|
THE DEPTH DISTRIBUTION, RELATIVE ABUNDANCE AND STRUCTURE OF THE
POPULATION OF THE CHAETOGNATHA SAGITTA SCRIPPSAE ALVARINO 1962, IN THE
CALIFORNIA CURRENT OF CALIFORNIA AND BAJA CALIFORNIA.
Trabajo recibido el
7 de junio de 1982 y aceptado para su publicación el 28 de septiembre de
1982.
ANGELES ALVARIÑO
National Marine Fisheries Service Fishery-Oceanography
Center, La Jolla, California 92037, USA,
Sagilta scrippsae habita la región de
Transición del Pacífico septentrional, principalmente en los 25 a
475 m de profundidad, pero puede aparecer a niveles más abisales frente
a California y Baja California. Su mayor abundancia se ha observado entre los
100 y los 350 m de profundidad, decreciendo gradualmente en número de
individuos y en espacio habitado, en los niveles por encirna de los 100 m y por
debajo de los 350 m de profundidad.
El material de plancton
estudiado se obtuvo en las cuatro estaciones del año 1969 en aguas de
California y Baja California, efectuando arrastres diurnos y nocturnos en todos
los niveles batimétricos correspondientes a cada localidad. Los
arrastres, con redes Bongo de apertura y cierre automáticos, ocuparon
los niveles batimétricos de 0-25, 25-50, 50-75, 75-100, 100-225,
225-350, 350-475, 475-600 m, durante los cruceros de invierno (febrero-marzo),
primavera (mayo-junio), verano (agosto-septiembre), otoño
(noviembre-diciembre).
Sagitta scrippsae
aparecía con mayor abundancia en el período, de
agosto-septiernbre, resultando así estas poblaciones de magnitud doble a
las obtenidas en el período de noviembre-diciembre y triplicando las
correspondientes a la primavera; mientras que las poblaciones de invierno
resultaron de escasa magnitud.
Las poblaciones de
S. scrippsae frente a California y Baja California se
extendían más hacia el oeste, el sur y niveles
batimétricos más profundos durante el período de
agosto-septiembre, que en las demás estaciones del año. Esta
especie se observó en el estrato de 0-25 m de profundidad, solamente
durante el invierno (febrero-marzo).
Sapitta
scrippsae aparecía más abundante en todas las
épocas del año en la enfilación de Estaciones de la parte
más nórdica de la zona explorada, frente a California, y era
menos abundante o estaba ausente de las Estaciones más meridionales,
frente a Baja California. El patrón de la distribución de este
organismo indica que las poblaciones que aparecen en aguas de California y Baja
California proceden de zonas al norte de esa región.
Los datos que se han obtenido señalan que S.
scrippsae se reproduce en todas las estaciones del año, con un
máximo en verano y un mínimo en invierno. Se ha analizado el
ciclo de vida de esta especie y se describen las cuatro fases de madurez
sexual, indicando las tallas correspondientes a cada fase, así como la
distribución diurna y nocturna para cada estación del año,
en relación con el nivel batimétrico y la localidad. Se discuten
las fluctuaciones en las tallas que presentan los individuos plenamente
maduros.
La distribución de S.
scrippsae frente a California y Baja California presenta una
correlación positiva con la intensidad de la Corriente de California,
disminuyendo progresivamente a medida que la Corriente va perdiendo su
identidad. Por lo tanto, esta especie podría utilizarse como indicadora
de la corriente nórdica que avanza frente a California y Baja
California, para identificar la extensión abarcada por esas aguas
procedentes del norte, y estimar la intensidad de la corriente y grado de
mezcla que se produce en el Sistema de la Corriente de California. Se
señala además la relación evidente entre la
distribución de este Quetognato y las caracteristicas que presentan
algunas de las pesquerías endémicas y de temporada, en esta
región.
Sagitla scrippsae is an inhabitant of
the Transition region of the North Pacific, occurring between 25 m and 475 m
depth (but occasionally deeper) off California and Baja California. It is
abundant in the strata between 100 and 350 m, decreasing gradually in numbers
and area occupied above and below those layers.
The
collections analyzed in this paper were obtained in 1969 off California and
Baja California, from day and night hauls at identical locations and depth
strata (0-25, 25-50, 50-75, 75-100, 100-225, 225-350, 350-475, 475-600m depth)
on winter (FebruaryMarch), spring (May-June), summer (August-September), and
fall (November-December) cruises.
The greatest number of
specimens was collected on the August-September cruise; the number was about
twice that of November-December cruise, and three times that of the spring
(May-June) cruise. The lowest number occurred in winter
(February-March).
Sagitta scrippsae
occurred off California and Baja California in waters deeper and farthest west
and south in August-September than during any other season of 1969. It was
present in the 25-0 m stratum only in February-March 1969.
Saggitta scrippsae was most abundant on the
northern Line of Stations in each season, and less abundant or absent on the
southernmost Lines. This distributional pattern indicates the source of the
California population of S. scrippsae is in the waters north
of California.
Present data indicate that S.
scrippsae breeds in every season, with a peak in the summer and a low
in winter. The life history of the species is analyzed, and the four Stages of
maturity are described. Size ranges of each Stage are indicated, together with
day and night distribution with respect to depth, location and season.
Fluctuations in the size range of fully mature individuals are
discussed.
The distribution of S.
scrippsae off California and Baja California shows a positive
correlation with the intensity of the California Current, gradually decreasing
in numbers as the California Current losses its distinctiveness. Therefore, the
species could be used as an indicator of the northern flow off California and
Baja California to identify the farthest extent of these waters from the north,
and to estimate the degree of mixing taking place in the California Current
System. Evidence is outlined on the distribution of this chactognath and the
characteristics of some endemic and seasonal fisheries in this
region.
The areal distribution of the Chaetognatha in the California Current region is well known (Alvariño 1964, 1965a, 1966), but their life histories, depth distribution, and their role or function in the biological community in relation to the various confluent water masses in this region has not been thoroughly analyzed. Sagitta scrippsae appears to be restricted to the Transition region (Alvariflo 1962, 1964, 1965b; Kotori 1972), between the Subarctic and Pacific waters, from about 35° N to about 49° N and from 160° E to 140° W (Eber 1971). It extends into the Gulf of Alaska via de Alaska Gyre, and southward off North America via the California Current. S. scrippsae is also carried by eddies from the Transition region into the Central Pacific waters north of Hawaii (author's unpublished data). A few stray specimens have been obtained in the Pacific Subarctic waters, but none were found in the Bering Sea collections analyzed by Kitou (1976) and Kotori (1972). Two specimens of S. scrippsae were observed (Kotori and Hara 1972) at 540° WN, 172° 00´W in the Bering Sea: one from 325 m deep and another from 669 m. Kotori (1967) also found this species from 0 m to 560 m depth at locations (39° 53' N to 49° 58' N and 154° 29' W to 155° 24´W) in the Transition region, with highest abundance between 56 in, and 88 m depth. The distribution of S. scrippsae off California appears to fluctuate with the strength of both the West Wind Drift in the Transition zone, and the California Current. It is most abundant at the northern end of the California Current, gradually decreasing in numbers as the Current flows southward (Alvariño 1962, 1964, 1965a, 1966). The abundance of this species could therefore be useful in identifying the influence of the northern waters as they extend southward off California and Baja California, and their various degrees of mixing with other waters of the California Current System. The objectives of this study were to analyze the vertical and horizontal distribution of S. scrippsae Alvariflo 1962, and to evaluate this species as an indicator of the northern flow into the California region. Fluctuations of numbers, the areas occupied, and the structure of the populations were studied to determine key locations and times at which the distribution pattern of S. scrippsae populations would indicate biotic and abiotic characteristics of the northern flow off California in spring and summer. This information may prove useful in studing the survival of California endemic fish larvae and the status of some seasonal fisheries in this region. METHODSThe plankton collections use for analysis in this study were obtainer in 1969 during the following four seasonal cruises: 6902-03 (February-March), 6905-06 (May-June), 6908-09 (August-September), and 6911-12 (November-December). The Stations at which the samples were obtained correspond to Station Lines 70, 90, and 120 of the cruise pattern regularly occupied by the California Cooperative Oceanic Fisheries Investigations (CalCOF1) off the west coasts of California (United States), and Baja California (México). The vertical distribution series occupied eleven Stations in those three Lines: Stations 70.601 70.75, 70.90, and 70.110 on Line 70; Stations 90.45, 90.60, and 90.90 on Line 90; and Stations 120.45, 120.55, 120.70, and 120.90 on Line 120. Specimens of Sagitta scrippsae were identified and enumerated from the whole of each sample (no aliquots were made). Each specimen was measured in millimeters, from the anterior end of the head to the end of the tail segment, excluding tail fin, and the Stage of maturity was determined. Opening-closing Bongo nets (paired nets set side-by side) were used, with paired covers that, when closed, resemble a bongo drum. The nets in each set are labelled left (port) and right (starboard). These nets were towed at the same depth at each Station during both the day (near noon) and the night (near midnight). Thus, about 12 hours elapsed between the daylight and night hauls, giving the opportunity to evaluate day/night differences in vertical distribution. The Bongo nets take paired hauls at depth sampled. Therefore, 16 samples were expected from each day or night series, or 32 daily samples from each Station. A total of 352 samples were expected from each cruise (176 pairs). Due to irregular sampling on cruise 6902-03, however, only 188 samples were taken, resulting in a lower overall total. During cruises 6905-06, 6908-09, and 6911-12, sampling was conducted in day and night hauls at four 125 m strata from 600 to 100 m (600 to 475 m, 475 to 350 m, 350 to 225 m, and 225 to 100 m), and at four 25 m strata from 100 m to the surface (100 to 75 m, 75 to 50 m, 50 to 25 m, and 25 to 0 m). The depths sampled in cruise 6902-03, however, were shallower than in the other cruises, and often covered differently spaced strata. Three depths, 350225 ni, 475-350 in, and 600-475 m were not sampled, and sampling within the upper 225 m was variable. Therefore, cruise 6902-03 data are not strictly comparable to those from other cruises, although the uneven hauls in the upper 225 m for this cruise could be compared to combined adjacent strata from other cruises. Net mesh size also varied from cruise to cruise: On cruise 6902-03, the net mesh was NITEX 333 cod end NITEX 223 (sometimes NITEX 102). On cruise 6905-06, net mesh was 505 cod end 333, or net mesh 333 cod end 223 for the pair. On cruises 6908-09 and 6011-12, net mesh was 333 cod end 223. Changes in mesh size used in the net and cod end probably accounted, is some instances, for certain variations in the numbers and sizes of the specimens collected. Flow meters were used to close the nets when the combined volume of water strained by the two nets at each sampled stratum equally - 1000 m³. The amount of water filtered per meter depth, however, was not identical for each stratum sampled. In the shallow hauls that sampled a 25 m depth stratum, for example, the amount of water filtered per meter of depth fished was 40 m³, whereas in the deeper hauls that sampled the 125 m depth stratum, the amount of water filtered per meter of depth fished was only 8 m³. To obtain the watercolumn abundance of S. scrippsae, the counts were standardized to equivalent volumes of water strained for each meter of depth fished for each depth stratum. For simplicity in comparing the abundance in the upper net hauls (numbers per 1000 nº across 25 m depth), the values selected represented the number under 40 square meters of sea surface (i.e. the sum of the numbers in 40 m³ per each meter of depth sampled, throughout the 600 m depth strata). It was determined that, in order to obtain the same volume of water strained by the nets in both the shallow and deep water series the time spent per meter of depth in the sampling for a 25 m stratum was five times that for a 125 m stratum. A standardization was therefore necessary to obtain quantitatively comparable data for the strata in the upper 100 m and the strata below this depth (Krarr.er and Alilstrom 1968). The counts were standardized to 1000 m³ of water for each 25 m depth (number beneath 40 m²) by the formula, N = n (opening depth- closing depth) ÷ 25 N= Standarized number of specimens n = Number of specimens, for port (left) and starboard (right). Bongo nets combined. If the sample from either net (port or starboard) in any haul was missing, it was estimated that an equal number of specimens would have been obtained in the missing haul if it had functioned properly. Therefore, the values of the good available sample, port or starboard, were simply doubled. Wen it was evident, either from the sample or from direct observation on shipboard, that the net did not function properly and sampled other than the intended depth, the data were not used. The information in the results corresponds only to the nets working properly (that is, opening and closing at the right depth layer). If a haul was not made, there is no way to interpolate the amount of individuals present at the location and time. A total of 704 hauls was made with the paired Bongo nets, equal to 1408 expected samples. The total number of actual samples taken corresponds to 440 hauls with both nets working (880 samples), plus 59 hauls with only one net working: a total of 939 samples. On the comparable cruises 6905-06, 6908-09, and 6911-12, there were Stations that were completely missed, either during the daylight or night series. The most complete set of collections corresponds to cruise 6905-06, and the least complete to 6902-03. Less than 38% of the data are missing or incomplete. In all, 438 of 608 levels of sampling attempted were successf ul, in that at least one of the two nets appeared to have functioned. The following series of hauls did not appear in the collections: Day series at Station 90.45 of cruise 6905-06, night series at Station 70.60 of cruise 6908-09, night series at Station 70.75, and both day and night series at Stations 70.90 and 70.110 of cruise 6911-12. Malfunctions involving both nets at a given depth occurred for 10 pairs during 6902-03, 20 pairs for 690506, 28 pairs for 6908-09, and 22 pairs, of 6911-12 (total, 80 pairs of hauls) . Gear malfunction involving only one net of the pair occurred for 6 depth strata for 6902-03, 9 depth strata for 6905-06, 27 depth strata for 6908-09, and 17 depth strata for 6911-12, resulting in a total of 59 depth strata. These results are sumarized below: 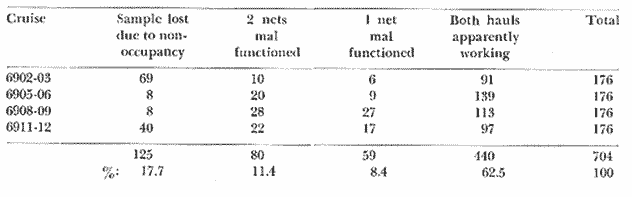 It is important to note that most of the missed Stations and gear malfunctions occurred on Line 70, the most valuable location for determination of the Sagitta scrippsae entering the California region: 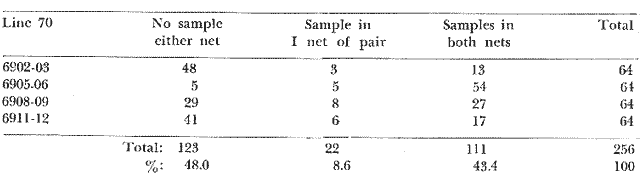 In addition, the deepest net seldom sampled below 300 m during 6902-03 cruise, so data are lacking for the deep strata during February and March 1969. Net failures were responsible for the occasional lack of samples from inter mediate levels within series. These gaps complicate the analyses of the material because of the attendant difficulties in presenting well defined vertical distribution patterns of the species. SEASONAL AND DIURNAL VARIATION IN THE BATHYMETRIC DISTRIBUTION OF THE SPECIESThe geographical and bathymetric distribution of S. scrippsae off California and Baja California is shown in Figures 1, 2, 3, and 4 for day and night sampling at each of the Stations on CaICOFI Lines 70, 90, and 120 for the four seasons of 1969. In addition, basic counts of abundance-per-1000 m3 of the different sizes of S. scroppsae (Table A) were averaged using values determined for each 25 m depth interval from 600 m to 0 m for each season (Tables B, C, D, E, and F). FEBRUARY-MARCH 1969 CRUISE (6902-03). Figure 1.
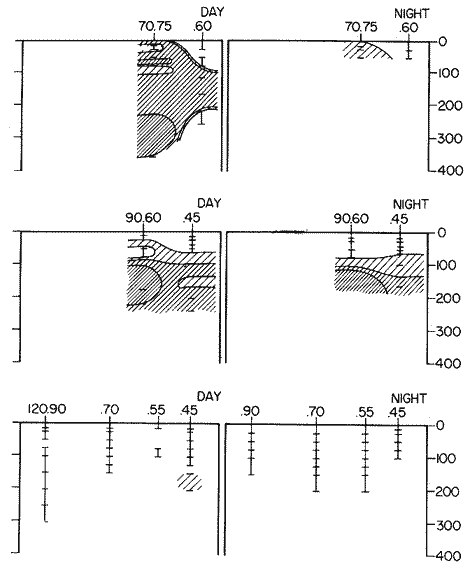 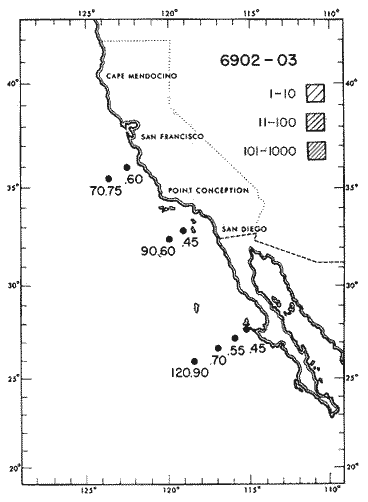 Fig. 1. Vertical days and night distribution of S. scrippsae on Lines 70, 90, 120, off California and baja California, Febrary-March, 1969 (6903-03). Vertical indicate were taken and the species was absent. 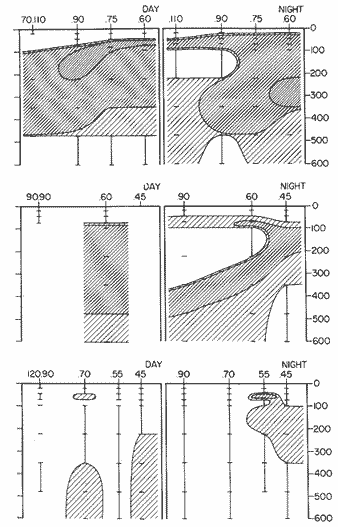  Fig. 2. Vertical day and night distribution of S. scrippsae on Lines 70, 90, 120, off California and Baja California, May June, 1969 (6905-06). Vertical lines indicate hauls were the especies eas absent. Line 70. Two Stations were-occupied with more daylight than night hauls. At Station 70.60, daylight hauls were made at seven depth strata, from 0 to 256 m in depth, and specimens were taken between depths of 208 to 75 m. No sample was obtained from the 50-25 m depth, and no S. scrippsae were collected at the 250-200 m depth. At Station 70.75, hauls were also made at seven depths, 0 to 350 m, and specimens were taken at all depths except 25-15 m. The highest concentrations were at 7550 m and 350-225 m, and next highest at three other strata (225-100 m, 50-25 m, an 15-0 m); the lowest (less than 10 speciments) occurred at 100-75 m. Night hauls were obtained only in shallow depths (50-0 m). Few specimens were obtained at Station 70.75. Line 90. Two Stations, 90.45 and 90.60, were occupied. Most hauls were taken during daylight. During the day, 10 depths were sampled from 240 to 0 m at Station 90.45. Sagitta scrippsae was absent both day and night in the upper hauls (levels in the 100-0 m strata), primarily from 60 to 0 m. Moderate numbers (11 to 100 specimens per 1000 m3) of S. scrippsae were collected during daylight at 240-168 m and 130-95 m depths, and at night at 168-130 m depth. Fewer than 10 specimens were obtained at other strata. At Station 90.60, eight hauls were made in daylight (213 to 0 m), and seven hauls at night (176 to 0 m). S. scrippsae was not collected in daylight hauls at 22-0 m or 75-34 m, nor in five hauls made in the 75-0 m strata at night. The highest concentration of this species was obtained at 213-100 m (daylight) and at 176-100 m (night), with moderate concentration at 100-75 m (daylight) ; the lowest numbers occurred at 34-22 m (daylight) and 100-75 m (night). 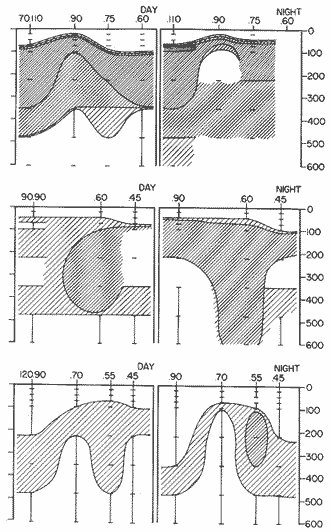  Fig. 3. Vertical day and night distribution of S. scrippsae on Lines 70, 90, 120, off California and Baja Califonrnia, Agust. September, 1969 (6908-09). Vertical lines hauls were taken and the species was absent. Line 120. Although four Station on Line 120 were sampled at various depths, only one specimen was taken at Station 120.45 in a 200-150 m depth haul during daylight hours. 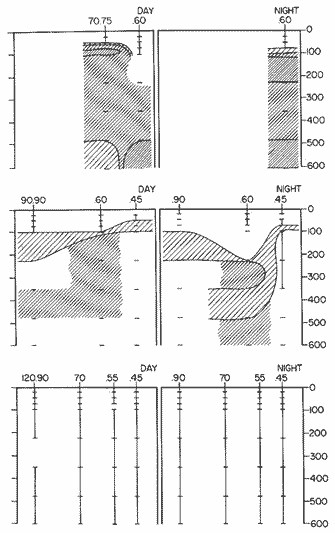 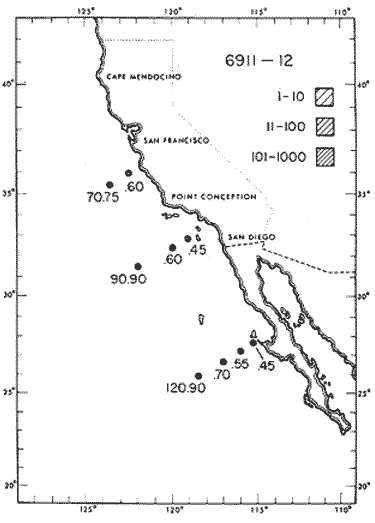 Fig. 4. Vertical day and night distribution of S. scrippsae on Lines 70, 90, 120, off California and Baja California, November-December, 1969 (6911.12). Vertical lines indicate hauls were taken and the species was absent. MAY-JUNE 1969 CRUISE (6905-06). Figure 2.Line 70. All Stations were occupied both during the day and at night. Eight strata, from 600 to 0 m, were sampled at four Stations. Daylight samples were not obtained at 350-225 m at Station 70.60, nor at depths 50-25 m, 75-50 m, 100-75 m, or 600-475 m at Station 70.110. At Station 70.60, no S. scrippsae appeared in the upper two hauls (at 50-0 m layers), at 600-475 m during daylight, nor in the 25-0 m at night. During daylight, high numbers (more than 100 specímens per 1000 m3 of water) were obtained at 75-50 m depth, and moderate numbers (11 to 100 specimens per 1000 m3 of water) at 225-100 m; few specimens (1 to 10) were obtained at 475-350 m depth. At night the highest concentration appeared at 350-225 m, with moderate numbers at 225-25 m; the lowest number occurred at 600-350 m depth. 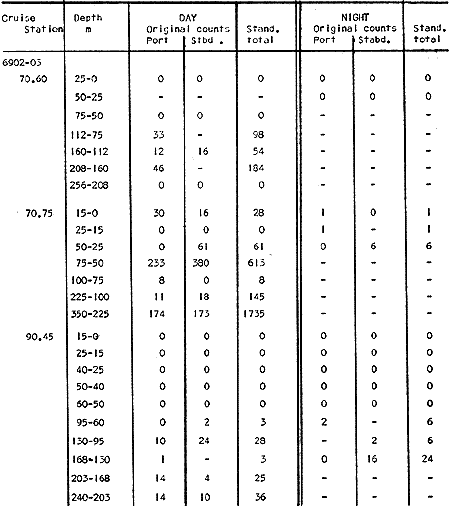 TABLE A THE BASIC COUNTS OF SAGITA SCRIPPSAE. STANDARIZED 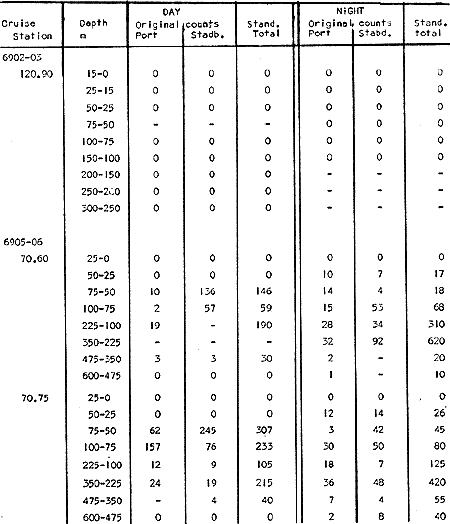 TABLE A (Continuation) 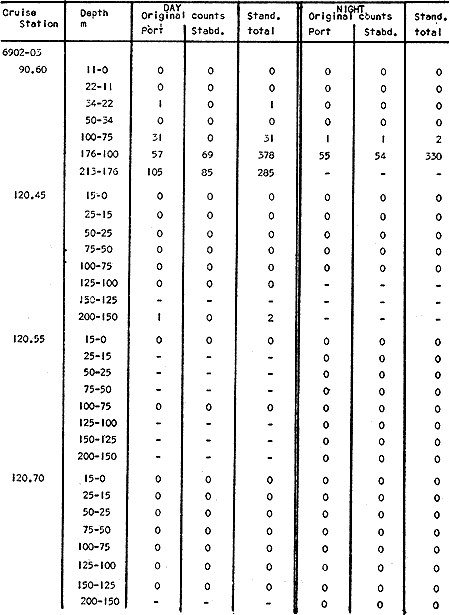 TABLE A (Continuation) 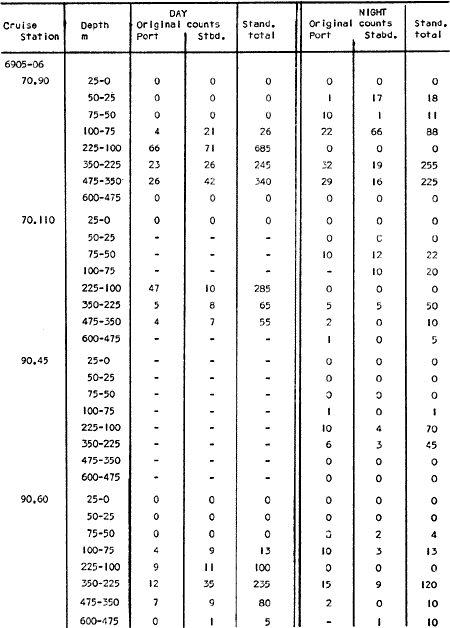 TABLE A (Continuation) 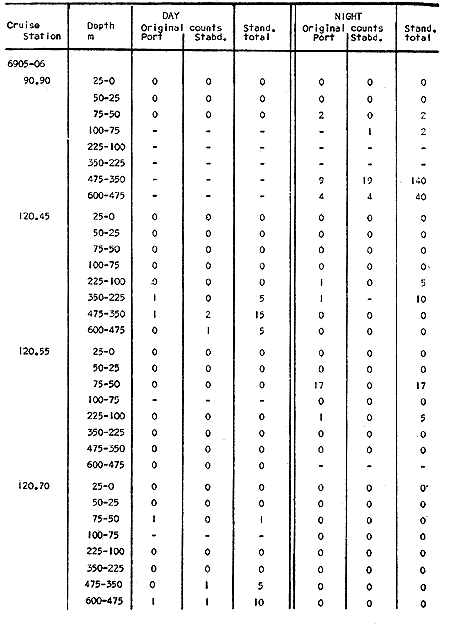 TABLE A (Continuation) 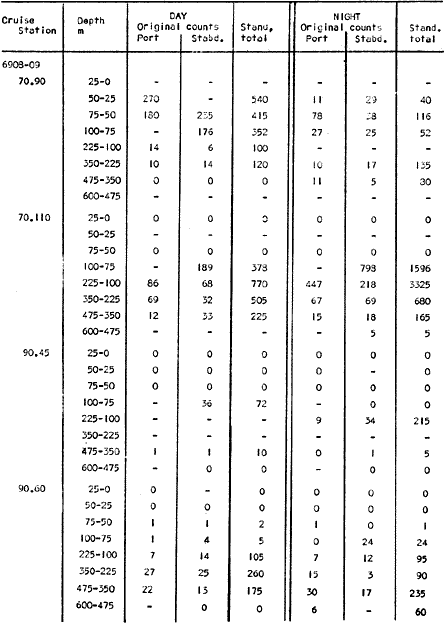 TABLE A (Continuation) 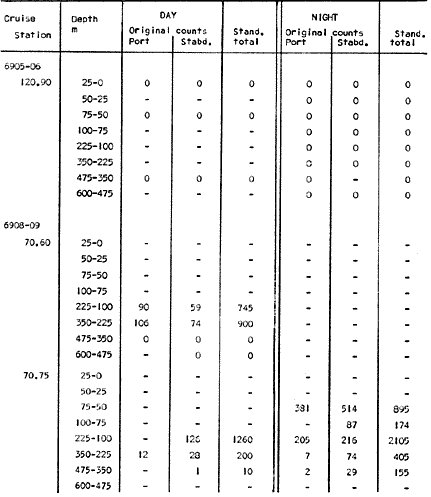 TABLE A (Continuation) 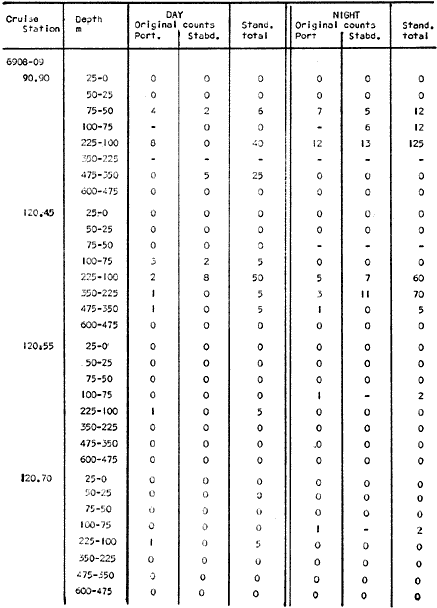 TABLE A (Continuation) 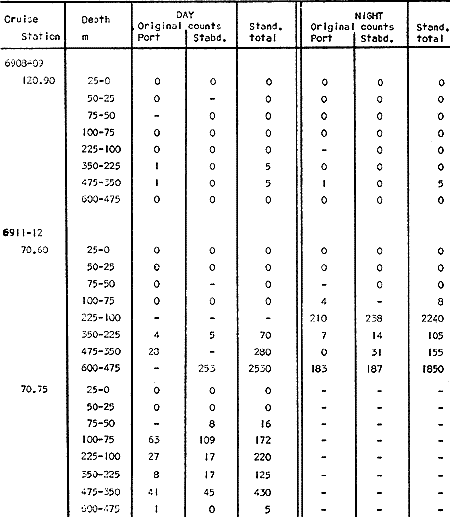 TABLE A (Continuation) 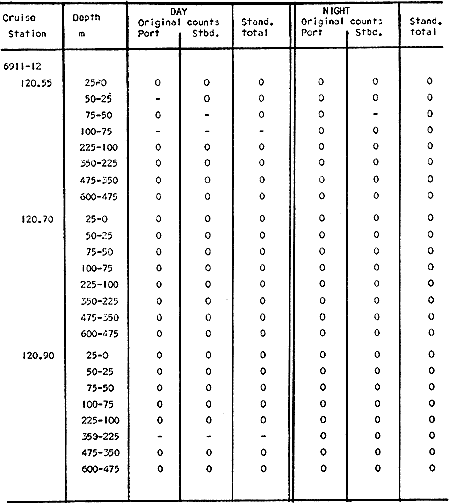 TABLE A (Continuation) 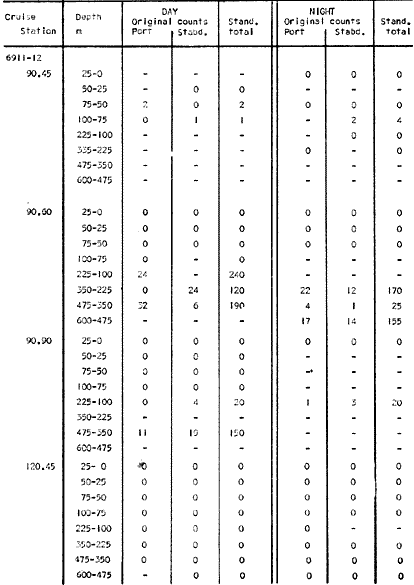 TABLE A (Continuation) At Station 70.75, S. scrippsae was absent in the upper two hauls (in strata 50-0 m3 , at 600-475 m during daylight, and at 25-0 m at night. The highest concentration during daylight was observed at 100-50 m, with moderate numbers at 350-100 m; the minimum was at 475-350 m. At night, moderate concentrations extended from 475 to 25 m, and the minimum occurred at 600-475 m depth. At Station 70.90, the species was absent during daylight in the upper three hauls (at 75-0 m strata) and at 600-475 m depth, and at night in the 25-0 m, 225100 m, and 600-475 m depth. The highest concentration during day light occurred at 225-100 m, with moderate e> numbers at 475-225 m and 100-75 m depth. At night, moderate numbers were found at 100-25 m and 475-225 m depth. At Station 70.110, S. scrippsae was absent in the uppermost hauls (in the 100-0 m strata) during daylight, and in the 50-0 m strata and 225-100 m at night. The species appeared in moderate numbers at daylight from 475 to 100 m depth. At night this concentration was restricted to 100-50 m depth; minimum numbers appeared at depths of 600 to 225 m. Line 90. Two Stations were occupied at daylight, but only Station 90.60 was completely sampled. At Station 90.90, samples corresponded to 25 m intervals from 75 to 0 m. At night, three Stations were occupied but samples were missed at 350-100 m at Station 90.90. No S. scrippsae appeared in the upper three daylight hauls (at 75-0 m strata). At night the speccies was absent from the upper three hauls at the Station closest to shore, and from the upper two hauls (50-0 m strata) at the other two Stations. The species was not observed at 600-350 m at Station 90.45, nor at 225-100 m at Station 90.60. During daylight, moderate numbers were found at 350-225 m, decreasing below and above these depths to 600 m and 75 m, respectively. At night, a moderate concentration occurred at 225-100 m at the Station closest to shore, and progressed offshore at two separate levels: 100-75 m and 350-225 m. At the Station farthest offshore, moderate numbers, occurred only at 475-350 m depth, with low concentrations at 100-50 m and below 475 m depth. At Station 90.45, the lowest number occurred at 100-75 m and 350-225 m depth. 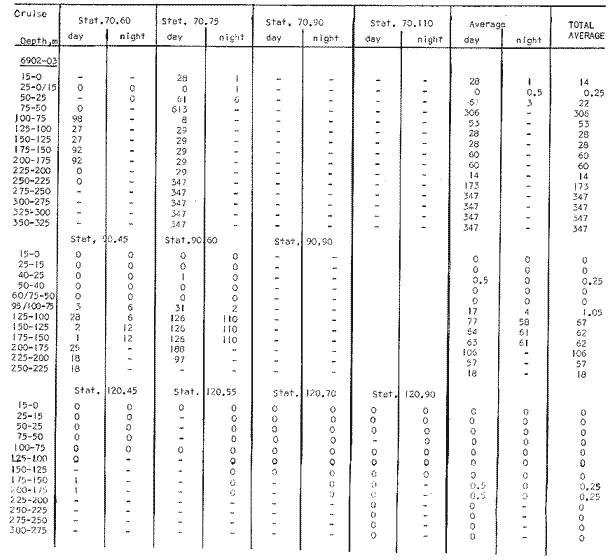 TABLE B DAY AND NIGHT AVERAGE AND TOTAL AVERAGE OF S. SCRIPPSAE POPULATION PER CRUISE, STATION AND DEPTH 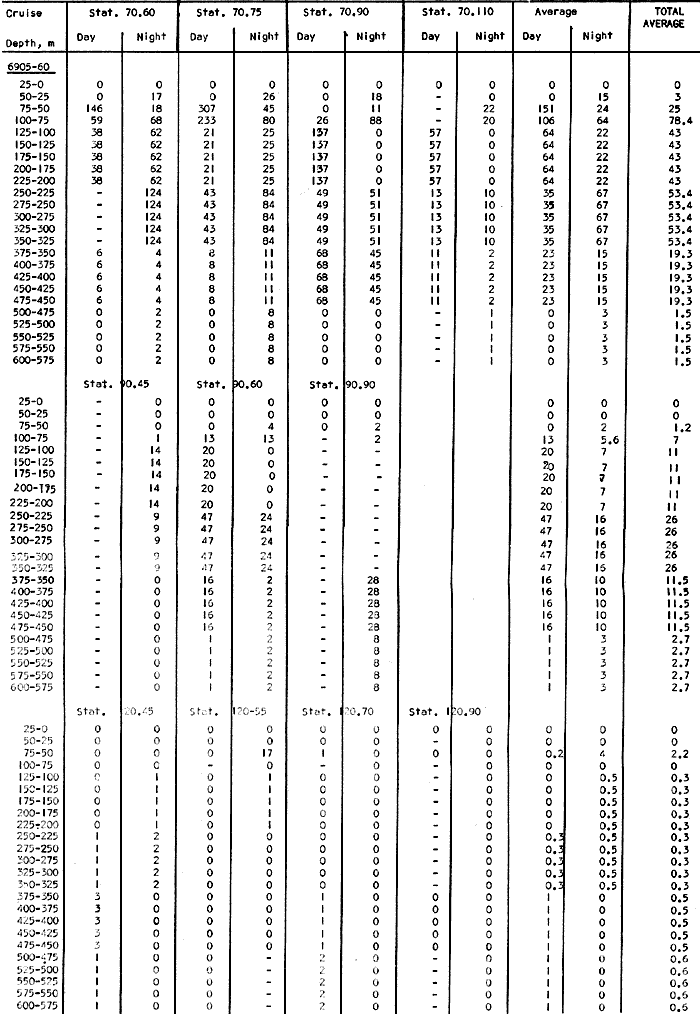 TABLE B (Continuation) 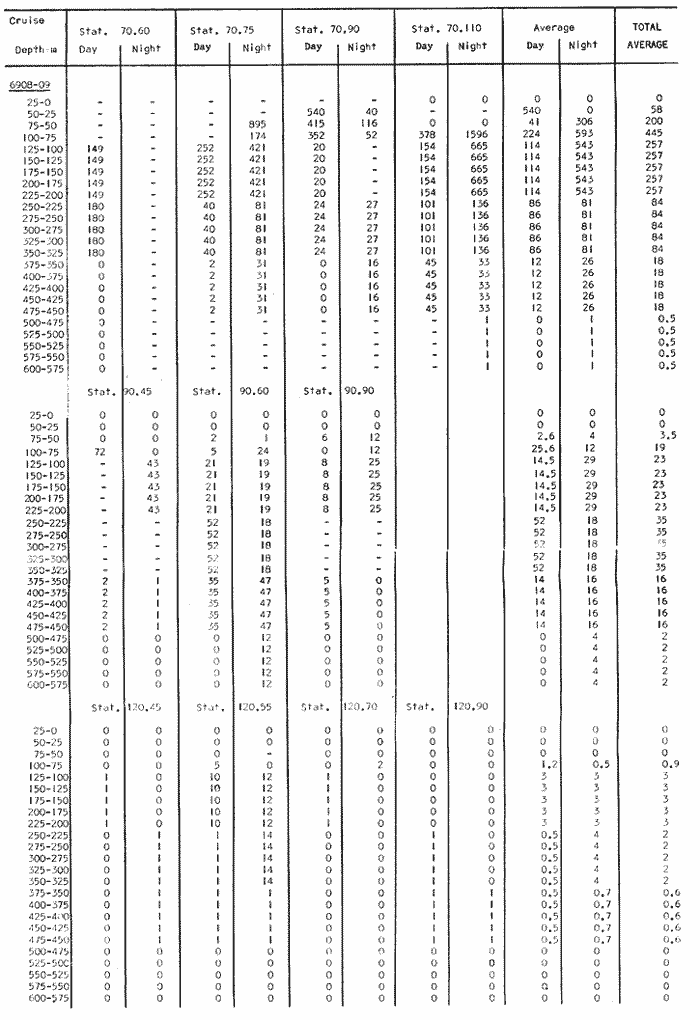 TABLE B (Continuation) 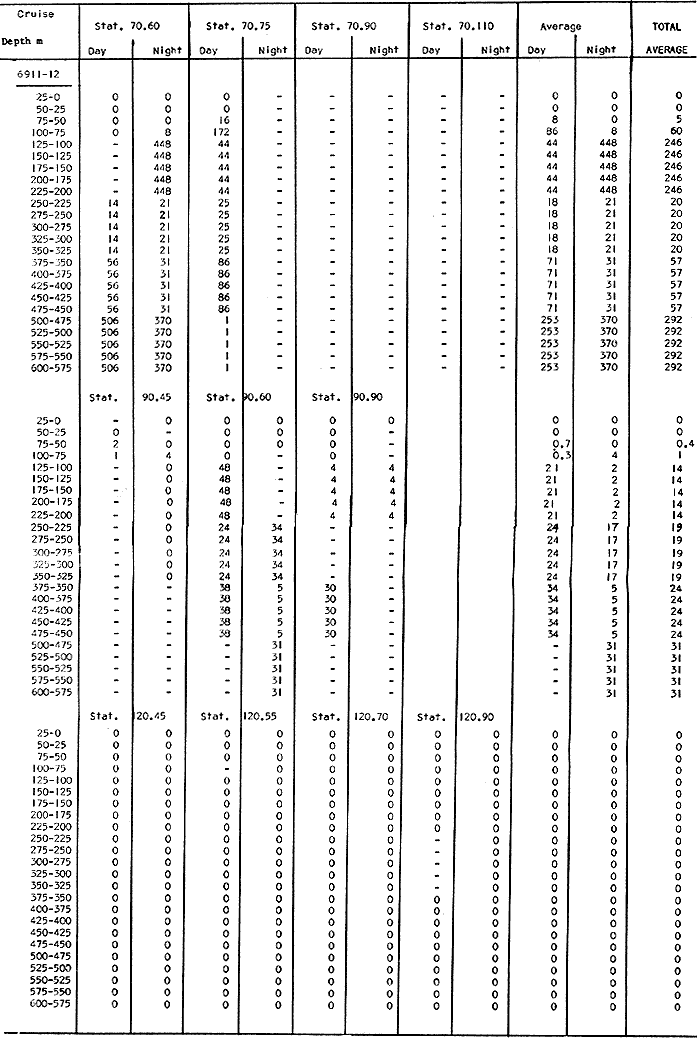 TABLE B (Continuation) 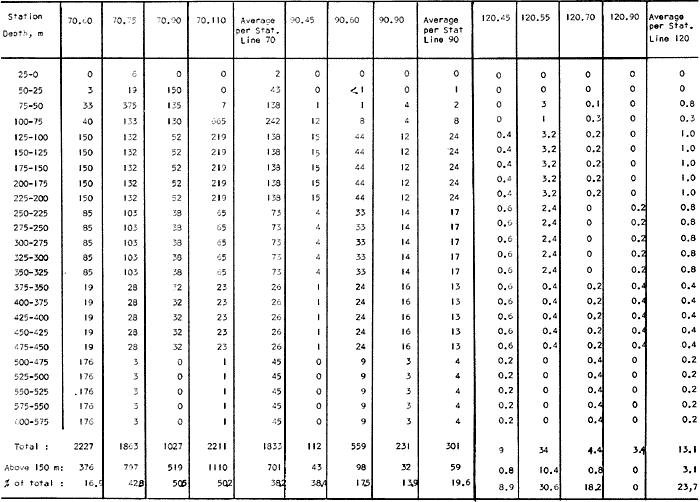 TABLE C AVERAGE TOTAL NUMBER OF SAGITTA SCRIPPSAE AT EACH 25 m DEPTH INTERVAL, FROM 0 TO 600 m 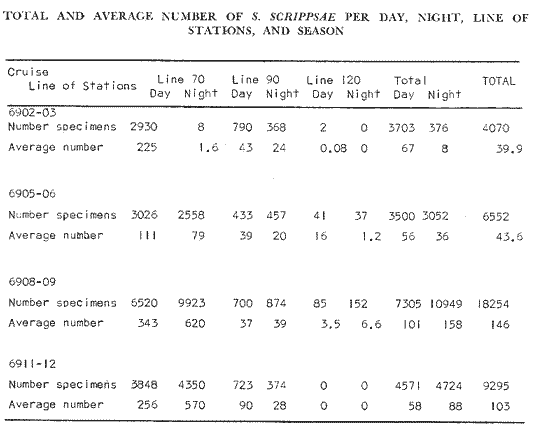 TABLE D TOTAL AND AVERAGE NUMBER OF S. SCRIPPSAE PER DAY, NIGHT, LINE OF STATIONS, AND SEASON
TABLE E TABLE E (Continuation) TABLE E (Continuation) TABLE E (Continuation) Line 120. All Stations on this Line were covered, both during daylight and at night. Daylight hauls, however, were missing at 10075 m for Stations 120.55 and 120.70; and at 50-25 m, 350-75 m, and 600-475 m for Station 120.90. Night hauls at 600475 m for Station 120.55 were also míssing. S. críppsae was present in 6 of the 25 daylight hauls made, and in 4 of 31 night hauls. During daylight, the species appeared in low concentrations at 600-225 m depth at Station 120.45, and at 75-50 m and 600-350 m at Station 120.70. It was found in low concentration at 350400 m at Station 120.45 during the night. At Station 120.55, it occurred in moderate concentration at 75-50 m, and low concentration at 225-100 m. This southerly extension of the S. scrippsae population appears relatively close to shore. AUGUST-SEPTEMBER 1969 CRUISE (6908-09). Figure 3.Line 70. During this cruise' all four Stations were occupied on this Line during daylight. At night, only three Stations were covered (no hauls were taken at Station 70.60). However, daylight hauls were missing for depths of 100 to 0 m from Station 70.60 and 70.75; at 600-475 m from Stations 70.75, 70.90, and 70.110; and at 25-0 m and 50-25 m from Stations 70.90 and 70.110, respectively Night hauls were missing from depths of 50-0 m, 25-0 m, and 50-25 m from Stations 70.75, 70.90, and 70.110, respectively; at 600-475 m from Stations 70.75 and 70.90; and at 225-100 m for Station 70.90. A high concentration of S. scrippsae occurred at 350-100 m depth of Station 70.60 during daylight. At Station 70.75, a high concentration appeared during daylight at 225-100 m, with moderate numbers at 350-225 m; minimum numbers occurred at 475-350 m. At night, the highest concentration remained at 225-50 m, with moderate numbers at 475-225 m. The highest concentration at Station 70.90 appeared in daylight at 100-25 m layers, with moderate numbers at 350-100 m. The high concentration at night occurred at 100-75 m, with moderate numbers above and below that layer. At Station 70.110, S. scrippsae was abundant at 350-75 m, with moderate numbers at 475-350 m during daylight. At night, the highest concentration observed for the whole year appeared at the 100-75 m depth. Below the 100 m layer, large numbers occurred at 350-100 m, with moderate abundance at 475-350 m. Few specimens were found at a depth of 600-475 m. The high and moderate concentration distribution pattern outlined a waved profile. 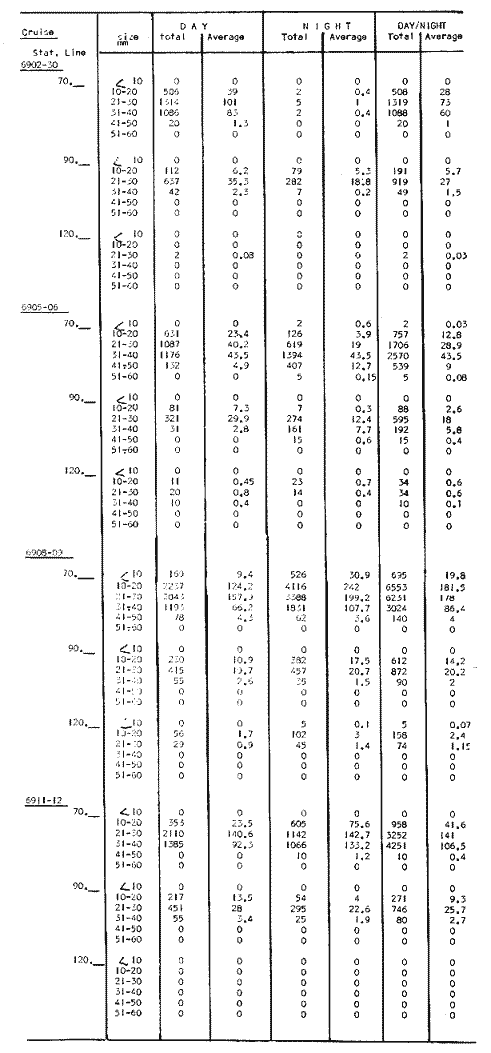 TABLE F AVERAGE DATA ON STRUCTURE OF S. SCRIPPSAE POPULATION DURING DAY AND NIGHT AT EACH LINE OF STATIONS DURING THE FOUR SEASONS OF 1969  TABLE G TOTAL AND AVERAGE STRUTURE OF THE POPULATION OF S. SCRIPSAE, DAY/NIGHT COMBINED, FOR THE REGION SURVEYED DURING THE FOUR SEASONS OF 1969. Line 90. The three Stations on this Line were all occupied at daylight and night, although samples at various layers were missing for Stations 90.45 and 90.90. S. scrippsae was absent from the three layers (75-0 m), two layers (50-0 m), and the 600-475 m depth in daylight, and the four and two upper layers at night. It was also missing at 600-475 m and 600350 m at Stations closest to, and farthest from . the shore, respectively. At Station 90.45, this species, appeared in daylight hauls in moderate concentrations at 100-75 m, with minimum numbers at 475-350 m. At night, moderate concentrations were found at 225100 in, and minimum numbers at 475-350 m depth. At Station 90.60, S. scrippsae was obtained in daylight in moderate concentration extending from 475-100 in, with a minimum at 100-50 m. At night, moderate concentrations were found in the layers from 600 to 75 m, with the lowest amounts at 75-50 m depth. The species appeared in low numbers in daylight at Station 90.90, while moderate numbers occurred at 225-50 m depth during the night. Line 120. All four Station on this Line were occupied both day and night at the eight stratified intervals per Station, except for level 75-50 m at Station 120.55 at night. S. scrippsae was absent from most of the upper and deep layers. At Station 120.45, it was present at 225-100 m and 475-225 m in small numbers in daylight and night, respectively. At Station 120.55, it occurred from 475 to 75 m in minimum concentrations during daylight; at night moderate concentrations appeared at 350 to 100 m depth, decreasing in number to the 475 m depth. At Station 120.70, the species occurred only in low numbers at 225-100 m in daylight and 100-75 m at night. It was found at Station 120.90 in minimum numbers at 475225 m and 475-350 m in daylight and night, respectively. The large wave mentioned in the distribution pattern of S. scrippsae in the profile of Line 70 was also evident at Line 90, and persisted in sharp outline at Line 120. NOVEMBER-DECEMBER CRUISE (6911-12). Figure 4.Line 70. Only three Stations were occupied on this Line: two during the day, and one at night. Samples were also missing. for 225-100 m layer at Station 70.60 during daylight. S. scrippsae was absent during daylight from the upper four and two layers for inshore and offshore locations, respectively, and from the upper three layers at night. At Station 70.60, high concentrations occurred during daylight at 600-475 in, and moderate numbers at 475-225 m; at night, two carters of high concentration appeared at 600-475 m and minimum at 100-75 m depth. A daylight low concentration occurred at Station 70.75 at 100-75 m depth, with moderate amount at 75-50 m and 475-100 m depth; the minimum occurred at 600-475 m depth. No collections were obtained at night from this locality. Line 90. The three Stations on this Line were occupied both during the day and at night, but several samples were missing: 25-0 m and 600-100 m during daylight and 50-25 and 600-350 m at night for Station 90.45; 600-475 m during daylight and 225-75 m at night for Station 90.70; and 600-475 m and 350-225 m during daylight, and 600-225 m and 100-25 m at night for Station 90.90. At Station 90.45, S. scrippsae was present in minimm amounts at 100-50 m during daylight and at 100-75 m at night. It occurred at Station 90.60 in moderate numbers at 475-100 m during daylight. At night, moderate numbers were found at 350-225 m and 600-475 in, with a minimum at 475-350 m. At Station 90.90, S. scrippsae occurred in moderate numbers at 475-350 in, with a minimum at 225-100 m during daylight, and a minimum concentration at 225-100 m at night. Line 120. All four Stations on this Line were occupied both during the day and at night, and at the eight stratified intervals, except for levels 100-75 m and 350-225 m at Stations 150.55 and 120.90 during the day However, S. scrippsae did not occur at any strata on this Line, either during daylight or at night. The population of S. scrippsae appeared at times more abundant in the upper layers. or at various depth strata at night, while in other instances this distributional abundance occurred during daylight. The greatest number of specimens (3325) were, obtained during a night haul on the August cruise at Station 70.110 (225 to 100 m depth). On the same cruise, 2105 individuals occurred at Station 70.75 at night and 1260 in daylight at the 225100 m depth. In November, 2539 specimens were collected in daylight Station 70.60 from the 600-475 m depth. On the same November-December cruise, a similar concentration of individuals was observed at night at the 225-100 m depth at Station 70.60. Slightly lower concentrations were noted here in the 600 to 475 m stratum (Figures 2 and 4, and Tables A and B). Unfortunately, daylight collecctions, from the 225 to 100 m stratum, which might have proved useful in comparing the night and day distribution of S. scrippsae, are missing for Station 70.60 of the November-December cruise. The greatest total number of S. scrippsae specimens was collected in August-September, followed by the number in November-December. The lowest numbers were obtained on the winter cruise. The amount of S. scrippsae obtained in August-September was about twice that of November-December, and three times that of spring (May-June) ; the lowest concentration occurred in winter (February- March), as seen in Table D. The seasonal day and night bathyraetric distributions of S. scrippsae are shown in Tables 13 and D. The greatest numbers of S. scrippsae appeared during summer, then fall, with the In lowest occurring during winter. It can be seen that day and night average, numbers are similar. The highest average numbers were present during daylight in winter and spring, while in summer and fall they were slightly greater at night. Comparing data at same depths for both 1 day an night series, indicates that equal numbers of S. scrippsae were taken, on the average, in both series. In February-March, the number of specimens decreased about 63% from Line. 70 to Line 90 in day and night populations combined, and decreased about 100%, from these Lines to Line 120. In May-June., the population extending to Line 90 was only 19.3% of that in Line 70, and the one reaching Line 120 was only 1.7%. In August-September, the population reaching the Lines 90 and 120 was 10.4% and 1.3% of Line 70, respectively, and in November-December, 13.3%, and 0% of Line 70 reached to Lines 90 and 120, respectively (Table D). A generalized pattern of the distribution of S. scrippsae was obtained by averaging the number of specimens at each 25 m depth interval from 0 to 600 m for all seasons, (Tables C and D). The highest concentrations occurred at level 100-75 m on Line 70, and at level 225-100 m on Line 90 and Line 120. The highest cencentration of the population of S. scrippsae occurred on Line 70, of which only 16% reached Line 90, and 0.7% to the Line 120. Stations 70.60 and 70.110 had the highest average number of specimens, with no strong difference between them. On Line 90, the average population appeared more concentrated at Station 90.60, and at Station 120.55 on Line 120. The data show that mortality, as indicated by the structure of the populations, is greater from spring through fall than in winter. Comparison of the present data for the upper 100 to 0 m with previous data off California for the seasonal cruises during 1954 and 1958 (Alvariflo, 1962, 1965a) indicates that the California Current was weaker during 1969 than in 1954, but stronger than in 1958. The main southward flow of S. scrippsae populations, as shown by its abundance, was at Stations 70.75 and 90.60 in February-March 1969; at 70.75, 90.60, and 120.70 in May-June; and at 70.75, 70.110, 90.60 and 120.55 in August-September. At this time it reached its highest abundance in a large area, extending both deeper and farther west and south than during any other season of 1969. For November-December (wit collections missing from several Stations) , the flow extended along Stations 70.60 and 90.60. The total number of tows were about the same for the combined seasons of spring and summer (578 hauls), and fall and winter (422 hauls) although those in winter cannot properly be compared with the other cruises. The numerical difference for both seasonal groups may not have statistical significance. The present data show that S. scrippsae generally inhabits depths between 25 m and 475 m, and occurs occasionally below that deeper level off California. It appears more abundant in the two strata between 350 and 100 m, gradually decreasing in abundance and in the area size inhabited above and below those combined strata. In February-March, S. scrippsae came closer to the surface layers than during any other season, appearing in the 25 to 0 m stratum. As expected, there is not a real remnant population of S. scrippsae restricted to the deep layers of the California waters during the winter months. However, the 1969 cruises covered only the southernmost portion of the distributional range of S. scrippsae along the northeast Pacific. The, sampling, therefore, did not extend far enough west off California and north of San Francisco to give a complete picture of the distribution of the species. In previous studies (Alvariño 1962, 1964, and unpublished data), it was observed that all life Stages of the populatign were present north of San Francisco and in the Transition region band. Thus, locations north of San Francisco, are strategic for the determination of the population structure of S. scrippsae entering the northeast Pacific flow of the California Current. The above analysis, of the distributional pattern presented by S. scrippsae off North America, indicates that the California Current population came from a northern source, sinking while progressing southwards. There remained the question as to whether the population could remain during the winter months in strata deeper than those reached by the oblique tows. The stratified hauls obtained in 1969 off California were used to determine the depth range and seasonal distribution of this species, and the results are presented here. The depth ranges of the hauls for the collections in this study do not coincide with those of former CaICOFI collections, which were made with a one-meter net in oblique tows to various depths as follows: from 1940 to 1950 ' with 100 m of wire out (to 70 m deep); from 1951 to 1968, with 200 m of wire out (about 140 m deep) , and from 1969 on, with 300 m of wire out (about 210 m deep). The plankton collections for 1954 and 1958 in the California Current region, which included integrated samples (oblique tows) generally from 0 to 140 m depth and, on a few occasions, to 400 in, were analyzed (Alvariño 1965a). In some instances S. scrippsae was present in the deep hauls, but it was not observed in the upper 140 m. However, a comparison of the catches of S. scrippsae obtained with Bongo nets during these studies, shown in Table C, includes standardized and average combined day and night number of specimens obtained per Station and season at the 25 m intervals. These data indicate that the largest numbers were usually obtained in the 225 to 100 m stratum. The data in this study indicate that while S. scrippsae appears in small numbers in the upper 100-m, the bulg of the population extends below those levels. Therefore, the CaICOFI standard hauls of 140 m depth could have not sampled most of the S. scrippsae population. No adequate reproduction needed to build up the population of S. scrippsae in the California and Baja California region was observed to take place at Lines 70, 90, or 120. It is therefore evident that, as the population of S. scrippsae drifts southward off California, it encounters environmental conditions which apparently are not conducive for its survival, and so gradually decreases in numbers until it disappears. PHSICO-CHEMICAL ENVIRONMENTThere is no clear evidence that physico-chemical factors such as temperature, salinity, and oxygen, or the presence of other populations such as phytoplankton blooms, might affect the population densities of S. scrippsae. The pattern of distribution of S. scrippsae shows a certain coincidence with the profiles of isohalines and oxygen isopleths. However, it appears that the environmental condition off California are not suitable for the normal spawning, growth and survival of this species inasmuch as only a few young and fully mature specimens were collected. It is known that the bulk of the population sinks, with the isothermal waters it inhabits in its progression southwards. In the Transition region, which is the proper habitat for this species (Alvariño 1962, Kotori 1976, and personal data from URSA MAJOR for August-October 1965, ZETES for January 1966 Expeditions, and in the California region) S. scrippsae appeared in waters between 50oC and 12oC Therefore, this species inhabits deeper strata off California than in its domain in the North Pacific, preferring waters of similar or closely-related temperature ranges. It is interesting to note that the peculiar wave-like distribution profile shown by S. scrippsae on Line 90 or cruise 6908-09, coincided closely with the isohaline profiles and oxygen isopleths (Figures 5, 6, 7, 8). However, in the locations where S. scrippsae sinks, the isotherms perform an inverse pattern, making an energence wave only in deep strata. "High oxygen uptake" is a known characteristic of arctic lamelibranch Species (Sparck 1936). Northern species displaced into warmer waters (Vernberg 1962) are known to increase their metabolic rates which may cause the animals to become deprived of food and oxygen. These factors may explain the high rate of mortality of S. scrippsae in the California region. In February-March, the population of S. scrippsae extended up to the 75 m depth at Station 70.75 (Figures 1 and 5) coincident with the upper displacement of the deep isotherms. In May-June, upwelling of oxygen contours and downward displacement of isohalines on Line 70 (Figures 2 and 6) fit roughly the distributional pattern of S. scrippsae. In August-September, the upward of isohalines and oxygen isopleths and downward and upward distribution of isotherms on Line 70 and Line 90, correlated with the distributional features of S. scrippsae. However, the distributional pattern of upward displacement and sinking of S. scrippsae on Line 120 does not agree with any physico-chemical profiles for that Line of Stations on that cruise (Figures 3 and 7). No explanation, however, can be made for the high concentration of S. scrippsae in the 600 to 475 m stratum at Station 70.60 in November-December for day and night collections (Figures 4 and 8). It is interesting to note that this welling up and sinking of S. scrippsae appears similar to the particular vertical distribution pattern of S. decipiens, a mesopelagic Chaetognatha collected in day and night hauls during the same cruises and Line of Stations. This peculiar distributional feature might have been produced by the large eddy (Wooster and jones 1970) formed by the warm deep undercurrent flowing northward off Baja California. Northward reverse of the transport by the California counter-current (Reid 1962, 1963) could not be detected by the distribution of S. scrippsae. However, this counter-current flows closer to the coast than the waters usually inhabited by S. scrippsae. The population of S. scrippsae reached the southernmost extension mainly at the 600-350 m or 350-100 m depth strata; below those levels the population virtually disappears before reaching areas off Punta Eugenia, Baja California. Unfortunately no calculations of the geostrophic flow off California are yet available for 1969; these would be of values as "the drift of the plankon may be better estimated from the geostrophic flow than from the instantaneous current measurements" (Wyllie 1966). Between consecutive day and night hauls, noticeable changes sometimes occurred in the vertical distribution of the species, while in other cases the bathymetric distributional pattern appeared similar or only slightly modified for both time and periods. Changes in the vertical distribution of the species at a location over so few hours could be due to variations in the environment caused by currents, eddies, mixing processes, etc., bringing about either concentration or dispersal of the populations. The fluctuations in the population density of this species may also be, due to certain undertermined favorable environmental factors which are present in the Transition waters. The, scarcity or absence of the species off California may suggest the absence of these favorable factors or the presence of environmental conditions unsuitable for the species survival. 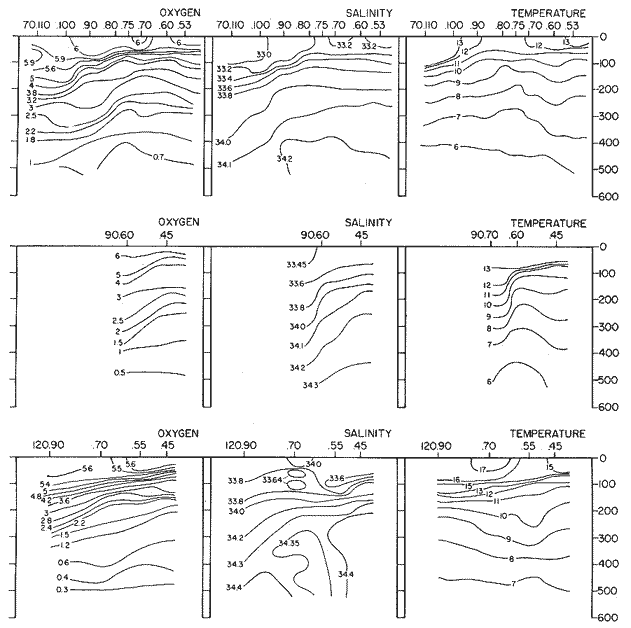  Fig. 5. Profiles of temperature, salinity, and oxygen for Lines of Stations 70, 90, 120, off California and Baja California,during February-March 1969 (6902-03) based on data University of California. 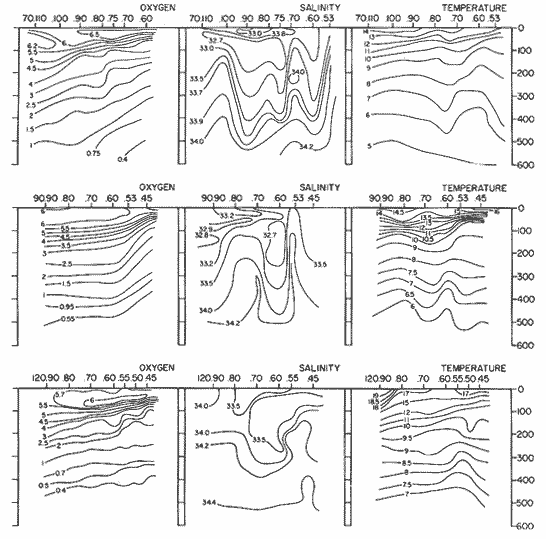  Fig. 6. Profiles of temperature, salinity, and oxygen for Lines of Stations 70, 90, 120 off California and Baja California, during May-June 1969 (6905-06) based on Data University of California. 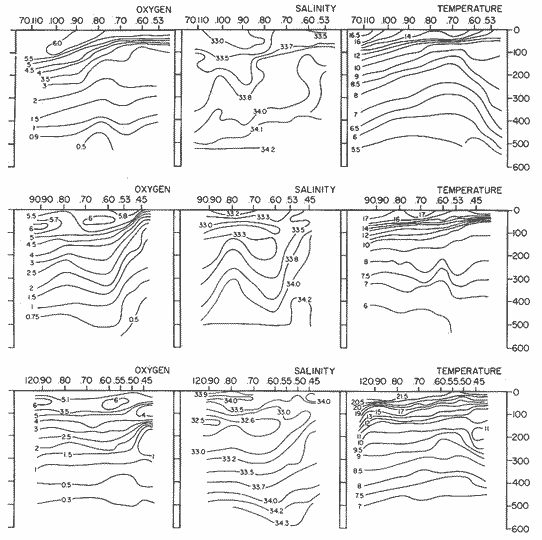  Fig. 7. Profiles of temperature, salinity, and oxygen for Starions 70, 90, 120 off California and Baja California, during August-Septembeer 1969 (6908-09), based on data University of California. 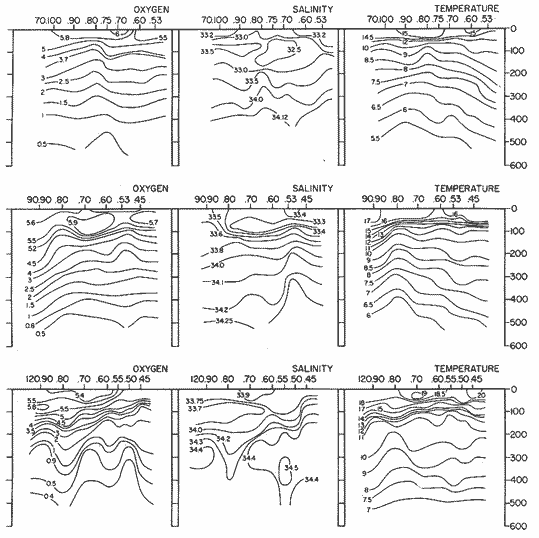  Fig. 8. profiles of temperature, salinity, and oxygen for Lines of Staions 70, 90, 120, off California and Baja California, during Agust-September 1969(6908-09), based on data University of California . BIOTIC CONDITIONS AND DAY AND NIGHT DISTRIBUTIONAL PATTERNCompetition with other zooplankters whould be considered as a factor in determining S. scrippsae. distribution. S. scrippsae inhabits the Transition region primarily with only two other species of Chaetognatha, S. elegans and Eukrohnia hamata; in the California waters, it occurs with more than 20 other species of this group. The eastern distributional boundary of S. scrippsae off California is roughly correlated with the westernmost extension of S. euneritica a typical neritic chaetognath of the North American Pacific (present data and Alvariño 1965a). S. euneritica mainly inhabits the uppei 100 m or 75 in, while S. scrippsae extends to layers below those levels. A comparative analysis of the vertical distribution of S. scrippsae and S. euneritica off California showed that where S. euneritica was abundant, few or no S. scrippsae were present. In February-March 1969 at Station 70.60, S. euneritica appeared abundant at the 75 to 0 m and the 260 to 200 m strata, with no S. scrippsae at those levels. It appears that S. scrippsae and S. euneritica inhabit different ecological environments, and the presence of S. euneritica may delineate the eastern boundary of S. scrippsae distribution off North America. Similarly, it was observed that the presence or abundance of S. enflata off California (information collected during the study of the collections for 1969) coincided with only scattered specimens of S. scrippsae. Differences in type, quality, and quantity of food in the Transition and the California regions should also be taken into account. Large concurrences of S. scrippsae (zooplankton predator) may be the result of an attraction to a concentration of food, and scattered numbers the reaction of the population to capture by predators: the typical behavior of predatory and prey organisms (rapid dispersal and sudden concentration). Decrease in numbers might also be due to increases in predation on these animals. If this is the case, distribution of predators (mainly fish and squids) of this species may have some bearing on the resulting distributional pattern. The data also indicate that some avoidance of the nets was taking place in daylight, with the larger specimens escaping by dodging the nets. Differences in the bathymetric distribution could be due to the seasonal effects of spawning. Breeding, however, does not appear to have been restricted to any particular season, since collections from different seasons had similar population structures. Thus, it is likely that differences in the diel vertical distribution might be brought about primarily by water movemetns, wherein water may he replaced at the Station location by waters with more dense or more scattered populations. Diel migration was shown to take place in some cases: some upward migration was observed at night in some instances, while in others upward migration appeared during daylight hours. However, there were some individuals which, remaining in the upper layers during daylight, may produce the resulted increase in the numbers captured in daylight hauls. The permanence of concentrated S. scrippsae in the upper layers during daylight may have been due to cloud cover, water turbidity or availability of food. The corresponding effect on the pattern of behavior of the animals, and the decrease in light intensity which influence the phototactic reaction of the animals, affected their vertical distribution pattern. LIFE STAGES AND THE STRUCTURE OF THE POPULATION OF SAGITTA SCRIPPSAE OFF CALIFORNIA AND BAJA CALIFORNIALittle is known of the life history of Chaetognatha, except for studies on S. elegans by Clarke, Pierce, and Bumpus, (1943), Dunbar (1940, 1941, 1962), Huntsman (1919), Huntsman and Reid (1940), Kotori (1976), McLaren (1966), Redfield and Beale (1940), Sameoto (1971, 197 3), and Sherman and Schaner (1968) : on S. hispida by Reeve (1970); on S. crassa by Murakami (1959); on S. nagae (Nagasawa and Marumo 1978); and on S. elegans and S. setosn by Pierce (1941), Russell (1932a, 1932b, 1933a, 1933b), and Wimpenny (1937). To learn more about the population structure of S. scrippsae four Stages of maturity were considered (Alvariño 1962) in order to determine reproduction, growth, and survival of the species: Stage I, specimens up to 30 mm long (ovaries starting to appear, or reaching up to anterior end of posterior fins); Stage II; specimens 31 to 40 mm long (ovaries reaching to midlenght of anterior fins); Stage III, specimens 41 to 50 mm long (ovaries reaching to anterior third of anterior fins); and Stage IV, specimens 51 to 60 mm long (ovaries reaching to neck region, ova large and mature filling the ovaries). Stage I includes three size groups (10 mm or less, 11 to 20 mm, and 21 to 30 mm). Stages II, III, and IV include the 10 ram size groups from 31 ram to 60 mm as listed. There may, of course, be exceptions to this classification scheme. For example, specimens 51 to 60 ram long could appear in Stage III, and specimens 31 to 40 mm may be fully mature or at Stage IV. From the observed conditions of the female gonads, there appears to be only one maturity cycle in this species. Prontandry is strong, with specimens as late as Stage 11 (39 to 40 mm long) having ripe seminal vesicles. Female gonads of the species are not ripe until the animal is more than 50 ram long, while the male gonads might go through several cycles of maturity in the life span of the animals. Evidence of this is shown in a large number of specimens up to 30 mm or 35 mm long which were at Stage I or II of maturity for ovaries and ova, while the tail segment was filled with sperm and the seminal vesicles were full or bursting. The fact that female gonads require a great ideal more time than male gonads to reach maturity, may be explained by the metabolic activity required to fully develop the female gonads and gametes. This activity consumes the reserves of the organism to such degree that, after maturity and spawning, the animal cannot recover and so dies. At the same time, there is evidence that male and female sexuality are separated in time for the same individuals; i.e., the hermaphrodite animals act first as males and later as females. Whit cross fertilization, the sperm is stored in the seminal receptacle of the ovaries of each of the coupled animals. Sperm reach the ripe stage to fecundate the ova at the time of spawning, when the female gametes are mature. Ovaries reach the neck region in the fully mature individuals (approaching 60 mm long) The ova are arranged in four rows (data from present studies and other previous analyses by the author, and Alvariño 1962). The total number of ova per individual ranges, from 700 to 1000. The length of each ovary ranges from 32 to 40 mm, reaching 48 mm in the largest individuals. The average number of ova in mature specimens can be used to determine ego, production and the apparent mortality at each stage of growth, and optimum conditions for reproduction, growth, and survival of the population in the species domain and along the margins of its distributional range. To accomplish this, data from the main distributional region are needed. However, with the data at hand, it appears that the highest mortality rate occurred in the smallest size group, based upon de numerical magnitude of that part of the population of S. scrippsae in relation to the high number of eggs the mature individuals presumable release. Tables E, F, G. It was found that S. scrippsae are present all year round in the Transition region of the North Pacific (the most stable domain of the species) where most reproduction takes place. Here the complete structure of the S. scrippsae population is present, from large fullymature, to young newly-hatched individuals (author's personal data, Alvariño 1962, 1964, 1966; Kotori 1976). The whole population structure is also represented in the northernmost location of the surveyed area, the region which approached, but is not close to, the main distributional domain of S. scrippsae. The number of mature individuals decreases progressively southward until the only ones present, as determined from these collections, are specimens at advanced Stage I of maturity. In the California waters, only a few very young and a few large mature specimens were occasionally observed in the northernmost part of the surveyed region. However, the structure of the population of S. crippsae in its main distributional region (where it reproduces) , has not been thoroughly studied. In the region surveyed off California, two phenomena are superimposed: the biological season, springsummer, when the major spawning occurs, and the maximum flow of the California Current coincident with that season, which conveys the population of S. scrippsae southrwards. In addition, data on the population structure obtained in port and sparboard samples for each haul, and the size of the individuals obtained in each net, shown evidence of patchiness, which could possibly be the result of the animals avoiding the net (Alvariflo, 1967). Stage I (including animals more than 10 mm in length) was present throughout the year, indicating that S. scrippsae spawns all year round in the Transition region and is brought into the California region with the flow of the California Current. Stage I specimens made up 90% of the population collected for each season of the sampling period. Stage II was also present throughout the year, but only a few specimens of Stage III. Some specimens of Stage IV appeared precociously mature, and were obtained off California in May. These were collected at 350-225 m adn only at night. Ontogenic stratification was observed, both bathymetrically and latitudinally, with the largest specimens appearing deeper and at the northernmost locations. The population fluctuation of this species is undobtedly related to changes in the California Current. Considering the rate of drift and the lifespan of the species, representatives of Stages II, III, and IV would be expected at the southernmost locations off California and Baja California. However, only specimens at Stage I were obtained. It could be suggested that some population replacement may occur locally, but this should be considered with caution because practically no fully-mature nor newlyhatched specimens were collected in those regions. The fact that only Stage I specimens were observed at the southernmost locations of the surveyed region indicates that the conditions may not be suitable to the survival of the species, and the surviving specimens may experience an arrest in their normal development. The abundance and size distribution range of S. scrippsae off California during day and night are presented in Tables E, F, and G, and Figures 9, 10, 11, 12 and 13. VERTICAL DISTRIBUTION OR. MATURITY STAGESSpecimens less than 6 mm in length were not observed in the surveyed region. Specimens less than 10 mm (9 mm individuals) were found in May 1969 at night at the most oceanic Station on Line 70 at 5025 m and 75-50 m depth at Stations 70.90 and 70.110, respectively. In August 1969, specimens 9 mm in length appeared at Station 70.90, and 6 ram. to 9 mm specimens were found at Station 70.110 in both daylight and at night. These small specimens were observed at 50-25 m and 225-75 m during daylight, and at 100-75 m and 225-100 m at night, being, more abundant at night than during daylight hours. In September, five specimens 9 mm in length were obtained at Station 120.55 during a night haul at 225-100 m depth (Table E). These data suggest that, in summer, spawning may extend into California waters, although the bulk of reproduction takes place north of Line 70. The production of young from that source is so large that some individuals are able to reach the surveyed region and survive there. Slight and occasional spawning may occur in the surveyed region, as indicated by the very young individuals obtained off Punta Eugenia. The fact that small specimens (6 mm long) reached the Stations on the northernmost surveyed Line suggests that gravid individuals were present close to that region in August, having been brought there by the strong flow from the northern waters during that season, and that only slight (almost nil) spawning can be considered taking place from Line 90 southwards. The largest size group of specimens (more than 50 mm long) were found only at Station 70.60, where five individuals 52 mm long were obtained in a night haul from 350-225 m depth. 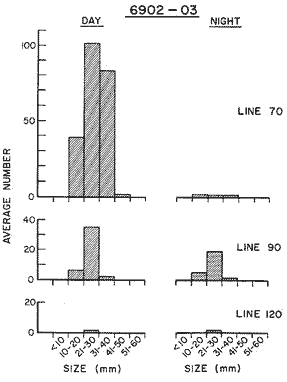 Fig. 9. Average day and night distribution of size group of the populationof S. scrippsae on Lines 70, 90, 120, off California and baja California, during February - March, 1969. SEASONAL DISTRIBUTION OF MATURITY STAGESDuring February-March 1969, the largest number of individuals during daylight and night hauls (size group 21-30 mm) appeared on Line 70, with decreasing abundance for the 31-40 ram , 10-20 mm, and 41-50 mm groups, the most abundant group, 21-30. mm, was also most numerous on Line 90, for both daylight and night. It was also the only size group represented at Line 120. However, the second most abundant group on Line 90 was 10-20 mm for both daylight and night, while group 3140 mm (which was second on Line 70) was scarce, and group 41-50 mm was absent. 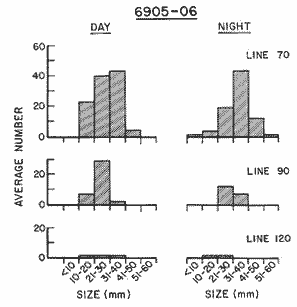 Fig. 10. Average day and night distribution of size group of the populationof S. scrippsae on Lines 70, 90, 120, off California and baja California, during May-June, 1969. During May-June, the highest average number of individuals occurring day and night on Line 70 belonged to size group 31-40 mm, followed (in decreasing abundance) by groups 21-30 mm, 10-20 mm, and 41-50 mm at daylight, and groups 21-30 mm, 41-50 mm, 10-20 mm, 50-60 mm and less than 10 ram at night. During the summer, the highest average number of individuals on Line 70 was from size group 21-30 mm, followed (in decreasing abundance) by size groups 1020 mm, 31-40 mm, less than 10 mm, and 41-50 mm. During night hauls, the most abundant size group was 1020 mm, followed by size groups 21-30 mm, 31-40 mm, less than 10 mm, 41-50 mm. How ever, the most abundant size group on Line 90, both day and night, was 21-30 mm (the most abundant group at night on Line 70), followed by 10-20 mm and 31-40 mm size groups. The remaining size groups were missing. On Line 120, the most abundant group was 10-21 mm, followed by 21-30 mm for both daylight and night. During the night hauls, five specimens 9 mm in length were obtained (indicative of an occasional slight spawining in the area). The sharp difference in the range of size groups observed for daylight and night hauls during the spring was not evident in the summer. 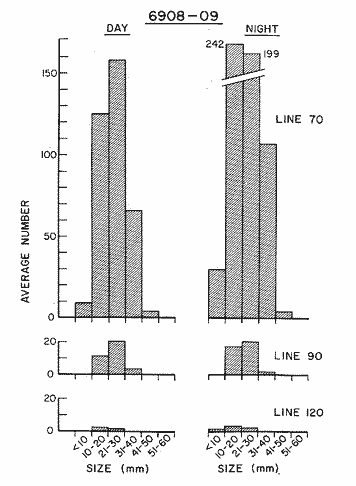 Fig. 11. Average day and night distribution of size group of the population of S. scripsae on Lines 70, 90, 120, off California and Baja California, during August-September, 1969. 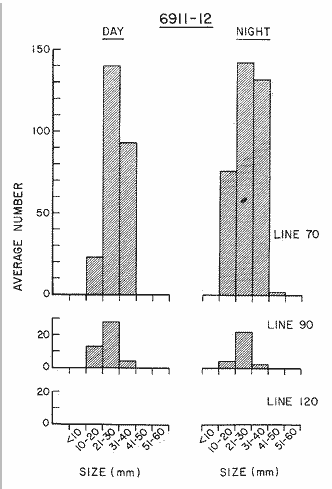 Fig. 12. Average day and night distribution of size group of the population of S. scripsae on Lines 70, 90, 120, off California and Baja California, during November - Dicember, 1969. During the fall, the most abundant size group on Line 70 was 21-30 mm, followed by size groups 31-40 mm and 10-20 mm for both daylight and night, and 41-50 mm at night. Specimens collected at 600-475 m at Station 70.60 ranged from 15 to 38 mm in daylight and 11 to 38 mm at night, with modes of 27 mm and 21 mm, respectively. Size group 21-30 mm was also the most abundant on Line 90, followed by the 10-20 mm and 31-40 mm groups. No representative of S. scrippsae reached Line 120 in the fall. There was also a wider range of size groups at night than at daylight during the fall. This phenomenon was not evident during winter and summer but was obvious during spring and fall when the turnover of biological and climatic periods occurs. On the whole, a wider range of size groups were captured during the night hauls than the daylight. The animals' behavior varied from night to day, being more alert to vibrations produced by the approaching net during daylight than at night, indicated by the size ranges obtained during both daily periods. Groups at both extremes of the size range appeared less abundant during daylight than at night. 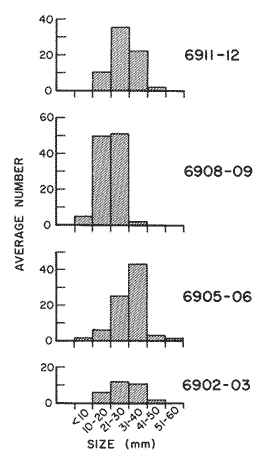 Fig. 13. Average size distribution (day and night crn bined) of S. scrippsae off California and Baja California during the seasons cobined of 1969. The structure of the population of S. scrippsae in the uppre 100 m during 1969 was analyzed to compare it with the population obtained in the standard 1954 and 1958 CAICOFI cruises. The population collected in the upper 100 m in the present studies included specimens ranging from 12 to 38 mm in FebruaryMarch, 9 to 45 mm in MayJune, 6 to 45 mm in August-September, and 10 to 38 mm in November-December. Specimens 44 to 45 mm long were occasionally obtained at 100-0 m levels during May and August 1969. These data and my unpublished notes on the sizes of S. scrippsae collected in the monthly cruises off California during 1954 and 1958 indicate that large mature specimens are virtually absent from the upper 100 m in this region, and that Bongo nets capture larger individuals than the one meter CaICOFI nets. The one meter net is therefore not adequate for collecting S. scrippae; Bongo nets without bridles are more effective. Although growth rates are known for S. hispida (Reeve 1970) and S. gazellae (David 1955), the closest relative of S. scrippsae, there is no knowledge of the growth rate of S. scrippsae because no studies have been made of its population structure in the main area of its distribution (i.e., the Transition region band of the North Pacific). Growth rates. may vary for the different sized individual, being higher for the young, and progressively decrease with increasing length. When development of the gonads occurs. food intake is increasin,oly channeled to this activity, with a proportionately decreasing amount given to body growth. The disappearance of fully mature individuals may be due to their inability to survive under adverse conditions. When the whole population declines, large mature individuals will occur in such small numbers that there will be little chance of their being represented in the collections. The consistently low percentage of the size group less than 10 mm long throughout the four surveys is remarkable, and may be due to several factors. The relatively rapid growth rate of the animals in the youngest generation while reaching the medium size Stage I, for example, is a possible cause of the reduced number of specimens of that size in the population at any one time. Gear limitations should also be considered in the loss of the individuals at both extremes of the size range, although this phenomenon may apply more to the larger sized animals than to the very small ones. The distributional domain of S. scrippsae is strictly bound to the Transition waters. Therefore, the temperatures or salinities encountered by this population off California should be adequate for spawning and survival of the species. However, practically no spawning took place south of San Francisco during 1969. Perhaps food quality or another factor inhibited sexual development. It has been considered that growth, maturity of the gonads, and production of ova or eggs, may be related to the characteristics of the food ingested. S. scrippsae devours every type of zooplankton (fish larvae, medusae, chaetognaths, copepods, euphausiids, and other crustaceans). Both the amount and quality of available food may be critical factors because this species is incapable of storing food reserves. Sameoto (1973) stated that food was not a limiting factor in determinine the erowth rate of S. elegans in the Bedford Basin, Nova Scotia, and also that growth was limited by the low temperature water. The present study does not support this statement. Growth rate is retarded by lack of food or other unfavourable conditions in the natural habitat (Russe II 1932a), and growth rate and maturity of the gonads are accelerated with increasing temperature (Bullock 1955). Some specimens of S. scrippsae were found to be retarded in sexual development; for example, 40 to 45 mm specimens normally corresponding to maturity Stage III appeared to be at Stage I or II. Such differences in the maturity of the gonads may be dependent on food and environmental conditions, which would account for an effect in the physiology of these animals. Temperature may be relevant when other factors remain constant, such as food supply or certain undetermined environmental factors. In this study, although temperatures encountered in the strata inhabited by the species off California were similar or within the higher segment of its normal temperature range in its main distributional region, gonad development did not appear directly related to this variable. Growth rate should not have been retarded but enhanced by such temperatures; however, this was not the case, as seen in the retardation and failure of gonad development. The few shorter specimens of S. scrippsae (35, 40, 41, and 45 mm long) at Stage IV of maturity (female gonads) obtained during May-june cruise, may have resulted from two possibilities: either those specimens encountered optimal conditions of food and some other factors, which, toogether with high temperatures permitted precocious development of the gonads, or, as observed by Reeve (1970) in laboratory studies on S. hispida, they experienced shrinkage in body length due to starvation, seemingly at Stage IV. A few S. scrippsae observed had parasites (nematodes and tremetodes) in the body cavity. In these specimens, development of the ovaries was retarded: individuals more than 40 ram in length were at maturity Stage I. The lack of adequate food uptake may be one of the important factors responsible for the retarded maturity of this species. An additional source of information on the distribution of S. scrippsae may be found in past records and studies done on some commercial fisheries. For example, Le Brasseur (1959) found that salmon catches in the northeast Pacific off Canada were inversely related to the oceanic flow as characterized by the occurrence of S. scrippsae (identified as S. lyra) into that region. This change in salmon catches, according to Le Brasseur, was found even at locations where change in physico-chemical characteristics was not detectable. It should also be noted that high catches of salmon in the northeastern Pacific coincide with low or nil catches of albacore, Thunnus alalunga (Powel and Peterson 1957). Evidence of positive correlation was observed between the distribution and abundance of S. scrippsae off North America and the albacore in these waters (author's unpublished date, Albacore Cruise 63, AO-7, June-July, 1971, R/V DAVID STARR JORDAN, NOAA, National Marine Fisheries Service, Southwest Fisheries Center, La Jolla, California). It was further observed, when analyzing the distribution of the longline catches of albacore in the Pacific (Alvariño 1970) that the fish were found in late September and in October in the northeastern part of the Transition region of the North Pacific waters, which might be their main westward migratory route. It has also been observed (Alvariño's unpublished data) that Trachurus symmetricus larvae survive better in waters inhabited by S. scrippsae off California. With data from locations north of the area surveyed during 1969 cruises, it could be possible to. follow the history of the broods of S. scrippsae, rate of growth, survival, interactions with other species, and their transport by the current, which would be of value in tracing the water movement and transport with time off California. The methods used to Predict the spawning season in chaetognaths. include: 1) Determining the number of individuals at early Stage 1, and 2) Comparing the percentage of adults with ripe gonads to the percentage of young. The latter cannot be applied in the present study because the area investigated is only the southernmost extension of the species distributional region. Information obtained during the various seasons in the Transition region include detailed observations on changes in the West Wind Drift flow, and the characteristics of eddies developing into the Central Pacific waters north of Hawaii. Comparison of these with Eber's (1971) hypothesis of standing waves in the current structure and changes in the characteristics and strength of the California Current through the years and seasons should provide necessary additional information. Wyllie (1966) reinforces the mentioned plan of study, implying that "the variation in strength of the California Current is probably controlled by the strength and direction of the West Wind Drift at the time it is influenced by the presence of the continent of North America." That information could provide answers and the clues needed to foretell the characteristics of the California Current, based on data collected in key location during the winter or early spring in the Transition region. This prognosis could be used to determine characteristics of fisheries in the North American Pacific. Sagitta scrippsae could be used to rapidly detect the presence of the Transition waters off the California coast and the degree of mixing of those waters. This species can be identified easily in a plankton sample with the naked eye if necessary, even by a nonspecialist, after a short training period. AgradecimientosI would like to honor the memory of Dr. Elbert H. Ahistrom by acknowledging my gratitude to him for the constructive discussion and advise on these studies. My heartfelt appreciation to Drs. Edward Brinton, Frank Ferrari, John R. Hunter, and to Mr. Richard Schwarzlose, for reading the manuscript and their valuable sugestions and encouragement. My thanks are also due to Shari Sitko for the great help provided in the editing of the manuscript, and Roy M. Allen for the final drawings of the maps and figures. LITERATURAALVARIÑO, A. Two new Pacific Chaetognatha; their distribution and relationships to allied species. Bull. Scripps Inst. Oceanogr., Univ. Calif. 1962 1-50 8(1) ---, Zoogeography of Chaetognatha, especially in the California region (Zoogeografía de los Quetognatos, especialmente de la región de California). Ciencia, Also, Contrib. Scripps Inst. Ocean., Univ. Calif., 34(1705):1677-1702. 1964 51-74 32(2) ---, Distributional atlas of Chaetognatha in the California Current region. CaICOFI monthly cruises of 1954 and 1958. Calif. Coop. Oceanic Fish. Invest. Atlas 1965 a. 1-291 3:I-XIII ---, Chaetognaths. In H. Barnes (ed.) Oceanography and Marine Biology, Annual Review. George Allen and Unwin Ltd. Publ. London 1965 b. 115-194 3 ---, Zoogeography of California: Chaetognatha (Zoogeografía de California: Quetognatos). Also, Contrib. Scripps Inst. Oceanogr., Univ. Calif., 37 (2139):487-531 Rev. Soc. Mex. Hist. Nat. 1966 199-243 27 ---, Bathmetric distribution of Chaetognatha, Siphonophorae, Medusae and Ctenophora off Saki Diego, California. Also, Contrib. Scripps Inst. Oceanogr. Univ. Calif., 37: (2171) :902-913. Pac. Sci. 1967 274-285 21 (4) ---, Studies of the albacore catches in the Pacific. Proc. 21st Tuna Conf., Lake Arrowhead, Calif., October 1970 1970 p. 8-9. ---, Plankton indicators and the oceanography of the California Region (Indicadores planctónicos y la oceanografía de la región de California). Mem. V Congr. Nacl. Oceanogr. México 1977 p. 52-74. BULLOCK, T. H. Compensation for temperature in the metabolism and activity of poikiloterms. Biol. Rev. 1955 311-342 30 CLARKE, G. L., E. L. PIERCE and D. F. BUMPUS The distribution and reproduction of Sagitta elegans on Georges Bank in relation to hydrographical conditions. Biol. Bull. Mar. Biol. Lab., Woods Hole, Mass. 1943 201-226 85 DAVID, P. M. The distribution of Sagitta gezellae RitterZahony. Discovery Rep. 1955 235-278 27 DUNBAR, M. J. On the size distribution and breeding cycles of four marine planktonic animals from the Arctic. J. Anim. Ecol. 1940 215-226 9 ---, The breeding cycle in Sagitta elegans arctica Aurivillius. Canada J. Res. Sect. D. 1941 258-266 19 ---, The life cycle of Sagitta elegans in the Arctic and Subarctic Seas, and the modifying effects of hydrographic differences in the environment. J. Mar. Res. 1962 76-91 20 EBER, L. E. Characteristics of the sea-surface temperature anomalies. U.S. Fish Bull. 1971 345-355 69 (2) HUNTSMAN, A. G. Some quantitative and qualitative plankton studies of the eastern Canadian plankton. Rep. Naval Serv. Canada, Fish. Exped. 1919 p. 405-485. 1914-15 and M. E. REID The success of reproduction of Sagitta elegans in the Bay of Fundy and the Gulf of St. Lawrence. Trans. R. Can. Inst. 1921 99-112 13 KITOU, M. Chaetognatha collected on the eight cruises of the Japanese expedition of Deep Seas. La Mer, Bull. Soc. Francojaponaise d'Oceanographie 1967 33-36 5 (2) KOTORI, M. Vertical distribution of chaetognaths in the northern North Pacific Ocean and Bering Sea. A. Y. Takenoute et al Biology and Oceanography of the Northern North Pacific. 1972 KOTORI, M. The biology of chaetognaths in the Bering Sea and the western North Pacific Ocean, with emphasis on Sagitta elegans. Mar. Fac. Fish. Hokkaido Univ. 1976 95-183 23 (2) and A. HARA On the Chaetognatha in the Bering Sea, with special reference to a new record of Sagitta scrippsae. Bull. Plankton Soc. Jap. 1972 5-12 19 (1) KRAMER, D. and E. H. AHLSTROM Distributional atlas of the fish larvae in the California Current region: Northern anchovy Engraulis mordax Girard, 1951 through 1965. Calif. Coop. Oceanic Fish. Invest. Atlas 1968 1-269 9:I-IX, LE BRASSEUR, R. J. Sagitta lyra a biological indicator species in the Subarctic waters of the eastern Pacific Ocean. J. Fish. Res. Bd. Canada 1959 795-805 16 (6) McLAREN, I. A. Effects of temperature and growth of zooplankton and adaptative value of vertical migration. J. Fish. Res. Bd. Canada 1963 685-727 20 ---, Adaptative significance of large size and long life of the chaetognath S. elegans in the Arctic. Ecology 1966 852-855 47 MURAKAMI, A. Marine biological study on the planktonic chaetognaths in the Seto Inland Sea. Bull. Naikai Reg. Fish. Lab. 1959 1-186 12 NAGASAWA, S. and R. MARUMO Reproduction and life history of the Chaetognatha Sagitta nagae Alvariño in Suruga Bay. Bull. Plankton Soc. Jap. 1978 67-89 25 (1) PIERCE, E. L. On the occurrence and breeding of Sagitta elegans Verrill and Sagitta setosa J. Müller in parts of the Irish Sea. J. Mar. Biol. Assoc. U. K. 1941 113-124 25 POWEL, D. E. and A. E. PETERSON Experimental fishing to determine distribution of salmon in the North Pacific Pacific Ocean, 1955. U.S. Fish Wildl. Serv., Spec. Sci. Rep. Fish. 1957 1-30 205 REDFIELD, A. K. and A. BEALE Factors determining the distribution of population of chaetognaths in the Gulf of Maine. Biol. Bull, Mar. Biol. Lab., Woods Hole, Mass. 1940 459487 79 REEVE, M. R. The biology of Chaetognatha, Quantitative aspects of growth, and egg production in Sagitta hispida. Marine Food chain J. H. Steele 1970 p. 168-189 REID, J. L. Measurements of the California Countercurrent at a depth of 250 meters. J. Mar. Res. 1962 134-137 20 (2) ---, Measurements of the California Countercurrent off Baja California. J. Geophys. Res. 1963 4819-4822 68 RUSSELL, F. S. On the biology of Sagitta. The breeding and growth of Sagitta elegans Verrill in the Plymouth area, 1930-31. J. Mar. Biol. Assoc. U. K. 1932a 131-145 18 ---, On the Biology of Sagitta II. The breeding and growth of Sagitta setosa J. Müller in the Plymouth area, 1930-31, with that of S. elegans Verrm. J. Mar. Biol. Assoc. U. K. 1932b 147-160 18 ---, On the biology of Sagitta III. A further observation on the growth and breeding of Sagitta setosa in the Plymouth area. J. Mar. Biol. Assoc. U. K. 1933a 555-558 18 (2) ---, On the biology of Sagitta. IV. Observations on the natural history of Sagitta elegans Verrill and S. setosa J. Müller in the Plymouth area. J. Mar. Biol. Assoc. U. K. 1933b 559-574 18(2) SAMEOTO, D. Life history, ecological production, and an empirical mathematical model of the population of Sagitta elegans in St. Margaret's Bay, Nova Scotia. J. Fish. Res. Bd. Canada 1971 971-985 28 ---, Annual cycle and production of the chaetognath Sagitta elegans in Bedford Basin. Nova Scotia. J. Fish Res. Bd. Canada 1973 333-344 30 (3) SHERMAN, K. and E. G. SCHANER Observations on the distribution and breeding of Sagitta elegans (Chaetognatha) in coastal waters of the Gulf of Maine. Limnol. Oceanogr. 1968 618-625 13 (4) SPARCK, R. Relation between metabolism and temperature in Marine Lamellibranchs. Danske Vidensbernes Selskab, Copenhahen Biologiske Medd. 1936 1-27 13 UNIVERSITY OF CALIFORNIA (In press) Physical and chemical data. CaICOFI cruises 1969 G. Bull. Scripps Inst. Oceanogr., Univ. Calif. VERNBERG, J. Comparative physiology; latitudinal effects on physiological properties of animal popuIations. Annual Rev. Physiol. 1962 518-546 27 WIMPENNY, R. S. The distribution, breeding and feeding of some important plankton organisms of the Southwest North Sea in 1934. Part 1. Calanus finmarchicus (Gunn.), Sagitta setosa J. Müller and Sagitta elegans Verrill. Minn. Agric. Fish., Fish. Invest. II 1937 1-53 15 (3) WOOSTER, W. S. and J. H. JONES California undercurrent off Baja California. J. Mar. Res. 1970 235-250 28 (2) WYLLIE, J. G. Geostrophic flow of the California Current at the surface and at 200 m. Calif. Coop. Oceanic Fish. Invest. Atlas 1966 1-268 4:I-XIII
|