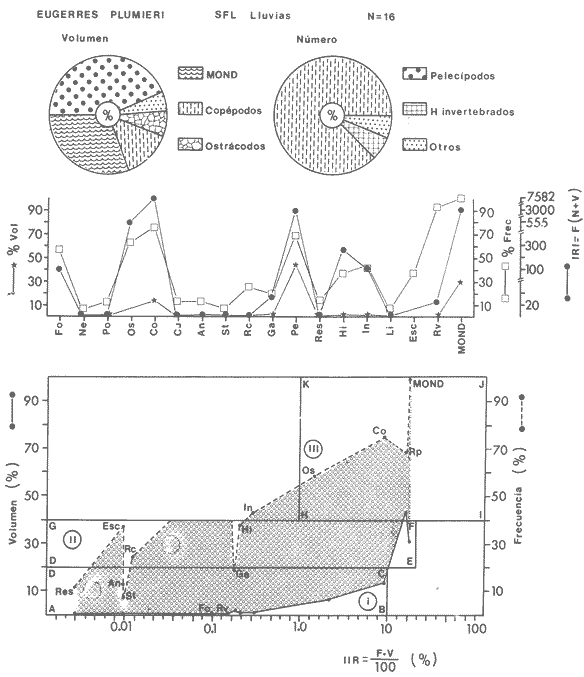 Fig. 53. Espectro trófico de la población de Eugerres plumieri en los Sistemas Fluvio-Lagunares durante la época de lluvias. Ver anexo de abreviaturas en Metodología. La Tabla 3, resume comparativamente los valores de los coeficientes a yb de todas las especies por época climática y subsistema. De igual manera, la tabla 4 resume el comportamiento de los rangos de clases de talla de las poblaciones a lo largo del año. En relación a los resultados presentados en estas dos tablas se encontró de manera general para todas las especies que las variaciones en el valor del coeficiente b de la relación pesolongitud se deben a las diferencias en los rangos de las clases de talla en función de la localidad y la época climática. Para poder comparar los resultados obtenidos de la relación peso-longitud de todas las especies se consideró conveniente uniformizar el rango de las clases de talla de cada una de las especies estudiadas en relación al mínimo y al máximo rango de talla encontrado durante el año para los dos subsistemas analizados. Los valores del coeficiente b encontrados para Eucinostomus gula reflejaron pequeñas variaciones a lo largo del año (Figs. 3 a 7) tanto en el litoral interno de la Isla del Carmen como en los sistemas fluvio-lagunares. En el litoral interno de la Isla durante ambas épocas climáticas el coeficiente b varió de 3.0 a 3.1 en comparación a los sistemas fluviolagunares donde fue de 2.9. En el primer subsistema el comportamiento del coeficiente b podría deberse a que la especie tuviera una mejor alimentación que en los sistemas fluviolagunres. Esto último debido a una mayor disponibilidad de alimento a lo largo del año, ya que el litoral interno de la Isla se caracteriza por presentar extensas praderas de pastos marinos en las cuales se encuentra asociada por sustrato una gran diversidad de macro y microfauna béntica. Además, en este subsistema se registran en ambas épocas climáticas individuos maduros que contribuyen con mayor peso (por el aumento en el tamaño de las gónadas) a la biomasa total de la población. Eucinostomus gula ingresa a la laguna con mucha probabilidad durante primavera y verano (fin de la época de secas y época de lluvias) al estado juvenil (menos de 30 mm de longitud total) y larvario. Fases larvarias deEucinostomus sp. han sido registradas abundantemente durante primavera y verano principalmente hacia la región este de la Laguna de Términos (Flores-Coto y Alvarez-Cadena, 1980). Las poblaciones de esta especie que migran a la laguna colonizan las áreas protegidas de manglar de los sistemas fluvio-lagunares y del litoral interno de la Isla del Carmen, encontrándose durante la época de lluvias poblaciones juveniles con rangos de talla entre 30 y 125 mm de longitud total (Tabla 4). En relación al análisis de frecuencia de clases de talla se encontró en el litoral interno de la Isla del Carmen una clase modal (sólo se consideró el valor máximo de las modas) de 61-65 mm, mientras que en los sistemas fluviolagunares fue de 71-75 mm (Tabla 4, Figs. 4 y 6). E. gula estuvo bien representada durante la época lluviosa. Sin embargo, se trata de una población de peces pequeños. Durante la época de secas, las poblaciones encontradas en el litoral interno de la Isla y en los sistemas fluvio-lagunares presentaron rangos de talla que variaron entre 30 y 160 mm de longitud total (Tabla 4). Las clases modales son para el primer subsistema de 71-75 mm y para el segundo de 86-90 mm (Tabla 4, Figs. 3 y 5). La especie estuvo bien representada durante el período de secas. Sin embargo, a diferencia de la época de lluvias, se encontraron en la de secas individuos maduros con mayor peso. La distribución total de la suma de frecuencias de las clases de talla para ambas épocas climáticas en los dos subsistemas (Fig. 7) muestra que la clase modal de 61-65 mm fue la más frecuente. Este último comportamiento podría deberse a la selectividad del arte de pesca utilizado (red de prueba camaronera) ya que, como se observa en la Tabla 4 y en las figuras 3 a 7 la especie es capturada a partir de los 30 mm de longitud total y con mayor frecuencia clases de talla juveniles. Es importante destacar también que la alta frecuencia de juveniles se debe a que los subsistemas ecológicos de la Laguna de Términos (principalmente el litoral interno de la Isla del Carmen) son utilizados como áreas de crianza. Lo anterior podría deberse a que las poblaciones de E. gula que ingresan principalmente durante la época de lluvias, crecen a lo largo de esta época, maduran y ganan peso en la época de secas para migrar hacía finales de la misma a la plataforma continental adyacente a desovar, probablemente en aguas someras con vegetación sumergida en la costa del Estado de Campeche para reiniciarse un nuevo ciclo en la laguna con la entrada de larvas y juveniles en primavera y verano. Por otra parte, este mismo comportamiento, presencia de poblaciones juveniles y patrones de migración ha sido estudiado por Springer y Woodburn (1960) en la Bahía de Tampa y por Springer y Mc Erlean (1962) en los Cayos de Matecumbe, ambos estudios de Florida, donde reportan abundantes poblaciones de individuos jóvenes a lo largo del año. Es probable que este patrón de comportamiento se realice en otras lagunas costeras del Golfo de México, sin embargo, no existen estudios al respecto. El planteamiento anterior se ve reforzado por los estudios realizados en la Sonda de Campeche adyacente a la Laguna de Términos por Sánchez-Gilet al. (1981) donde se analizan los patrones poblacionales de E. gula en verano. Las poblaciones que allí se encuentran están constituidas principalmente por individuos subadultos y adultos, siendo razonable suponer que se trata de poblaciones maduras. 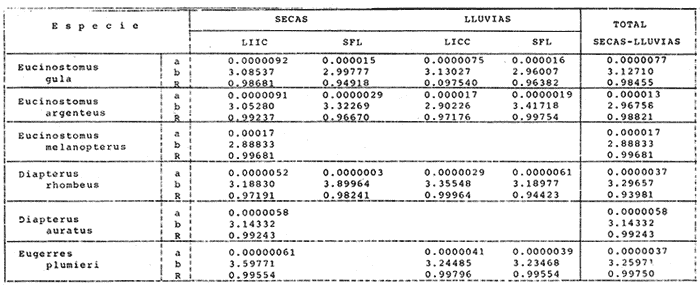 TABLA 3 VALORES DE LAS CONSTANTES A Y B Y EL COEFICIENTE DE CORRELACIÓN R POR ESPECIE,LOCALIDAD Y EPOCA CLIMATICA 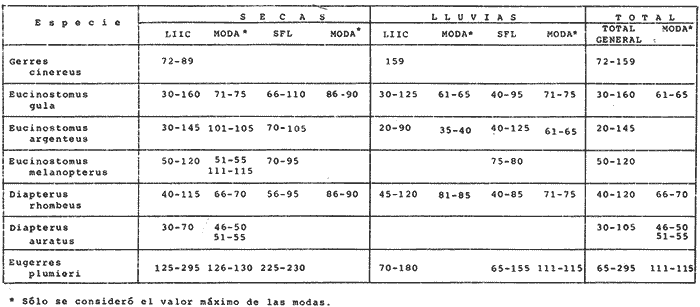 TABLA 4 RANGOS DE TALLA POR ESPECIE, LOCALIDAD Y ÉPOCA CLIMÁTICA Los valores del coeficienteb calculados para Eucinostomus argenteusvariaron de acuerdo con la época del año y la localidad (Tabla 3, Figs. 15 a l9). Durante las épocas de secas y lluvias en el litoral interno de la Isla del Carmen el coeficiente bvarió de 2.9 a 3.0 mientras que en los sistemas fluviolagunares fue de 3.3 a 3.4 (Tabla 3). Podría pensarse que las diferencias encontradas en el comportamiento del coeficiente b de un subsistema a otro se debieran a una mejor alimentación en los sistemas fluvio-lagunares. Sin embargo, en este último subsistema se observa que la distribución de las clases de talla de los rangos encontrados (Tabla 4, Figs. 15 a 19) no es homogénea, por lo que, las variaciones del coeficiente b estarían mas bien dadas por las diferencias en los rangos de talla y no por diferencias en la alimentación. E. argenteus se encuentra representado en la laguna principalmente por individuos juveniles. Durante la época de secas en el litoral interno de la Isla del Carmen se encontraron individuos de tallas entre 30 y 145 mm de longitud total (Tabla 4, Fig. 15). En relación a la figura 15 se observa en el histograma de frecuencia de tallas que se diferencian al menos cuatro clases modales, 41-45; 71-75; 101-105 y 131-135 mm. Mientras que, en la época de lluvias en el mismo subsistema solo se observa una clase modal de 36-40 mm. En los sistemas fluvio-lagunares (Figs. 16 y 18) se encontraron individuos entre 70 a 95 mm durante la época de secas y de 40 a 125 mm en la de lluvias, sin embargo, en ambas épocas climáticas no se observan clases modales o al menos no están bien definidas. De lo anterior podría pensarse que el litoral interno de la Isla del Carmen ofrece condiciones más propicias tanto ambientales como ecológicas a esta especie. La distribución total de las clases de talla de la población en toda la laguna (Fig. 19) refleja nuevamente una separación de las clases modales encontradas en el litoral interno de la Isla del Carmen durante la época de secas. No obstante, es importante observar en la gráfica que la clase modal 36-40 mm fue la que presentó la mayor frecuencia. Puesto que E. argenteus presenta un patrón de comportamiento estacional o cíclico en la laguna, la composición de clases de talla varía a lo largo del año. La laguna es utilizada por esta especie como un área de crianza y alimentación. Un comportamiento similar con presencia principalmente de poblaciones juveniles ha sido observado en otras lagunas costeras del Golfo de México (Chávez, 1972), por Springer y Woodburn (1960) y por Springer y Me Earlen (1962) en estuarios de Florida, donde nuevamente encuentran un patrón cíclico de la especie reflejándose en la estructura de la población. Sánchez-Gil et al. (1981) ha observado también un comportamiento estacional de E. argenteus en la Sonda de Campeche adyacente a la Laguna de Términos. Eucinostomus melanopterus es una especie poco frecuente en la Laguna de Términos por lo que no pudo realizarse un análisis comparativo de la relación peso-longitud a lo largo del año (Tabla 3). Aunque estuvo poco representada en la laguna es más abundante en el litoral interno de la Isla del Carmen. Dada la baja abundancia de la especie en la laguna, es razonable suponer que su presencia en ella se deba a necesidades tróficas, ya que fue encontrada mejor distribuida en las áreas de pastos marinos del litoral interno de la Isla del Carmen donde existe buena disponibilidad de alimento y salinidades mayores a 26 ppm. La población de E. melanopterus está constituida principalmente por individuos juveniles con tallas entre 50 y 120 mm, y en fases tempranas de maduración gonádica. Esta especie ha sido reportada en otras lagunas costeras del Golfo de México en rangos de talla mayores con mayor abundancia (Reséndez, 1970 y 1973). En estudios posteriores Reséndez (1981 b) ha reportado a esta especie en la Laguna de Términos también con baja abundancia y en tallas juveniles. En un estudio realizado en la Sonda de Campeche por Sánchez-Gil et al. (1981) señalan que E. melanopterusestá representada principalmente por individuos juveniles. Los valores del coeficiente bcalculados para Diapterus rhombeus variaron con la época del ano y la localidad (Tabla 3, Figs. 30 a 34). Durante las épocas de secas y lluvias en el litoral interno de la Isla del Carmen y en los sistemas fluviolagunares el coeficienteb vario de 3.1 a 3.8. Analizando la Tabla 3 se observa que los valores más altos de b se encontraron en los sistemas fluviolagunares en la época de secas (b1 = 3.8) y en el litoral interno de la Isla en la época de lluvias (b = 3.3). En estos dos subsistemas y en las dos épocas climáticas fueron registrados individuos adultos y maduros (Figs. 35 y 36). Las poblaciones de D. rhombeus que se encuentran en la laguna a lo largo del año son en su mayoría juveniles. Se encontró representado durante la época de lluvias en el litoral interno de la Isla del Carmen en rangos de talla de 45 a 120 mm de longitud total (Tabla 4) y en los sistemas fluvio-lagunares entre 40 y 85 mm, sin embargo, la población de la Isla del Carmen estuvo pobremente representada. En los sistemas fluviolagunares se encontró una clase modal de 7175 mm (Tabla 4, Fig. 33) y la especie se encontró bien representada. Durante la época de secas las poblaciones encontradas en el litoral interno de la Isla del Carmen y en los sistemas fluviolagunares presentaron rangos de clases de talla que variaron entre 40 y 115 mm (Tabla 4). Las clases modales fueron para el primer subsistema de 66-70 mm y para el segundo de 86-90 mm (Tabla 4, Figs. 30 y 31). En general, la especie estuvo mejor representada en los sistemas fluviolagunares que en el litoral interno de la Isla. La distribución total de la suma de frecuencias de las clases de talla para ambas épocas climáticas en los dos subsistemas (Fig. 34) muestra una clase modal de 66-70 mm. D. rhombeusse comporta como un visitante cíclico en la laguna, desovando probablemente en la plataforma de Campeche adyacente a la Laguna de Términos durante primavera-verano, migrando a la laguna para crecer, madurar y alimentarse regresando a la plataforma continental a reproducirse nuevamente. D. rhombeus se encuentra bien representado en las ictiocomunidades de la Sonda de Campeche, sin embargo, a la fecha no se han realizado estudios sobre su biología en esta área (Sánchez-Gil et al., 1981). D. auratus es una especie ocasional en la Laguna de Términos y sólo ha sido capturada en el litoral interno de la Isla del Carmen. La Tabla 3 muestra el valor del coeficiente bcalculado para esta especie. D. auratus se encuentra representado por individuos juveniles e inmaduros. Es importante hacer notar su presencia en áreas de pastos marnos donde existen salinidades mayores a 26 ppm. Esta especie ha sido reportada en otras lagunas costeras del Golfo de México con gran abundancia (Reséndez, 1970 y 1973; Chávez, 1972; CastroAguirre, 1978). Los valores del coeficienteb encontrados para Eugerres plumierireflejaron poca variación a lo largo del año (Tabla 3 y Figs. 45 y 48) tanto en el litoral interno de la Isla como en los sistemas fluviolagunares. En el litoral interno de la Isla durante ambas épocas climáticas el coeficiente bvarió de 3.2 a 3.5, mientras que en los sistemas fluvio-lagunares fue de 3.2. Las poblaciones de E. plumieriestán constituidas en su mayoría por individuos subadultos y adultos (Tabla 4, Figs. 45 a 48), por lo que las diferencias del coeficiente b podrían estar dadas por la proporción de adultos maduros encontrados en cada subsistema a lo largo del año. E. plumierise encontró representado durante la época de lluvias en los sistemas fluviolagunares en tallas entre 65 y 155 mm de longitud total (Tabla 4, Fig. 47) con una clase modal de 111-115 mm. Durante la época de secas las poblaciones encontrdas en el litoral interno de la Isla y en los sistemas fluvio-lagunares presentaron tallas entre 125 y 295 mm de longitud total (Tabla 4, Fig. 45), con una clase modal de 126-130 mm sólo para el litoral interno de la isla. La especie estuvo mejor representada en el litoral interno de la isla que en los sistemas fluviolagunares. La distribución total de la suma de frecuencias de las clases de talla para ambas épocas climáticas en los dos subsistemas (Fig. 48) muestra que la clase modal de 111-115 mm fue la más frecuente. E. plumieri es un visitante cíclico y penetra a la laguna para alimentarse y protegerse. Reséndez (1981b) la encuentra, al igual que este estudio, poco representada a lo largo del año, sin embargo, es la mojarra que alcanza las mayores tallas en la laguna, por lo mismo, constituye un recurso pesquero local. No ha sido reportada todavía para ambientes neríticos del Sur del Golfo de México, mientras que, en otras lagunas costeras del Golfo destaca por su gran abundancia durante el año donde también constituye un recurso comercial (Reséndez, 1970 y 1973), por lo que es importante el seguir incrementando los estudios sobre su biología y ecología en aguas costeras de México. MADUREZEl estudio de la madurez gonádica en los peces es un aspecto esencial para el entendimiento de sus cielos biológicos, por lo que, debe estudiarse desde un punto de vista temporal y espacial. Un análisis de este tipo permite entender la dinámica de una población de una especie determinada en un sistema lagunar-estuarino, ya que, dentro de esa dinámica se encuentran incorporadas estrategias que le permiten a las especies estar adaptadas a dicho sistema. Es por lo tanto, prioritario el conocer dentro de esas estrategias, áreas de desove, reclutamiento, crianza, maduración y ciclos migratorios. Así, mediante el conocimiento de estos aspectos se delimitan, proporción de sexos, tallas de primera madurez, distribución de fases sexuales, deriva larvaria entre otros. Todo lo anterior lleva a proponer áreas de protección y/o explotación en las lagunas costeras y estuarios de acuerdo al cielo biológico de las especies. Esto último conduce a un mejor uso, aprovechamiento y administración de los peces como recurso en la zona costera (Yáñez-Arancibia, 1982). En relación a estudios sobre la madurez gonádica y cielos reproductivos en especies de la Familia Gerreidae, existen muy pocas investigaciones al respecto. Yáñez-Arancibia (1978), estudia algunos aspectos sobre la madurez de Gerres cinereus(Walbaum), Eugerres lineatus (Humboldt y Valenciennes) yDiapterus peruvianus (Cuvier y Valenciennes) en lagunas costeras del Estado de Guerrero en el Pacífico de México. Bravo-Núñez y Yáñez-Arancibia (1979), en un trabajo sobre la ecología de las comunidades de peces de la Boca de Puerto Real en la Laguna de Términos, estudian algunos aspectos sobre la madurez deEucinostomus gula y Eucinostomus argenteus.Finalmente Aguirre León et al. (1982), estudian principalmente la biología de 8 especies de mojarras de la Laguna de Términos. Todas estas investigaciones han servido como marco de referencia a los estudios que aquí se presentan. La Tabla 5, resume comparativamente la distribución de fases sexuales de todas las especies por época climática y subsistema. De igual manera la Tabla 6 resume el análisis de proporción de sexos durante el año.  TABLA 5 DISTRIBUCIÓN DE FASES SEXUALES POR ESPECIE, LOCALIDAD Y ÉPOCA CLIMÁTICA Gerres cinereus es una especie muy poco frecuente en la Laguna de Términos, por lo que, no pudo ser estudiada en sus aspectos de madurez. Sin embargo, se cuenta con información sobre su biología en otras latitudes tropicales de México. Así, Yáñez-Arancibia (1978), entre otras consideraciones demanda la necesidad de efectuar estudios sobre la biología de esta especie en lagunas costeras mexicanas. En este último estudio se ha señalado que G. cinereus es una de las mojarras con grandes perspectivas como recurso pesquero en el Pacifico central de México. Dentro de las observaciones hechas sobre esta especie en las lagunas costeras del Estado de Guerrero se encontró que G. cinereusestuvo representada en tallas de 61 a 185 mm de longitud total, con predominancia de individuos en fases de desarrollo gonádico II y III. De acuerdo a estas observaciones, la especie alcanza la madurez gonádica después de los 145 mm de longitud total. Sus poblaciones inmaduras penetran en las lagunas para completar parte de su desarrollo gonádico, alimentarse y crecer. Los ejemplares capturados en Laguna de Términos oscilaron entre 89 y 159 mm de longitud total y de acuerdo a lo anterior es probable se encontrarán en fases de maduración I, II y III, sin embargo no fue posible determinarlo. Gerres cinereus es definitivamente rara en la Laguna de Términos, por lo que, su cielo biológico esta muy poco conocido en el Sur del Golfo de México. 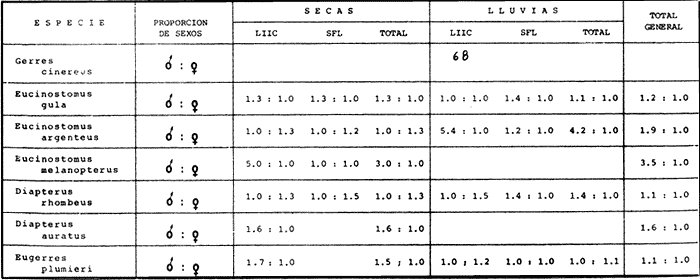 TABLA 6 PROPORCIÓN DE SEXOS POR ESPECIE, LOCALIDAD Y ÉPOCA CLIMÁTICA Del análisis realizado en Eucinostomus gula, se encontró que esta especie está representada en su mayoría por individuos en estadios tempranos de maduración gonádica (fases I y II) durante las épocas climáticas definidas para el área (Figs. 8 y 9). Sin embargo, comparando las dos épocas climáticas, se observa que en el período de secas (Fig. 8), se encuentra mejor representada esta especie. Asimismo se encontró que los machos presentan mayor frecuencia que las hembras, presentando la población una proporción total macho: hembra en esta época de 1.2: 1.0. A pesar de la amplia distribución y abundancia de esta especie en la laguna, el total de la población sólo alcanzó la fase de maduración IV, la cual se encontró para la época de secas en individuos de 140 mm de longitud total. No se encontraron individuos en fases III y IV en los sistemas fluvio-lagunares en cualquiera de las dos épocas climáticas lo cual sugiere que estas áreas sólo son utilizadas por individuos inmaduros que migran a ellos para crecer y alimentarse buscando aguas someras con vegetación de manglar y sumergida. Los registros de fases en maduración y maduras (fases III y IV) encontradas en el litoral interno de la Isla del Carmen sugiere por otro lado que esta especie necesita salinidades elevadas (mayores a 26 ppm) para alcanzar su madurez. Esto último se refuerza con el hecho de queE. gula se ha colectado al estado adulto en la plataforma adyacente a la Laguna de Términos en tallas generalmente mayores a 120 mm, y a pesar de que no se ha realizado análisis gonádico en estas poblaciones es factible que sean maduras. Las máximas tallas reportadas en esa área son de 146 mm de longitud total con gran abundancia hacia la región este de la Sonda de Campeche en profundidades de 13 m y en salinidades de 35.6 ppm (Sánchez-Gil et. al., 1981). Apoyando lo anterior, Hildebrand (1955) en un estudio de la fauna de acompañamiento del camarón rosado en el Golfo de México, encuentra ejemplares maduros de E. guladurante la epoca de secas. Asimismo Bravo-Núñez y Yáñez-Arancibia (1979) reportan para la Boca de Puerto Real en la Laguna de Términos, ejemplares en tallas entre 30 y 124 mm de longitud total, encontrando que están representadas las fases sexuales I, II, y III correspondiendo la fase III a ejemplares machos mayores a 120 mm, colectados en salinidades de 28 a 35 ppm durante la época de secas. Lo anterior ha sido también observado en las costas del norte del Golfo de México en los estados de Texas, Louisiana y Florida, donde E. gula ha sido colectada en la plataforma adyacente a las lagunas costeras de esos Estados, en tallas grandes y maduras, encontrando generalmente etapas juveniles en los sistemas costeros (Kilby, 1955; Springer y Bullis, 1956; Springer y Woodburn, 1960; Springer y Me Erlean, 1962). Estos últimos autores han señalado que E. gula migra hacia las aguas abiertas del Golfo de México desde las estuarinas para desovar. Este último comportamiento es similar al observado en el Sur del Golfo de México, ya que, la escasa presencia de tallas adultas de esta especie en la Laguna de Términos refuerza aún más un comportamiento de este tipo (Vargas Maldonado et al, 1981 y Aguirre León et al., 1982). Es importante destacar que, se ha reportado la presencia de larvas de gérridos de la especie Eucinostomus sp en gran parte de la cuenca central de la laguna, en las bocas de marea y áreas adyacentes a las mismas en la plataforma continental durante primavera y verano, siendo los registros en otoño e invierno escasos o nulos (Flores-Coto y Alvarez-Cadena, 1980; Méndez Velarde y Velarde Méndez, 1982). Ya se ha discutido en otros estudios que las larvas deEucinostomus sp, corresponden con mucha probabilidad aEucinostomus gula, especie que al estado juvenil es sumamente abundante en la laguna (Vargas Maldonado et al., 1981 y Aguirre León et al., 1982. De igual manera Ayala Duval (1980) en un estudio sobre el ictioplancton del Sur del Golfo de México, reporta la presencia de larvas de gérridos frente a la Laguna de Términos y en mayor abundancia hacia la región este de la Sonda de Campeche. Sin embargo, la identificación de las larvas sólo se hizo hasta familia. No obstante, la abundancia de larvas en esa región coincide con la abundancia de adultos de E. gulareportada por Sánchez Gil et. al. (1981) en la Sonda de Campeche para la misma región. Podría considerase que esas larvas correspondieran nuevamente a E. gula, dados los antecedentes presentados para la Laguna de Términos, sus bocas de marea y la plataforma continental adyacente. Es deseable que futuras investigaciones arrojen luz sobre este aspecto. Los dos estudios anteriormente referidos, fueron realizados aunque por separado en el mes de junio del mismo año en la época de verano y por lo mismo, existe correspondencia en los resultados encontrados. Es de esperarse que la distribución y abundancia de larvas se corresponda con la abundancia de adultos, si se trata de la época de desove, aunque dicha correspondencia no es obvia, ya que se esperaría mayor abundancia de huevos y no de larvas. Con base en estos resultados y de acuerdo a la presencia estacional de larvas de la especieEucínostomus sp en la Laguna de Términos, así como en la plataforma adyacente a esta, E. gulano desova dentro de la laguna, efectuando su reproducción cerca de la línea de costa y con mucha seguridad durante la época de secas que es cuando se registra la mayor abundancia de larvas tanto en la plataforma como dentro de la laguna. E. gula está actualmente siendo estudiada particularmente dada su importancia ecológica y parcialmente económica en el área. Estos estudios arrojarán mayor información sobre su biología y ecología. Eucinostomus argenteusestá representado principalmente por individuos juveniles e inmaduros (fases I y II) durante las épocas climáticas para el área (Figs. 20 y 21). Sin embargo, comparando las dos épocas climáticas se observa que en el período de secas (Fig. 20), se encuentra mejor representada esta especie. Asimismo se observa que las hembras presentan mayor frecuencia que los machos, presentando la población una proporción macho: hembra de LO: 1.3. La población total de esta especie sólo alcanzó la fase de maduración III durante la época de secas en individuos de 130 mm de longitud total. Al igual que en E. gula, las fases de maduración III en ejemplares colectados en el litoral interno de la Isla del Carmen, hace suponer que esta especie necesita similares condiciones ambientales para alcanzar su madurez sexual. Sánchez-Gil et al. (1981), han encontrado aE. argenteus en la plataforma de Campeche en tallas de 42 a 153 mm de longitud total, sin embargo, no se ha realizado análisis gonádico de estas poblaciones; no obstante, es factible pensar que los individuos mayores a 120 mm sean maduros. La mayor abundancia de tallas grandes se registró hacia la región oeste de la Sonda de Campeche en profundidades de 18 m y salinidades de 35.5 ppm. Springer y Bullis (1956) destacan la presencia de E. argenteus en el norte del Golfo de México en gran abundancia en profundidades de 18 a 65 m. Hildebrand (1954) considera rara la presencia de E. argenteus en las capturas de camaron café en las costas de Texas, sin embargo, la encuentra mejor representada en la Sonda de Campeche (Hildebrand, 1955). Las anteriores observaciones hacen suponer que esta especie se encuentra madura en aguas abiertas asociada a salinidades altas. No obstante, Springer y Woodburn (1960), encuentran en el Río Santa Lucía, Florida, 2 ejemplares hembras de 51.5 y 53.3 mm en estado avanzado de desarrollo gonádico, sin embargo, no mencionan la fase sexual de estos ejemplares. Esto último no es un comportamiento extraño, ya que E. argenteustambién ha sido colectado en la Laguna de Términos en fases de maduración III. Finalmente Bravo Núñez y Yañez-Arancibia (1979), han reportado la presencia de E. argenteus en la Boca de Puerto Real en la Laguna de Términos en tallas de 55 a 88 mm de longitud total en fases I y II y también de sexo indeterminado. No se cuenta para esta especie con registros de ictioplancton en el área, por lo que es factible pensar que E. argenteus realiza la mayor parte de su cielo biológico en la plataforma continental adyacente a la Laguna de Términos. Eucinostomus melanopterus está Pobremente representada en la Laguna de Términos, por lo que, los estudios sobre la madurez de esta especie en el área son muy incompletos. Esta especie sólo está representada por individuos en fases I y II. Es importante resaltar que, la mayoría de los ejemplares analizados fueron machos reflejando ésto en el análisis de proporción de sexos cuya relación fue de 3.5: 1.0. Igualmente, esta especie está escasamente representada en la Sonda de Campeche en tallas pequeñas (Yáñez-Arancibia et al., 1982b). Al igual que otras mojarras, E. melanopterus debe de realizar su reproducción en aguas costeras de la plataforma continental adyacente a la laguna. Es necesario realizar más estudios sobre la biología de esta especie en el Sur del Golfo de México. Aún cuando se encuentra en otras lagunas costeras del Golfo en mayor abundancia tampoco existen estudios al respecto. En relación a estudios sobre la madurez y reproducción deDiapterus rhombeus en el Sur del Golfo de México, se cuenta con poca información al respecto. En este trabajo, D. rhombeus esta representado en su mayoría por individuos en fases I y II durante la época de secas y sólo por la fase I durante la de lluvias. La fase III fue registrada en ambas épocas climáticas (Figs. 35 y 36), sin embargo, está representada por muy pocos individuos en ambos subsistemas. Las fases I y II se registraron en individuos menores a los 110 mm, mientras que la fase III se alcanza después de los 120 mm de longitud total, por lo que la especie alcanza la madurez después de esta longitud. La proporción de sexos macho: hembra, varía de una época climática a otra, siendo para la época de secas de LO: 1.3, y para la de lluvias de 1.4: 1.0. A pesar de su amplia distribución en la laguna, el total de la población sólo alcanzó la fase de maduración III.D. rhombeus, como otras mojarras utiliza la laguna como área de crianza y maduración. Se ha sugerido que esta especie migra a la zona nerítica del Golfo de México a realizar su desove al comienzo de la época de lluvias (Aguirre León et al., 1982), sin embargo, son necesarios más estudios al respecto. Sánchez-Gil et al., 1981, ha reportado aD. rhombeus en la Sonda de Campeche en tallas mayores a 114 m de longitud total. Estudios posteriores en la misma área la han reportado en tallas hasta de 135 mm (Yáñez-Arancibia et al., 1982b). Cabe suponer que, las tallas reportadas en la Sonda de Campeche representan individuos maduros. Sin embargo, la distribución y abundancia de D. rhombeus a lo largo del año es muy irregular. No cuenta con registros de ictioplancton en la laguna, ni en la Sonda de Campeche. Al igual que en otras mojarras son necesarios estudios sobre el ictioplancton de estas especies en cuerpos costeros. Es importante mencionar que Diapterus peruvianus (Cuvier y Valenciennes), especie homóloga a Diapterus rhombeus, pero del Pacífico central de México, ha sido objeto de estudios parciales sobre su biología y dinámica de sus poblaciones. Este estudio constituye un marco de referencia para el entendimiento de los cielos reproductivos de especies homólogas en ecosistemas costeros del Golfo de México (YáñezArancibia, 1978). Diapterus rhombeus ha sido también estudiado en aspectos parciales de su biologia en aguas costeras del litoral suroeste de Puerto Rico, en el Océano Atlántico (Austin, 1971). Se destaca que esta especie realiza la mayor parte de su cielo biológico en aguas someras fuera de la costa. Alcanza su madurez sexual a los 125 mm de longitud total y presenta una época de reproducción que se extiende por 8 meses a lo largo del año principalmente durante la época de secas y lluvias. Algunas de las observaciones realizadas en el presente trabajo son similares a las encontradas por Austin (1971) en aguas de Puerto Rico. Diapterus auratus es una especie afin a Diapterus rhombeus, sin embargo, está pobremente representada en la Laguna de Términos, por lo que, los estudios sobre su madurez gonádica en esta área son muy incompletos. El análisis gonádico de esta especie sólo reveló individuos en fase I en tallas no mayores a 70 mm de longitud total. Mientras que, el análisis de proporción de sexos macho: hembra, fue de 1.6: 1.0. Ha sido reportada con mayor abundancia en la Sonda de Campeche y en tallas mayores a 130 mm de longitud total, siendo probable que se trate de individuos maduros (Yáñez-Arancibia et al., 1982b). Por la importancia comercial que reviste en otras lagunas costeras del Golfo de México es importante realizar estudios sobre su reproducción con vistas a un mejor aprovechamiento de esta especie como recurso actualmente explotado. En la Laguna de Términos esta especie no tiene importancia económica. Eugerres plumieri es la mojarra de mayor talla presente en la Laguna de Términos, reviste por lo tanto, importancia económica local en el área. Está representada por fases inmaduras I y II en ambos subsistemas hasta los 184 mm de longitud total. Por fases en maduración (fase III) y maduras (fases IV), postreproductivas y en descanso (fases VI y VII), sin embargo, su distribución no es regular a lo largo del año en ambos subsistemas (Figs. 49 y 50). De acuerdo con estos resultados, E. plumieri madura después de los 180 mm de longitud total. La población de esta especie presenta una proporción de sexos macho: hembra de 1. 1: 1. 0. A pesar de que esta especie está representada por fases maduras y postreproductivas, son muy pocos los individuos que representan estas fases a lo largo del año, por lo que, la presencia de individuos maduros en la laguna se debe principalmente a necesidades tróficas más que a reproductivas. Lo mismo puede decirse de los individuos jóvenes ya que los ejemplares analizados presentaron el estómago lleno. Además esta especie se le encuentra con más frecuencia en zonas protegidas de manglar en fondos limosos con abundancia de detritus y aguas turbias. No cuenta con registros de ictioplancton en la laguna y todavía no ha sido reportada para la Sonda de Campeche, por lo que, aspectos sobre el cielo biológico de esta especie en el Sur del Golfo de México son poco conocidos. Otros estudios en lagunas costeras del Golfo de México reportan a E. plumieri en su mayoria con tallas comerciales (Darnell, 1962; Reséndez, 1970, 1973, 1979 y 1981b; Chávez, 1972 y Castro-Aguirre, 1978). E. plumieri como especie típica de lagunas costeras mexicanas del Golfo de México, carece casi por completo de estudios sobre su reproducción y ecología, requiere por lo tanto, de estudios urgentes sobre su cielo biológico. Especies homólogas a E. plumieri en el Pacífico central de México,i.e., Eugerres lineatus (Humboldt) y Eugerres axílaris han sido estudiadas parcialmente en sus aspectos biológicos y dinámica de poblaciones en lagunas costeras del Estado de Guerrero (YáñezArancibia, 1978). De acuerdo a este estudio, estas especies penetran a estas lagunas costeras con finalidades tróficas en tallas adultas principalmente, siendo su abundancia considerable a lo largo del año. ALIMENTACIÓNEl estudio del contenido estomacal de los peces constituye un aspecto muy importante para la comprensión de los hábitos alimenticios, del hábitat donde ocurren las especies, y también de la ecología y relaciones fisiológicas entre especies afines (Yasuda, 1960). Sin embargo, aún más importante en el análisis cuantitativo de la alimentación y hábitos alimenticios de los peces, son las relaciones que se establecen entre espectro trofico, trama trófica y flujo de energía, ya que finalmente estas interrelaciones tienden a definir el papel ecológico de las especies en un ecosistema determinado (Yáñez-Arancibia y Nugent, 1977). Estas interrelaciones son de mayor -complejidad en las lagunas costeras y estuarios por su alta diversidad faunística y de habítat (Day y Yáñez-Arancibia, 1982) y por la dinámica de los espectros tróficos (Sheridan y Livingston, 1979). La ubicación de los peces dentro de la trama trófica en los estuarios y lagunas costeras mediante el conocimiento de la alimentación y hábitos alimenticios determina que por la biología de las especies y sus relaciones ecológicas, transformen energía a partir de fuentes primarias, la conduzcan activamente a través de la trama trófica, que por migraciones intercambien energía con ecosistemas vecinos a través de la importación y exportación de ella, ademas, constituyan una forma de almacenamiento de tal energía dentro del ecosistema, así como también, funcionen como agentes de regulación energética (Yáñez-Arancibia y Nugent, 1977). Todo lo anterior está enfocado hacia una interpretación y comprensión de la ecología de los estuarios y lagunas costeras para efectuar recomendaciones para la administración óptima de sus recursos pesqueros (Yáñez-Arancibia, 1982). Dentro de este último enfoque las mojarras de la familia Gerreidae, están siendo objeto de diversos estudios sobre su alimentación y hábitos alimenticios en lagunas costeras y estuarios del Sur del Golfo de México por ser especies que actualmente constituyen un recurso pesquero en explotación. La baja frecuencia y abundancia de Gerres cinereus en la Laguna de Términos no ha permitido realizar estudios profundos sobre su alimentación y hábitos alimenticios desde un punto de vista cuantitativo. Se ha señalado que G. cinereus es una especie de suma importancia en estuarios y lagunas costeras del norte del Golfo de México donde ha recibido mayor atención al respecto (Tabbet al. 1962 y Hoese y Jones, 1963). El estudio cualitativo del contenido estomacal de esta especie indica que puede alimentarse de diversos grupos bentónicos, entre ellos: pelecípodos, gasterópodos, poliquetos, además ostrácodos y materia organica de origen diverso, así como también fragmentos de vegetales, siendo los grupos tróficos más, abundantes la materia organica y los pelecípodos. Los individuos analizados variarion entre 89 y 159 mm de longitud total y por tratarse de ejemplares subadultos y adultos su alimentación está bien definida. Esto último puede reforzarse por lo descrito por Randall (1967), el cual encuentra en el Mar Caribe, que los grupos tróficos principales que incorpora a su dieta G. cinereus son pequeños decápodos en un 27 % y pelecípodos con un 23 % siguiendo en importancia los gasterópodos, poliquetos y sipuncúlidos, encontrándose un patrón similar al que presenta la especie en Laguna de Términos. Los ejemplares analizados variaron entre 95 a 280 mm de longitud patrón. Asimismo se observó que esta especie puede alimentarse sobre substratos duros y en claros de arena entre arrecifes de coral. Beebe y Tee-Van (1928) y Longley e Hildebrand (1941) han encontrado que G. cinereus se alimenta principalmente de poliquetos, moluscos y crustáceos, además de partes de pequeños peces. Especies afines del género Gerres han sido estudiadas en otras latitudes por la importancia comercial que revisten. Autores como Francis (1878), Job (1940), Chacko (1949), Bapat y Bal (1952), Basheeruddin y Nagappan Nayar (1961), Jhingran et al. (1963), Jhingram y Natayaran (1966) y Prabhakara Rao (1968), han discutido ampliamente diversos aspectos de la alimentación y hábitos alimenticios de Gerres oyena (Forskal),Gerres oblongus Cuvier, Gerres filamentosus Cuvier, Gerres setifer (Hamilton) y Gerres abbreviatus Cuvier, en el Indo Pacífico. En conjunto, estos estudios han coincidido en señalar lo variado de los grupos tróficos en la alimentación de las diferentes especies. Estas son comedoras de fondo con hábitos alimenticios similares, encontrando en su amplio espectro trófico, pelecípodos, detritus, anfipodos, poliquetos, gasterópodos, copépodos, pequeños decápodos y vegetales entre otros. Se ha señalado también en estos estudios que los grupos tróficos cambian segUn la talla de los ejemplares, encontrando que. los ejemplares juveniles se alimentan preferentemente de copépodos y diatomeas, mientras que al estado adulto estos grupos tróficos son substituidos por otros diferentes. Recientemente Blaber (1982) ha estudiado la oesteología de los aparatos bucales de 3 especies del género Gerres de las aguas costeras del sureste de Africa en relación a los hábitos alimenticios de estas especies, encontrando que, la morfología de sus aparatos bucales está íntimamente relacionada al tipo de alimento que ingiere. Asimismo Cyrus y Blaber (1983) han estudiado la ecología trofodinámica del géneroGerres en estuaríos de Natal, India. Otros estudios sobre la alimentación de Gerres cinereus han sido hechos por Yáñez-Arancíbia (1978) en lagunas costeras del Pacífico central de México. Dicho estudio reveló que esta especie se alimenta de yegetales, peces, moluscos, crustáceos, insectos, anélidos, ostrácodos, copépodos, foraminíferos, anfipodos, briozoos, materia orgánica y sedimentos inorgánicos. En este estudio también se destaca la variación que sufre el espectro trófico al estado juvenil y adulto. Los ejemplares examinados variaron entre 61 y 185 mm de longitud total y son considerados como consumidores de primer y/o segundo orden. En el presente estudio, Gerres cinereus se considera como un consumidor.de primer orden. Futuras investigaciones sobre la biología de la especie en el Golfo de México reforzarían los ya existentes y arrojarían mayor información al respecto. La alimentación deEucinostomus gula fue analizada de acuerdo a las épocas climáticas definidas para el área tanto en el litoral interno de la Isla del Carmen así como en los sistemas fluviolagunares (Figs. 11 a 14). No se encontraron diferencias apreciables en el espectro trófico de E. gula de una época climática a otra ni de un subsistema al otro. De acuerdo al análisis trófico combinado realizado en esta especie y al índice de importancia relativa IIR se encontró que los poliquetos son el alimento principal a lo largo del año. Aparece también como grupo trófico abundante numéricamente durante la época de secas en el litoral interno de la Isla (Fig. 11). En menor proporción incorpora materia orgánica y una gran diversidad de grupos bénticos de pequeño tamaño. Puede notarse que el espectro trófico de E. gula estacionalmente es muy homogéneo en cuanto a grupos preferenciales y al mismo tiempo variado en cuanto a grupos tróficos secundarios y circunstanciales. Los ejemplares analizados-de esta especie variaron de 35 a 112 mm de longitud total. De acuerdo a este rango de tallas sólo individuos menores a 40 mm, ingieren preferencialmente copépodos y esta preferencia fue detectada del análisis numérico realizado en ejemplares de esas tallas. Sin embargo, la- mayoría de los ejemplares analizados corresponden a individuos de mayor talla. Es importante hacer notar que E. gula presenta variaciones en su dieta de acuerdo a la talla, no obstante, las tallas analizadas en este estudio no permiten cuantificar completamente esas variaciones. En función de estos resultados se considera a E. gula como un consumidor de primer orden en la Laguna de Términos. Bravo Núñez y YáñezArancibla (1979) han encontrado que esta especie en la Boca de Puerto Real en la Laguna de Términos se alimenta de una gran variedad de grupos tróficos entre los que destacan como importantes: copépodos, pequeños crustáceos decápodos, poliquetos, oligoquetos, isópodos y anfípodos. E. gula ha sido objeto de estudios sobre su alimentación en lagunas costeras del Norte del Golfo de México donde es importante por su abundancia a lo largo del año. Reid (1954) analizó estómagos de ejemplares cercanos a Cedar Key, Florida, encontrando en ellos, copépodos con un 65.8 % y políquetos con 51.2 % ; camarones y misidáceos en un 7.3 %. No encontró copépodos en individuos mayores a 45 mm. Springer y Woodburn (1960) en la Bahía de Tampa, Florida, encontraron que estómagos deE. gula contenían en su mayoría poliquetos y en menor cantidad copepodos; otros más con algas filamentosas, ostrácodos, gamaridos, holoturoideos, pelecípodos y partes de crustáceos decápodos. Odum y Heald (1972) han estudiado la alimentación de ejemplares juveniles de E. gula (19 a 70 mm de longitud total) en el suroeste de Florida cerca de Cabo Sable. Analizaron el espectro trófico de esta especie durante la época de secas y de lluvias de esa área encontrando que durante la época de secas predominan en el espectro de E. gula, copépodos harpacticoides, anfipodos y en menor proporción, pequeños moluscos, poliquetos, misidáceos y detritus. En la época lluviosa, los copépodos, moluscos y poliquetos nereidos desaparecen de la dieta de esta especie para ser reemplazados en gran porcentaje por larvas de chironómidos, manteniéndose los anfipodos durante todo el año. Comportamientos similares en la alimentación de E. gula han sido encontrados por Linton (1905), Smith (1907), Reid (1954), Springer y Woodburn (1960), Randall (1967), Odum (1971), Carr y Adams (1973), Adams (1976) y Brook (1977). La mayoría de estos autores destacan el cambio que sufre la alimentación de E. gula conforme crece. Además coinciden en que individuos hasta los 40 mm, ingieren preferentemente copépodos siendo substituidos gradualmente por poliquetos como grupo preferencial en tallas mayores a 50 mm. Del análisis anterior se desprende que esta especie como otras mojarras se alimenta entre y sobre las hojas de pastos marinos y en la interfase aguasedimento, predando sobre organismos del micro y meiobentos. El espectro trófico deEucinostomus argenteus fue analizado también a lo largo de las épocas climáticas en el año y en función de los dos subsistemas definidos en Laguna de Términos (Figs. 23 a 25). De acuerdo a este análisis, se observan variaciones en la alimentación de esta especie de una época climática a otra y de un subsistema a otro, incluso durante una misma época climática. Mientras que en la época de secas, el alimento principal en el litoral interno de la Isla del Carmen son los poliquetos, y en menor proporción la materia orgánica (MOND), en los sistemas fluvio-lagunares el alimento principal es la materia orgánica (MOND) y en menor medida los tanaidáceos. Durante la época de lluvias en el litoral interno de la isla el alimento principal lo constituyen los poliquetos y en menor medida los tanaidáceos, mientras que en los sistemas fluvio-lagunares, el grupo trófico principal son los tanaidáceos y en menor medida los poliquetos y la materia orgánica. Al mismo tiempo, durante ambas épocas climáticas en el litoral interno de la Isla del Carmen, los poliquetos aparecen como el grupo preferencial, mientras que en los sistemas fluvio-lagunares la materia orgánica (MOND) que aparece como principal en secas es substituida por una gran abundancia de tanaidáceos en la época lluviosa. Los ejemplares analizados deE. argenteus variaron de 27 a 122 mm de longitud total, sin embargo, la.mayoría de ellos oscilaba entre 30 y 90 mm. Los individuos menores a 40 mm ingieren preferentemente copépodos, ostrácodos y otros grupos del microbentos. Individuos mayores a 40 mm cambian gradualmente su dieta a tanaidáceos, materia organica y poliquetos. Al parecerE. argenteus tiene un espectro más heterogéneo en cuanto a grupos principales en comparación con E. guladurante el año, no obstante es similar en cuanto a consumo de grupos secundarios y circunstanciales. Esto pudiera deberse a que E. argenteus presenta competencia por alimento con E. gula, ya que estas dos especies son las más abundantes del género Eucinostomus en la Laguna de Términos, además de que pueden encontrarse juntos en el mismo hábitat (Aguirre Leónet al., 1982). De acuerdo al análisis trófico realizado en E. argenteus, se considera a esta especie como consumidor de primer orden en este estudio. Bravo-Núñez y Yáñez-Arancibia (1979) han analizado estómagos de E. argenteus en la Boca de Puerto Real en la Laguna de Términos encontrando cefalocordados (Amphioxus), protocordados, poliquetos, nemátodos, copépodos, fragmentos vegetales y materia orgánica. Los ejemplares analizados variaron de 55 a 88 mm de longitud total. Springer y Woodburn (1960), encuentran poliquetos como alimento principal en E. argenteus en un estudio de la Bahía de Tampa, Florida, además encontraron también, copépodos, ostrácodos, anfípodos y pequeños pelecípodos. Estos autores destacan también que no existen casi copepodos en la dieta de esta especie después de los 45 mm. Odum y Heald (1972) analizan la alimentación de E. argenteus en individuos de 19 a 63 mm de North River, Florida, encontrando durante la época de secas, principalmente anfípodos y copépodos harpacticoides, siendo substituidos estos últimos por larvas de chironómidos en la época de lluvias, mientras que los anfípodos permanecen en su dieta. Otros grupos como pequeños moluscos, misidáceos, ostrácodos y detritus son consumidos en menor cantidad. Estos autores destacan que E. argenteus y E. gula no presentan ninguna separación ecológica entre ellos ya que sus espectros tróficos son casi idénticos a lo largo del año. Similar comportamiento ha sido observado por Brook (1977) en Card Sound, Florida. Springer y Woodburn (1960) sí encuentran una separación ecológica en estas dos especies de acuerdo a sus espectros tróficos en la Bahía de Tampa, Florida. Estas observaciones son similares a las encontradas en este estudio. Finalmente Randall (1967) ha encontrado anfipodos, poliquetos y pequeños cangrejos en la alimentación de E. argenteus en el Mar Caribe. E. argenteus como otras moj arras presenta un espectro trófico muy amplio y variado dependiendo de la disponibilidad de alimento, época del año, localidad y relaciones ecológicas con especies afines. Eucinostomus melanopterus carece por completo de estudios cuantitativos sobre sus relaciones tróficas en lagunas costeras y estuarios del Golfo de México, a pesar de que está bien representada en lagunas costeras de Veracruz y Tamaulipas (Darnell, 1962; Reséndez, 1970 y 1973; Chávez et al., 1976; Castro-Aguirre, 1978). En Laguna de Términos es poco frecuente, de lo cual, sólo ha sido posible realizar estudios cualitativos. Así, el análisis indicó que puede alimentarse de tanaidáceos, poliquetos, copépodos, ostrácodos, cumáceos, nemátodos, foraminíferos, materia orgánica, restos de pequeños decápodos y restos vegetales. De estos grupos tróficos la materia orgánica (MOND), los copépodos y los restos vegetales fueron los más abundantes. Las tallas de los ejemplares analizados variaron entre 50 y 120 mm de longitud total. E. melanopterus se considera como un consumidor de primer orden de acuerdo al análisis efectuado. Pocas observaciones sobre la alimentación de esta especie se han hecho en otras latitudes del Atlántico occidental. Perry y Perry (1974) han analizado estómagos de E. melanopterus en la Costa Atlántica de Costa Rica, encontrando principalmente poliquetos, moluscos y también en menor proporción peces pequeños. Los estudios sobre la alimentación y hábitos alimenticios de Diapterus rhombeus son en general también muy escasos en latitudes tropicales de América. En la Laguna de Términos D. rhombeus es la segunda especie más abundante y está bien representada durante la mayor parte del año, por lo que, el análisis de sus relaciones tróficas se realizó en función de la estacionalidad de la propia especie.D. rhombeus es importante también en otras lagunas costeras del Golfo de México, sin embargo, no ha recibido atención en estudios sobre su alimentación. Los resultados del análisis trófico combinado para esta especie demostraron que su espectro de alimentación es muy amplio como el de otras mojarras afines en la Laguna de Términos, presentándose ligeras variaciones en función de la epoca climática en el año y la localidad. En cuanto a grupos tróficos principales se refiere, el espectro de D. rhombeus es homogeneo a lo largo del año. Durante la época de secas, tanto en el litoral interno de la Isla del Carmen como en los sistemas fluvio lagunares, el alimento principal lo constituye la materia orgánica (MOND). Acompañando a este grupo trófico con similar importancia se encuentran poliquetos en los sistemas fluvio-lagunares. En la época lluviosa vuelve a aparecer como grupo importante, la materia orgánica en ambos subsistemas. Cabe destacar que a lo largo del año los copepodos constituyen un complemento importante en la dieta de D. rhombeus. Aunque no tienen alta significancia volumétrica, numéricamente son ímportantes, ya que como grupo trófico siguen apareciendo en tallas mayores a 90 mm de longitud total. Ests últimas observaciones difieren de las encontradas para especies del género Eucinostomus, en donde los copépodos sólo aparecieron en ejemplares no mayores a 35 mm, cambiando en gran medida su dieta a organismos del meiobentos en individuos mayores a 45 mm. Los ejemplares analizados variaron entre 50 y 120 mm y aunque no se analizaron ejemplares pequeños de D. rhombeus, es muy probable que como otras mojarras, su dieta esté constituida casi exclusivamente de copépodos, y el que sigan apareciendo en tallas aún mayores como se encontró en este caso indica que la especie seguiría comportándose en mayor medida que otras mojarras como zooplanctófago. Por otro lado, la presencia mayoritaria de materia orgánica a lo largo del año en esta especie podría ser producto de la digestión de otros grupos tróficos, o la ingestión directa del medio. Al parecer es más probable la primera opción. De acuerdo a estos resultadosD. rhombeus se considera consumidor de primer orden. En relación a otros estudios sobre la alimentación del género Diapterus,Yáñez-Arancibia (1978) ha analizado las relaciones tróficas de Diapterus peruvianus en lagunas costeras del Pacífico Central de México. Esta última especie es homóloga a D. rhombeus del Golfo de México. De acuerdo al estudio cualitativo de estómagos de D. peruvianus se encuentra que la especie se comporta con hábitos preferentemente carnívoros predando sobre pequeños peces, anélidos, crustáceos y moluscos, y eventualmente vegetales y detritus. Sin embargo, en algunas de estas lagunas se encontró que el detritus y la materia orgánica constituían una parte importante en la dieta de D. peruvianus con un 55 % en volumen. Además se observó que durante períodos hiposalinos en estas lagunas, la dieta puede componerse principalmente de copépodos con un 60 % en volumen y 80 % de frecuencia. Estos resultados son similares al patrón encontrado en los hábitos alimenticios de D. rhombeus ya que los grupos tróficos que lo caracterizan tienen un comportamiento similar en ambas especies. Las tallas de los ejemplares de D. peruvianusexaminados variaron entre 28 y 128 mm de longitud total. D. rhombeus requiere necesariamente de estudios sobre su ecología trofodinámica en lagunas costeras del Golfo de México. Diapterus auratus es otra mojarra que en la Laguna de Términos tiene poca representatividad. Su análisis trófico sólo se realizó durante la época de secas en el litoral interno de la Isla del Carmen. Su alimento principal durante esa época fue la materia orgánica (MOND). Destacan también numéricamente los copépodos en la dieta de esta especie aún cuando no tuvieron una alta significancia volumétrica. Otros grupos tróficos que puede ingerir esta especie en menor proporción son: restos vegetales, ostrácodos, nemátodos, foraminíferos, quinorrincos, huevos de invertebrados y restos de peces. Esta especie es considerada como consumidor de primer orden. Los ejemplares examinados variaron entre 30 y 70 mm de longitud total. Los estudios sobre la alimentación de Diapterus auratus en el G olfo de México son m uy escasos o no se tiene referencia de ellos, a pesar de que es una de las mojarras que reviste gran importancia comercial en lagunas costeras del Golfo de México, principalmente en Veracruz y Tamaulipas (Hildebrand, 1958; Darnell, 1962; Resendez, 1970, 1973 y 1979; Chávez, 1972; Chávezet al., 1976; C astro-Aguirre, 1978). D. auratus no cuenta tampoco con estudios sobre su alimentación en otras latitudes del Atlántico de América. Sin embargo, las observaciones hechas por Yáñez-Arancibia (1978) sobre D. peruvianus en lagunas costeras del Pacífico Central de México sirven como marco de referencia en las investigaciones sobre la alimentación y hábitos alimenticios del géneroDiapterus en México. Son escasos los estudios sobre la alimentación y hábitos alimenticios deEugerres plumieri en el Golfo de México. En la Laguna de Términos se analizó la alimentación de esta especie en el litoral interno de la Isla del Carmen durante la época de secas y en los sistemas fluvio-lagunares durante la de lluvias. De acuerdo al análisis trófico combinado realizado para E. plumierien la época de secas, se encontró que su alimento principal es la materia organica (MOND). Numéricamente, destacan también durante esta epoca los ostrácodos y en menor medida los copepodos. En la época de lluvias permanece como grupo trófico principal la materia orgánica y en menor medida pequeños pelecípodos. Numéricamente los copépodos tienen alta significancia en esta época. E. plumieri ingiere diversos grupos del meiobentos como alimento secundario o circunstancial. Los ejemplares analizados de esta especie variaron entre 69 y 295 mm de longitud total. Se observó que individuos menores a 80 mm ingerían preferentemente copépodos con alta significancia numérica. Después de esta talla y hasta los 200 mm aproximadamente su dieta varía consumiendo principalmente pequeños crustáceos, moluscos y ostrácodos. Individuos mayores presentan un comportamiento preferentemente carnívoro pues pueden predar pequeños peces, aunque no deja de ser importante la materia orgánica. Con base en estos resultados, E. plumieri es considerado como un consumidor de primer orden. Algunas observaciones en relación a la alimentación deE. plumieri han sido hechas por Springer y Woodburn (1960) en la Bahía de Tampa, Florida. El análisis estomacal reveló que se alimenta principalmente de poliquetos. Los ejemplares examinados variaron entre los 176 mm de longitud total. Otros individuos del Río Santa Lucía de 71 a 78 mm contenían en sus estómagos ostrácodos y copépodos. Harrington y Harrington (1961), han analizado también la alimentación de juveniles (5 a 35 mm) de E. plumieri encontrando copépodos en un 86 % en volumen e insectos con un 9 %. Odum (1971) y Odum y Heald (1972) han señalado que no existen diferencis marcadas en la alimentación de esta especie entre juveniles y adultos. Analizaron ejemplares de 35 a 172 mm. En los estómagos examinados, determinaron volumétricamente 44 % de misidáceos, 36 % de anfípodos, 6 % de larvas de chironomidos, 4 % de copépodos harpacticoides, 4 % de detritus vegetal, 3 % de ostrácodos y 3 % de pequeños bivalvos. Sin embargo, estos resultados difieren de los encontrados para E. plumieri en la Laguna de Términos donde su alimentación varía notablemente de etapas juveniles a las adultas. En lagunas costeras del Pacífico Central de México se presentan dos especies homólogas a Eugerres plumieri que sonEugerres lineatus y Eugerres axillaris, sin embargo, la última especie es poco frecuente en las lagunas de esa área del Pacífico y E. lineatus es la que por su abundancia reviste importancia comercial y mayor interés en el estudio de sus relaciones tróficas (Yáñez-Arancibia, 1978). Se precisa en este último estudio que E. lineatus es una especie consumidora de primer orden predominantemente omnivora. Su alimento está constituido por vegetales, pequeños moluscos peces y crustáceos, poliquetos, insectos, detritus, ostrácodos, copepodos y foraminíferos y sedimentos inorgánicos. Ecológicamente reviste importancia el pastoreo que realiza sobre algunos vegetales específicos, comportamiento que ya ha sido estudiado en otros peces por Randall (1965) y Odum (1970). Las tallas de los ejemplares examinados variaron entre 134 y 187 mm de longitud total. Mojarras del género Eugerres en México representan un recurso comercial de buena calidad, por lo que, se necesita mayor atención en estudios sobre su biología en lagunas costeras mexicanas. Conclusiones1. La familia Gerreidae está bien representada en la Laguna de Términos por su diversidad, distribución y abundancia. Siete especies juegan un papel ecológico muy importante en la estructura y función de las comunidades ictiofaunísticas del área. 2. Las poblaciones de todas las especies están constituidas en su mayoría por individuos juveniles, a excepción de Eugerres plumieri, que está además representada por individuos adultos. 3. En general, se encontró para las especies analizadas, que las variaciones que sufre el coeficiente b de alometría de la relación pesolongitud se deben a las diferencias en los rangos de las clases de talla en función de la localidad y la época del año. 4. Todas las especies están representadas por fases tempranas de maduración gonádica con poca representatividad de individuos maduros a lo largo del año. 5. Eugerres plumieri es la especie que estuvo mejor representada por fases maduras y postreproductivas en la laguna, sin embargo, no es abundante durante todo el año. 6. En general, para todas las especies existe una proporción ligeramente mayor de machos que de hembras. 7. La mayoría de las especies alcanzan la madurez sexual después de los 120 mm de longitud total, a excepción de Eugerres plumíeri que la alcanza después de los 180 mm. 8. Eucinostomus gula es la única especie que tiene registros de ictioplancton en la laguna durante primavera y verano. 9. Las fases adultas registradas en Eucinostomus gula y Eucinostomus argenteusfueron encontradas en áreas con persistente influencia marina, mientras que, para Diapterus rhombeus y Eugerres plumieri,fueron registradas indistintamente en la laguna. Las demás especies no presentan un patrón regular de distribución de fases sexuales. 10. Por todo lo anterior, se considera que la Laguna. de Términos es utilizada por las mojarras de la f amilia Gerreidae como un área de protección, crianza, maduracion y alimentación. 11. El espectro trófico de todas las especies es muy amplio y diverso. Son consumidores de primer orden. Presentan variaciones particulares en su alimento principal., secundario y ocasional de acuerdo a la edad, la época climática, la localidad y la disponibilidad de alimento en la laguna. 12. Las mojarras presentan una alimentación zooplanctófaga en estadíos juveniles, representada principalmente por copépodos y ostrácodos. En individuos subadultos y adultos su alimentación es preferentemente omnívora, representada por poliquetos, pequeños decápodos, moluscos y detritus. 13.Eucinostomus gula presenta patrones estacionales de migración hacia la Sonda de Campeche que obedecen a estrategias reproductivas. 14. La importancia biológica y ecológica de Eucinostomus gula determina que esté siendo estudiada de manera particular en relación a la dinámica de sus poblaciones y cielo biológico. AgradecimientosAl Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México por el apoyo institucional en actividades de campo y laboratorio para la realización del presente estudio. Asimismo al Consejo Nacional de Ciencia y Tecnología (CONACYT) el apoyo económico a través del Proyecto Estudios Ecológicos de la Laguna de Términos, Campeche, México, con Referencia Especial a los Recursos Pesqueros y al Impacto Potencial del Hombre"PCMABEU - 005322. A Agustín Ayala-Castañares, Manuel Gallardo Cabello, Alfredo Laguarda-Figueras y Leonardo Lizárraga-Partida por la cuidadosa revisión del manuscrito y sus valiosos comentarios y sugerencias. A los compañeros del Laboratorio de Ictiología y Ecología Estuarina por la colaboración en las distintas campañas para colecta de peces y parámetros ambientales de la Laguna de Términos. Especial agradecimiento a Silvia Díaz Ruiz por su constante ayuda en la elaboración de tablas. gráficas y comentarios generales al trabajo. A Eduardo SáinzHernández por su ayuda en la elaboración de los programas de computación para el análisis estadístico de los datos. LITERATURAADAMS, S.M., Feeding ecology of eelgrass fish communities Trans. Amer. Fish. Soc., 1976. 514-519 105: AGUIRRE LEÓN A., A. YÁÑEZ-ARANCIBIA y AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Taxonomía, diversidad, distribución y abundancia de las mojarras de la Laguna de Términos, Campeche. (Pisces: Gerridae). 1982. 213-250 9 (1): ÁLVAREZ GUILLÉN, H., A. YÁÑEZ-ARANCIBIA y A. L. LARA-DOMÍNGUEZ, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de la Boca del Carmen, Laguna de Términos. El habitat y estructura de las comunidades de peces. 1985. 107-144. (1): 12 AMEZCUA-LINARES F. y A. YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El hábitat y estructura de las comunidades de peces. 1980. 70-118. (1): 7 AUSTIN, M. H., Bull., Some aspects of the biology of the rhomboid mojarra Diapterus rhombeus in Puerto Rico. Mar. Sci., 1971. 886-902. (4): 21 AYALA-DUVAL, E., Tesis Profesional. Contribución al conocimiento del ictioplancton de la región suroccidental del Golfo de México. Fac. de Ciencias, Univ. Nal. Autón. de México, 1980. 61 p. BAPAT, S. V. y D. V. BAL, The food of some young fishes from Bombay. Proc. Indian Acad. Sci., 1952. See. B. 78-92. 35 (2): BASHEERUDDIN, S. y K. NAGAPPAN NAYAR, A preliminary study of the juveniles fishes of the coastal waters off the Madras City. J. Fish., Indian 1961. 169-188. (1): 8 BAEZ HIDALGO, M. y E. GUEVARA CARRIO, Estudio morfométrico y merístico de Eugerres brasiliensis (Cuvier in Cuvier et Valenciennes, 1830) (Pisces: Gerreidae). Revista de Investigaciones Marinas, 1983. 63-90 (1): 4 BEEBE, W. y J. TEE-VAN, The fishes of Port-andPrince-with a summary of the known species of marine fishes of the island of Haiti and Santo Domingo. Zoologica, 1928. 1-279 10 (1): BEEBE, W. y J TEE-VAN, Field Book of the Shore Fishes of Bermuda and the West Indies. Inc., Dover Publications, New York, U.S.A., 1970. 327 p. BRAVO-NUÑEZ, E. y A. YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de la Boca de Puerto Real, Laguna de Términos. I. Descripción del área y análisis estructural de las comunidades de peces. 1979. 125-182 6 (1): BROOK, J. M., Trophic relationships in a seagrass community (Thalassia testudinum), in Card Sound, Florida. Fish diets in relation to macrobenthic and cryptic faunal abundance. Trans. Amer. Fish. Soc., 1977. 219-229 106 (3): CARR, W. E. S. y C. A. ADAMS, Food habits of juvenile marine fishes occupying seagrass beds in the estuarine zone near Crystal river, Florida. Trans. Amer. Fish. Soc., 1967. 511-540 102 (3): CARRANZA, J., Informe preliminar sobre la alimentación y hábitos alimenticios de las principales especies de peces de la zona de los Planes Pilotos Yavaros y Escuinapa. 3er. Informe del Contrato de Estudios No. EI-69-51, Univ. Nal. Autón. México, Inst. Biol. y SRH, 1969. 28p. CARVAJAL, J., Contribución al conocimiento de la biología de los robalos Centropomus undecimalis y C. poeyi en la Laguna de Términos, Bol. Inst. Oceanogr. Univ. Oriente, Campeche, México 1975. 51-70 14 (1): CASTRO-AGUIRRE, J. L., Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México con aspectos zoogeográficos y ecológicos. Dir. Gral. Inst. Nal. Pesca, México, Serie Científica, 1978. 1-298. 19: CERVIGÓN, M. F., Los Peces Marinos de Venezuela. Estación de Investigaciones Marinas de Margarita, Fundación la Salle de Ciencias Naturales, Caracas, 1966. 1-951. Monogr. 11 y 12: CHACKO, P.I., Food and feeding habits of the fishes of the Gulf of Mannar. Proc. Indian Acad. Sci., 1949. 83-97 29 (3), See. B: . CHARLES, R., Thesis, Aspects of the biology of the mojarra Eucinostomus gula (Qouy y Gaimard), in the Biscayne Bay, University of Miami, Florida. M. Sc. U.S.A. 1975. CHAVANCE, P., D. FLORES HERNÁNDEZ, A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología, Biología y dinámica de las poblaciones de Bairdiella chrysoura en la Laguna de Términos, Sur del Golfo de México. (Pisces: Scianidae). 1984. 123-162 11 (1): CHAVANCE, P. A. YÁÑEZ-ARANCIBIA, D. FLORES HERNÁNDEZ, A. L. LARA-DOMÍNGUEZ y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecology, biology and populations dynamics of Archosargus rhomboidalis (Pisces- Scianidae) in a tropical coastal lagoon, Southern Gulf of Mexico. 1986. 13 (2): (en prensa). CHÁVEZ, E., Notas acerca de la ictiofauna del estuario del río Tuxpan y sus relaciones con la temperatura y la salinidad. In: Carranza, J. (Ed.). Mem. IV. Congr. Nal. Ocenogr. México: 1972. 177-199. CHÁVEz, E., B. A. VELEZ, F. A. SÁNCHEZ y J. S. CHÁVEZ, Prospección ecológica de las Lagunas de Mandinga, Veracruz. In: Mem. I Reunión Latinoamericana sobre Ciencia y Tecnología de los Océanos, Veracruz, México: 1976. 140-162. CYRUS, D. P. y S.J.M. BLABER, Species identification, distribution and a bundance of Gerridae (Telostei) Bleeker 1958, in the estuaries of Natal. S. Afr. 1. Zool., 1982a. 105-116. 17 (3): CYRUS, D.P. y S.J. M., BLABER, Mouthpart structure and function and the feeding mechanisms of Gerres (Telostei). S. Afr. I. Zool., 1982b. 117-121. 17 (3): CYRUS, D. P. y S.J.M., BLABER, The food and feeding ecology of Gerridae (Bleeker, 1859) in estuaries of Natal. four. Fish. Biol., 1983. 373-394. 22 (4): DARNELL, R. M., Fishes of the Rio Tamesi and related coastal lagoons in East-Central Mexico. Publ. Inst. Mar. Sci., Univ. Texas, 1962. 299-365. 8: DARNELL, R. M., R. E. DEFENBAUGH y D. MOORE, Northwestern Gulf shelf bio- atlas: a study of the distribution of demersal fishes and peneid shrimp of soft bottons of the continental shelf from. the Rio Grande to the Mississippi River Delta. Open File Report No. 82-04. Metairie, L. A. Minerals Management Service, Gulf of Mexico. Regional Office: 1993. 438 p. DAY, F., Fishes of India. William Dawson and Sons Ltd., London (Rep. in 1958). 1878. DEEGAN, L. A. y B. A. THOMPSON, Ecology and fish communities of the Mississippi River Deltaic plain region. In: Fish Community Ecology in Estuaries and Coastal Lagoons: Towards and Ecosystem Integration. Yáñez-:Arancibia, A. (Ed.). Editorial Universitaria, UNAM/PUAL/ICML, México. 1985. DÍAZ RUIZ, S., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Taxonomía, diversidad, distribución y abundancia de los pomadásidos de la Laguna de Términos, Campeche (Pisces: Pomadasyidae). 1982. 251-278. 9 (1): FISCHER, W. (Ed.), FAO Species Identification Sheets for Fishery Purpose. Western Central Atlantic (Fishing area 31) FAO, 2. Roma, 1977. FLORES-COTO, C. y J. ALVAREZ-CADENA, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Estudios preliminares de distribución y abundancia del ictioplancton en la Laguna de Términos, Camp. 1980. 67-78. 7 (2): GONZALEZ SANSON, G. y L. RODRÍGUEZ VIÑAS, Alimentación natural de Eugerres brasilianus (Cuvier) y y Gerres cinereus (Walbaum) (Pisces: Gerreide) en las lagunas costeras de Tunas de Zaza, Cuba, Revista de Investigaciones Marinas, 1983. 91-134. 4 (1): GUITART, J. D., Sino psis de los Peces Marinos de Cuba. serie Oceanológica, Academia de Ciencias de Cuba, 1977. 325-608. 3: HARRINGTON, R. W. y E. A. HARRINGTON, Ecology, Food selection among fishes invading a high subtropical salt marsh; from onset of flooding throught the progress of a mosquito brood. 1961. 646-660. 42 (4): HILDEBRAND, H. H., A study of fauna of the brown shrimp (Penaeus aztecus Ives) graunds in the western Gulf of México. Publ. Inst. Mar. Sci. Univ. Texas, 1954. 233-366. 3 (2): HILDEBRAND, H. H., A study of fauna of the pink shrimp (Penaeus duorarum Burkenroad) grounds in the Gulf of Campeche. Publ. Inst. Mar. Sci. Univ. Texas, 1955. 151-173. 4 (1): HILDEBRAND, H. H., Ciencia. Estudios biológicos preliminares sobre la Laguna Madre de Tamaulipas. México, 1958. 151-173 17 (7-9): HOESE, H. D. y R. S. JONES, Seasonality of larger animals in a Texas turtle grass community. Publ. Inst. Mar. Sci. Univ. Texas, 1963. 37-48. 9: HOESE, H. D. y R. H., MOORE, Fishes of the Gulf of Mexico, Texas, Louisiana and Adjacent Waters. Texas A. M. University Press College Station and London, 1977. 327 p. JHINGRAN, V. H., Bull. Report on the fisheries of the Chilka Lake 1957-60. Centr. Inl. Fish. Res. Inst., 1963. 1-113. 1: JHINGRAN, V. y G. A. V. NATAJARAN, Bull. Final Report on the fisheries of the Chilka Lake (1957-65). Centr. Inl. Fish. Res. Inst., 1966. 8. JOB, T. J., An investigation on the nutrition of the perches of the Madras coast. Rec. Indian Mus., 1940. 289-364. 42: KILBY, J. D., The fishes of the Gulf coastal marsh areas the Florida. Tul. Stud. Zool., 1955. 176-247. 2 (8): LAEVASTU, T., Manual de Métodos de Biología Pesquera. Publicación FAO. Ed. Acribia, España, 1971. 243 p. LARA-DOMINGUEZ, A. L., A. YAÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Biología y ecología del bagre Arius melanopus Günther, en Laguna de Términos, Sur del Golfo de México. 1981. 267-304. 8 (1): LINTON, E., Bull. Parasites of the fishes of Beaufort. U. S. Bureau of fisheries Comm., 1905. 321-428. 24: LONGLEY, W. H. y S. F. HILDEBRAND, Systematic catalogue of the fishes of Tortugas Florida. Pap. Tortugas Lab., 1941. 1-331 34 (535): MALLARD COLMENERO, L.A., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Taxonomía, diversidd, distribución y abundancia de los tetraodóntidos de la Laguna de Términos, Sur del Golfo de México. (Pisces: Tetraodontidae). 1982. 161-211. 9 (1): MEEK, E. S. y S. F. HILDEBRAND, The marine fishes of Panama. Field. Mus. Nat. Hist. Publ., Zool. Ser., 1925. 1-1045. 15 (226): MÉNDEZ, V. S. F. y A. VELARDE MÉNDEZ, Tesis Profesional Estudios del Ictioplancton en la Boca del Carmen, Laguna de Términos, Campeche. Fac. Ciencias, Univ. Nal. Autón. México, 1982. 77 p. MILLER, R. R., Geographical distribution of central american freshwater fishes. In: T. B. Thorson (Ed.) Investigations of the Ichthyofauna of Nicaraguan Lakes. School of Life Sciences, University of Nebraska-Lincoln, Lincoln, Nebraska: 1976. 125-156. NIKOLSKY, G. V., The Ecology of Fishes. Academic Press Inc. New York. 1963. 352 p. ODUM, W. E., Utilization of the direct grazing and plant detritus food chains by the striped mullet Mugil cephalus. In: Steek, J. (Ed.). Proc. Symposium on Marine Food Chains, Edimburgo. Oliver and Boyd, London: 1970. 222-240. ODUM, W. E., Bull. Pathways of energy flow in a South Florida estuary. Sea Grant Program Tech. Univ. Miami, 1971. 1-162 7: ODUM, W. E. y J. HEALD, Bull. Trophic analysis of an estuarine mangrove community. Mar. Sci., 1972. 671-738. 22 (3): PERRY, J. A. II y S. D. PERRY, Los peces Comunes de la Costa Atlántica de Costa Rica. Univ. de Costa Rica. Fac. Cienc. y Letras. Departamento de Biología 1974. 1-224 p. (7): PINKAS, L., M. S. OLIPHANT, y I. L. K. IVERSON, Bull., Food habits of albacore, blue fin tuna and bonito in California waters. Dept. Fish. and Game Cal. Fish. 1971. 1-105. 152: PRABHAKARO RAO, A.V., Observations on the food and feeding habits of Gerres oyena (Forskal) and Gerres filamentosus Cuvier from the Pulicat Lake with notes on the food allied species. J. Mar. Biol. Ass. India, 1968. 332-346. 10 (2): RANDALL, J. E., Ecology, Grazing effect on sea-grass by herbivorous reef fishes in the West Indies. 1965. 255-260. 46 (2): RANDALL, J. E., Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr. Miami, 1967. 665-847. 5: REID, G. K., Bull. An ecological study of the Gulf of Mexico fishes in the vecinity of Cedar Key. Florida. Mar. Sci. Gulf. Caribb., 1954. 1-94. 4 (1): RESENDEZ A., An. Inst. Biol. Univ. Nal. Autón. México, Ser, Cienc. del Mar y Limnol., Estudio de los peces de la laguna de Tamiahua, Veracruz, México. 1970. 79-146. 41 (1): RESENDEZ, M. A., Estudio de los peces de la laguna de Alvarado, Veracruz, México. Revta. Soc. Mex. His. Nat., 1973. 183-281 34: RESENDEZ M. A., Biótica, Estudio de los peces de la Laguna de Términos, II. Campeche, México. 1981. 345-430. 6 (4): RICKER, W. E., Linear regressions in Fishery Research. J. Fish. Res. Board Can., 1973. 409-434. 30: RICKER, W. E., Bull. Computation and interpretation of biological statistics of fish populations. Fish. Res. Board. Can., 1975. 1-382. 191: RODRÍGUEZ CASTELLANOS, V. E., Contribución al conocimiento biológico de las mojarras (Pisces: Gerreidae) de la Cíénega de la Virgen. Boletín Facultad de Biología Marina, 1983. 8. (1): RUBIO, C. E., Crecimiento, sexualidad y desarrollo gonadal de la mojarra rayada, Eugerres plumieri (Cuvier) de la Ciénega Grande de Santa Marta con anotaciones sobre su biología. Divul. Pesq., 1975. 37. 9 (1): SÁNCHEZ GIL, P., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LiNARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Diversidad, distribución y abundancia de las especies y poblaciones de peces demersales de la Sonda de Campeche (Verano, 1978). 1981. 209-240. 8 (1): SHERIDAN, P. F. y R. J. LIVINGSTON, Cyclic throphie relationship of fishes in an unpolled, river-dominated estuary in north Florida. In: Ecological Processes in Coastal and Marine Livingston, R.J. (Ed.). Sci. Plenum Press. N.Y., 1979. 143-161. 10: SMITH, H. N., The fishes of North Carolina Raleigh. North Carolina, Geology and Economics, Survey, 1907. 14-53. 2: SPRINGER, S. y H. R. BULLIS, Collections by the "Oregon" in the Gulf of México, U.S., Fish. Wildl. Serv., Spec. Sci. Rept., 1956. 1-134 196: SPRINGER, V.G. y K.D. WOODBURN, An ecological study of the fishes of Tampa Bay Area. Fla. St. Bd. Conserv. Mar. Lab. Prof. Pap., 1960. 1-104 1: SPRINGER, V. G. y A. J. MCERLEAN, Bull. Seasonality of fishes on a South Florida shore. Mar. Sci. Gulf Caribb., 1962. 39-60 12 (1): TABB, D.C., D.L. DUBROW y R. B. MANNING, The ecology of northern Florida Bay and adjacent estuaries. Florida State Board of Conservation, 1962. 1-80 39: TORAL, S., y A. RESENDEZ, Los cíclidos (Pisces: Perciformes), de la Laguna de Términos y sus afluentes. Rev. Biol. Trop., 1973. 259-279. 21 (2): VARGAS MALDONADO, I., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología y estructura de las comunidades de peces en áreas de Rizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos, Sur del Golfo de México. 1981. 241-246. 8 (1): WALDINGER, F. J., Thesis. Relationships of environmental parameters and catch of three species of the mojarra family (Gerridae), Eucinostomus gula, E. argenteus and Diapterus plumieri collected in 1963 and 1964 in Buttonwood Canal, Everglades National Park, Florida, M. Sci. Univ. 1969. WALLS, G.J., Fishes of the Northern Gulf of Mexico. T.F.H. Publications, Inc. Ltd., U.S.A. 1975. 417 p. YÁÑEZ-ARANCIBIA, A., An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Sobre los estudios de peces en las Lagunas costeras: Nota Científica. 1975. 53-60. 2 (1): YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Taxonomía, ecología y estructura de las comunidades de peces en lagunas costeras con bocas efímeras del Pacífico de México. Publ. Esp., 1978. 1-306. 2: YÁÑEZ-ARANCIBIA, A., Ecology in the entrance of Puerto Real Términos Lagoon. IL Discussion on trophic structure of fish community in banks of Thalassia testudinum. In: Present and Future Research on Coastal Lagoons. Lasserre, P., H. Postma, J. Costlow and M. Steyaert (Eds.). II UNESCO/IABO. Tech. Pap. Mar Sci. UNESCO, 1981. 191-232. 33: YÁÑEZ-ARANCIBIA, A., Usos, recursos y ecología de la zona costera. Ciencia y Desarrollo (CONACYT), 1982. 58-63. 43 (VII): YÁÑEZ-ARANCIBIA, A. y R. S. NUGENT, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, El papel ecológico de los peces en estuarios y lagunas costeras. 1977. 287-306. 5 (1): YÁÑEZ-ARANCIBIA A. y F. AMEZCUA LINARES, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de Urolophus jamaicensis (Cuvier) en Laguna de Términos, un sistema estuarino al Sur del Golfo de México. (Pisces: Urolophidae). 1979. 107-114. 6 (2): YÁÑEZ-ARANCIBIA, A. y J. W. DAY, Jr., Ecological characterization of Terminos Lagoon a tropical lagoonestuarine system in the southern Gulf of México. In: Coastal Lagoons. Oceanologica Acta. Lasserre, P. y H. Postma (Eds.). 1982. 417-429. Vol. Spec., 5 (4): YÁÑEZ-ARANCIBIA, A. y A. L. LARA-DOMÍNGUEZ, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Dinámica ambiental de la Boca de Estero Pargo y estructura de sus comunidades de peces en cambios estacionales y cielos de 24 hrs (Laguna de Términos, Sur del Golfo de México). 1983. 85-116. 10 (1): YÁÑEZ-ARANCIBIA, A., J. CURIEL-GÓMEZ y V. LEYTON, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Prospección biológica y ecológica del bagre marino Galeichthys caerulescens (Günther) en el sistema lagunar costero de Guerrero, México. (Pisces: Ariidae). 1976. 125-180. 3 (1): YÁÑEZ-ARANCIBIA A., F. AMEZCUA LINARES y J. W. DAY, Jr., Fish community structure and function in Terminos Lagoon, a tropical. estuary in southern Gulf of Mexico. In: Estuarine Perspectives. Kennedy, V. (Ed.), Academic Press Inc. New York: 1980. 465-482. YÁÑEZ-ARANCIBIA, A., F. AMEZCUA LINARES y M. TAPIA GARCÍA, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Prospección ictioecológica del estuario del Rio Champotón, Verano 1979. Campeche, México. 1982a. 395-398. 9 (1): YÁÑEZ-ARANCIBIA, A. y A. L. LARA-DOMÍNGUEZ y H. ALVAREZ GUILLÉN, Fish community ecology and dynamics in estuarine inlets. In: Fish Community Ecology in Estuaries and Coastal Lagoons: Towards an Ecosystem Integration. Yáñez-Arancibia, A. (Ed.). Editorial Universitaria, UNAM/PUAL/ICML, México. 1985a. YÁÑEZ-ARANCIBIA, A., A. L. LARA DOMÍNGUEZ, P. CHAVANCE y D. FLORES HERNANDEZ, An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Environmental behavior of Términos Lagoon ecological system, Campeche, México. 1983a. 137-176. 10 (1): YÁÑEZ-ARANCIBIA, A., P. SÁNCHEZ-GIL., M. TAPIA GARCÍA y M. DE LA C. GARCÍA ABAD, Ecology, community structure and evaluation of tropical demersal fishes in the southern Gulf of Mexico. Cahiers de Biologie Marine, 1985. 137-163. 26: YÁÑEZ-ARANCIBIA A., A. L. LARA-DOMÍNGUEZ, A. AGUIRRE LEÓN, S. DÍAZ Ruiz, F. AMEZCUA LINARES, D. FLORES HERNANDEZ y P. CHAVANCE, Ecology of dominant fish populations in tropical estuaries: Environmental Factors Regulating Biological Estrategies and Production. In: Fish Community Ecology in Estuaries and Coastal Lagoons: Towards an Ecosystem Integration. Yáñez-Arancibia, A. (Ed.). Editorial Universitaria, UNAM/PUAL/ICML. México. 1985c. YÁÑEZ-ARANCIBIA, A., F. AMEZCUA LINARES, P. SANCHEZGIL, I. VARGAS MALDONADO, A. L. LARADOMÍNGUEZ, S. DÍAZ Ruiz, A. AGUIRRE LEÓN y P. CHAVANCE, Estuaries, Estuary-shelf fish community interactions in Southern Gulf of Mexico: Fish-habitat comparisons. 1981. 295. 4 (3): YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. SANCHEZ-GIL, I. VARGAS MALDONADO, P. CHAVANCE, F. AMEZCUA LINARES, A. AGUIRRE LEÓN y S. DIAZ Ruiz, Coastal Lagoons. O. Ecosystem dynamics and nichthemeral and seasonal programming of fish community structure in a tropical estuarine inlet, México. In: Lasserre, P. y H. Postma (Eds.). Oceanologica Acta. 1982b. 431-44 Vol. Spec., 5 (4): YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. SANCHEZ CIL, I. VARGAS-MALDONADO, MA. DE LA C. CARCÍA ABAD, H. ALVAREZ GUILLÉN, M. TAPIA GARCIA, D. FLORES HERNANDEZ y F. AMEZCUA LINARES, Ecology and evaluation of fish community in the Campeche Sound and Terminos Lagoon: Estuary-shelf interactions in the Southern Gulf of México. Atlantica Rio Grande, 1982c. 130. 5 (2): YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. SANCHEZ-GIL, I. VARGAS MALDONADO, MA. DE LA C. GARCÍA-ABAD, H. ALVAREZ GUILLÉN, M. TAPIA GARCÍA, D. FLORES HERNANDEZ y F. AMEZCUA LINARES, Ecology and evaluation of fish community in the Campeche Sound and Terminos Lagoon: Estuary-shelf interactions in the Southern Gulf of Mexico. In: . Fish Community Ecology in Estuaries and Coastal Lagoons: Towards and Ecosystem Integration. México. Yáñez-Arancibia, A. (Ed.) Editorial Universitaria, UNAM/PUAL/ ICML, 1985d. YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMÍNGUEZ, P. SÁNCHEZ-GIL, H. ALVAREZ GUILLÉN, I. VARGAS MALDONADO, A. AGUIRRE LEÓN, M. TAPIA GARCÍA, MA. DE LA C., GARCÍA ABAD, D. FLORES HERNÁNDEZ, P. CHAVANCE, F. AMEzcuA LINARES, S. DíAZ RUIZ, M. ALVAREZ RuBio y J. L. ROJAS GALAVIZ, Informe Final. Caracterización ambiental del sistema ecológico y análisis comparativo de las poblaciones de peces de la Sonda de Campeche y de la Laguna de Términos antes y después del derrame petrolero del Pozo IXTOC-I. PCEESC/Sría. d e Marina/Sría de Pesca/PEMEXIUNAM-ICML (IF), 1982d. 4 partes, 221 p., 22 tablas, 49 figs. YASUDA, F., Bull., The type of food habits of fishes assured by stomach contents examination. Japanese Soc. Sci. Fish., 1960. 653-662. 26 (2):
|