|
CONTRIBUCIÓN AL CONOCIMIENTO DE LOS ANÉLIDOS POLIQUETOS
DE LAS PRADERAS DE THALASSIA TESTUDINUM Y HALODULE SP DEL SECTOR DE PUNTA
GALETA (PANAMÁ, PROVINCIA DE COLÓN)
Trabajo recibido el 6 de
noviembre de 1981 y aceptado para su publicación el 18 de agosto de
1982. Centro de Ciencias del Mar y Limnología.
RAFAEL VÁSQUEZ MONTOYA
Universidad de Panamá.
BERNARD THOMASSIN
Station
Marine d'Endoume, Marsella, Francia.
Dos praderas
de fanerógamas (Thalassia testudinum y
Halodule sp) en Punta Galeta (Colón, República
de Panamá), fueron muestreadas con un cilindro para sedimento. Dos
grupos de anélidos poliquetos, cada uno perteneciente a una pradera
diferente, fueron diferenciados por sus índices de afinidad, coeficiente
de rangos Spearman y coeficiente de Jaccard (porcentaje de presencia-ausencia);
las características del 51% de la población global
demostró su pertenencia a la fauna de los substratos arenosofangosos,
comparables con las biocenosis infralitorales de tipo "sedimento fangoso de
aguas tranquilas". El 27% de la población global demostró tener
afinidad con la fauna de substratos duros y comparables.
Une
étude écologique du peuplement annélidien des herbiers de
Thalassia Testudinum et d'Halodule du
secteur de Punta Galeta (coté caraïbe de la Réplublique de
Panama) est presentée. La liste faunistique, et une analyse du
rôle des principaux facteurs édaphiques dans la distribution des
espèces, sont données. La faune des armélides
observées dans l'herbier de Thalassia est différente et moins
abondante que celle d'Halodule) phanérogame qui croît dans
l'étage infralittoral (aux niveaux supérierurs) et qui
présente, d'aprés le coefficient d'affinité, et de
I'indice de diversité de Shannon Weaver, un degré dequilibre et
de structuration supérieurs.
Este estudio, basado en muestras recogidas en una zona cercana a Punta Galeta (República de Panamá), pretende contribuir al conocimiento actual sobre la fauna de anélidos de las praderas de Thalassia testudinum y de Halodule sp., fanerógamas que, en el Caribe "colonizan" el substrato, y el sedimento, y llegan a constituir la especie dominante de un biotopo que en consecuencia es muy productivo y se caracteriza por la presencia de numerosas especies que allí encuentran su habitat. SITUACIÓN Y CARACTERÍSTICAS DE LAS PRADERAS ESTUDIADASUna pradera dominada por Thalassia testudinum fue localizada dos kilómetros al Sur del Smithsonian Tropical Research Institute (STRI) de Punta Galeta (9°23´2"N;79°50´W), precediendo los manglares (Fig. 1) . Se encuentra sobre una plataforma de pendiente ligera en una zona protegida de los vientos y de las olas por los manglares litorales y por las formaciones arrecifales. Estas formaciones no han alcanzado aún el estado de arrecife barrera propiamente dicho, lo cual no permite considerar la depresión, donde se encontró la pradera, como una laguna post arrecifal sensu stricto sino más bien como un canal de embarcación.
La pradera de Halodule sp. está situada en una laguna post arrecifal de poca profundidad a proximidad del laboratorio (9°23´9"N;79°51´8"W). Está dividida en dos sectores: uno limitado por el Sur por el dique artificial que lleva al laboratorio y al Oeste, por los manglares y una pequeña playa arenosa; el otro sector limita al Norte con el mismo dique (que le sirve de protección) y, al Sur, con los manglares. Ambos están bordeados por una franja de Thalassia que casi no se mezcla con la pradera de Halodule. Es frecuente encontrar fragmentos de Acanthophora (Rodophyta) entre los rizomas de la pradera lo cual, como en el caso de la pradera de Thalassia donde se encuentran pedazos de Porites, hace el muestreo más difícil. Estos fragmentos de Acanthophora provienen de la zona frontal del arrecife, colonizada por esta alga. [1] Es conveniente señalar que la terminología utilizada es la recomendada por Elementos de Terminología Arrecifal (Battistini [2] Estos fragmentos de madera correspondían a despojos de antiguos manglares MATERIALES Y MÉTODOS
MUESTRAS DE INFAUNA10 estaciones (cada una aportando 38.2 dm3 de sedimento) en zonas homogéneas fueron muestreadas con un cilindro de 15.5 cm diámetro y 22.5 cm altura introducido en el substrato. En cada tipo de pradera un total de 382.10 dm3 fueron recogidos. Este volumen fue considerado arbitrariamente puesto que los escasos trabajos sobre las infaunas de sedimentos realizados anteriormente en la región del Caribe y del Golfo de México no definen un volumen mínimo (sensu Pérès y Picard, 1964) para estas praderas tropicales.
Las muestras fueron tamizadas sobre una malla cuadrada de 1.8 mm de lado, es decir, 2.5 mm de diagonal. Los organismos fueron fijados en solución de formaldehído al 10%, e identificados posteriormente. [3] O'Gower y Wacasey (1967) reunieron 100 muestras de 3.9 dm ESTIMADO DEL PESO EN CENIZAS DE LAS FANERÓGAMASA fin de estimar el peso en cenizas y el porcentaje de la materia orgánica vegetal, se escogió algunos rizomas y hojas de Thalassia y de Halodule, los cuales fueron cuidadosamente lavados para eliminar el sedimento. Su peso fresco y su peso seco (luego de permanencia en el secador a 65°C durante 24 horas) fueron anotados; las muestras (2 g cada una) fueron posteriormente incineradas a 560ºC durante dos horas y finalmente pesadas. 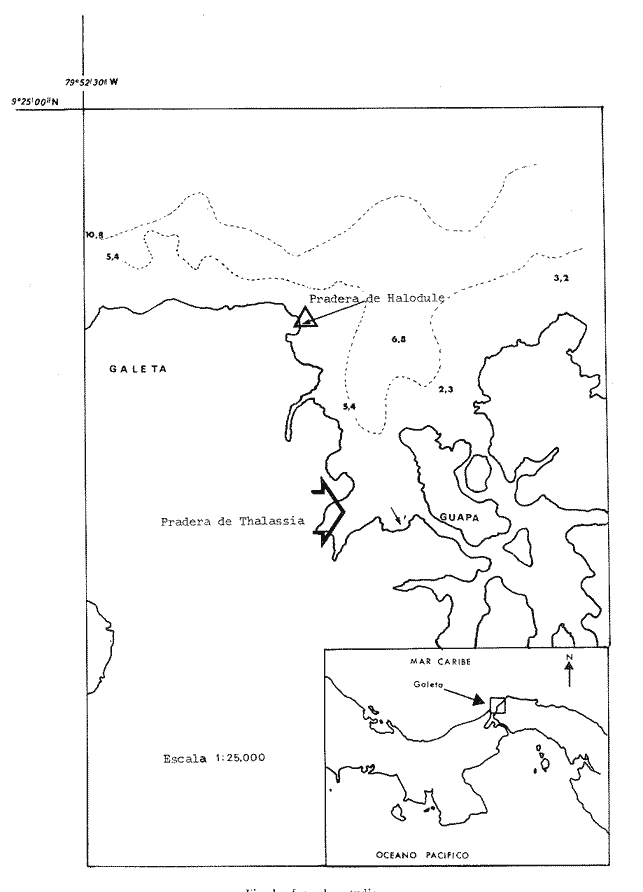 Fig. 1 Área de estudio CARACTERÍSTICAS DEL SEDIMENTOSe hizo igualmente una estimación de la granulometría de las dos praderas sobre una columna de tamices AFNOR. Las muestras secadas en la estufa fueron luego pesadas y lavadas sobre el tamiz de 63 μ, con el fin de recuperar la fracción fina (63 11μ) (arcilla-limo); secadas nuevamente en la estufa y pasada por la columna de tamices. 250 g de la fracción fina fueron utilizadas para un análisis de, calcimetría Bernard. Las muestras para el estudio granulométrico fueron escogidas, no según la estación estudiada, sino en función del tipo de substrato observado. En la pradera de Thalassia la presencia de más de una zona de sedimentos diferentes fue evidente. En la pradera de Halodule se tomaron muestras de las dos partes diferentemente expuestas al hidrodinamismo con el propósito de determinar si existía una diferencia significativa. TRATAMIENTO MATEMÁTICOEl coeficiente de correlación de punto 0 modificado (Sokal y Sneath, 1963), basado sobre el criterio de presencia-ausencia de las especies en cada una de las estaciones fue utilizado con el objeto de cuantificar la afinidad entre las especies y las estaciones. Los resultados de este análisis están representados bajo la forma de dendrograma (Cuadro l). Como la matriz de datos presentó muchas clases expeditas se empleó el coeficiente de correlación de Spearman (r) para corroborar la relación existente entre las estaciones de las dos praderas juntas. El índice de diversidad Shannon-Weaver (H) fue aplicado en el estudio de la estructura de la población. Estos resultados son presentados juntos con el índice de Pieles y el coeficiente de Jaccard al tratar la fisonomía y la estructuración de la población. 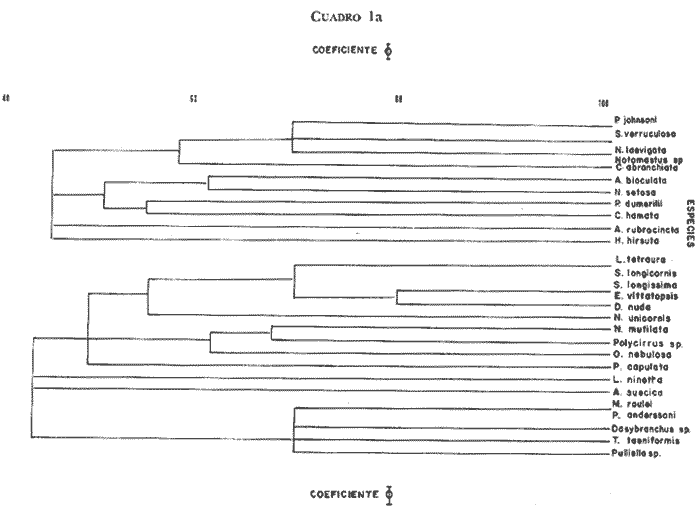 Cuadro 1a 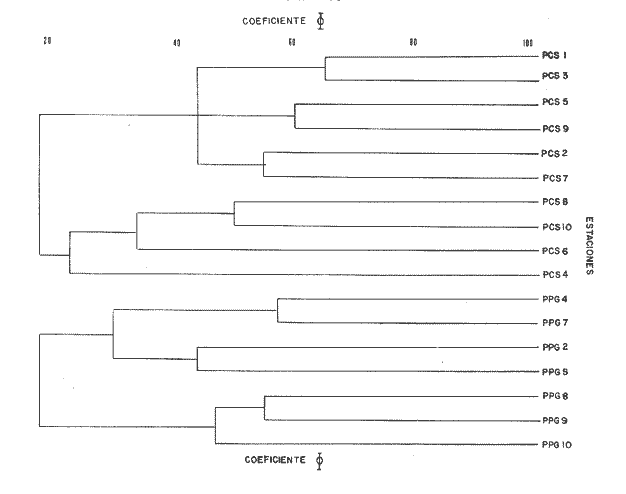 Cuadro 1b [3] O'Gower y Wacasey (1967) reunieron 100 muestras de 3.9 dm RESULTADOS Y DISCUSIÓN
1. CARACTERÍSTICAS DE LOS SEDIMENTOS Y BIOMASA DE LAS FANERÓGAMASLas muestras para el estudio granulométrico provienen de las dos praderas estudiadas y fueron escogidas en función del tipo de substrato observado y de la fanerógama que lo recubría. En cuanto a los análisis de carbonatos y cenizas, éstos se efectuaron de muestras de las fanerógamas de las praderas en cuestión. Los resultados discutidos a continuación se presentan en el cuadro 2. Granulometría: Existe una diferencia entre IGS valores granulométricos de algunos grupos de muestras. Esto se debe a la presencia de importantes fragmentos de coral o mangle, así como también a diversos grados de exposición al hidrodinamismo (e.g. fracción gruesa más importante en estaciones situadas cerca del límite externo de la pradera hacia el arrecife). Carbonatos: Se encontró que el contenido global de carbonatos es, en general, bastante elevado, lo cual demuestra la importancia de los organismos productores de carbonato en este medio próximo al arrecife, toda vez que la orientación de las corrientes, del arrecife hacia la costa, favorece el aporte de los carbonatos. También se nota la importancia del lavado costero en las muestras que presentan características terrígenas, (generalmente menores de 40% de carbonatos). Cenizas: El resultado de los análisis de las plantas (porcentaje ponderal de cenizas) indican que el peso de la materia viviente en las muestras de Thalassia testudinum es ligeramente superior al paso de la materia viviente en las muestras de Halodule sp. Estos valores son más elevados que aquellos reportados por D'Croz et al. (1977) sobre las muestras de Thalassia recogidas al Este de Punta Galeta. Esto es posiblemente debido a que sus muestras comprendían las hojas y frondas de las plantas y no los rizomas. 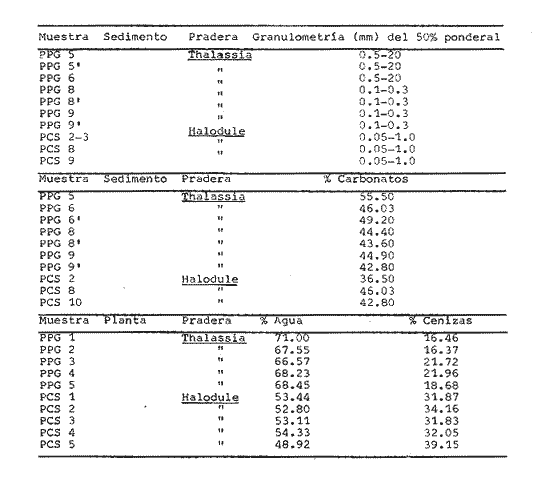 Cuadro 2 2. COMPOSICIÓN DE LA POBLACIÓNLas muestras para el estudio faunístico fueron recogidas en las praderas de Thalassia testudinum y de Halodule sp de Punta Galeta. Los anélidos poliquetos encontrados se presentan, en el cuadro 3, bajo la forma de frecuencia por estación. Estos resultados indican que, según los grados de presencia y de constancia, la fauna de las praderas de Thalassia testudinum está caracterizada por: Marphysa depressa, Onuphis nebulosa, Axiothella rubrocincta, Nereis riseii y Arabella mutans como especies constantes (C > 50%)
Con referencia al mismo criterio, la fauna de la pradera de Halodule sp, está caracterizada por: Malacoceros indicus, Arabella mutans, Isolda pulchella, Terebellides stroemi, Scolopos armiger y Nereis riseii como especies constantes; Armandia bioculata y Axiothella rubrocincta como especies accesorias. Malacoceros indicus y Terebellides stroemi no fueron encontradas en la pradera de Thalassia testudinum. [4] c = grado de constancia (%,). 3. CARACTERÍSTICAS DE LA POBLACIÓNSegún los resultados de la prueba ø (tomando 95% como restricción, a la altura del coeficiente 0.44) y aquellos del cálculo de correlación Spearman, dos grupos de afinidad cierta se destacan. El primero (Cuadro 1) está compuesto por las siguientes especies por orden de afinidad: Poecilochaetus johnsoni, Sthenelais verruculosa, Naineris laevigata, Notomatsus sp, Glycera abranchiata, Armandia bioculata, Naineris setosa, Platynereis sp af. dumerilii, Caulleriella hamata, Axiothella rubrocincta y Harmothoe hirsuta. El segundo grupo (Cuadro 1) está constituido por: Lumbrineris tetraura, Schistomeringos longicornis, Syllis longissima, Eunice vittatopsis, Drilonereis nuda, Nematonereis unicornis, Naineris mutilata, Polvcirrus sp, Onuphis nebulosa, Pherusa capulata Lysidice ninelta, Aricidea suecica, Megalomma roulei, Perinereis sp af. anderssoni, Dasybranchus sp, TrypanosyIlis taeniformis y Pulliella sp. Además tres especies están más o menos bien correlacionadas entre ellas: Isolda pulchella, Terebellides stroemi y Malacoceros indicus. 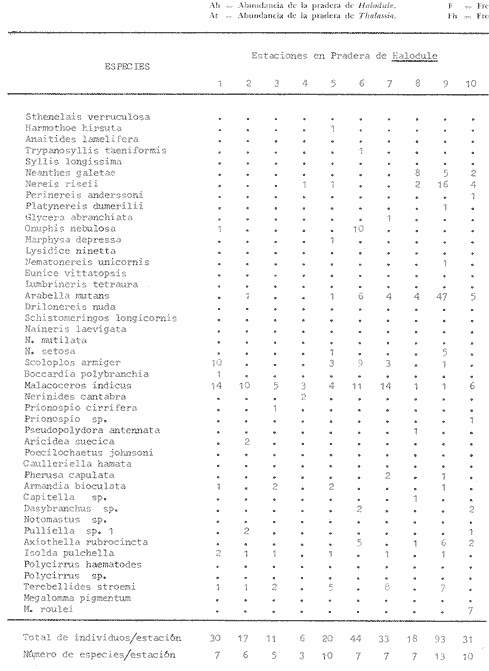 CUADRO 3 LISTA FAUNIÍSTICA (EN ESTUDIO /0,038M3/0,17M2)  CONTINUACION CUADRO 3 Sin embargo se observa que estos dos grupos principales presentan un grado de afinidad más elevado (Cuadros 1, 3) en el seno de la pradera de Thalassia, en las estaciones más alejadas de la orilla. El tercer grupo está localizado en la estación 10 de la pradera de Halodule, la cual es la estación mas expuesta y, al mismo tiempo, la más cercana a la laguna post arrecifal donde aún crecen porciones aisladas de Thalassia. Al comparar la infauna de la pradera de Thalassia testudinum con aquella de Halodule sp, según los resultados de la prueba * y del coeficiente de correlación de Spearman, parece que las poblaciones de las diferentes praderas son cualitativamente distintas. Sólo 17 especies de las 42 halladas en total son comunes a los dos tipos de praderas, es decir, un solo grado de afinidad (según el coeficiente de Jaccard) de sólo 40%. 4. ESTRUCTURA DE LAS POBLACIONESEn la pradera de Thalassia testudinum la población anelidiana varía entre 9 y 21 indviduos /38.2 dm³/0.17m² con una densidad media de 15.2 ± 20.18 individuos/38.2 dm³/O. 17 m². Las especies dominantes, por orden decreciente, son Onuphis nebulosa (2.6 individuos/ 0.17m²; Dr. = 17%,).
En la pradera de Halodule sp, la densidad de los poliquetos es más elevada; ella varía de 6 a 93 individuos/38.2 dm3/0.17m² con una mediana de 30.3 -4- 24.77 individuos/38.2 m³/ 0.l7m². Las especies dominantes son Malococeros indicus (6.9 individuos, Dr = 22.4%), y Arabella mutans (6.8 individuos; Dr = 22.4%), presentando aspectos sobresalientes de la población, aspectos que parecen reemplazarse; están seguidos por Scoloplos armiger, Terebellides stroemi y Nereis riseii (abundante al mismo tiempo que Arabella mutans). La riqueza específica (Sr) varía de 4 a 10 especies /38.2m³/O. 17m², con una mediana de Sr = 7.7 +=L 2.16, en los sedimentos de la pradera de Thalassia testudinum. Ella es ligeramente diferente en la pradera de Halodule sp, de 3 a 13 especies/38.2m³/ O.17m²; mediana de Sr = 7.5 ± 2.84. Las familias mejor representadas son: Onuphidae (17%) Ariciidae (16.5%), Nereidae (15%) y Arabellidae (9%) en los sedimentos de la pradera de Thalassia; contraria mente, son los, Spionidae (25%), Arabellidae (22%), Nereidae (13.5%), Aricúdae (10.5%), Trichobranchidae (8%) y los Onuphidae (4%) los mejores representados en la pradera de Halodule. Para el conjunto de estos dos tipos de praderas las dominaciones más fuertes están representadas por los Arabellidae (18%), los Spionidae (16%), los Nereidae (14%) los Ariciidae (12%), los Onuphidae (8%) y los Trichobranchidae (5%). Desde un punto de vista global, aproximadamente el mismo número de especies ha sido encontrado en las dos praderas (30 spp en la pradera de Thalassia, 29 spp en la pradera de Halodule). No obstante, desde el punto de vista de la estructura de las poblaciones, la infauna de la pradera de Thalassia aparece como ligeramente más estructurada (H` = 4.137 bits/ind),
[5] Dr = Dominancia relativa media [6] H = índice de diversidad de Shannon-Weaver 5. OBSERVACIONES SOBRE SUCESIÓN DE COMUNIDADESEn cuanto a los aspectos de sucesión de comunidades (sensu stricto), la información aquí presentada, referente a las características fisionómicas de la población anelidiana, y tomando en cuenta datos análogos anteriores (Vasquez, 1979) sobre la fauna general de las praderas, sefialan que la pradera de Halodule sp. no puede ser considerada como una comunidad, sino como una fase transitoria que con frecuencia tiende hacia la pradera de Thalassia testudinum, comunidad en donde la angiosperma constituye un aspecto de epiflora y la especie dominante. Esta pradera de Thalassia, tal como fue contemplado por Welch (1972), puede ser un escalón más en la evolución de los sedimentos coralinos hacia la serie terrestre o climax (Cuadro 4). 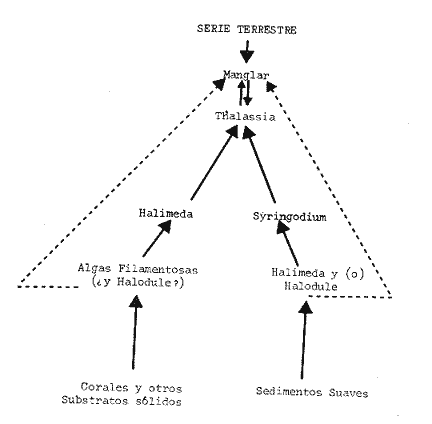 CUADRO 4 [4] c = grado de constancia (%,). [5] Dr = Dominancia relativa media [6] H = índice de diversidad de Shannon-Weaver ConclusionesEn el plano de la fisonomía de las poblaciones, tres grupos faunísticos han sido individualizados; los dos principales presentan su grado de afinidad más elevado en los sedimentos de la pradera de Thalassia testudinum situados mas hacia el mar. Las poblaciones de estas dos praderas, que constituyen una fase transitoria del sedimento, son diferenciables. Si se analiza la distribución de las 45 especies de poliquetos encontrados durante este estudio (cf. lista faunística) se puede constatar que 13 especies, es decir el 29%, de la población global, ya han sido observadas en los sedimentos de otras praderas (Vàsquez, 1979). A éstas se les puede agregar 10 especies, esto es un 51% de la población global, encontradas en los sedimentos arenoso-fangosos, lo cual demuestra la pertenencia de la fauna anelidiana encontrada a aquella de los substratos suaves arenoso-fangosos. Esta observación está en acuerdo con el hecho de que en otros tipos de praderas mediterráneas y tropicales de Cymodea y Halodule conocidas (True-Schlenz, 1965; Le Gall, 1969; Thomassin, 1978), la infauna es comparable a las biocenosis infralitorales de tipo "sedimentos fangosos de aguas tranquilas" (sediments vaseux de mode calme) sensu Peres y Picard (1964). Por otro lado, se observa la presencia de 12 especies, sobre las 45 recogidas, es decir, 27% de la población global, que han sido reportadas colonizando los substratos duros, las bases de algas macroscópicas o fragmentos de gran talla, lo cual corrobora que la fauna de anelidos poliquetos de estas praderas muestra cierta afinidad con la fauna de los substratos duros y de aquellos que son comparables (como estipes de algas y rizomas de fanerogamas). Esta observación también confirma aquellas que han sido anteriormente efectuadas en las praderas con sistema complejo de rizomas (caso de las praderas de Posidonias; cf. Harmelin, 1964). AgradecimientosEs placentero expresar reconocimiento a todos aquellos que, de una manera u otra, han colaborado en la preparación del presente trabajo: P. Glynn e I. Rubinoff quienes recibieron al autor en el Smithsonian Tropical Research Institute en Punta Galeta; a A. González por su muy valiosa ayuda en el terreno y laboratorio; a C. Retiere (Universidad de Rennes/Museo Nacional de Historia Natural de París) y L. Amoureux (Universidad Católica de Angers) quienes permitieron efectuar una estadía en el Laboratorio Marítimo de Dinard y en el Laboratorio de Zoología de Invertebrados de la Universidad Católica de Angers y finalmente a G. Bellan y R. Plante (Estación Marina de Endoume) por sus consejos y puntos de vista y al colega L. D'Croz por sus comentarios. Este trabajo ha sido realizado bajo los auspicios de una beca francesa del Ministerio de Relaciones Exteriores (AF22780), de los programas del Centro Nacional para la Investigación Científica/LA no. 41 (Francia) y del contrato del Centro Nacional para la Explotación de los Océanos (Francia). LITERATURABATTISTINI, R., F. BOURROUILH, J. P. CHEVALIER, J. COUDRAY, M. DENIZOT, G. FAURE, J. C. FISHER, A. GUILCHER, M. HARMELIN-VIVIER, J. JAUBERT, J. LABOREL, L. MONTAGGIONI, J. P. MASSE, L. A. MAUGE, M. PEYROT-CLAUSADE, M. PICON, R. PLANTE, J. C. PLAZIAT, Y. B. PLESSIS, G. RICHARD, B. SALVAT, B. A. THOMASSIN, P. VASSEUR Y P. WEYDERT Tethys Elements de Terminologie Récifale Indopacifique. 1975 1-111 7 (1) D'CROZ, L., J. B. DEL ROSARIO, C. CHANG y G. YEE Oceanografía Biológica un Simposio. Proc. II Latinoamerican Symp. Biol. Ocean Ecología de las Praderas de Thalassia en la Costa Atlántica de Panamá. Universidad de Oriente, 1975 Venezuela 1977 1 37-46 HARMELIN, J. C. Etude de I'Endofaune des "Mattes" d'Herbiers de Posidonia oceanica Delile. Recl. Trav. St. Mar. Endoume-Marseille, Fasc. 51, Bull. 1964 43-105 35 LE GALL, J. Tethys Etude de I'Endofaune des Pelouses de Zosteracées Superficielles de la Baie de Castiglione (Algéric). 1969 395-420 1 (2) LOT-HELGUERA:, A. Estudios sobre Fanerógamas Marinas en las Cercanías de Veracruz. An. Inst. Biol. Univ. Nac. Auton. Mex. Ser. Bot. 1968 1-48 1 O'GOWER, A. K. y J. W. WACASEY Animal Communities Associated with Thalassia, Diplanthera and sand beds in Biscayne Bay. I Analysis of Communities in Relation to Water Movements. Bull. Mar. Sci. 1967 175-210 17 (1) PÉRÈZ, J. M. y J. PICARD Nouveau Manuel de Bionomie Bentique de la Mer Mediterranée. Recl. Trav. St. Mar. Endoume-Marseille, Fasc. 43, Bull. 1964 5-137 31 PIELOU, E. C. Species Diversity and Pattern in the Study of Ecological Succession. J. Theor. Biol. 1966 131-144 13 S0KAL, R. R. y P. H. A. SNEATH Principles of Numerical Taxonomy W. H. Freeman and Co. San Francisco y Londres 1963 359 p. THOMASSIN, B. Peuplements des Sédiments Coralliens de la Région de Tuléar (S. W. Madagascar) et leur Insertion dans le Contexte Côtier Indo-Pacifique. These du Doctorat ès Sciences. AO 1581 Universidad de Aix-Marsella II. Abril 22, 1978, Archivos Originales del CNRS. 1978 TRUE-SCHLENZ, R. Données sur les Peuplements des Sédiments a Petites Phanérogames Marines (Zostera nana Roth et Cymodocea nodosa Ascherson) Compar& a ceux des Habitats Voisins Dépourvus de Vegétation. Recl. Trav. St. Mar. EndoumeMarseille, Fasc. 55, Bull. 1965 96-125 35 VÀSQUEZ-MONTOYA, R. Peuplements des Herbiers de Thalassia testudinum et d'Halodule sp. de la Cote Caraïbe de Panama. Tésis Doctoral Universidad de Aix-Marsella II, Septiembre de 1979. 1979 VOSS, G. N. y N. A. VOSS An Ecological Survey of the Marine Invertebrates of Bimini, Bahamas, with a Consideration of their Zoogeographical Relationships. Bull. Mar. Sci. Gulf Caribb. 1960 96-116 10 WELCH, B. Thesis Aspects of Succession in Shallow Waters in the Caribbean, Ph. D. Florida State University. 1962
|