|
LOS CROMOSOMAS DE DORMITATOR LATIFRONS.
(PISCES: GOBIIDAE)
Trabajo recibido el 22 de marzo de 1982 y aceptado
para su publicación el 24 de septiembre de 1982.
MANUEL URIBE ALCOCER, JULIO ARREGUÍN ESPINOSA, ALEJANDRO TORRES PADILLA y ARTURO CASTRO PÉREZ
Universidad Nacional Autónoma de México, Instituto
de Ciencias del Mar y Limnología. Contribución 230 del Instituto
de Ciencias del Mar y Limnología, UNAM.
Se informa
sobre un estudio realizado sobre el cariotipo de Dormitator
latifrons, pez muy abundante en las lagunas costeras del
Pacífico. Utilizando tejido epitelial de branquias, se obtuvieron campos
mitóticos suficientes para determinar el número diploide, 46, y
el número fundamental, 90, de esta especie. Estos rasgos
cariotípicos y la estructura de los cromosomas se asemejan notoriamente,
si no son idénticos, a los de la especie gemela D.
maculatus que habita en las costas del Golfo. Se discuten estas
características a la luz de sus relaciones filogenéticas y de las
tendencias evolutivas manifestadas en la familia Gobiidae, y la
conclusión principal es que la posición taxonómica de
quienes afirman que el género Dormitator es
monotípico parece ser más acertada, de acuerdo a los rasgos
citogenéticos encontrados.
The karyotype of Dormitator
latifrons, a fish abundant in the coastal lagoons of the Pacific
Coast of Mexico, is reported. Gill epithelium was used to obtain enough mitotic
cells to determine the diploid number, 46, and the fundamental number, 90, of
this species. These karyotypical features and the form. and size of chromosomes
are very similar, if not identical, to those of D.
maculatus, a sibling species inhabiting the Gulf coast. These
characteristics are discussed from the view point of their phylogenetic
relationships and the evolutionary trends shown in the Gobiidae. The main
conclusion is that the taxonomical position which considers the genus
Dormitator as monotypic, seems to be more accurate, in
accordance to our cytogenetical data.
La familia Gobiidae está constituida por numerosos representantes que habitan en las Costas de México. La especie Dormitator latifrons tiene un ámbito de distribución que abarca desde el Golfo de California hasta Perú (Ancieta y Landa, 1977; Miller, 1966 y AmezcuaLinares, 1977) y se presenta particularmente en aguas salobres y corrientes turbias (Miller, 1966). Este pez es sumamente abundante en las lagunas costeras del Pacífico de México, debido a algunas características que le confieren extraordinaria resistencia fisiológica, entre las que se pueden mencionar su capacidad para resistir variaciones notables de salinidad y temperatura, y para sobrevivir en ambientes deficientes de oxígeno (Ancieta y Landa, 1977). En tal situación los especímenes presentan una inflamación en la parte anterosuperior de la cabeza que, al parecer, funciona como órgano de respiración aérea (Todd, 1973) siempre que se encuentre en condiciones adecuadas de humedad. Otro rasgo relacionado con su extraordinaria resistencia es el incremento de la concentración de hemoglobina en ambientes con poca acreación (Graham, 1967), característica frecuente en las lagunas costeras de México. Los organismos de esta especie tienen un papel ecológico importante en las lagunas costeras al convertir la energía de los detritus en formas útiles para organismos de estratos tróficos superiores (Yáñez-Arancibia y Díaz-Gonzalez, 1977). Su abundancia ha propiciado que, en el Estado de Guerrero, se haya establecido una planta procesadora de la carne de Dormitator latifrons, a fin de producir con ella harina de pescado. El objetivo del presente trabajo fue, en primer lugar, establecer el cariotipo de D. latifrons, lo que fue motivado por el hecho de conocer el cariotipo de la especie gemela que habita en las costas del Golfo de México (Maldonado-Monroy, 1981 y Uribe Alcocer, et al., 1979), y mediante ello aportar criterios para la correcta ubicación taxonómica de la especie, puesto que según varios autores, entre ellos Jordan y Evermann (1898), Jordan, Evermann y Clark (1930) y Castro-Aguirre (1978), los peces de las poblaciones de ambas costas del continente deben ser incluidas en una misma especie: Dormitator maculatus o bien Dormitator latifrons (Yáñez-Arancibia, 1977), mientras que otros especialistas, entre ellos Álvarez del Villar (1970), Yáñez-Arancibia (1978) y Miller (1976) insisten en la validez de la especie presente en el Pacífico Mexicano. MATERIAL Y MÉTODOSLos ejemplares estudiados fueron dos machos y cinco hembras, colectados en las inmediaciones de Agua Verde, Sin., en dos lagunas formadas por brazos temporales del Río Baluarte: Lagunas El Marino y El Pozo, durante los meses de junio y agosto de 1980. Los ejemplares fueron colectados por metodos artesanales y transportados a las instalaciones de la Estación Mazatlán del Instituto de Ciencias del Mar y Limnología, UNAM, en dicho lugar con objeto de llevar a cabo la fase inicial de las técnicas cromosómicas. Esto consistió en la elaboración de dispersiones cromosómicas, de acuerdo al siguiente procedimiento: a) Se inyectó a los especímenes una solución de cloruro de calcio a una concentración de 0.1% A los peces cuya longitud total era de 5 a 10 cm se les aplicó 0.5 cc; a los de 10-15 cm, 0.75 cc, y a los de 15-10 cm, 1 cc.b) Tres horas después se les inyectó una solución de colchicina al 0.1% en una proporción de 0.1 ml/10 g de peso corporal. c) Tres horas después se procedió a la extirpación de las branquias, las que fueroncolocadas en solución hipotónica de cloruro, de Potasio a una concentración de 0.75 M a 37ºC. d) A fin de producir una descamación del epitelio branquial, se raspó con un bisturí el material celular, y una vez desprendido fue mantenido en suspensión en la solución hipotónica durante media hora. e) Se separaron las células de la solución hipotónica mediante centrifugación y decantación del sobrenadante. f) Se procedió a la fijación del material celular utilizando metanolacetico (3:1), como medio para la resuspensión del botón celular. g) Se separó nuevamente el material celular mediante centrifugacion. h) Este último paso se repitió, cuando menos dos veces mas, a fin de obtener una fijación completa. i) Se goteo la suspensión celular sobre portaobjetos previamente limpiados con una mezcla de xileno-éter (1:1) y se secaron al aire. En las instalaciones del Laboratorio de Genética del Instituto de Ciencias del Mar y Limnología, situadas en la Ciudad Universitaria, D. F., se continuó el proceso de acuerdo a los siguientes pasos: a) Se tiñieron las preparaciones en una solución de Giemsa, en buffer de fosfatos pH 6.8. Algunas preparaciones fueron teñidas con aceto, carmín, aunque los resultados de estas tinciones fueron inferiores a las logradas con Giemsa. Posteriormente fueron montadas las preparaciones con resina sintética. b) Se realizó un examen microscópico de las preparaciones a fin de determinar el número diploide del organismo, así como para buscar campos microscópicos cuyos cromosomas fueran adecuados al análisis cariotípico. c) Se fotografiaron los mejores campos utilizando un microscopio Zeiss con aditamentos para la microfotografía, rollo "kodak" de alto contraste HC 16. El revelado de este rollo y su impresión en papel Kodabromide F 5 fueron realizados de acuerdo a las técnicas convencionales. d) Se escogieron las amplificaciones mejor logradas a fin de elaborar los cariotipos. Se hicieron por lo menos tres de cada especimen procesado. RESULTADOSEl número modal en las cuentas cromosómicas fue de 46, por lo cual se puede considerar este número como número diploide. Este mismo número fue encontrado igualmente en campos mitóticos aislados y con cromosomas bien definidos. El número fundamental (número total de brazos cromosómicos) es de 90. Se midieron los cromosomas de 12 cariotipos, 8 provenientes de hembras y 4 de machos y siguiendo los criterios de Levan et al. (1964) se clasificaron los cromosomas según la posición del centrómero: De los 46 cromosomas doce son metacéntricos, veintidós son submetacéntricos diez subtelocéntricos y un solo par acrocéntrico (Tabla l). No se encontró evidencia de heterocromosomas en ningún sexo. La figura 1 muestra un campo mitótico y un cariotipo representativo de los especímenes estudiados. 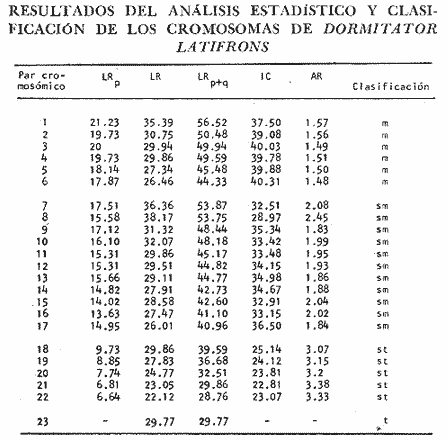 TABLA 1. RESULTADOS DEL ANÁLISIS ESTADÍSTICOS Y CLASIFICACIÓN DE LOS CROMOSOMAS DE DORMITATOR LATIFRONS DISCUSIÓNLos números diploides encontrados en las especies del género Dormitator coinciden en aspectos importantes: tienen el mismo número diploide: 46 la morfología y tamaño de los cromosomas son notoriamente semejantes, si no idénticos. Estas características compartidas pueden ser más fácilmente explicadas por su presencia en el antecesor común que por ser rasgos producto de evolución convergente. Basados en estos datos, citogenéticos, parece ser más adecuada la posición de los autores que sostienen que las poblaciones del género Dormitator de las costas del Golfo y del Pacífico deben ser incluidas dentro de una sola especie, ya que son poblaciones que comparten un cariotipo idéntico o extremadamente similar. De demostrarse que existe efectivamente un aislamiento reproductivo distinto del geográfico, se debera probablemente buscar en mecanismos distintos de los citológicos. 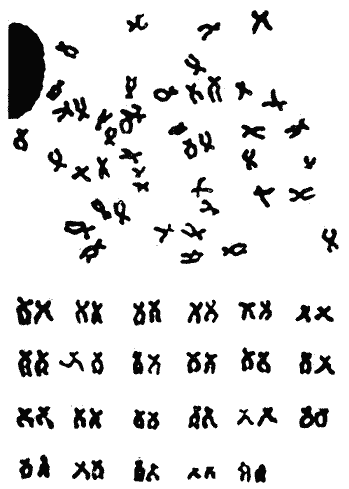 Fig 1. Campo mitótico y cariotipo representativo de los especímenes pertenecientes a la especie Dormitator latifrons. Existe la posibilidad de verificar con mayor exactitud la identidad o extremado parecido de los cariotipos de las poblaciones de ambas costas, mediante el estudio de las bandas cromosomicas "G" presentes en dichos cariotipos. Es muy probable que, además, se puedan llegar a detectar divergencias microevolutivas a través del establecimiento de los patrones electroforé de diversas proteínas de ambas poblaciones. Estos estudios permitirán detectar diferencias electroforéticas resultantes de la modificación de las secuencias de aminoácidos que constituyen las proteínas estudiadas, que cambien el punto isoeléctrico de las mismas y que por no llegar a alterar su función hacen difícil su detección por otros métodos. En la tabla 2, se presenta un cuadro de especies de la familia Gobiidae, cuyos cariotipos son conocidos. Cook (1978) afirma que las cuatro primeras forman una entidad natural de evolución cuyo grupo ancestral identifica como Gillichtys mirabilis. Propone que a partir del cariotipo de este organismo se haya derivado el de Quietula y-cauda, involucrando en el proceso un número muy elevado de reacomodos cromosómicos: 4 fusiones céntricas seguidas de 14 inversiones pericentricas o bien cuatro fusiones en tandem seguidas de doce inversiones pericentricas. Propone asimismo que el cariotipo de Q. y-cauda se haya originado el de Q. guaymasie a través de 10 inversiones pericéntricas. Chen y Ebeling (1971), por su parte han propuesto que el cariotipo de G. mirabilis, o uno muy semejante, es el ancestral de G. seta, al que se llego por medio de 4 inversiones pericéntricas. Cook propone que el fenómeno geológico de las glaciaciones fue el factor que favoreció el establecimiento de los distintos aislamientos requeridos para la fijación de los diversos cariotipos. Las ilustraciones presentadas en los respectivos informes, al parecer no sustentan las opiniones de los autores antes mencionados, puesto que el cariotipo de G. mirabilis aparentemente está formado por únicamente seis cromosomas subtelocéntricos y 38 acrocéntricos, de acuerdo a la clasificación de los cromosomas propuesta por el propio Chen (1967). 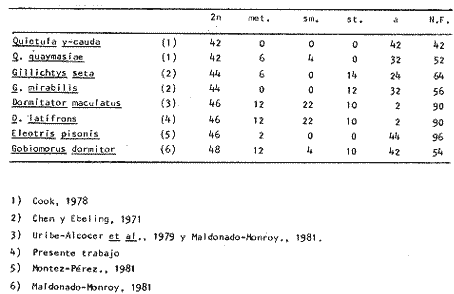 TABLA 2. MORFOTIPOS CROMOSÓMICOS DE PECES DE LA FAMILIA GOBIIDAE Esta interpretación del cariotipo de G. mirabilis simplifica grandemente el esquema evolutivo del grupo en cuestión, y presenta un cuadro más acorde a los esquemas evolutivos cariotípicos conocidos. De esta manera las modificaciones del cariotipo de G. mirabilis al de Q. y-cauda podrían ser una fusión céntrica y 4 inversiones pericéntricas. Esta interpretación no alteraría lo propuesto por Cook para explicar la derivación de Q. guaymasiae a partir de Q. y-cauda. Los cinco eventos mutacionales invocados para convertir el cariotipo de Q. y-cauda al de Q. quaymasiae, implicarían un mecanismo de aislamiento reforzado, que evoca el caso de distintas poblaciones de Peromyscus maniculatus y P. melanotis, en el que poblaciones que habitan regiones vecinas tienen fuertes mecanismos de aislamiento que evitan su hibridación. Estos mecanismos de aislamiento son mas laxos en poblaciones que habitan regiones lejanas, y experimentalmente se han podido establecer cruzas entre ambas especies (Bowers, et al, 1973). Es necesario apuntar que en los cuadros evolutivos mencionados por Chen y Ebeling (1971) y Chen (1967) no se requiere de dos mutaciones cromosómicas idénticas afectando a los dos miembros de un par de homólogos para explicar la modificación de un par de cromosomas homólogos de un cariotipo a los de otro. Es más bien necesario invocar un solo evento mutacional y su paso a estado homocigoto en el seno de una población. Para poder explicar este paso a estado homocigoto es necesario invocar una serie de circunstancias, acompañantes que determinen o cooperen a la extensión del reacomodo cromosómico en la población, tales como pudieran ser una modificación espacial de los genes que alterara sus interrelaciones y produjera cambios en los procesos regulatorios o bien introdujeran efectos de posición entre los genes involucrados en los reacomodos cromosómicos. Pudieran también influir en el establecimiento de un reacomodo cromosómico su aparición junto a una caracteristica adaptativa importante no dependiente directamente del reacomodo cromosómico, que determinara que fuera sujeto de selección positiva conjunta. Otra circunstancia coadyuvante podría ser la reducción del tamaño de la población y/o una fuerte endogamia en el se de ella. Dadas las circunstancias anteriores, o algunas de ellas, un evento mutacional cromosómico podría establecerse homocigóticamente en una población y despues expanderse a otras poblaciones de la misma especie. Por otra parte el hecho de invocar dos eventos mutacionales idénticos, implica que dos eventos de probabilidad independiente sumamente baja se han dado en forma casi simultánea lo que es muy dificil de aceptar lógicamente, existiendo explicaciones alternativas más satisfactorias (Wilson et al., 1975). Por lo que respecta a las especies del Género Dormitator cabe afirmar que forman una línea evolutiva diferente a la formada por el grupo Gillichtys-Quietula, puesto que el ámbito de distribución de aquél es mucho más extenso abarcando gran porción de ambas costas del continente americano, y el hecho de compartir gran cantidad de rasgos fenotipicos, entre ellos el cariotipo, implica que desde antes que pudiera establecerse divergencias locales, el genero ya existía y estaba abundantemente representado cuando ambos océanos estaban aun conectados. El número de cromosomas birrameo es importante en este género, puesto que es indicativo de un modo de evolución particular, ya que las pautas de evolución propuestas para la familia de los Gobiidae (Chen, 1967; Ohno et al., 1968; Chen y Ebeling, 1971) no son seguidas por el Género Dormitator. Éste presenta un número cercano al número primitivo: 46, y al mismo tiempo, presenta rasgos de adelanto evolutivo en el número de cromosomas birrámeos, que son casi su totalidad. A fin de analizar las relaciones de evolución cromosómica con mayor exactitud existen posibilidades que deberán ser usadas en futuros estudios. Tales posibilidades están dadas por las bandas cromosomicas G y C que pueden ser utilizadas como marcadoras de los cromosomas o de regiones cromosómicas. Al presente estas tecnicas, por razones desconocidas, raramente han podido ser aplicadas con éxito a cromosomas provenientes de epitelio branquial de peces, fuente de las células estudiadas en el presente trabajo. Conclusiones1. Existe una concordancia, si no identidad, entre los cariotipos de las dos especies del género Dormitator, en cuanto a su número diploide, 46, y en cuanto a la estructura del cariotipo: 12 m, 20 sm, 12 st y 2 a. Esta similitud, o identidad, apoya la opinión de que el género Dormitator es monotípico. 2. Las especies estudiadas forman un grupo evolutivo distinto del propuesto por Chen y Ebeling (1971) por una parte, y por Cook (1978), por otra. 3. Se deben obtener bandas cromosómicas "G" y "C" de estos grupos a fin de tener una visión más precisa de los grados de relación existente entre los cromosomas de las especies de Dormitator. Se deben efectuar asimismo estudios de los perfiles electroforéticos de algunas proteínas de los organismos en cuestión, a fin de precisar las relaciones existentes entre estos grupos. LITERATURAÁLVAREZ DEL V. J. Peces mexicanos (Claves). Inst. Nal. Inv. Biol. Pesq., Com. Nal. Consultiv. Pesca 1970 156 p. 62 figs. ANCIETA, D. F. y A. LANDA C. Reseña taxonómica y biológica de los peces cultivados en el área andina incluyendo la costa del Perú. FAO Inf. Pesca 1977 106-113 159 (2) AMEZCUA-LINARES, F. Generalidades ictiológicas del sistema lagunar costero de Huizache-Caimanero, Sinaloa, México. An. Centro Cienc. del Mar y Limnol. Univ. Nal. Auton. México 1977 1-26 1 (1) BOWERS, J., R. J. BAKER Y M. SMITH Chromosomal, electrophoretic and breeding studies of selected populations of deer mice (Peromyscus maniculatus) and black-eared mice (P. melanotis) Evolution 1973 378-386 27 (3) CASTRO-AGUIRRE, J. L. Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México con aspectos zoogeográficos y ecológicos. Serie Científica Dir. Gral. Inst. Nal. Pesca, Depto. de Pesca, México 1978 1-298 19 COOK, P. C. Karyotypic analysis of the gobiid fish genus Quietula (Jordán y Evermann). J. Fish. Biol. 1978 173-179 (2) CHEN, T. R. Comparative karyology of selected deep-sea and shallowwater teleost fishes. Ph. D. Dissertation Yale University. 1967 CHEN, T. R. y EBELING, A. U. Chromosomes of the goby fishes in the genus Gillichthys. Copeia 1971 171-174 (1) GRAHAM, J. B. Hemoglobin concentrations of air breathing fishes Am. Zool. 1976 192 16 (2) JORDAN, D. S. y B. W. EVERMANN The fishes of north and middle America. Bull. U.S. Nat. Mus. 1898 2196 17 (3) LEVAN, A., K. FREDGA y A. SANDBERG Nomenclature for Centroameric position on chromosomes. Hereditas 1964 201-220 52 MALDONADO-MONROY, M. del C. Tesis profesional Estudio Cariotipico de Dormitator maculatus Bloch y Gobiomorus dormitor Lacepede (Gobiidae, Pisces: Perciformes). Fac. de Ciencias. Univ. Nal. Autom. México. 1981 MILLER, R. R. Geographical distribution of central American freshwater fishes. Copeia 1966 (4) 773-802 ---, An evaluation of Seth E. Meek's contribution Mexican ichthyology. Fieldiana Zoology Field Mus. Nat. Hist. 1976 131 69 (1) MONTES-PÉREZ, R. Tesis profesional Estudios citogenéticos en Eleotris pisonis (Gobiidae-Perciformes). Facultad de Ciencias. Univ. Nal. Autom. México. 1981 OHNO, S., U. WOLF y N. B. ATKIN Evolution from fish to mammals by gene duplication. Hereditas 1968 169-187 59 TODD, E. S. Positive buoyancy and air breathing: a new piscine gas bladder function. Copeia 1973 461-464 (3) URIBE-ALCOCER, M., M. DEL C.. MALDONADOMONROY, A. CASTRO-PÉREZ y A. LAGUARDAFIGUERAS Estudios cromosómicos en Dormitator maculatus (Gobiidae, Perciformes). Actas IV Congreso Latinoamericano de Getética, Mendoza, Argentina, 1:28. 1979 WILSON, A. C., BUSH, G. L., CASE, S. M. y KING, M. C. Social structuring of mammalian populations and rate of chromosomal evolution. Proc. Natl. Acad. Sci. USA 1975 5061-5065 72 (12) YÁÑEZ-ARNCIBIA, A. Piscicultura en lagunas costeras: perspectivas en México. FAO Fish. Rep. 1977 529-547 200 YÁÑEZ-ARANCIBIA, A. Taxonomía, ecología y estructura de las comunidades de peces en lagunas costeras con bocas efímeras del Pacífico de México. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Publ. Esp. 1978 1-306 2 YÁÑEZ-ARANCIBIA, A. y G. DÍAZ-GONZÁLEZ Ecología trofodinámica de Dormitator latifrons (Richardson) en nueve lagunas costeras del Pacífico de México (Pisces: Eleotridae). An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México 1977 125-140 4 (1)
|