|
ANÁLISIS DEL CRECIMIENTO DE LA ANCHOVETA
ENGRAULIS MORDAX GIRARD, EN AGUAS DE BAJA CALIFORNIA NORTE (PISCES:
ENGRAULIDAE)
Trabajo recibido el 23 de octubre de 1983 y aceptado para su
publicación el 1 de diciembre de
1983.
MANUEL GALLARDO- CABELLO
Instituto de Ciencias del Mar y Limnología,
Universidad Nacional Autónoma de México. Contribución 365
del Instituto de Ciencias del Mar y Limnología,
UNAM.
En el presente
trabajo, se determinan las constantes del crecimiento de la ecuación
de von Bertalanffy para dos poblaciones de anchovetaEngraulis mordaxde
Baja California, denominadas para los fines de este estudio, "Norte"
y "Sur". Asimismo, se proporcionan datos sobre el crecimiento ponderal,
por sexos y edad límite o longevidad. Los parámetros del
crecimiento para la población "Norte" son L∞ = 123.32 mm;
W∞ = 17.437 gr; k = 0.5167; to = -1.7933; A (
0.95) = 4 años. Los parámetros del crecimiento para
la población "Sur" son L∞ = 119.96 mm; W∞ . = 15.885
gr; k = 0.5227; to = -1.8581; A(0,95) = 3.87 años.
In the present
paper, parameters from von Bertalanffy's growth equation were determinated
for twoEngraulis mordaxpopulations located in the Baja California
coast, that for the purpose of this study has been named "North" and
"South". Also differences in growth between sexes, weight growth and
longevity were detected. Growth parameters for the northern population
were: L∞ = 123.32 mm; W∞ . = 17.437 gr; k = 0.5167; to
= -1.7933; A(0.95) = 4 years. Growth parameters for the southern population
were: L∞ = 119.96 mm; W∞ = 15.882 gr; k = 0.5227; to
= -1.8581; A(0.95)= 3.87 years. Las familias o especies de peces, presentan tasas potenciales del crecimiento determinadas genéticamente. Dentro de cada población pueden ocurrir variaciones del índice catabólíco k, asociadas con cambios en las condiciones ambientales y los ciclos de producción biológica que a su vez, traen como consecuencia cambios en la disponibilidad del alimento y en las interrelaciones de depredación y competencia intra e interespecífica. Las diferencias en el índice catabólico k, indican diferencias en la longevidad, talla de primera captura, índice de mortalidad natural y otros parámetros de dos o más poblaciones en un área determinada. Dichos cambios se traducen en variaciones de la biomasa de las distintas poblaciones, es decir, el peso ganado para cada población debido al crecimiento, y el peso de cada población, perdido por la mortalidad natural. En el presente trabajo, se han aplicado diversas técnicas para determinar las constantes del crecimiento de la ecuación de von Bertalanffy. Para ello, se han empleado los datos de edad-longitud de dos poblaciones de la anchovetaEngraulis mordax de Baja California denominadas para los fines de este estudio, "Norte" y "Sur". Los diferentes métodos empleados para la evaluación de los parámetros del crecimiento, han sido ampliamente desarrollados y comparados con los siguientes propósitos: a) Realizar un análisis minucioso de la meto dología empleada por los diversos autores, para este tipo de estudios, y b) Establecer las ecuaciones del crecimiento que mejor se, ajusten a las poblaciones "Norte" y "Sur" de Engraulis mordax. Por otra parte, se proporcionan datos sobre el crecimiento ponderal, por sexos y edad límite o longevidad, para cada una de las poblaciones en estudio. MATERIAL Y MÉTODOSSe obtuvieron los datos biológicos de 21,309 ejemplares deEngraulis mordaxfrente a las costas de Baja California Norte, desde las Islas Coronado hasta Punta Santo Tomás. Diariamente de abril a diciembre de 1982 se llevaron a cabo las muestras de la captura comercial en los buques "Calafia", "Ciprés", "Concepción" "Kino", "Libertador del Sur", "Loreto", "Sauzal", "Tortugas" y "Vizcaíno", pertenecientes a la Compañía Pesquera Zapata. Debido a que durante los meses de enero, febrero y marzo las poblaciones de la especie mencionada se dispersan y no son capturadas por la flota comercial, no se poseen registros para dichos meses. De cada captura se separó un kilogramo de peso, elegido al azar por cada barco. De cada uno de los lotes, se consignaron en las hojas de muestreo los datos siguientes: nombre del barco, área y fecha de la captura, volumen de la captura, número de la muestra y peso total de la misma. De los ejemplares obtenidos se registró: la longitud standard, el peso entero y el sexo. Por otra parte, se efectuó la extracción de 3,400 pares de otolitossagitta. El análisis de los anillos de crecimiento y la determinación del tiempo de formación de las bandas de crecimiento rápido y lento, permitió la identificación de cinco grupos de edad (Gallardo 1985). Por otra parte, en base a las tallas promedio para cada edad y a los datos merísticos -como son: longitud, anchura total, longitud del núcleo y amplitud de las bandas del crecimiento- se determinaron dos poblaciones en las costas de Baja California Norte, que para los propósitos del presente trabajo se denominaron "Norte" y "Sur", detectándose, asimismo diferencias del crecimiento entre sexos (Gallardo, 1983). Para el análisis del crecimiento, en base a los resultados obtenidos, se empleó la ecuación de Brody (1927) y von Bertalanffy (1938). La obtención de los parámetros de dicha ecuación se realizó aplicando las técnicas de Ford (1933) y Walford (1946) Gulland (1964) Tomlinson y Abramson (1961) Allen (1966) y Beverton (1954). Para la obtención de la edad límite o longevidad se empleó la ecuación de Taylor (1958, 1959, 1960, 1962). RESULTADOS Y DISCUSIÓN
EXPRESIONES MATEMÁTICAS DEL CRECIMIENTO ECUACIÓN DE BRODY VON BERTALANFFY
Método de Ford-Walford Empleando los datos obtenidos a partir de la lectura de Otolitos, se tienen las siguientes longitudes de 1t y 1t+1 para cada una de las poblaciones en estudio: 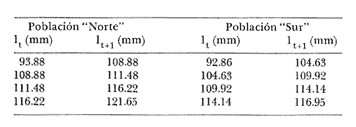 Las rectas de regresión encontradas para todos los puntos por el método de mínimos cuadrados son los siguientes: 1t+1= 0.5069 1t + 60 para la población "Norte"; 1t+1= 0.5769 1t + 50.61 para la población "Sur". Los valores de los coeficientes k y L∞ y los encontrados por este método, aparecen a continuación: k = 0.6794 L∞ = 121.68 mm, para la población "Norte" y; k = 0.55 L∞ = 119.62 mm, para la población "Sur", respectivamente. No se empleó el método gráfico para la obtención de los valores de L8, por considerar dicho método muy sensible a las variaciones de k pudiéndose obtener de esta forma resultados erróneos. Por ello, resultó más práctico obtener los valores de este parámetro, aplicando las ecuaciones de regresión correspondientes. Método de Gulland Para el cálculo de los valores de to se empleó el método de Gulland (1964), obteniéndose los resultados siguientes: to = -2.1867 para la población "Norte"; y to= -1.71692 para la población "Sur" Método de Tomlinson y Abramson Los valores de Z obtenidos a partir de las tablas de polinomiales para J 5j (z) son los siguientes: Z = 0.5965 para la población "Norte" y Z = 0.5929 para la población "Sur", respectivamente. En las tablas 1 y 2 se muestra el método empleado para cada caso.  TABLA 1 CÁLCULO DE 5/Sigma;/j = 1 lj J 5j (z )DEL MÉTODO DE TOMLINSON Y ABRAMSON (POBLACIÓN NORTE) 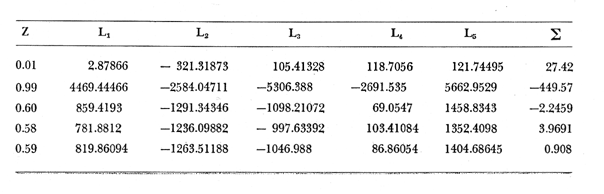 TABLA 2 CÁLCULO DE 5/Sigma j=1lj 5j (z) DEL MÉTODO DE TOMLINSON Y ABRAMSON (POBLACIÓN SUR) Aplicando y = 1-q loge z-1 se tienen los valores de k: 0.5167 para la población "Norte" y 0.5227 para la población "Sur". Aplicando las ecuaciones: 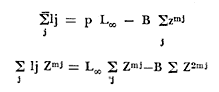 se obtienen los valores de ΣZmj, ΣZ2mj y Σ1j Σ1j ΣZmj, que aparecen en las tablas 3 y 4 para cada una de las poblaciones en estudio. 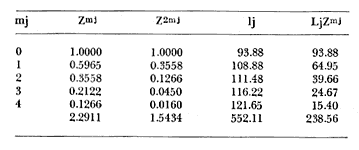 TABLA 3 RESOLUCIÓN DEL SISTEMA DE ECUACIONES DEL MÉTODO DE TOMLINSON Y ABRAMSON (POBLACIÓN NORTE) 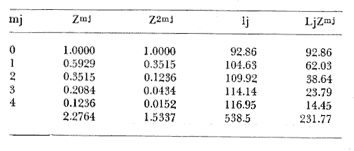 TABLA 4 RESOLUCIÓN DEL SISTEMA DE ECUACIONES DEL MÉTODO DE TOMLINSON Y ABRAMSON (POBLACIÓN "SUR") Empleando las siguientes ecuaciones: 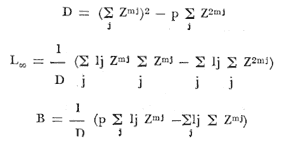 se obtienen los valores de L∞ = 123.82 mm. para la población "Norte" y; L∞= 119.96 mm para la población "Sur". Aplicando: 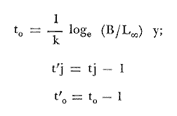 se obtienen to = –1.7933 para la población "Norte" y, to = –1.8581 para la población "Sur". Método de Allen Se tomaron como valores de trabajo del índice k, los estimados por el método de Tomlinson y Abramson para cada una de las poblaciones en estudio. Aplicando las ecuaciones: 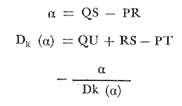 se obtuvieron como nuevos valores del índice k los siguientes: 0.52 para la población "Norte" y 0.5134 para la población "Sur". A partir de las ecuaciones, 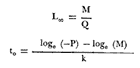 se obtuvieron los valores siguientes: L∞ = 123.82 mm; to = -1.7821 para la población "Norte", y L8 = 119.97 mm; to = -.891 para la población "Sur", respectivamente. Método Beverton Empleando los valores de L∞ obtenidos por medio de los métodos de Ford-Walford, Tomlinson-Abranison y Allen, se aplicó la siguiente ecuación: 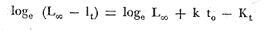 obteniéndose los resultados siguientes:  Selección y valoración de los coeficientes del crecimiento De acuerdo con las estimaciones realizadas para cada parámetro, mediante los métodos propuestos por los diversos autores, se han, desarrollado las curvas de crecimiento de Engraulis mordáx para: cada una de las poblaciones en estudio, y obtenido los valores calculados de las tallas, para cada edad, respecto a los observados. Los valores calculados y la suma de los cuadrados de las diferencias, aparecen en la tabla 5 para la población "Norte" y en la tabla 6 para la población "Sur". 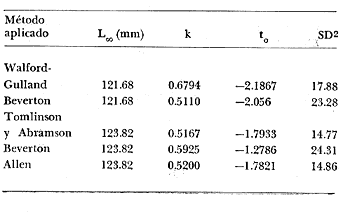 TABLA 5 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE" 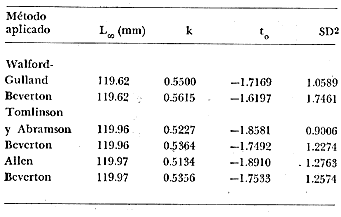 TABLA 6 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" Población Norte. Los parámetros L8 y k obtenidos por el método de Ford-Walford, presentaron diferencias significativas en comparación a los obtenidos por medio de las, otras técnicas. En el desarrollo de la curva de von Bertalanffy, los valores calculados que mostraron una mayor diferencia con respecto a los observados, fueron los obtenidos por medio de la aplicación de los parámetros de- Ford-Walford. Es decir, el método de Ford-Walford no proporcionó un buen ajuste de los datos con respecto a los otros métodos. Lo anterior puede deberse a que en el desarrollo de la curva de crecimiento se empleó el valor de to, determinado por el método de Gulland. Dicho valor presenta una mayor diferencia en relación a los obtenidos por Tomlinson Abramson y Allen. Considerando que el valor de to obtenido por el método de Gulland, podría estar relacionado con un error de muestreo al estar mejor representados algunos grupos de edad que otros; y basándose en las recomendaciones de este autor (Gulland 1964) que sugiere la aplicación de la fórmula en los grupos de edad ampliamente muestreados, se repitieron los cálculos para los grupos de edad de "0" a "3" años obteniéndose como valor de to, -0.926275. El desarrollo de la curva de von Bertalanffy, para este nuevo valor, mostró un mejor ajuste de, los valores calculados, correspondientes a los grupos de edad " 2", "3" y "4" Sin embargo, para el resto de los grupos de 'edad, "0" y "1", se observó una mayor diferencia entre los valores calculados y observados, en comparación a los obtenidos de la ecuación aplicada inicialmente en la que se consideran todos los valores de to de cada grupo de edad para la obtención de to promedio. Por lo tanto, las modificaciones aplicadas en el método de Gulland, tan sólo ajustan una parte de la curva del crecimiento de Engraulis mordaxno tomando en cuenta los valores restantes. Posiblemente los métodos de Tomlinson y Abramson proporcionan mejores valores de to que el método anterior, por ser técnicas más elaboradas que ajustan los errores que se pueden presentar entre la curva calculada y los datos observados. En el estudio del crecimiento de la anchoveta de la población "Norte", tanto la técnica de Tomlinson-Abramson como la de Allen, proporcionaron una buena corrección de los datos. La aplicación de la ecuación de Beverton a los métodos anteriormente citados, no proporcionó en ninguno de los casos un mayor ajuste, sino todo lo contrario, observándose una mayor diferencia entre los valores calculados y los observados, en comparación a los obtenidos por las otras técnicas. Población "Sur". El valor del índice k obtenido por medio del método de Ford-Walford, presentó diferencias significativas en relación a los valores obtenidos por Tomlinson-Abramson y Allen. Sin embargo, al aplicar los parámetros obtenidos por Ford-Walford y Gulland en el desarrollo de la curva de von Bertalanffy se obtuvieron valores similares a los observados, es decir, la aplicación de dichas técnicas presentó un buen ajuste de los datos. La técnica de Tomlinson y Abramson fue la que permitió una mayor corrección entre la curva calculada y los datos observados. Los valores calculados por medio del método de Allen, fueron los que presentaron una mayor diferencia con respecto a los observados. La aplicación de la ecuación de Beverton, proporcionó una optimización de los valores calculados por el método de Allen, al corregir el valor del índice k. Para el resto de los métodos aplicados la ecuación de Beverton, no proporcionó un mayor ajuste, observándose, al igual que en la población "Norte", una mayor diferencia entre los valores calculados y los observados. Ecuación del crecimiento de Engraulis mordax Las curvas calculadas que más se ajustan a los valores observados son los correspondientes a los coeficientes: L∞=123.82 mm, K= 0.5167 y to=-1.7933 años para la población "Norte", y L∞=119.96 mm, k=0.5227 y to=-1.8581 años para la población "Sur". por lo tanto, la ecuación de Brody-Bertalanffy adoptará para la anchoveta Engraulis mordaxde la población "Norte" la forma siguiente: 1t = 123.82 (1 - e-0.5167 (t + 1.7933) ) y para la población "Sur": 1t = 119.96 (1 - e-0.5227 (t + 1.8581) ) Los valores de los constantes de ambas ecuaciones, son los calculados por el método de Tomlinson y Abramson. Empleando dichas ecuaciones se han calculado las longitudes teóricas de las poblaciones "Norte" y "Sur" de la anchoveta para diferentes edades (Tablas 7 y 8). 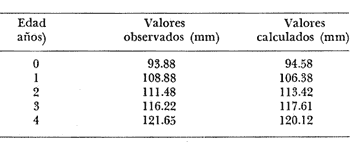 TABLA 7 LONGITUDES TEÓRICAS DE LA ANCHOVETA ENGRAULIS MORDAX A DIFERENTES EDADES EN LA POBLACIÓN "NORTE"  TABLA 8 LONGITUDES TEÓRICAS DE LA ANCHOVETA ENGRAULIS MORDAX A DIFERENTES EDADES EN LA POBLACIÓN "SUR" En la figura 1 se muestran las curvas teóricas del crecimiento en longitud de la anchovetaEngraulis mordax para las poblaciones "Norte" y "Sur" correspondientes a las edades de "0" a "4" años. En las poblaciones estudiadas, se observa que durante los primeros meses de vida, la anchoveta presenta un crecimiento rápido. La gran mayoría de los ejemplares capturados durante el mes de junio de 1982, presentaron un otolito formado por el núcleo, una banda de crecimiento lento y otra de crecimiento rápido, por lo tanto pertenecen a la clase de edad "F'. Por ello, el crecimiento es muy rápido durante los primeros meses. A partir del primer año de vida, la velocidad del crecimiento disminuye, incrementándose 7.04 mm. de la longitud entre el primero y el segundo año para la población "Norte"; y 6.5 mm de la longitud entre el primero y segundo año para la población "Sur". Entre el segundo y tercer año, de edad, el incremento de la longitud es de 4.19 mm para la población "Norte" y 3.86 mm para la población "Sur". Entre el tercer y cuarto afio el incremento en longitud es de 2.51 mm para la población "Norte" y 2.28 mm para la población "Sur". Valores extraordinariamente pequeños comparados con los alcanzados durante el primer año, de 108.88 mm para la población "Norte" y 104.63 mm para la población "Sur" (Fig. 2). Con el objeto de analizar las características del crecimiento de Engraulis mordax y de establecer algunas relaciones con otras poblaciones de esta misma especie, a continuación se presentan las longitudes medias para cada edad de la anchoveta, en distintas localidades del Pacífico Oriental. 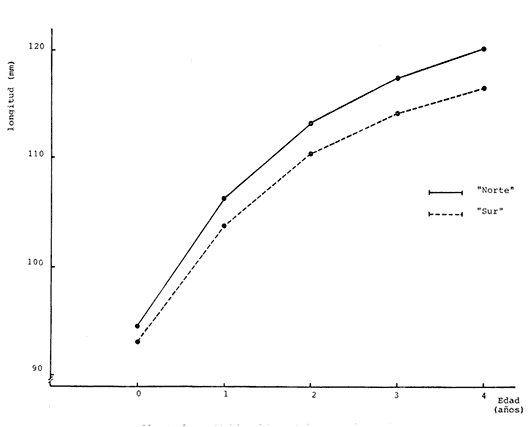 Fig. 1. Curva Teórica del crecimiento en longitud. 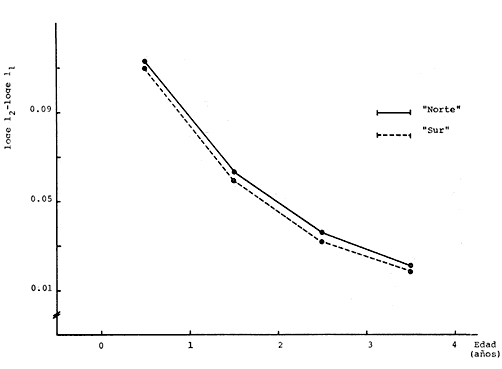 Fig. 2. Rango instantáneo del incremento en longitud. 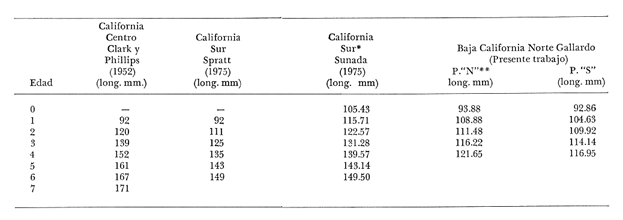 Sunada (1975)presenta datos de edad registrados para cada año durante 6 años, de 1965 a 1973, a excepción del año de 1967. En el presente trabajo, se presentan las longitudes medias obtenidas de promediar los valores de cada edad para los seis años con el objeto de facilitar su interpretación ** P. "N" población "Norte" P. "S" Población "Sur" De acuerdo con lo anterior, se observa que la anchovetaEngraulis mordax del área central de California, alcanza longitudes mayores a una misma edad a partir del tercer año, que la anchoveta del área Sur de California, diferencia que se incrementa conforme aumenta la edad. Similarmente, la anchoveta del área Sur de California alcanza longitudes mayores a una misma edad que la ancnoveta de Baja California. A continuación se hace una exposición de los valores de los parámetros de la ecuación de Brody-Bertalanffy, para las distintas localidades.  Se observa, que cuanto más elevado es el valor del coeficiente k , menor es el valor de L8 y por lo tanto, más rápido será alcanzado éste por la especie en cuestión, como ocurre con la anchoveta de la población "Sur" de Baja California. Las diferencias del valor de k entre las distintas especies, pueden estar relacionadas con lo establecido por Wheatherly (1966). Este autor pone de manifiesto, que las familias o especies, pueden llegar a presentar tasas potenciales de crecimiento determinadas genéticamente. Dichas tasas pueden variar dentro de cada especie o población de acuerdo con las condiciones ambientales de un área determinada. Como son la temperatura, la disponibilidad del alimento y la intensidad de la competencia, las cuales producen variaciones. Lo anterior sugiere, que la anchoveta habita en un área donde las condiciones de temperatura y alimentación son mejores para el desarrollo de la especie, y conforme las poblaciones se desplazan hacia el Sur, se desarrollan en áreas donde las condiciones son menos apropiadas para su crecimiento. Variaciones semejantes se observan dentro de una familia o especie como ocurre con el bacalao Gadus callarias en diferentes áreas del Atlántico Norte (Saemundson, 1923; Jensen y Hansen, 1931; Graham 1934; Trout, 1954; Martín, 1953; y Rollefsen 1934). En Merluccius merluccius en las áreas de Castellón y Cantábrico (Figueras, 1955; Robles et al, 1975) y otras especies. También es importante señalar que las tasas de crecimiento de una familia o especie, pueden presentar importantes variaciones debidas a la explotación pesquera. La cual puede producir una disminución en la talla comercial 7 de las especies sometidas a explotación, y por lo tanto, desviaciones del valor del coeficiente k. CRECIMIENTO SEGÚN EL SEXO
Crecimiento en los machos de Engraulis mordax Utilizando los datos obtenidos por medio de la lectura de otolitos, se obtuvieron los valores de k, L(infinito) y to por medio de la aplicación de las diversas técnicas tanto para la población "Norte", como para la población "Sur". En las tablas 9 y 10 pueden observarse los resultados obtenidos. Como puede deducirse de las sumas de los cuadrados de las diferencias, los valores más ajustados a los observados, corresponden a aquellos obtenidos mediante el método de Tomlinson y Abramson. De tal manera, puede proponerse como ecuación del crecimiento en longitud de los machos de Engraulis mordax,la forma siguiente: 1t = 119.86 (1 - e-0.5505 (t + 1.8325) ) para la población "Norte", y 1t = 116.49 (1 - e-0.5682 (t + 1.8326)) para la población "Sur" 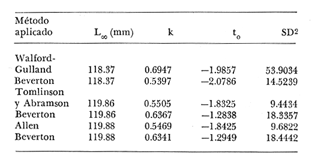 TABLA 9 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LOS MACHOS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE" 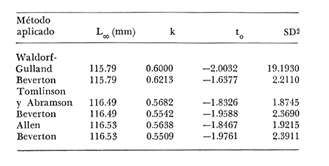 TABLA 10 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LOS MACHOS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" Las longitudes teóricas a diferentes edades aparecen en la Tabla 13 para la población "Norte", y en la Tabla 15 para la población "Sur". Crecimiento en las hembras de Engraulis mordax Para probar el ajuste de los coeficientes obtenidos por medio de los métodos empleados, se han desarrollado las curvas correspondientes, y comparado con los datos observados por medio de la suma de los cuadrados de las diferencias. Los resultados obtenidos, aparecen en la Tabla 11 para las hembras de la población "Norte" y en la Tabla 12 para las hembras de la población "Sur". 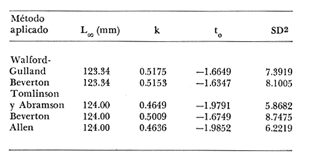 TABLA 11 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LAS HEMBRAS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE" 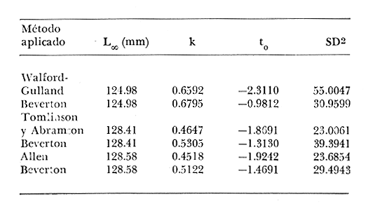 TABLA 12 VALORES DE LOS COEFICIENTES DEL CRECIMIENTO OBTENIDOS POR DIVERSOS MÉTODOS PARA LAS HEMBRAS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" En ambos casos, se observa, que la curva calculada que más se ajusta a los valores observados, es la correspondiente a los coeficientes obtenidos por medio del método de Tomlinson y Abramson. Por lo tanto, la ecuación de Brody-Bertalanffy adoptará en el caso de las hembras de Engraulis mordax la forma siguiente: 1t = 128.41(1-e-0·4647(t+1·8691)) para la población "Norte", y 1t = 124.0 (1 - e-0.4649 (t + 1.9791) ) para la población "Sur'. Las longitudes teóricas correspondientes a las diferentes edades, aparecen en la Tabla 14 para la población "Norte" y en la Tabla 16 para la población "Sur".  TABLA 13 LONGITUDES TEÓRICAS PARA LOS MACHOS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE"  TABLA 14 LONGITUDES TEÓRICAS PARA LAS HEMBRAS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE" 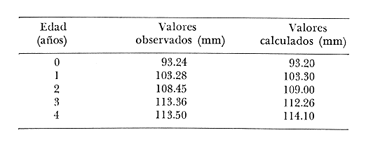 TABLA 15 LONGITUDES TEÓRICAS PARA LOS MACHOS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" 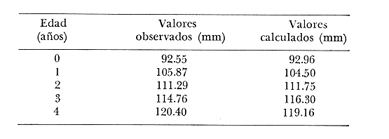 TABLA 16 LONGITUDES TEÓRICAS PARA LAS HEMBRAS DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" Diferencias del crecimiento entre sexos Con respecto a los parámetros del crecimiento entre sexos, se observa que el valor de k, es mucho más elevado en los machos que en las hembras, por lo que éstos alcanzarán más rápido una talla cercana a L∞. 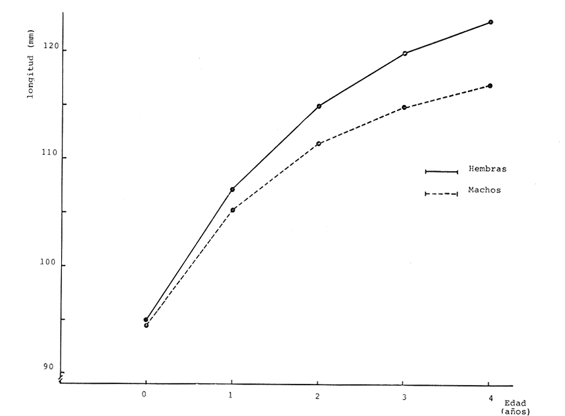 Fig. 3. Diferencias del crecimiento entre sexos para la anchoveta Engraulis mordax de la población "Norte". 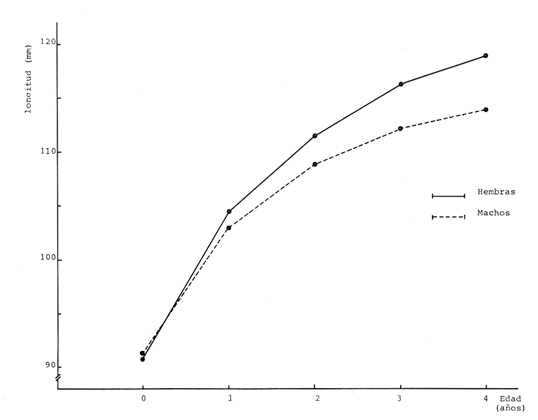 Fig. 4. Diferencia del crecimiento entre sexos para la anchoveta Engraulis mordax de la población "Sur" En las figuras 3 y 4 se observan las curvas teóricas de crecimiento en longitud, para machos y hembras de las poblaciones "Norte" y "Sur". Las diferencias del crecimiento entre sexos, aparecen a partir del primer año de edad. Entre el primero y segundo año de edad las hembras incrementan su longitud: 7.86 mm y 7.25 mm para las poblaciones "Norte" y "Sur" respectivamente. En ese mismo intervalo de tiempo, los machos incrementan su longitud: 6.16 mm y 5.7 mm, para las poblaciones "Norte" y "Sur", respectivamente. Al alcanzar su tercer año de vida, los incrementos en longitud para las hembras son de 5.0 mm. y 4.55 mm. respectivamente, mientras que el incremento para los machos de esa misma edad es de 3.52 mm y 3.26 mm respectivamente. Como puede observarse, las diferencías entre las tasas de crecimiento entre machos y hembras aumentan con la edad. Valores similares, son encontrados por Sunada (1975) para la anchoveta Engraulis mordax de California Sur. Es muy probable que las diferencias del crecimiento entre sexos deEngraulis mordax estén asociadas con la edad de madurez de uno y otro sexo. Es posible que los machos maduren a una edad más temprana que las hembras, por lo que éstos deben emplear una cantidad mayor de energía en la producción de los productos sexuales, mientras que las hembras de la misma edad, emplean la mayor parte de la energía en el crecimiento. CRECIMIENTO EN PESO
Relación Talla-Peso La línea de regresión entre la longitud y el peso, viene dada por la siguiente ecuación: W = 0.0000116882 X2.95 En la tabla 17 aparece la relación entre el peso y la longitud. En la figura 5, se muestra la recta de regresión de la relación entre el logaritmo del peso y el logaritmo de la longitud, calculada por medio de mínimos cuadrados. 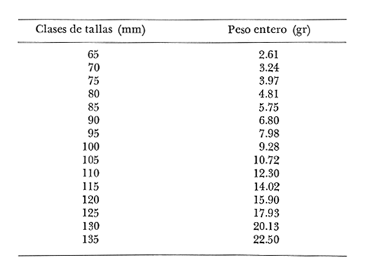 TABLA 17 RELACIÓN PESO-LONGITUD Peso a cada edad De acuerdo con la relación entre la talla y el peso, se obtienen los pesos a cada edad, a partir de los datos del crecimiento en longitud. En las tablas 18 y 19, se muestra el crecimiento en peso absoluto, y los valores del peso relativo, para cada una de las poblaciones en estudio. 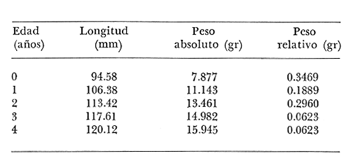 TABLA 18 CRECIMIENTO EN PESO, ABSOLUTO Y RELATIVO DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "NORTE" 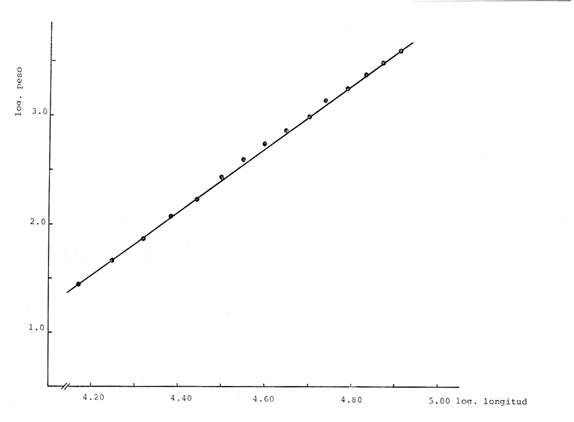 Fig. 5. Relación logaritmo del peso y logarítmo de la longitud. 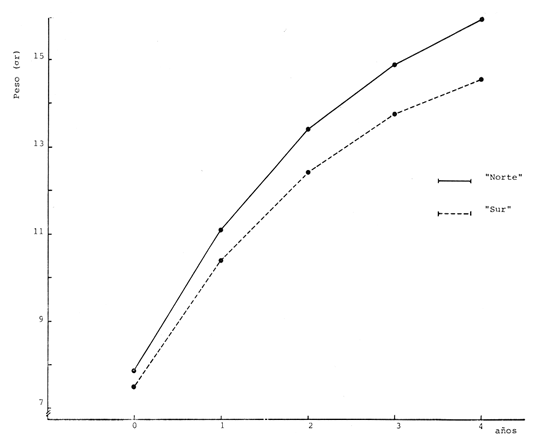 Fig. 6. Curva teórica del crecimiento en peso. 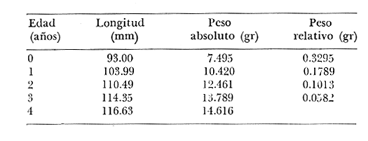 TABLA 19 CRECIMIENTO EN PESO, ABSOLUTO Y RELATIVO DE LA ANCHOVETA ENGRAULIS MORDAX EN LA POBLACIÓN "SUR" De acuerdo con los datos obtenidos, podemos apreciar que el incremento instantáneo en peso disminuye conforme aumenta la edad. Crecimiento teórico en peso Las ecuaciones del crecimiento en peso son las siguientes: wt = 17.437 (1 - e-0.5167 (t + 1.7933) ) 3 para la población "Norte", y wt = 15,882 (1 - e-0.5227 (t+1.8581) ) 3 para la población "Sur". Empleando dichas ecuaciones, se han calculado los pesos teóricos de Engraulis mordax a diferentes edades que aparecen en la Tabla 20 para las poblaciones "Norte" y "Sur". Las diferencias de crecimiento en peso por sexos, aparecen en la Tabla 21 para las dos poblaciones en estudio. Edad límite o longevidad La edad límite encontrada para la anchoveta Engraulis mordax, para cada una de las poblaciones en estudio y por sexos aparece en la Tabla 22. 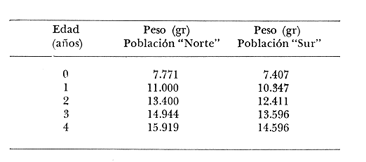 TABLA 20 RELACIÓN ENTRE LA EDAD Y EL PESO SEGÙN LA ECUACIÒN DE VON BERTALANFFY PARA LA ANCHOVETA ENGRAULIS MORDAX EN LAS POBLACIONES "NORTE" Y "SUR" 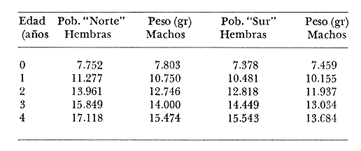 TABLA 21 RELACIÒN ENTRE LA EDAD Y EL PESO, POR SEXOS EN LAS POBLACIONES "NORTE" Y "SUR", SEGÙN LA ECUACIÒN DE VON BERTALANFFY PARA LA ANCHOVETA ENGRAULIS MORDAX La edad límite encontrada para Engraulis mordax, coincide con los datos expuestos por NikoIsky (1963) quien establece una longevidad de 2 a 5 años para peces cuya L∞ es de 100 a 200 mm. Los ejemplares de la población "Norte" alcanzan una longevidad mayor que los de la población "Sur". Similarmente, se observan diferencias entre la longevidad que pueden llegar a alcanzar las hembras con respecto a los machos. 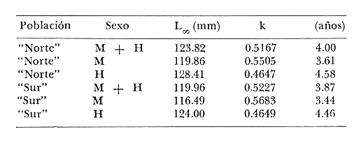 TABLA 22 EDAD LIMITE 0 LONGEVIDAD, PARA LAS POBLACIONES "NORTE" Y "SUR" Y POR SEXOS DE LA ANCHOVETA ENGRAULIS MORDAX Por otra parte, aplicando la ecuación de Taylor a los datos del crecimiento, obtenidos por Spratt (1975) se obtiene una longevidad para la anchoveta de California Sur de 8.3 años. La longevidad varía grandemente de una población a otra, como resultado de una adaptación a condiciones particulares. Es muy probable que la anchoveta de Baja California Norte, presente un crecimiento inicial más rápido, pero una menor longevidad, que los ejemplares de la misma especie en California Centro y Sur que habitan en aguas más frías. Esto quiere decir, que estos últimos tienen un coeficiente k menor y una edad límite mayor. Taylor (1968) establece que el coeficiente k guarda una relación directamente proporcional con el log. de la temperatura media del agua superficial; y una relación inversamente proporcional al valor de la temperatura con respecto a L∞ y la edad límite. También es de suponerse, que la longitud y edad límite dependen especialmente de la cantidad del alimento, del desove y de los efectos de la explotación pesquera. ConclusionesEn las poblaciones "Norte" y "Sur", las curvas teóricas del crecimiento que más se asemejan a los valores observados por medio de las lecturas de lassagittas, son las obtenidas aplicando el método de Tomlinson y Abramson (1961). Los parámetros del crecimiento para la población "Norte", son los siguientes: Talla máxima L∞ = 128.32 mm. Peso entero máximo W∞ = 17.437 gr. Coeficiente catabólico k = 0.5167 Edad teórica a la longitud cero (to) = -1.7933 Edad límite o longevidad (A(0,95) = 4 años. Los parámetros del crecimiento para la población "Sur", son los siguientes: Talla máxima L∞. = 119.96 mm. Peso entero máximo W∞ = 15.882 gr. Coeficiente catabólico k = 0.5227 Edad teórica a la longitud cero (to = -1.8581 Edad limite o longevidad A(0.95) = 3.87 años. El periodo de mayor crecimiento lineal ocurre durante los primeros meses de vida. A partir del primer año de edad, la velocidad del crecimiento disminuye considerablemente. La anchoveta de las áreas de Baja California Norte, en comparación con la anchoveta de las áreas de California Centro y Sur, alcanza longitudes menores a una misma edad, a partir del segundo año de vida, así como una menor longevidad. En Baja California los individuos de la población "Norte", presentan tallas y una longevidad mayores en comparación con los individuos de la población "Sur". En las poblaciones de anchoveta de Baja California Norte, el crecimiento de los machos, a partir del primer año de vida, es menor que el de las hembras de esa misma edad. Esta diferencia aumenta en proporción directa con la edad. Los parámetros del crecimiento por sexos, para la población "Norte", son los siguientes:  Los parámetros del crecimiento por sexos, para la población "Sur", son los siguientes:  El valor de k es mucho más elevado ' en los machos que en las hembras, por lo que éstos alcanzan más rápido una talla cercana a L∞ y presentan una longevidad menor. La relación peso-longitud es: W = 0.00001.16882 X2.95 AgradecimientosA Alfredo Laguarda Figueras, investigador del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, por la revisión y la ayuda para la publicación del presente. A Humberto Lobo Morales y Armando Charles Lobo por haber facilitado el apoyo de los recursos humanos y materiales de la Pesquera Zapata, S. A. de C. V. A Ernesto Ruffo, Jorge Camargo, Rubén Bouchez, Víctor Pérez y al personal del laboratorio de "Control de Calidad" y de la flota de la empresa mencionada, Por el apoyo técnico prestado. LITERATURAALLEN, K. R., J. Fish. Res. Bd. Canadá, A method of fitting growth curves of the von Bertalanffy type to observed data.1966.163-277233 BEVERTON, R. J. H., Notes on the use of theorical models in the study of the dynamics of exploited fish populations. U.S. Fish and Wildl. Serv. Fish. Lab., Beaufort, Misc. Contrib.,1954.1-1812 BRODY, S., Univ. Missouri Agric. Exp. Sta. Bull., 97: 101Growth rates.1927101-21597 CLARK, F. N. y J. B. PHILLIPS, The Northern anchovy Engraulis mordax mordax in the California FisheryCalif. Dept. Fish and Game1952.189-207382 FIGUERAS, A., Inv. Pesq., Datos sobre la edad y crecimiento de la pescadilla (Merluccius merluccius L.) del Levante (Sector Castellón) determinados por medio de los otolitos1955.73-861 FORD, E., An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933 J. Mar. Biol. Ass.UK, NS1933.305-38419 GALLARDO-CABELLO, M., An Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México,Determinación de la edad de la anchoveta Engraulis mordax Girard, en aguas de Baja California Norte (Pisces: Engraulidae).1985.221-234121 GRAHAM, M., Fish. Invest. Lond. Ser.Report on the North Sea Cod.1934.13-222 GULLAND, J. A., Manual of Methods of fish population analysis.FAO Fish. Tech. Pap.,1964.1-6040 JENSEN, A. S., y P. M. HANSEN, Investigations on the Greenland cod (Gadus callarias L.) with an introduction on the history of the Greenland cod fishery. Cons. Int. Explor. Mer. Rapp. et Proc. Verb.,1931.120-13472(1): MARTIN, W. R., Vital statistics of the cod and haddock in Subarea 4. ICNAF, 3rd Ann. Meet.,1953.1-2039 NIKOLSKY, G. W., Biological basis of mathematical simulation of fish population dynamics. Vopr. Ikhtiol1963.1-2934 ROBLES, R.; S. IGLESIAS; F. J. PEREIRO y J. A. PEREIRO, Bol. Inst. Esp. Ocean., Primeros datos sobre la dinámica del "stock" de merluza europea (Merluccius merluccius L.) frente al litoral gallego.1975.1-21196 ROLLEFSEN, G., The cod otolith as a guide to race, sexual development and mortality. Cons. Int. Explor. Mer. Rapp. et Proc. - Verb,1934.177-190882 SAEMUNDSSON, B., On the age and growth of the cod (Gadus callarias) in Iceland waters. Med. Komm. Danmarks Fisk. og Havunders., Ser. fish.,1923.32-5073 SPRATT, J. D. Calif. Dept. Fish. and Game Fish Bull.,Growth rate of the northern anchovy Engraulis mordax, in Southern California waters, calculated from otoliths.1975.116-126613 SUNADA, J. S. Calif. Dept. Fish. and Game Fish. RullAge and length composition of northern anchovies Engraulis mordax in the 1972-1973 season,1975.133-143613 TAYLOR, C. C. Cod growth and temperature.J. Conseil,1958.366-370233 TAYLOR, C. C.,Temperature and growth - The Pacific razor clamJ. Conseil1959.93-101251 TAYLOR, C. C., Temperature, growth and mortality - The Pacific cockle.J. Conseil,1960.177-124261 TAYLOR, C. C., Growth equations with metabolic parameters. J. Conseil,1962. TOMLINSON, P. K., y N. J. ABRAMSON, Fish Bull., Calif.Fitting a von Bertalanffy growth curve by least squares.1961.3-69116 TROUT, G. C., V. Reun. Cons. Int. Explor Mer.,Otolith Growth of the Barents sea Cod. Rapp. p. -1954.89-102136 VON BERTALANFFY, L,, Human Biology, A quantitative Theory of organic growth.1938.181-213102 WALFORD, L. A., Biol. Bull., A new graphic: method of describing the growth of animals.1946.141-147902 WEATHERLY, A. H., Nature, Ecology of fish growth.1966.1321-1324212
|