|
BIOGEOGRAFÍA DE CORALES
HERMATÍPICOS: EL ISTMO CENTRO AMERICANO
Trabajo recibido el 3 de
mayo de 1984 y aceptado para su publicación el 28 de octubre de
1984.
JORGE CORTÉS N.
CIMAR, Universidad de Costa Rica San Pedro, Costa Rica.
Contribución del Centro de Investigación en Ciencias del Mar y
Limnología (CIMAR).
Las faunas actuales de
corales hermatípicos a ambos lados del istmo centro americano son muy,
diferentes. Dos hipótesis han sido propuestas para explicar estas
diferencias: 1- Heck y McCoy; modificación de una fauna coralina
pan-Tética por eventos tectónicos especiacion Y-
extinción. 2- Dana, eliminación de las faunas coralinas del
Pacífico Este en el Pleistoceno y posterior transporte
transoceánico de plánulas desde el Indo-Pacífico oeste.
Las dos hipótesis son discutidas y evidencia en favor de la
hipótesis de Dana es presentada.
Palabras
clave: Biogeografía, corales hermatípicos, Centro
América, Pacífico Este.
Present day,
faunas of hermatylpic corals on both sides of Middle America are very,
different. Two hypothesis have been proposed to explain the differences: 1-
Heck and McCoy.'s, modification of a pan-Tethyan coraline fauna by tectonic
events, speciation and extinctions. 2- Dana's; elimination of the Eastern
Pacific coraline faunas during the Pleistocene, and hence, transoceanic
transport of planulae accounts for the present day fauna. The two hypothesis
are discussed and evidence is marshalled in favor of Daria's
position.
Key Words: Biogeography,
hermatypic corals, Central America, Eastern Pacific.
Los océanos Pacífico y Atlántico se hallan separados en la zona neotropical por una estrecha faja de tierra (50 km en Panamá, Leschine, 1981); Centro América. Esta área ha constituido un puente para organismos terrestres (Simpson, 1964; Whitmore y Stewart, 1965; Marshall et al., 1979, 1982; Rich y Rich, 1983), pero una barrera para organismos marinos (Woodring, 1954, 1966; Newell, 1971; Hallam, 1973). La provincia faunística Caribeña del Terciario incluía el Caribe y el Pacífico Este (Woodring, 1966). Después de la separación de los océanos, Ekman (1953) consideró ambas costas de Centro América como parte de una misma provincia debido a la similitud de algunos grupos marinos. Posteriormente, Briggs (1974), divide el área en dos regiones: Pacífico Este (separado del Pacífico Oeste) y Atlántico Oeste (se- parado del Atlántico Este). Sin embargo, al considerar las faunas coralinas modernas, el surgimiento del istmo de Centro América divide los océanos en dos grandes provincias biogeográficas (Fig. 1): 1- Provincia del Indo-Pacífico, que se extiende desde la costa este de Africa y el Mar Rojo, a través del Mar Indico y el Pacífico, hasta la costa oeste del continente americano. 2- Provincia del Caribe o Atlántico, que se extiende desde la costa este de Norte América a través del Mar Caribe y la costa este de América del Sur, hasta la costa oeste de Africa (Stehli y Wells, 1971; Goreau et al., 1979). La separación del Pacífico Este Tropical y del Mar Caribe, es tan corta en distancia como relativamente reciente, habiendo existido por solo 3 millones de años (Emiliani et al., 1972; Keigwin, 1978, 1982 a, b; Fischer, 1981). Para una explicación amplia de la historia tectónica y la paleogeografía de Costa Rica, revísese Rich y Rich (1983). En la figura 2, se resumen esquemáticamente las conecciones entre el Caribe y el Pacífico Este. Durante el Cretácico Superior la separación de estos océanos era parcial, posteriormente aumenta el intercambio hasta el Eoceno. Durante el Oligoceno la separación es parcial y en el Mioceno Superior se termina la conexión de aguas profundas. Perdura la conexión somera hasta el Plioceno. La emersión total de Costa Rica y Panamá hace unos 3 MA separa al Pacífico y el Atlántico hasta nuestros días. 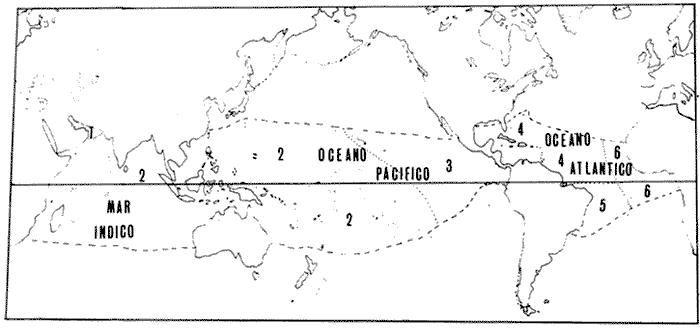 Fig. 1. Mapa del Mundo en el que se indican las dos grandes provincias biogeográficas de corales: 1-3 Indo-Pacífico y 4-6 Caribe-Atlántico; y las subprovincias- 1- Mar Rojo, 2- Indo-Pacífico Oeste y Central; 3- Pacífico Este; 4- Caribe; 5- Brasil, 6- Africa. Basado en Stehli y Wells, 1971; Laborel 1974; Goreau et al., 1979. Los planes para la construcción de un canal a nivel en Panama han promovido mucha investigación en el área del istmo. Los resultados de algunas de estas investigaciones fueron presentados en un simposio sobre la Biota Panámica (Jones, 1972). Posteriormente Glynn (1982) ahonda sobre el tema, ofreciendo excelentes comparaciones entre los arrecifes del Caribe y del Pacífico Este, proporciona además un cuadro sinóptico donde se dan características físicas y biológicas de los arrecifes y comunidades coralinas a ambos lados del istmo centro americano. En el presente trabajo se hace énfasis en las especies de corales hermatípicos (corales formadores de arrecifes) y se presentan las dos hipotésis biogeográficas que tratan de explicar la distribución actual de estos corales. COMPARACIÓN DE LAS FAUNAS CORALINAS Y ORGANISMOS ASOCIADOS A ARRECIFES DEL CARIBE Y DEL PACÍFICO ESTE
CORALES HERMATÍPICOSLas comunidades de corales hermatípicos a ambos lados de Centro América son sorprendentemente diferentes (Porter, 1974). Solamente hay dos géneros de corales en común entre el Pacífico Este y el Caribe: Porites y Millepora; no hay ninguna especie en común. Los arrecifes del Caribe tienen cerca de 75 especies de corales pertenecientes a quince familias del Orden Scleractinia: Acroporidae, Agariciidae, Astrocoeniidae, Caryophylliidae, Dendrophylliidae, Faviidae (familia dominante en el Caribe), Flabellidae, Guyniidae, Meandrinidae, Mussidae, Oculinidae, Pocilloporidae, Poritidae, Rhizangiidae, Siderastreidae e hidrozoarios: Milleporidae (Wells, 1973; Smith, 1976; Caims, 1982). El Pacífico Este tiene cerca de 22 especies de corales escleractíneos en cinco familias: Agariciidae, Dendrophylliidae, Pocilloporidae (dominante en los arrecifes de Panamá), Poritidae, Thamnasteriidae y el hídrozoario: Milleporidae (Durham, 1966; Glynnet al., 1972; Glynn et al.,1983). 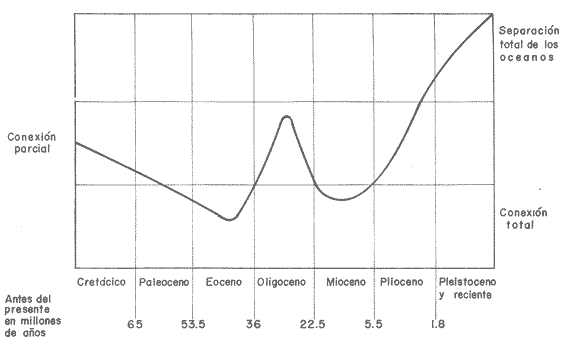 Fig. 2 Resumen esquemático de las conexiones entre el Caribe y el Pacífico Este. Basado en Simpson, 1964; Savage, 1966; Windely, 1978. Los corales hermatípicos del Pacífico Este Tropical están asociados a los corales modernos del Indo-Pacífico Oeste (Stehli y Wells, 1971; Glynn et al., 1972; Dana, 1975). Los corales del Atlántico por su parte, se consideran una reliquia de la fauna coralina del Terciario Medio del Mar de Tetis (Wells, 1956). OTROS ORGANISMOSAl analizar otros grupos de organismos asociados a arrecifes coralinos encontramos lo siguiente: 1- organismos presentes sólo en el Caribe o en el Pacífico y 2- organismos presentes en el Caribe y en el Indo-Pacífico Oeste, pero no en el Pacífico Este. Dentro de la primera categoría se puede citar el equinodermo asteroideo deprededor de corales, Acanthaster planci. Acanthaster se encuentra en el Indo-Pacífico y en los arrecifes de la costa Pacífica de Costa Rica y Panamá, pero no en el Caribe (Glynn, 1982). Los peces loro (Scaridae) del Pacífico Este no están relacionados con los del Atlántico, con la excepción de Nicholsina denticulata. Esto contrasta con los patrones de distribución de otras familias de peces de aguas someras no asociadas directamente con corales representados en el continente americano (Rosenblatt y Hobson, 1969). Se encuentran muchos ejemplos de especies de aguas someras con un representante en el Atlántico y su cognado (o pariente más cercano) en el Pacífico (W. Bussing, 1981, com. pers.) Las faunas de moluscos del Caribe y del Pacífico Este empiezan a divergir en el Mioceno, concordante con la separación de los mares por el istmo centroamericano (Kruckow, 1982). En el Pacífico Este, se han registrado 41 especies de gasterópodos prosobranquios y 7 especies de bivalvosde afinidad Indo Pacífica. Casi todas estas especies tienen formas larvales con períodos planctónicos largos (Emerson, 1978). Garth (1974) describe decápodos comensales de corales hermatípicos del Indo-Pacífico presentes en los corales del Pacífico Este, pero atisentes en el Caribe. El segundo patrón de distribución consiste en grupos presentes en el Caribe y en el IndoPacífico Oeste pero no en el Pacífico Este. Por ejemplo, el congrejo Phymodius maculatus, se encuentra en él Caribe y en el Indo-Pacífico Oeste pero no en el Pacífico Oeste (J. Garth, 1981, com. pers.) Otro ejemplo es el llamado "pasto de tortuga", Thalassia, el cual se encuentra abundantemente en las lagunas de los arrecifes del Caribe (T.testudinum) y del Pacífico Oeste (T. hemprichii) pero no en el Pacífico americano (Pettittet al., 198 l). Es más, probablemente esta fanerógama marina estaba presente en el Pacífico Este en el Eceano (Eva, 1980). HIPÓTESIS PROPUESTASe han propuesto dos hipótesis para explicar las diferencias entre los corales a ambos lados de Centro América: 1- Eliminación completa de las comunidades coralinas del Pacífico Este en el Pleistoceno y subsiguiente transporte transoceánico de plánulas de corales desde el Indo-Pacífico Oeste (Dana, 1975). 2- Modificación de las comunidades coralinas pan-Téticas, pre-emersión del istmo, por eventos tectónicos, especiación y extinciones (McCoy y Heck, 1976; Heck y MeCoy, 1978). HIPÓTESIS DE DANALa hipótesis que propone Dana (1975) requiere ante todo una eliminación completa de las comunidades coralinas del Pacífico Este en el Pleistoceno. Apoyan esta suposición las siguientes evidencias: 1-Después del surgimiento del istmo centroamericano, las condiciones para la existencia de corales en el Pacífico Este se deterioraron. La temperatura del agua disminuyó debido a la circulación de los océanos y a fenómenos de surgencia de aguas (Emiliani et al.,1972; Birkeland, '1977; Keigwin, 1982 a). Al mismo tiempo, la franja tropical se fue reduciendo desde el Terciario Temprano (Kurtén, 1972; Frenzel, 1973; Goudie, 1977; Revelle y Thompson, 1982). 2- Las fluctuaciones en el nivel del mar durante el Pleistoceno contribuyeron al deterioro de las con diciones vitales de los corales en el Pacifico Este. Debido a que el Pacífico americano tiene una plataforma muy angosta, un cambio de unos 100 m en el nivel del mar, dejaría muy pocos lugares habitables para los corales. Al mismos tiempo un cambio igual en el Atlántico no ha de haber tenido un impacto tan drástico en la reducción del hábitat de los corales (P. Sprechman, com. Pers., 1983). 3- La evidencia fósil presentada por Durham (1966) apoya esta idea de la eliminación de la fauna coralina del Pacífico Este, empezando desde el Plioceno. Algunos géneros del Atlántico-Tetis Oeste presentes en el Pacífico Este, aparecen por última vez en el Plioceno, igual que varios géneros del Indo-Pacífico (Durham, 1966). 4- No se han encontrado arrecifes del Pleistoceno en la costa oeste de América y los arrecifes actuales del Pacifico Este empezaron a desarrollarse hace 4500 a 5600 años sobre rocas volcánicas y sedimentos calcáreos (Glynn y Macintyre, 1977). La segunda premisa de la hipótesis de Dana establece que los corales hermatípicos del Pacífico Este actual derivan de las invasiones de larvas del Indo-Pacífico (Pacífico Central e Islas del Pacífico Ecuatorial) desde finales del Pleistoceno. Evidencias: 1- La evidencia actual sobre la duración del período planetónico de las Plánulas es muy limitada (Connell, 1973), sin embargo, podría ser de hasta vanos meses. 2- Existen corrientes trans-Pacíficas, la contracorriente ecuatorial, que mueve agua del Pacífico Central al Pacífico Este (Wyrtki, 1973). 3- La dispersión a larga distancia de larvas al azar es una posibilidad que se debe considerar (Carlquist, 1981). 4- Se ha sugerido el transporte transoceanico para explicar la presencia de otros grupos por ejemplo gastrópodos prosobranquios y bivalvos del Pacífico Este con afinidad IndoPacífica, cuyas formas larvales pueden permanecer hasta diez meses en el plancton (Emerson, 1978). HIPÓTESIS DE HECK Y McCOYLa hipótesis propuesta por Heck y MeCoy (1978) requiere primero: una comunidad coralina pan-Tética ampliamente distribuida. Apoya esta idea la siguiente evidencia: 1- Durante el Cretácico, y el Terciario temprano, había muchos géneros de corales en común entre el Pacífico Este y Atlántico-Tetis (durham, 1966). Cuatro genéros del Caribe actual (Dichocoenia, Diplora, Eusmilia y Siderastrea) han sido encontrados en depósitos del Plioceno en el Mar de Córtes (Durham y Barnard, 1952). 2 - Especímenes de Pocillopora, género del Indo-Pacífico, hansido encontrados en depósitos de Pleistoceno Superior del Caribe (Geister, 1977) y fragmentariamente en laFormación Río Banano (L. D. Gómez, com. pers., 1982). La segundo premisa de la hipótesis de Heck y McCoy propone que la comunidad coralina pantética, pre-emersión del istmo, fue modificada por eventos tectónicos, especiación y extinción. Evidencias: 1- Eventos tectónicos y climáticos a largo plazo, causan cambios en el ambiente, los cuales redundan en especiaciones y extinciones (Vallentine, 1971; Croizat et al., 1974; Rosen, 1975). 2- Corales y rudistas del Caribe empezaron a diferenciarse de las del Mediterráneo durante el Cretácico Medio, debido al aislamiento geográfico producido por la expansión del Océano Atlántico (Coates, 1973). 3- Estudios de las faunas de moluscos paleocaribeñas muestran claramente cómo por eventos tectónicos se fueron diferenciando, primero las faunas del Caribe de las de Europa-Mediterráneo, y luego las faunas del Caribe y las del Pacífico Este (Kruckow, 1982). DISCUSIÓNLa interpretación vicarista de la distribución tal como la presentan Heck y McCoy (1978), incorpora las ideas biogeográficas, evolucionistas y geofísicas más recientes. Sin embargo, no es la explicación más realista por las siguientes razones: 1. Los fósiles de corales del Terciario tienen poca relación con las formas actuales presentes en el Pacífico Este (Durham, 1966; Glynn et al., 1972). 2. La fauna coralina de la costa Pacífica de América pre-Plioceno era típicamente pantética, mientras que la actual está más relacionada con la fauna Indo-Pacífica (Durham, 1966). La fauna actual del Atlántico tiene restos de una fauna pan-Tética (Wells, 1956; Glynn, 1982). Un análisis de grupo ("Cluster Analysis") de los géneros de corales hermatípicos muestra que tienen más relación los corales del Pacífico-Este con los del IndoPacífico, que con los del Atlántico (Stehli y Wells, 1971). 3. Los corales tienen un registro paleontológico relativamente bueno, y ha sido posible determinar las edades de muchos géneros y especies. Gracias a esto, se ha comprobado que más del 50 % de las especies que existieron hace diez millones de años, están vivas hoy (Stanley, 1979). Más aún, además de vivir muchos años, los corales tienen el potencial de producir varias generaciones que se traslapan y producen pocas generaciones en períodos de tiempo muy largos (Potts, 1983). Esto significa que los tres millones de años desde que se separaron el Atlántico y el Pacífico, no han sido suficientes para justificar la diferencia entre las especies de corales en ambos lados de Centro América (Birkeland), 1977). La interpretación biogeográfica de centro de origen y dispersión para corales hermatípicos (Dana, 1975), parece ser la más acertada por las siguientes razones: 1 . La eliminación completa de las faunas coralinas del Pacífico Este en el Pleistoceno es probable debida a los cambios en la temperatura y en el nivel del mar. La evidencia paleontológica apoya esta premisa: No hay arrecifes del Pleistoceno, y el registro de muchas especies llega hasta el Plioceno o Pleistoceno. 2. El transporte transoceánico de plánulas es factible aunque casi imposible de probar. 3. El período de separación parece ser muy corto para que se haya dado suficiente divergencia evolutiva que pueda justificar la diferencia entre las faunas coralinas del Caribe y del Pacífico Este (Birkeland, 1977; Stanley, 1979; Potts, 1983). ConclusionesLas faunas coralinas del Pacífico Este posiblemente derivan de emigrantes del Indo-Pacífico como lo propone Dana (1975), y no representan restos de una fauna ampliamente distribuida que sufrió modificaciones como lo indican Heck y McCoy (1978). "The biogeographer who deals with patterns probably created by long distance dispersal must use evidence that is circunstantial, indirect and subjective, and therefore vulnerable. However, if that kind of evidence leads to plausible answers, we cannot afford to rule out of court" (Carlquist, 1981, p. 515). AgradecimientosUn primer borrador de este manuscrito fue redactado mientras disfrutaba de una beca otorgada por McMaster University, Hamilton, Canadá y un complemento económico del CONICIT. Mike Risk leyó ese manuscrito y motivó a mejorarlo. M. M. Kandler ayudó a todo lo largo de la producción de este documento. Finalmente agradezco a las siguientes personas que leyeron este manuscrito y aportaron mejoras: Ricardo Soto, CIMAR; Peter Sprechman, Escuela Centroamericana de Geología; Luis Diego Gómez, Museo Nacional de Costa Rica. Norma Bermúdez hizo el trabajo de mecanografía. NOTAAl terminar de preparar este trabajo, se recibió una comunicación de Henry von Prahl en la que menciona el haber encontrado un coral del género Acropora en Gorgona, Colombia. De confirmarse este hallazgo, el número de géneros en común entre el Pacífico Este y el Caribe sería de tres y habría que agregar la familia Acroporidae a la lista de cinco familias del Pacífico Este. Todavía falta el estudio de esta especie para determinar su relación con otras especies deAcropora. LITERATURABIMELANED, C., The importance of rate of biomass accumulation in cartly successional stages of benthic communities to the survival of coral recruits. Proc. 3rd. Int. Coral Reef Symp., 1977. 15-21 1 BRIGGS, J. C., Marine Zoogeography. MeGraw-Hill Book Company. E. U. A. 1974. 475 p. CAIRNS, S. A., Stony corals (Cnidaria: Hydrozoa, Scleractinia) of C arrie Bow Cay, Belize: In: Smithsonian Contribution to the Marine Sciences, K. Rützler and I. G. Macintyre (Eds.). 1982. p. 271-302. 539 p. 12: CARLQUIST, S., Chance, dispersal. Amer. Scient. 1981. 509-516. 69: COATES, A. G., Cretaceous Tethyan coral-rudist biogeography related to the evolution of the Atlantic Ocean. Palentological Association of London. Special Papers in Paleogtology, 1973. 169-174. 12: CONNELL, J. H., Population ecology of reef building corals. In: Biology and Geology of Coral Reefs. O. A. Jones and R. Endean, Bol. II, Biol. 1973. 205-246. 1: CROIZAT, L., C. NELSON y D. E. ROSEN, Centers of origin and related concepts. Syst. Zool., 1974. 265-278. 23: DANA, T. F. Development of contemporary Eastern Pacific coral reefs. Mar. Biol., 1975. 355-374. 33: DURHAM, J. W., Coelenterates, especially stony corals, from the Galápagos and Cocos Islands, In: The Galápagos. University of California Pre R. I. Bowman (Ed.) 1966. p. 123-135. ss. DURHAM, J. W. y J . L. BARNARD, Stony corals of the Eastern Pacific collected by the Velero III and Velero IV. Allan Hancock Pacific Expeditions, 1952, 1-110. 16 (1): EKMAN, S., Zoogeography of the sea. Sidgwick and Jackson, London. 1953. 417 p. EMMERSON, K.W. Mollusks with Indo-Pacific affiníties in the Eastern Pacific Ocean. The Nautilus, 1978. 91-96. 92 (2): EMILIANI, C., S. GARTNER y B. LIDZ, Neogene sedimentation on the Blake Plateau and the emergence of the Central American Isthrnus. Paleogeo., Paleoclim, Paleoecol., 1972. 1-10. 11: EVA, A.N., Paleontology, Pre-Miocene seagrass communities in the Caribbean. 1980. 231-236. 23: FISCHER, R., The formation of the Central American Isthmus in the Costa Rican Miocene. Zbl. Geol. Palaont. Teil I., H. 1981. 210-221. 3/4: FRENZEL, Climatic Fluctuations of the Ice Age. The Press of Case Western Reserve University, Cleveland, Ohio. 306p. GARTH, J. S., On the occurrence in the Eastern Tropical Pacific of Indo West Pacific Decapod crustaceans commensal with reef-building corals. Proc. 2nd. Int. Coral Reef Symp., 1974. 397-404. 1: GEISTER, J., Ocurrence of Pocillopora in late Pleistocene Caribbean coral reefs. 2nd. Symp. Int. Coraux et Recifs Coral. Fossiles. Mém. Bur. Rech. Géol. min., París. 1977. 378-388. 89: GLYNN, P. W., Coral communities and their modifications relative to past and prospective Central American Seaways. Adv. Mar. Biol., 1982. 91-132. 19: GLYNN, P. W. y I. G. MACINTYRE, Growth rate and age of coral reefs on the Pacific coast of Panamá. Proc. 3rd. Coral Reef Symp., 1977. 251-259. 2: GLYNN, P. W., R. H. STEWART y J. E. McCOSHER, Pacific coral reefs of Panamá: Structure, distribution and predators. Geol. Runds., 1972. 483-519. 61: GLYNN, P. W., E. M. DRUFFEL y R. B. DUNBAR, A dead Central American coral reef tract: Possible link with the little Ice Age. J. Mar. Res., en prensa. 1983. GOREAU, T. F., N. I. GOREAU y T. J. GOREAU, Corales y arrecifes coralinos. Inv. Cien., 1979. 48-60. 37: GOUDIE, A., Environmental Change. Claredon Press., Oxford. 1977. 244 p. HALLAM, A., Distributions patterns in contemporary terrestrial and marine animals. In: organisms and Continents Through Time. N. F. Hughes (Ed): Special Papers in Palaentology, 1973. 93-105. 12: HECK, K. L. y E. D. McCoy, Long distance dispersal and the reef building corals on the Eastern Pacific. Mar. Biol., 1978. 349-356 48: JONES, M. L. (Ed.), Bull, The Panamic Biota: Some Observations Prior to a Sea level Canal. Biol. Soc. Wash., 1972. 1-220. 2: KEIGWIN, L. D., Geology, Plioceno closing of the Isthmus of Panamá, based on biostratigraphic evidence from nearby Pacific Ocean and Caribbean Sea cores. 1978. 630-634. 6: KEIGWIN, L. D., Neogene planktonic Foraminifers from Deep Sea Drilling Project sítes 502 and 503. In: W. L. Prell, J. V. Gardner et al.: Init. Repts. DSPP, 1982a. 269-288 68: KEIGWIN, L. D., Stable isotope stratigraphy and paleoceanography of sites 502 and 503. In: W. L. Prell, J. V. Gardner et al.: Init. Repsts. DSPP, 1982b. 445-453. 68: KRUCKOW, T., Brenesia, Some aspects of the development of paleocaribbean Molluscan fauna. 1982. 353-357. 19/20: KURTEN, B., The Ice Age. G. P. Putnams Sons. New York. 1972. 179. p. LABOREL, J. West African reef corals: An hypothesis on their origin. Proc. 2nd. Int. Coral Reef Symp., 1974. 425-443. 1: LESCHINE, T. M., Oceanus, The Panamaniam sea-level canal. 1981. 20-30. 24 (2): MARSHALL, L. G., R. F. BUTLER, R. E. DRAKE, G. H. CURTIS y R. H. TEDFORD, Science, Calibration of the great American interchange: A radioisotope chronology for late Tertiary interchange of terrestrial faunas between the Americas. 1979. 272-279. 204: MARSHALL, L. G., S. D. WEBB, J. J. SEPKOSKI y D. M. RAUP, Science, Mamalian evolution and the great Amerícan interchange 1982. 1351-1357. 215: McCoy, E. D. y K. L. HECK, Biogeography of corals, seagrasses, and mangrove: An alternative to the center of origin concept. Syst. Zool., 201-210. 25: NEWELL, N. S., An outline history of tropical organic reefs. Amer. Mus. Nov., 1971. 1-37. 2465: PETTITT, J., S. DUCKER y B. KNOX, Submarine pollination. Scient. Amer., 1981. 134-143. 244 (3): PORTER, J. W., Science, Community structure of coral reefs on opposite sides of the Isthmus of Panamá. 1974. 543-545. 186: POTTS, D. D., Bull., Evolutionary disequilibrium among IndoPacific corals. Mar. Sci., 1983. 619-632. 33 (3): REVELLE, R. y B. THOMPSON, Los océanos y el clima. Cien. Desg., 1982. 28-36. 43: RICH, P. U. y T. H. RICH, The Central American Dispersal Route: Biotic History and Paleogeography: In: Costa Rican Natural History. The University of Chicago Press. D. H. Janzen (Ed.), 1983. 12-34. ROSEN, D. E., A vicariance model of Caribbean biogeography. Syst. Zool., 1975. 431-464 24: ROSENBLATT, R. H. y E. S. HOBSON, Copeia, Parrotfish (Searidea) of the Eastern Pacific, with a generic rearrangement of the Scarinae. 1969. 434-453 3: SAVAGE, J. M., Copeia, Central American Herpetofauna. 1966. 719-766 4: SIMPSON, G. G., Evolución y Geografía. Editorial Universitaria de Buenos Aires. 1964. 87 p. SMITH F. G. W., Atlantic Reef Corals. University of Miami Press. Coral Gables, Florida. 1976. 164 p. STANLEY, S. M., Macroevolution: Patterns and Process. W. H. Freeman and Co., San Francisco. 1979. 332 p. STEHLI, F. G. y J. W. WELLS, Diversity and age patterns in herm atypic corals. Syst. Zool., 1971. 115-126 20: VALLENTINE, J. W., Plate tectonics and shallow marine diversity and endemism, an actualistic model. Syst. Zool, 1971. 253-264 20: WELLS, J. W., Seleractinia, In: Treatise on Invertebrate Paleontology, R. C. Moore (Ed.), Univ. Kansas Press. and Geol. Soc. Amer. 1956. p. F328-F444. Vol. F. WELLS, J. W. Bull, New and old Scleractinians corals from Jamaica. Mar. Sci., 1973. 16-59 23: WHITMORE, F. C. y R. H. STEWART, Science, Miocene mammals and Central American seaways. 1965. 180-185 148: WINDLEY, B. F., Chichester. The Evolving Continents. john Wiley and Sons, 1978. 385 p. WOODRING, W. P., Bull., Caribbean land and sea through the ages. Geol. Soc. Amer., 1954. 719-732 65: WOODRING, W. P., The Panama land bridge as a sea barrier. Proc. Amer. Phil. Soc., 1966. 425-433 10: WYRTKI, K., Science., Telecormections in the Equatorial Pacific Ocean. 1973. 66-68 180:
|