|
UN MÉTODO PARA ESTIMAR
FLUJOS DE NITRÓGENO RECICLADO EN LAGUNAS COSTERAS SOMERAS NOTA
CIENTÍFICA
Trabajo recibido el 17 de octubre de 1983 y aceptado
para su publicación el 15 de noviembre de
1983.
ROBERTO LUIS ESCALONA
Instituto de Ciencias del Mar y
Limnología, Universidad Nacional Autónoma de México. Apdo.
Postal 70-305, 04510 México, D. F. México. Contribución
367 del Instituto de Ciencias del Mar y Limnología,
UNAM.
Se
investigó una técnica para estimar los principales flujos de
nitrógeno inorgánico y su impacto sobre la productividad primaria
en lagunas costeras someras. La técnica se basa en el uso de estructuras
plásticas que aislan una columna de agua de 1 m² de
sección transversal, permitiendo en algunos casos contacto entre el agua
y los sedimentos, y evitándolo en otros por medio de un fondo de
plástico. Los flujos de nitrógeno reciclado se estimaron a partir
de los cambios observados en las concentraciones de ión amonio y
también de datos de producción primaria. En pruebas efectuadas en
el estero "El Verde", Sin., se calculé que los peces, jaibas y otros
macro-organismos pueden contribuir hasta con el 53% del flujo de
nitrógeno reciclado a la columna de agua, los sedimentos con aprox. 31%
y los microorganismos suspendidos con aprox. 16%. La tasa promedio de
liberación de amonio de los sedimentos se estimó en 6.9 mg at N.
m-² day-¹, cigra que es comparable con la obtención en
otros ambientes costeros en el verano. Esta liberación podría
proporcionar potencialmente más de la tercera parte del nitrógeno
requerido para mantener la producción primaria
observada.
A
technique for estimating the main inorganic nitrogen flows and their impact on
primary productivity in shallow coastal lagoons was investigated. The technique
makes use of plastic enclosures that isolated a water column of 1 m²
surface area, allowing in some cases contact between the sediments and the
overlying water, and preventing it in others by means of a plastic bottom.
Recycled nitrogen fluyes were estimated from the observed changes in ammonium
ion concentrations and also from primary production data. in tests carried out
in "El Verde" estero, Sin., México, it was calculated that fish, crabs
and other macro-organisms can contribute with up to 53% of the recycled
nitrogen flow to the water column, sediments with ca. 31% and suspended
microorganisms up to 16%. The mean release rate of ammonium. from the sediments
was estimated at 6.9 mg at N. m-². day-¹, and is comparable to
that measured in other coastal environments in summer. This release could
potentially provide more than one third of the nitrogen required to maintain
the observed primary
productivity.
Una porción significativa de los litorales del mundo está ocupada por cuerpos de agua que caben dentro de una definición amplia de laguna costera. En áreas tropicales es poco probable que la productividad primaria de estos cuerpos de agua, que típicamente es elevada, esté limitada ya sea por la temperatura o por la disponibilidad de luz. Estudios químicos realizados en diversas lagunas costeras (Mee, 1978; Ryther y Dunstan, 1971; Gilmartin y Revelante, 1978) revelan consistentemente razones nitrógeno: fósforo bajas en la fase in orgánica disuelta, indicando que el nitrógeno es el nutriente limitante. Muchas de las lagunas costeras mexicanas se encuentran en regiones de clima árido o semi-árido, siendo poco común el que una laguna reciba un aporte constante de nutrientes de origen continental suficiente para mantener los altos niveles de producción primaria observados (hasta 600 mg C m-³. día-¹, Mee. 1977). Por tanto, una fracción importante del nitrógeno asimilado por el fitoplancton debe provenir del reciclaje de este elemento dentro de la misma laguna. Este reciclaje puede llevarse a cabo en la columna de agua, por oxidación bacterial del nitrógeno orgánico disuelto o por excreción animal directa, o bien en los sedimentos, por la acción bacterial sobre los detritos orgánicos. Desde hace tiempo se ha reconocido la importancia de la liberación de nutrientes regenerados de los sedimentos hacia el agua suprayacente en regiones someras, y si bien se han reportado valores para las tasas de liberación en algunos ambientes costeros, no se dispone de información comparable para las lagunas costeras. Okuda (1960) midió flujos de fósforo y nitrógeno entre núcleos de sedimento de la Bahía de Matsushima (Japón) y el agua suprayacente. Varios investigadores (Hale, 1975; Fisher et al., 1982; Boynton et al., 1981; Davies, 1975; Rowe et al., 1975 y 1977; Hartwig, 1976) han realizado este tipo de mediciones in situ, en estuarios, bahías y en la plataforma continental, utilizando cámaras cerradas de diversas formas colocadas sobre los sedimentos. El volumen de tales cámaras siempre ha sido pequeño (entre 7.8 y 62 litros), de manera que la relación entre el suministro de nutrientes y proveniente de los sedimentos y la producción primaria de las aguas suprayacentes necesariamente ha sido establecido en base a cálculos independientes. Smetacek et al. (1976), han tratado de aislar una columna de agua completa, incluyendo los sedimentos, para medir flujos de nutrientes. Desafortunadamente, se detectaron instrusiones de agua a través del fondo en el curso de estos experimentos. El objetivo del presente trabajo fue el desarrollo de un método para evaluar la importancia relativa de la contribución de los sedimentos a la productividad en lagunas costeras someras. Estos estudios forman parte de un proyecto cuya finalidad es el desarrollo de un modelo general de reciclaje del nitrógeno en lagunas costeras tropicales. ÁREA DE ESTUDIOPor su cercanía al puerto de Mazatlán y por su reducida influencia humana, se escogió al estero "El Verde" como sitio de prueba de la técnica desarrollada en este trabajo. Este estero está situado en la costa de Sinaloa aproximadamente 30 Km al norte de la ciudad de Mazatlán, a los 23'25' de latitud norte y 106°34' de longitud aproximada de 7 Km, una anchura promedio de El estero yace paralelo a la costa, teniendo una longitud aproximada de 7 km, una anchura promedio de 70 m y una profundidad media de 1.7 m. Durante la mayor parte del año, una barra arenosa de aproximadamente 200 m de anchura separa al estero del mar abierto, pero en los meses de mayor precipitación pluvial (agosto y septiembre) el río, que sólo lleva un caudal apreciable en esta época, abre una boca temporal en la barra. La salinidad en el estero varia normalmente entre 5 y 28%o a lo largo del año. Las orillas están ocupadas casi totalmente por manglar. La actividad pesquera es moderada, capturándose principalmente camarón, lisa, pargo y bagre. La hidroquimica general de esta laguna ha sido descrita por Galindo (1981). 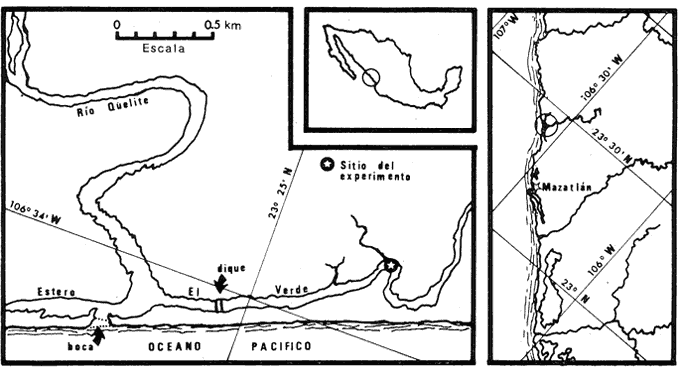 Fig. 1 .Área de estudio. Para la instalación de las estructuras plásticas desarrolladas en este trabajo se escogió un sitio fácilmente accesible a una distancia de 1.3 km de la boca del río, pero separado de ella a partir de julio de 1981 por un dique de arena que ha restringido la circulación considerablemente. La altura de la columna de agua (1.5 m) y el tipo de sedimento (limo fino de color negro) son representativos de la laguna en general. MATERIAL Y MÉTODOS
ESTRUCTURAS PLÁSTICASPara aislar una columna de agua completa dentro de la laguna, se construyeron unas estructuras de vinilo laminado flexible con grosor de 0.8 mm. Este material es transparente y puede sellarse por medio de calor. Se utilizaron dos diferentes diseños: las estructuras de fondo abierto (Fig. 2) permiten el contacto entre los sedimentos y el agua. El borde inferior consiste en un marco rígido de acrílico que penetra casi completamente en el sedimento. Las estructuras de fondo cerrado son de hecho bolsas de plástico de fondo cuadrado. Ambos tipos de estructura son sostenidos por armazones de madera, que permiten además la instalación de las estructuras con un mínimo de perturbación de los sedimentos o de la estratificación de la columna de agua, si la hay. En el caso de las estructuras de fondo cerrado, la instalación se realizó plegando totalmente las paredes de la bolsa y sujetándolas temporalmente con cinta adhesiva. La bolsa plegada, que es relativamente plana, se sumergió (sostenida por su armazón) casi hasta el fondo de la laguna. A continuación la estructura se desplazó lateralmente varios metros. Las paredes plegadas se izaron rápidamente por medio de cables al mismo tiempo que el armazón de madera era hundido en el lodo. La instalación de ambos tipos de estructuras usando los armazones de madera causa. muy poca o nula perturbación de los sedimentos, lo cual quedó comprobado al no observarse incrementos anómalos en la concentración de ion amonio en el interior de las estructuras al término de la instalación. 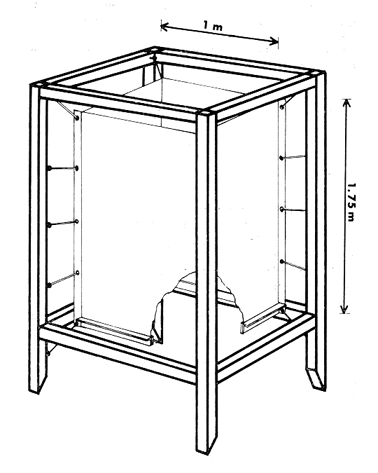 Fig. 2. Estructura de plástico de fondo abierto cortada para mostrar las características del borde inferior. La hermeticidad de las estructuras se comprobó de tres maneras: a) disolviendo Rodamina B en el interior de unas estructuras de prueba y verificando por análisis fluorométrico de las aguas exteriores la ausencia de fugas; b)elevando artificialmente la salinidad interior hasta un cierto valor y midiendo este parámetro durante varios días para detectar cualquier decremento debido a la entrada de agua a las estructuras; y c) supervisando la forma de todas las estructural, tanto de prueba como experimentales. La forma debe responder claramente a toda variación en el nivel de agua de la laguna (± 0.1 m durante los experimentos). Los tres procedimientos demostraron la completa hermeticidad de todas las estructuras. MUESTREOSe llevaron a cabo dos experimentos con duración de 10 días cada uno y por duplicado en cada ocasión: el primero del 14 al 23 de mayo de 1981, al final de la época de secas, y el segundo del 10 al 19 de septiembre del mismo año, en la época de lluvias. Se tomaron muestras a dos profundidades (0.1 y 1.3 m) al amanecer (5:00-6:00 horas) y al atardecer (18:00-19:00 horas) de cada día. Se desarrolló un sistema de' muestreo para tomar agua a una profundidad precisa, capaz de tomar muestras para análisis de oxígeno directamente con una botella B.O.D. sin diferenciales de presión y sin contacto con la atmósfera. ANÁLISISTodos los análisis se llevaron a cabo, salvo pequeñas modificaciones, según los procedimientos recomendados por Strickland y Parsons (1972). Las muestras para análisis de oxígeno se "fijaron" inmediatamente con los reactivos apropiados y se analizaron antes de 3 horas por el método Winkler. Las muestras para análisis de nutrientes se filtraron inmediatamente a través de filtros de fibra de vidrio Whatman GF/C y se congelaron. Las muestras acumuladas se analizaron cada dos o tres días. Para el amonio el procedimiento recomendado (Solórzano, 1969) dio resultados poco satisfactorios, pero una reducción en los reactivos usados (a la décima parte de las cantidades recomendadas) permitió obtener blancos bajos y reproducibles. Las muestras para análisis de pigmentos fotosintéticos se colocaron en botellas de plástico de 500 m1 y se filtraron de 1 a 2 horas después del muestreo. Los filtros se cubrieron de acetona al 90%, completándose el análisis 24 horas más tarde. Las mediciones de producción primaria se realizaron incubando botellas B.O.D. de 300 ml claras y obscuras durante tres horas (de las 10.00 a las 13.00 horas) a las profundidades de 0.1 y 1.3 m. La fotosíntesis bruta (P(z, t), en mgC, m-³. hr-¹) se calculó en base a los cambios en las concentraciones de oxígeno disuelto utilizando un cociente fotosintético de 1.2 (Williamset al., 1979). De cada pareja de mediciones se derivó el valor de la fotosíntesis máxima que se lleva a cabo bajo esas condiciones (Popt), asi como la intensidad luminosa a la que se lleva a cabo (Ip) . Este último valor está dado como una fracción de la luz que penetra la superficie del agua al mediodía local aparente (Ip.). Los cálculos se hicieron utilizando la ecuación de Vollenweider (1965a) en una forma que ha sido aplicada por Mee (comunicación personal) a ambientes similares, donde demostró describir razonablemente los datos experimentales: 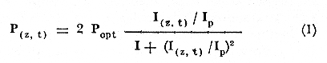 I (z, t), la intensidad de luz a la profundidad z y al tiempo t puede calcularse con la ecuación de Ikushima (1967): 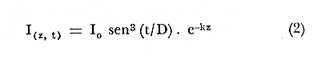 donde D es el total de horas de luz en el día, t es el punto medio del periodo de incubación (con t = 0 al amanecer) y k es el coeficiente de extinción del medio, aproximado aquí por 1.44 dividido entre la profundidad del disco Secchii (Holmes, 1970). Asi, la forma integrada de la ecuación (1) 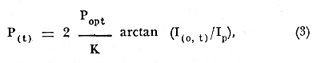 Puede usarse para estimar la producción bruta bajo un metro cuadrado de superficie del agua. Es obvio, sin embargo, que en el caso de estos experimentos el fondo (a una profundidad de 1.5 m) queda dentro de la zona eufótica, de manera que es necesario calcular P(t) para I(o, t)y para I(1.5 t) y sacar la diferencia. No es necesario considerar la producción por algas sésiles en los sedimentos, ya que éstos son demasiado blandos y/o anóxicos para permitir el crecimiento de tales organismos. Los valores de P(t) obtenidos se extrapolaron a un día completo multiplicándose por 9. Esta cifra se justifica en base a la consideración de Vollenweider (1965b) de que durante el segundo quinto del periodo de luz de día se lleva a cabo aproximadamente el 30% de la fotosíntesis diaria total. RESULTADOS Y DISCUSIÓN
CAMBIOS OBSERVADOS DENTRO DE LAS ESTRUCTURASUna característica notable de los resultados obtenidos es la poca diferencia que se observa entre las mediciones a 0.1 m y 1.3 m de profundidad para la mayoría de los parámetros, y también entre los experimentos de mayo y septiembre. En los casos en que dicha diferencia no era evidente, se calculó el promedio de las diferencias individuales y se consideró que existía una diferencia significativa cuando dicho promedio era mayor del 5% del promedio de las mediciones. Este análisis revelé diferencias significativas entre los valores a ambas profundidades solamente para el amonio en las estructuras de fondo abierto y para el oxígeno disuelto. Se encontraron diferencias estacionales sólo en el caso de la salinidad, la temperatura y el ortofosfato, mientras que para los nitratos y nitritos afuera de las estructuras se pudieron establecer diferencias tanto estacionales como de profundidad. Por lo tanto, excepto cuando se indica lo contrario, las curvas de todas las figuras han sido dibujadas promediando los valores a ambas profundidades y de ambos experimentos. Al avanzar los experimentos, la salinidad y la temperatura mostraron únicamente pequeñas variaciones, indicando que en estos periodos no se presentaron cambios hidrodinámicos importantes en esa parte del estero. En mayo, la salinidad varió entre 20.9 y 21.1 ‰ tanto en el exterior como en el interior de las estructuras, mientras que la temperatura fue de 27.3 ± 1.1ºC al amanecer y 30.4 ± l °C al atardecer. En septiembre la salinidad, debido a lluvias ocasionales, varió en un rango más amplio, de 16.3 a 17.1 ‰ afuera de las estructuras y de 16.5 a 16.9 en el interior de las estructuras. La temperatura promedió 29.6 ± 0.7ºC al amanecer y 33.0 ± 1.2 ºC al atardecer. Aunque Hale (1975) detectó una correlación entre la liberación de amonio de los sedimentos y la temperatura, las variaciones en estos rangos no son al parecer un factor significativo en lagunas costeras tropicales. 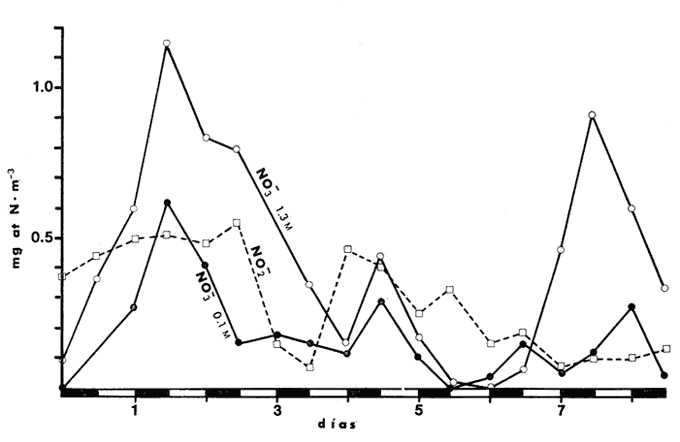 Fig. 3. Concentraciones de nitratos y nitritos en el estero "El Verde, afuera de las estructuras durante el experimento de mayo, 1981. El tiempo es cero al atardecer del 14 de mayo. Las concentraciones de nitratos y nitritos en el interior de las estructuras durante el experimento de mayo, y tanto dentro como fuera de las estructuras durante el experimento de septiembre, fluctuaron entre no detectables y 0.1 mg at N m-³. Solamente en mayo, y únicamente afuera de las estructuras, se detectaron niveles más altos, especialmente a 1.3 m durante la primera parte del experimento (Fig. 3). Ya que este fenómeno no se observó en septiembre, es poco probable que estos nutrientes tuvieran un origen externo. La razón nitritos: nitratos es relativamente alta, lo que sugiere algún proceso bacterial (probablemente nitrificación) que por alguna razón aún no determinada no se lleva a cabo dentro de las estructuras (ni siquiera en las de fondo abierto) , y no siempre en la laguna misma. Adicionalmente, se esperaban concentraciones altas de nitratos durante el experimento de septiembre, pero al parecer el dique de arena construido en julio impidió que entraran cantidades apreciables de agua de río a la parte sur del estero. Las concentraciones de ortofosfato presentaron variaciones hasta de 0.4 mg at P m-³ de un día a otro, pero no fue posible distinguir alguna tendencia definida en el interior de las estructuras, donde las concentraciones promedio fueron de 4.01 ± 0.35 mg at P m-³ en mayo y 16.61 ± 0.29 mg at P m-³ en septiembre. Fuera de las estructuras si parece haber una ligera tendencia ascendente a lo largo de los periodos de muestreo, como puede verse en la Tabla 1. Esto pudiera ser parte de un ciclo general de movilización de fósforo en la laguna y sus alrededores a lo largo de un año. Los tres parámetros que muestran el comportamiento más interesante son las concentraciones de amonio, clorofila a (Figs. 4 y 5) y oxígeno disuelto. En el exterior de las estructuras, los tres parámetros presentaron fluctuaciones cíclicas considerables, con máximos de oxígeno y clorofila a y mínimos de amonio al atardecer, y mínimos de oxígeno y clorofila a y máximos de amonio al amanecer. Sin embargo, no parece existir una tendencia definida a largo plazo, como puede verse por los análisis de regresión lineal separados de los datos del atardecer y del amanecer (Tabla 1) que dan coeficientes de correlación extremadamente bajos. Por tanto, parece razonable el considerar que durante los experimentos el estero permaneció en un estado de "estabilidad dinámica": durante la noche el amonio se acumula en el agua, pues las tasas de asimilación por el fitoplancton son muy reducidas o nulas. De día su concentración decrece considerablemente al ser asimilado por los productores primarios, y es posible que hacia el final del día se establezca un equilibrio entre las tasas de asimilación y de suministro a la columna de agua. Las tasas fotosintéticas medidas son otras, ocasionando un incremento neto de la biomasa fitoplanctónica (representada por la concentración de clorofila a)., mientras que de noche hay una reducción neta por pastoreo, hundimiento, etc. Aparentemente, un equilibrio entre ambos procesos mantiene a la población fitoplanctónica del estero en un nivel relativamente constante a lo largo de varios días. 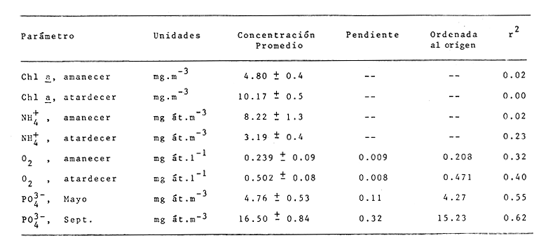 TABLA 1 ANÁLISIS DE REGRESIÓN LINEAL (CONCENTRACION VS TIEMPO) DE CLOROFILA a, AMONIO, OXÍGENO DISUELTO Y ORTOFOSFATO EN EL ESTERO "ELVERDE" FUERA DE LAS ESTRUCTURAS EN MAYO Y SEPTIEMBRE DE 1981. EL TIEMPO ES IGUAL A CERO DIAS AL ATARDECER DEL 14 DE MAYO Y AL ATARDECER DEL 10 DE SEPTIEMBRE. 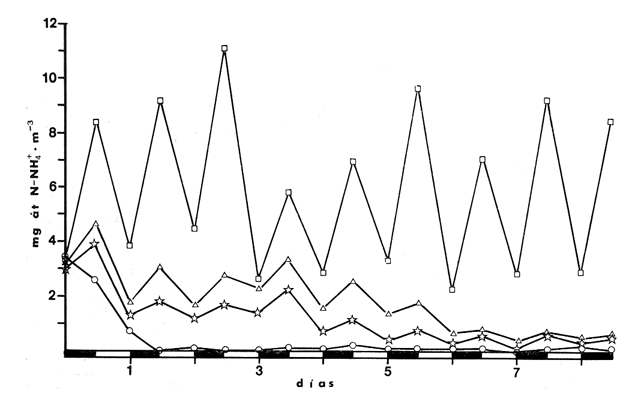 Fig. 4. Variaciones en la concentración de ión amonio durante los experimentos con estructuras de plástico en el estero El 'Verde. Los triángulos y las estrellas corresponden a los dat os de las estructuras de fondo abierto, a una profundidad de 1.3 y 0.1 metros respectivamente. Los círculos representan datos de las estructuras de fondo cerrada- y los cuadrados datos de fuera de las estructuras. El tiempo es 0 al atardecer del 14 de mayo y del 10 de septiembre de 1981. 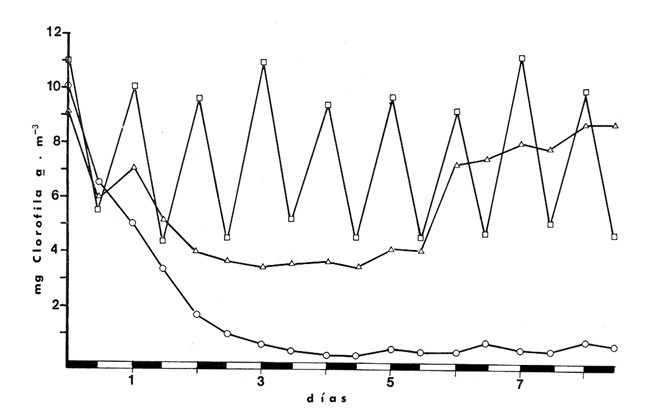 Fig.5.Variacionesenlaconcentracióndeclorofila a durantelosexperimentosconestructurasdeplásticoenelestero"ElVerde".Lostriángulosycírculoscorrespondenadatosdelasestructurasdefondoabiertoy cerradorespectivamente,yloscuadradosadatosdefueradelasestructurasEltiempoes0alatardecerdel14demayoydel10deseptiembrede1981 En las estructuras de fondo abierto, la desaparición gradual del amonio parece indicar que se ha eliminado una fuente importante de este nutriente. El efecto es más pronunciado cerca de la superficie del agua, lo cual es razonable si la mayor parte del suministro de amonio a la columna de agua proviene ahora de los sedimentos. Las fuentes posibles del amonio faltante son: a) Escurrimientos continentales. b) Advección desde otras partes del estero. c) Macro-organismos tales como peces, camarones, can grejos, aves, manglares, etc., ya sea por excreción directa o por perturbación de los sedimentos. Los niveles tan bajos de nitratos detectados, y la similaridad de los resultados obtenidos en ambos experimentos sugieren que durante estas pruebas los dos primeros factores no tuvieron un papel importante. Actualmente se está llevando a cabo un estudio en esta misma laguna, con objeto de estimar las tasas de excreción de amonio de las lisas (Mugil cephalus). Los resultados preliminares indican que dichas tasas pueden proporcionar fácilmente todo el amonio faltante de las estructuras de fondo abierto. El comportamiento de las concentraciones de clorofila a (Fig. 5) no puede explicarse de manera tan sencilla. Durante los primeros días, los procesos de reducción de la biomasa (pastoreo, hundimiento, etc.) parece predominar sobre la producción. Más adelante, estos procesos aparentemente se detienen, y la concentración de clorofilaapermanece relativamente constante (3.8 ± 0.2 mg. m-³ entre el tercer y sexto días (esta cifra es para las mediciones de mayo a 0.1 m, pero el comportamiento es esencialmente el mismo a 1.3 m y en septiembre). Sin embargo, las fluctuaciones en el oxígeno disuelto indican que se está llevando a cabo una cierta cantidad de fotosíntesis. A partir de este momento, hay un incremento neto en las concentraciones de clorofila a durante el día, sin la disminución nocturna correspondiente. Es probable que esta tendencia no se prolongue por mucho tiempo, pero no había desaparecido aun en el momento de terminarse los experimentos. En las estructuras de fondo cerrado la producción primaria cesa casi por completo en unos cuantos días, sugiriendo que la contribución de amonio por la excreción del zooplancton y por la acción bacterial sobre el nitrógeno orgánico suspendido o disuelto es relativamente insignificante. Hacia el final de los experimentos hay una ligera recuperación de la biomasa fitoplanctónica, lo cual posiblemente se deba a la remineralización del nitrógeno orgánico particulado por las bacterias, proceso que es relativamente lento. Los niveles de oxígeno disuelto, con su patrón cíclico muy atenuado, muestran una reducción del 50% relativa a los máximos del primer día, mientras que a partir del tercer día sólo pueden detectarse cantidades traza de amonio (Fig. 4) . ESTIMACIÓN DE FLUJOS DE NITRÓGENOEn este tipo de experimentos, la mejor forma de estimar flujos de nitrógeno es a partir de los incrementos nocturnos en la concentración de amonio ( Δ NH4+)N , ya que el error introducido por la necesidad de adoptar valores de las tasas de asimilación de amonio es mucho menor que durante el día. Por tanto el flujo total de amonio a la columna de agua (F, en mg at N m-³ hr-¹) estará dado por: 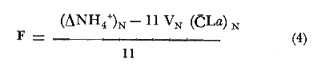 donde (CLa)N es la concentración promedio de clorofila a durante la noche, 11 son las horas de oscuridad y VN es la velocidad de asimilación de amonio en la oscuridad, con un valor promedio de 0.025 mg at N mg Cla-¹ hr-¹ (D, S. = 0.004). Esta cifra es el resultado de experimentos efectuados en el estero "El Verde" durante 1981 y 1982 (Escalona, datos no publicados), durante los cuales se agregaban diferentes cantidades de nutrientes (nitrato y amonio) a estructuras de fondo cerrado. A continuación, se midió la concentración de nutrientes y clorofila a a intervalos cortos (30 min. de noche, 10 min. de día) durante dos días. VN es prácticamente independiente, dentro de un rango razonable, de la concentración de amonio en el medio, por lo que se usó el valor promedio tanto dentro como fuera de las estructuras. Los experimentos mencionados produjeron también valores para las tasas de asimilación de amonio durante el día, pero éstos varían dentro de un rango tan amplio (dependiendo de la biomasa inicial, historia de disponibilidad de nutrientes, condiciones de iluminación, etc.) que sería imposible utilizarlos para estimar flujos de amonio a partir de los cambios en la concentración de amonio durante el día. Por ejemplo, a una concentración de amonio de 3 mg at. m-³ se obtuvieron valores de VD en el rango 0.03-0.90 mg at N mg Cl a-¹ hr-¹. Los flujos de amonio calculados usando la ecuación (4) se muestran en la Tabla (2). Se han combinado los resultados de mayo a septiembre, tanto a 0.1 m como a 1.3 m, de manera que cada valor de F es el promedio de 8 mediciones. La excepción son las estructuras de fondo abierto, donde se tomaron en cuenta únicamente las mediciones hechas a 1.3 m. En esta tabla puede verse a primera vista que mientras que fuera de las estructuras los flujos de amonio variaron muy poco a lo largo de los experimentos, en el interior de las estructuras hay un decremento notable después del primer día. Esto probablemente esté relacionado con el comportamiento inesperado de las concentraciones dentro de las estructuras (especialmente las de fondo abierto) mencionado anteriormente. Si es verdad que la actividad del zooplancton se reduce considerablemente después del segundo día (cuando la curva de la clorofila a se vuelve horizontal) también se esperaría una reducción en su excreción de nitrógeno. De hecho, la diferencia entre el flujo de amonio calculado para el primer día en las estructuras de fondo abierto (0.32 ‰ 0.03 mg at N m-³ hr-¹) y el flujo promedio de amonio desde el tercero hasta el noveno día (0.20 ± 0.02) es 0.12 mg at N m-³ hr-¹. Esta cifra no es muy diferente del flujo de amonio calculado para el primer día en las estructuras de fondo cerrado (0.11 ± 0.02), donde es de suponerse que la excreción de zooplancton es la principal fuente de amonio. 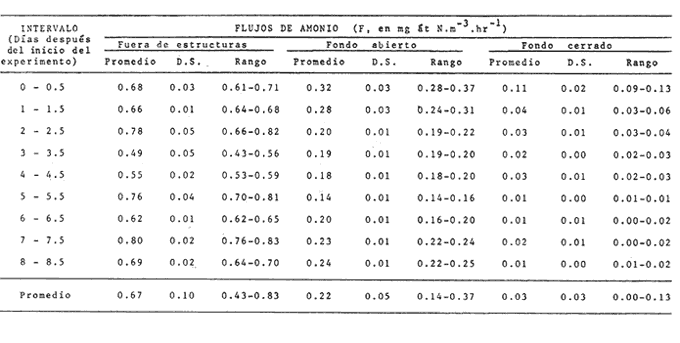 TABLA 2 FLUJOS DE AMONIO A LA COLUMNA DE AGUA ESTIMADOS EN LOS EXPERIMENTOS CON ESTRUCTURAS DE PLÁSTICO EN EL ESTERO "ELVERDE" En consecuencia, quizá sería más correcto el estimar la contribución relativa de cada fuente al flujo de nitrógeno utilizando únicamente los valores promedio obtenidos para el primer día de los experimentos. Dichas estimaciones se presentan en la Tabla 3, donde "contribución externa" se refiere a la "contribución de macro-organismos" (por excreción o bioturbación) así como el flujo advectivo de nitrógeno de otras partes de la laguna. En el presente caso es probable que este último haya sido relativamente insignificante. Para verificar la magnitud de los valores de F obtenidos, se estimaron éstos en forma independiente a partir de los datos de producción primaria fuera de las estructuras (Tabla 4). El flujo de nitrógeno (llamado aquí F') será así el nitrógeno necesario para mantener la producción observada, ya que se considera que el estero permanece estable a mediano plazo. Por tanto, la fotosíntesis diaria total (P, en mgC. m-². día-¹) se dividirá entre 24 horas, la altura de la columna de agua (1 .5 m) , el peso atómico del carbono y una razón atómica de asimilación carbono: nitrógeno de 3 para dar F. Este valor bajo de la razón de asimilación C:N se escogió en vista de la alta productividad observada y la relativa abundancia de amonio en el medio. Los valores de F' asi calculados se presentan en la Tabla 4 junto con los promedios de los valores de F para la noche anterior y la siguiente. Aunque los valores de F' resultan un tanto bajos, quedan todos dentro del margen de error esperado. 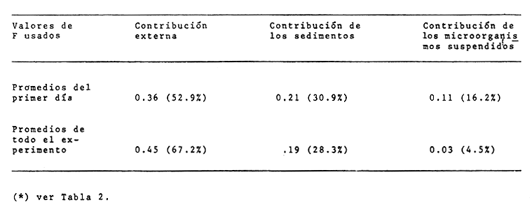 TABLA 3 CONTRIBUCIÓN DE LAS DIFERENTES FUENTES DE NITRÓGENO RECICLADO A LAS AGUAS DEL ESTERO "ELVERDE (EN MG AT N-NH4+M-3hr-1, Y COMO PORCENTAJE DEL TOTAL ENTRE PARÉNTESIS) 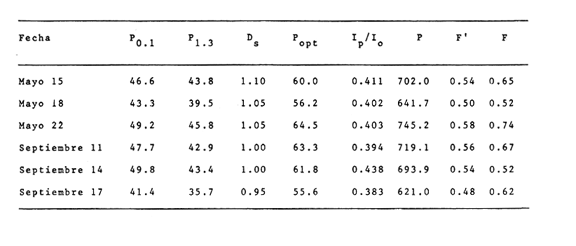 TABLA 4 DATOS DE PRODUCCIÓN PRIMARIA DE LESTERO ELVERDE EN MAYO Y SEPTIEMBRE,1981.LASTASAS FOTOSINTÉTICAS (Popt,P0.1YP1.3)ESTÁNDADASENMGC.M-3.HR-1LAFOTOSINTESISTOTAL(P)ENMGC.M-2.DÍA1;LAPROFUNDIDADDELDISCOSECCHII(D)ENMETROS;LOSFLUJOSDENITRÓGENO(FYF')ENMGT NM-3HR-1 LA CONTRIBUCIÓN DE LOS SEDIMENTOSEl objetivo principal de este tipo de experimentos es evaluar la importancia relativa de la contribución de los sedimentos no perturbados a la productividad de una laguna costera. Dicha contribución se estimará aquí a partir de la diferencia en los flujos de amonio calculados para las estructuras de fondo abierto y de fondo cerrado, que tiene un valor promedio de 0.19 mg at N m-³ hr-¹ (D.S. = 0.04). Esto corresponde a una tasa de liberación de amonio de 6.9 (D.S. = 1.4) mg at N día-¹ por metro cuadrado de superficie de los sedimentos. En la Tabla 5 estos resultados se comparan con los estimados por otros investigadores para diversos ambientes costeros. Puede verse que los valores Obtenidos en este trabajo son, aunque altos, del mismo orden que los otros. Además, la mayoría de los valores promedio presentados han sido calculados con datos de todo un ciclo anual en zonas templadas, y los valores más altos corresponden a mediciones hechas en verano. En varios casos se ha establecido una relación directa entre la temperatura del agua y la liberación de amonio (p. ej. Hale, 1975), por lo que no es de sorprender que en "El Verde", donde la temperatura media es por lo menos tan alta como las temperaturas de verano en las otras localidades las tasas de liberación de amonio a 30ºC (que es la temperatura media de "El Verde" en primavera y verano), se obtiene un rango de 4.22 a 7.66 mg at N m-³ día-¹, que incluye el valor promedio de 6.9 estimado en este trabajo. En cuanto a la importancia relativa de esta contribución a la producción primaria de las aguas suprayacentes, en la Tabla 5 puede verse que el porcentaje promedio de los requerimientos diarios de nitrógeno que puede ser cubierto por el amonio liberado de los sedimentos varía ampliamente, desde 5,8% (Hartwig, 1976) hasta 96% (Rowe et al.,1975). Por lo tanto, la estimación hecha aquí representa un valor intermedio. Esta contribución es aún más importante ya que probablemente no varía mucho a lo largo del año, mientras que en las zonas templadas en invierno el flujo de amonio de los sedimentos al agua puede llegar a invertirse. 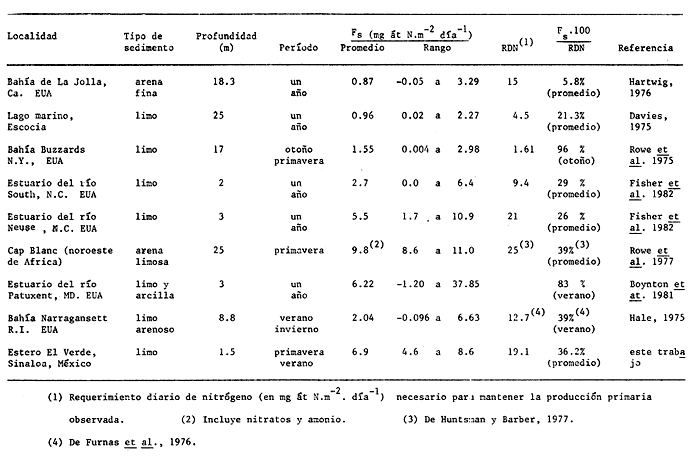 TABLA 5 TASAS DE LIBERACIÓN DE AMONIO DE LOS SEDIMENTOS (F) EN DIVERSOS AMBIENTES COSTEROS Y SU CONTRIBUCIÓN POTENCIAL A LA PRODUCTIVIDAD PRIMARIA ConclusionesLa técnica de estructuras de plástico presentadas aquí promete ser la utilidad para evaluar flujos de nitrógeno y su efecto sobre la productividad primaria de lagunas costeras bajo condiciones no muy diferentes de las naturales. Sin embargo, es evidente la conveniencia de realizar estimaciones simultáneas y precisas de la razón de asimilación C:N y la velocidad de asimilación de amonio utilizando técnicas más sofisticadas, tales como la espectrometría de masas para este último parámetro. Mientras tanto, los resultados obtenidos indican que, por lo menos en lagunas costeras tropicales que reciben aportes negligibles de nutrientes "nuevos", los peces, crustáceos, aves y otros macro-organismos tienen un papel más importante de lo que generalmente se reconoce en el reciclaje del nitrógeno, mientras que la excreción del zooplancton y la actividad de bacterias suspendidas son relativamente poco importantes en este contexto. En cuanto a los sedimentos, las tasas de liberación de amonio estimadas aquí son aparentemente del mismo orden de las que prevalecen en la mayoría de los ambientes costeros de las zonas templada., en verano, y pueden contribuir potencialmente con más de la tercera parte del nitrógeno necesario para mantener la producción primaria de una laguna. AgradecimientosEl autor agradece a Gilberto Díaz y Pedro Ortega su ayuda en la instalación de las estructuras y durante los muestreos, y a Lawrence Mee sus útiles sugerencias. LITERATURABOYNTON, W. R., W M., KEMP y C. G. OSBORNE Estuarine Perspectives.Nutrient fluxes across the sediment-water interface in the turbid zone of a coastal plain estuary. In; Kennedy, J. P. (Ed.)Academic Press,Londres1980.93-110 DAVIES, J. M., Mar Biol., Energy flow through the benthos in a scottish sea loch.1975.353-362314 FISHER, T. R., P. R. CARLSON y R. T. BARBER, Sediment nutrient regeneration in three North Carolina estuaries.Estuar. Coast. Shelf Sci.,1982.101-116141 FURNAS M. J., G. L. HITCHCOCK y T. L. SMAYDA, : Estuarine Processes 1. Nutrien tphytonplank ton relationships in Narragansett Bay during the 1974 summer bloom. In: Wiley, M. (Ed.)Academic Press,New York1976.118-113 GALINDO, J. G., Tesis de Maestría en Ciencias del Mar, Univ. Nal. Autón. México, Estudio preliminar sobre la productivídad primaria y la dinámica de los nutrientes en el estero, El Verde, Mazatlán, Sin. México.1981.134 GILMARTIN M. y N. REVELANTE, Estuar. Coast. Mar. Sci., The phytoplankton characteristics of the barrier island lagoons in the Gulf of California,1978.29-4771 HALE, S. S., Mineral Cycling in Southeastern Ecosystems.The role of benthic communites in the nitrogen and phosphorus cycles of an estuary, In: Howell, F. G., J. B. Gentry y M. H. Smith (Ed.)ERDA Symposium Series 1975 (CONF-74-0513): 291308.1975,291-308 HARTWIG, E. O., : Estuarine Processes 1. The impact of nitrogen and phosphorus release from a siliceous sediment on the overlying water. In: Wiley, M. (Ed.)Academic Press,New York1976.103-117 HOLMES, R. W., Limnol. Oceanog., The Secchii disk in turbid coastal waters.1970.688-694155 HUNTSMAN S. A. y R. T. BARBER, Primary production off North West Africa: the relationship to wind and nutrient conditions.Deep Sea Res.,1977.25-33241 IKUSHIMA, I., Ecological studies on the productivity of acquatic plant conimunites. III. Effect of depth on daily photosynthesis in submergel macrophytes. Bot. Mag.Tokyo,1967.56-67801 MEE, L. D., Tesis doctoral, Universidad de Liverpool,The chemistry and hydrography of some coastal lagoons in the Pacific coast of Mexico. Reino Unido.1977. MEE, L. D., Chemical oceanography, 7, Coastal lagoons, In: Riley, J. P. y Chester, R. (Eds.) Academic Press, Londres2a. Ed.1978.441-486 OKUDA, T., Trab. Inst. Oceanogr. Univ. RecifeMetabolic circulation of phosphorus and nítrogen in Matsushima Bay (Japan) with special reference to exchange of these elements between estuarine water and sediments.(Brazil),1960.7-15321 ROWE, G. T., C. H. CLIFFORD, K. L. SMITH JR., Y P. L. HAMILTON,Nature Benthic nutrient regeneration and its coupling to primary productivity in coastal waters.1975.215-217255 ROWE, G. T., C. H. CLIFFORD y K. L. SMITH, JR., Nutrient regeneration in sediments off Cap Blanc, Spanish Sahara.Deep-Sea Res.1977.57-63241 RYTHER, J. H. y W. M. DUNSTAN, Science Nitrogen, phosphorus and eutrophication in the coastal marine environment.1971.1008-1012171 SMETACEK, V., B. VON BODUNGEN, K. VON BROECKEL y B. ZEITSCHEL, The plankton tower II. Release of nutrients from sediments due to changes in the density of bottom water. Mar. Biol.,1976.373-378344 SOLÓRZANO, L. Limnol. Oceanog., Determination of ammonia in natural waters by the phenol-hypochIorite method,1969.799-801146 STRICKLAND, J. D. H. y T. R. PARSONS, A practical handbook for seawaler analysis,Fisheries Research Board of Canada. 2a. ed.Ottawa,1972.310 VOLLENWEIDER, R. A., Calculation models of photosynthesis.In: Goldman, C. R. (Ed.)Primary productivity in aquatic environments. Mem. Inst. Ital. Idrobiol., 18 suppl., University of California Press,Berkley1965a.415-457 VOLLENWEIDER, R. A., Materiali ed idee per una idrochimica delle acque insubriche. Mem. Inst. Ital. Idrobiol.,1965b.213-286192 WILLIAMS, P. J., R. C T. RAINE y J. R. BRYAN, Oceanol. Acta. Agreement between the carbon-14 and oxygen methods of measuring phytoplankton production: reassessment of the photosynthetic quotient.1979.411-41624
|