|
ESTIMACIÓN DE LA EDAD EN UN ORGANISMO COLONIAL PLEXAURA
HOMOMALLA (ESPER). (OCTOCORALLIA: PLEXAURIDAE)
Trabajo recibido el 25 de
febrero de 1981 y aceptado para su publicación el 5 noviembre de
1981.
OSCAR MORENO
Instituto de Ciencias del Mar y Limnología, Ap. Postal
70-305, México, 20, D. F. Contribución 285 del Instituto de
Ciencias del Mar y Limnología, UNAM.
MARTÍN MERINO
Instituto de
Ciencias del Mar y Limnología, Ap. Postal 70-305, México, 20, D.
F. Contribución 285 del Instituto de Ciencias del Mar y
Limnología, UNAM.
ERIC JORDAN DAHLGREN
Instituto de Ciencias del
Mar y Limnología, Ap. Postal 70-305, México, 20, D. F.
Contribución 285 del Instituto de Ciencias del Mar y Limnología,
UNAM.
Se estudia el crecimiento de
Plexaura homomalla en las costas de Quintana Roo con la
finalidad de poder predecir la edad de las colonias para estudios poblacionales
de la especie.
Para ello se utilizan los anillos de
crecimiento presentes en las colonias de este octocoral. Se verifica que dichos
anillos presentan periodicidad y que ésta es anual. Se elabora un modelo
matemático que permite predecir la edad indirectamente, utilizando la
altura de las colonias. Se observa una alta variabilidad en las formas de
crecimiento colonial, la cuál aumenta con la edad. Sin embargo, para
grupos de colonias, el modelo de crecimiento permite hacer predicciones de la
edad promedio con una precisión aceptable para su uso en estudios
poblacionales. Se discuten las causas de la alta variabilidad observada,
considerándose el carácter colonial de la especie como una de las
más importantes.
The growth of
Plexaura homomalla is studied, searching for a method to
estimate the age of the colonies indirectly. This is required for the studies
on population dinamics of the species in the coast of Quintana Roo,
Mexico.
Analizing the relation between size of the colonies
and growth rings present in their sclerotized skeleton, the annual periodicity
of sudi growth rings is verified. A mathematical model that relates the height
of the colony to its age is constructed. Although a wide range of variability
is evident in the growth froms of the colonies, the model allows indirect
determination of the mean age of groups of colonies, with an acceptable
accuracy for its use in the population dinamics studies. The variability
observed is discussed and considered to he due mainly to the colonial nature of
the organism.
Hasta fines de la década de los 60's existía apenas un conocimiento elemental acerca del octocoral Plexaura homomalla. Sin embargo, en 1969 Wenheimer y Spraggins comunicaron el aislamiento de prostaglandinas y algunos de sus derivados a partir de ejemplares de esa especie colectados en Florida. Debido a la importancia de este tipo de sustancias, Plexaura homomalla pasó a ser considerada uno de los recursos más valiosos entre las especies marinas de importancia farmacéutica.
En
1975, con el apoyo del CONACYT,
Consejo Nacional de Ciencia y
Tecnología.
En los organismos como los corales, cuyo crecimiento es muy lento e irregular (Grigg, 1974), no es factible en términos del tiempo que implicaría, conocer la edad por observación directa. De aquí que es deseable encontrar un método que no requiera de dicha observación, el cual puede estar basado en la presencia de marcas de crecimiento en el organismo por estudiar. Este tipo de método ha sido muy utilizado en estudios de bosques, y en el caso particular de especies marinas también es común. Cabe mencionar los anillos en escamas de peces (Van Oostan, 1957), las bandas en conchas de moluscos (Evans, 1972), y los anillos presentes en corales preciosas del género Antipathes (Grigg, 1976). Los octocorales muestran también estas bandas de crecimiento, en forma de anillos, y fueron mencionadas por Linneo (según Johnton 1847) al describir el esqueleto axial de una "gorgonia". Asimismo, Goldberg (1976) reporta la existencia de anillos de crecimiento enPseudopterogorgiaspp. y Pterogorgia spp., y en las especies Gorgonia ventalina, Swiffla excerta y Muricea muricata. Grigg (1974) no solo reporta los anillos deMuricea californica y M. fruticosa de las costas de California, U.S.A., sino que también demuestra su carácter anual. Dado quePlexaura homomalla también presenta marcas de crecimiento en forma de anillos en el eje axial (Opresko, 1974), se pretende en este trabajo utilizar estos anillos para estimar la edad de las colonias. Sin embargo, para poder contar dichas marcas de crecimiento, se requiere obtener una sección del eje de la colonia en su parte basal. Esto implica desprender la colonia del substrato al cual se encuentra adherida; lo que generalmente causa la muerte de ésta. Debido a que en un estudio poblacional se requiere estimar la edad de un gran número de individuos, es necesario utilizar una forma de determinación indirecta de la edad que no dañe al organismo, lo cual en octocorales elimina la posibilidad de la estimación indirecta a través de biomasa. Generalmente se recurre a encontrar una relación entre las marcas de crecimiento y alguna característica del organismo que permita estimar su edad sin dañarlo, y sin una gran cantidad de esfuerzo. Siendo el objetivo principal de este trabajo el encontrar un método lo más sencillo posible para determinar la edad de un gran número de individuos sin dañarlos, y con la precisión necesaria para un estudio poblacional; se decidió generar un modelo de crecimiento que, en base a la medición de una característica merística de la colonia y su relación con los anillos de crecimiento, permitiese lograr la estimación de edad en estos términos. ÁREA DE ESTUDIOLos estudios poblacionales de Plexaura homomalla en las costas mexicanas se están llevando a cabo inicialmente en la barrera arrecifal del noreste del estado de Quintana Roo, limitada por la Punta Nizuc y el poblado de Tulum. De aquí que es deseable un modelo de crecimiento que sea adecuado para estimar la edad de colonias provenientes de cualquier punto de esta zona. Para lograr lo anterior, se decidió utilizar colonias de cuatro localidades distintas: Puerto Morelos, Punta Maroma, Yakul y Akumal (Fig. 1), las cuales fueron estudiadas en 1976 durante la evaluación poblacional de la especie, y que pueden considerarse representativas de la zona (Jordán y Nugent, 1978). Otras características que llevaron a seleccionar estas localidades fueron la abundancia y la accesibilidad de las colonias, con la idea de facilitar el muestreo y dañar lo menos posible a la población. 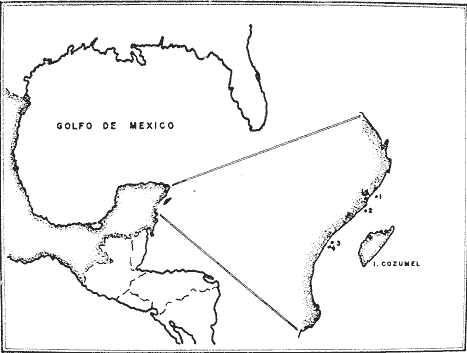 Fig. 1. Área de estudio y localización de los muestreos. (1) Puerto Morelos, (2) Punta Maroma, (3) Yakul, (4) Akumal. MÉTODOS
CONTEO DE LOS ANILLOS DE CRECIMIENTOPara poder contar el número de anillos es necesario desprender la colonia haciendo un corte en la base del esqueleto axial. Una sección de éste, lo más cercana posible al disco basal, se utiliza para llevar a cabo el conteo de los anillos en el laboratorio. Las secciones del eje se mantienen constantemente en una solución de Na Cl de 35 gr/lt, ya que de secarse los ejes cortados, se presentan deformaciones que dificultan la estimación de los anillos. Los anillos son aparentes a simple vista (Fig. 2), pero con el fin de lograr la máxima precisión y repetibilidad en el conteo se ensayaron diversos procedimientos, desde rebajar las secciones hasta un espesor de aproximadamente 0.5 mm y contar por transparencia, hasta la utilización de rayos X y Microscopio Electrónica de Barrido. De los métodos que aportaron los mejores resultados se seleccionó el menos costoso,, que consiste en pulir cada sección con una lija para madera de grano fino, utilizando para el conteo un microscopio de disección, iluminando desde diversos ángulos para resaltar las líneas anulares y contando los anillos a lo largo de diversos radios de la pieza. La incertidumbre promedio con el conteo es de ± 0.6 anillos variando desde ± 0 para las muestras con las estructuras anulares más fácilmente distinguibles hasta ± 2.5 en 3 casos con un alto número de anillos relativamente difusos entre sí. CARACTERÍSTICAS MERÍSTICASEntre las características estudiadas se incluyeron la altura de la primera bifurcación a la parte más alta de la colonia (h), el diámetro mayor de la fronda vista desde arriba (D), el diámetro menor tomado desde el mismo ángulo (d), el área de la sección transversal del eje de la colonia en su parte basal (A), y los índices de cobertura (I.C.) utilizados anteriormente en octocorales por Jordán y Nugent (1978).  Fig. 2. Sección transversal del eje de una colonia de Plexaura homomalla en su parte basal, utilizando para el conteo de los anillos. La medición de la altura y los diámetros se realizó "in situ" mediante un bastón de aluminio graduado en intervalos de 5 cm, puesto que el oleaje, y las condiciones bajo el agua no permiten una precisión mayor. El área de la sección transversal del eje fue medida con un planímetro sobre un dibujo en papel, transferido del esqueleto correspondiente. Los índices de cobertura se calcularon en base a las siguientes ecuaciones (Jordán y Nugent, 1980): 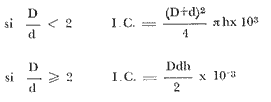 CÁLCULOSPara los cálculos necesarios en el análisis estadístico se utilizó la computadora Burroughs 6700 del Centro de Servicios de Cómputo de la UNAM, y los programas necesarios fueron elaborados por los autores. RESULTADOS Y DISCUSIÓN
PERIODICIDAD DE LOS ANILLOS Y MODELO DE CRECIMIENTOPara verificar el posible carácter periódico de las estructuras anulares y con el fin de seleccionar cuál de las características merísticas de las colonias sería la más adecuada para elaborar el modelo de crecimiento necesario, se hizo un primer estudio con 50 colonias. En este estudio se calculó el coeficiente de correlación entre el número de anillos y la altura (h), los diámetros (d,D), el índice de cobertura (I.C.) y el Área de la sección basal (A) de las colonias. En la tabla 1, se muestran los resultados del análisis de correlación. 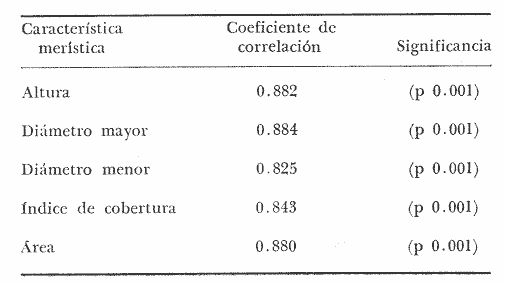 TABLA 1. CORRELACIÓN ENTRE EL NúMERO DE ANILLOS Y LAS CARACTERÍSTICAS MERISTICAS ESTUDIADAS (n = 50) Resalta el hecho de que para todos los casos el valor de los coeficientes de correlación es aceptable y positivo. Esto indica que el aumento en el número de anillos es de carácter periódico y esta ligado a un aumento en las diversas dimensiones de la colonia. Así los anillos que estas colonias presentan pueden considerarse un estimador válido de su edad relativa. Estos resultados no son excepcionales. Los estudios que Grigg llevó a cabo en California conMuricea muricata y Muricea californica(1974) evidencian el carácter anual de estas marcas. Por otro lado, Opresko (1974), en sus estudios sobre Plexaura homomalla en las aguas de Florida, elaboró una curva de crecimiento en base a la biomasa de las colonias y su edad relativa para esa región. A partir de esta curva asignó una periodicidad anual a los anillos de crecimiento de la especie en las aguas de esa región. Es importante destacar que la presencia de anillos de crecimiento en múltiples especies de aguas tropicales como son Muricea spp. (Verril, 1968-79),Corallium y Keratosis de regiones someras de Argelia (Dantan, 1928), así como de bandas en conchas de balanos y corales solitarios de regiones abisales (Grigg, 1974), han llevado a considerar que en estos casos el mecanismo que da lugar a estas estructuras es de origen principalmente biológico, como pueden serlo variaciones en la fuente alimenticia o el ciclo reproductivo (Grigg, 1974). En particularPlexaura homomalla sigue un ciclo reproductivo anual en el Caribe Mexicano, y se ha encontrado que la época crítica de reproducción abarca precisamente los meses de julio y agosto (Martín, 1980). Por otro lado, no sé debe descartar por completo la influencia que los cambios estacionales en las condiciones ambientales pueden tener directa o indirectamente sobre el crecimiento. En base a los resultados de los análisis de correlación y dado el ciclo reproductivo anual marcado que presenta la especie en las costas de Quintana Roo, se puede considerar que los anillos de crecimiento que presenta Plexaura homomalla en esta zona también son de carácter anual, lo que los hace sumamente útiles como un indicador de la edad de las colonias. Sin embargo, los anillos muestran una alta dispersión en sus relaciones con las características merísticas estudiadas, siendo esta dispersión considerable aún en el caso de las características que la presentan en un grado menor como son el diámetro mayor y la altura. Esta dispersión puede ser intrínseca al fenómeno o debida parcialmente al muestreo, por lo que para generar un modelo con una mayor representatividad se aumentó el número de elementos muestreados a 120, lo que teóricamente reduciría la variabilidad generada por el muestreo y las técnicas de medición. Este nuevo esfuerzo fue concentrado en una sola de las características merísticas estudiadas. La repetibilidad de su determinación in situ, así como su uso por otros autores (Grigg, 1974), (Opresko, 1974), (Kinzie, 1974), y la alta correlación que presentó, situaron a la altura (h) de la colonia como la mejor característica para la elaboración de un modelo de crecimiento. La expresión gráfica de los 120 valores de altura y sus respectivos números de anillos puede observarse en la figura 3. Se observa que sigue existiendo una fuerte dispersión de los datos. Otra posible fuente de dispersión residiría en la existencia de comportamientos de crecimiento distintos en las cuatro localidades estudiadas, lo cual puede ser estimado cuantitativamente. La figura 4 muestra gráficamente la relación entre el número de anillos de cada colonia y su altura, agrupados por localidad. Para averiguar si los cuatro grupos de datos puede ser considerados como provenientes de la misma población se utilizó una prueba F para muestreo múltiple (Zar, 1974), tomando como hipótesis nula (Ho) la igualdad de ajuste de cada muestra a una regresión lineal. El valor obtenido fue de F = 1.76, que corresponde en las tablas a una probabilidad (P) de 0.10 < P < 0.50 (F (0.05 (2) 6, 111) = 2.53. 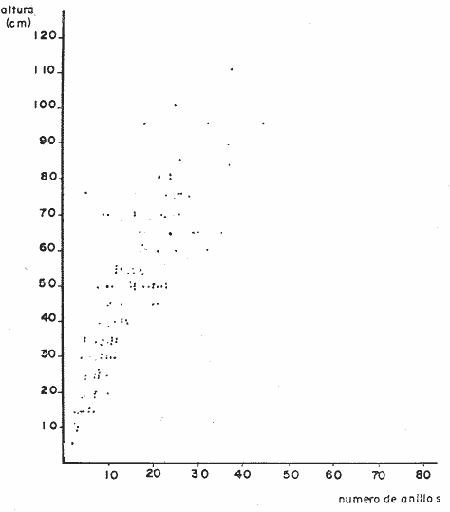 Fig. 3. Parejas de valores (anillos, altura) para las 120 colonias estudiadas. Este resultado nos permite afirmar que no se puede rechazar (Ho) con confianza, por lo que para poder aceptar la hipótesis nula se calculó el riesgo de error tipo II (β). Utilizando las curvas de potencia para la prueba de F (Winer, 197 1) se obtiene que la probabilidad de que la varianza entre las pendientes verdaderas de los cuatro grupos de valores sea mayor a 0. 12 veces la varianza de los errores de, regresión es menor al 10%. Esto quiere decir que se puede afirmar, con una probabilidad de equivocarse menor al 10%, que la dispersión interna de los grupos es 8.3 veces más grande que la dispersión entre los grupos; lo cual indica que no hay una diferencia significativa entre ellos. 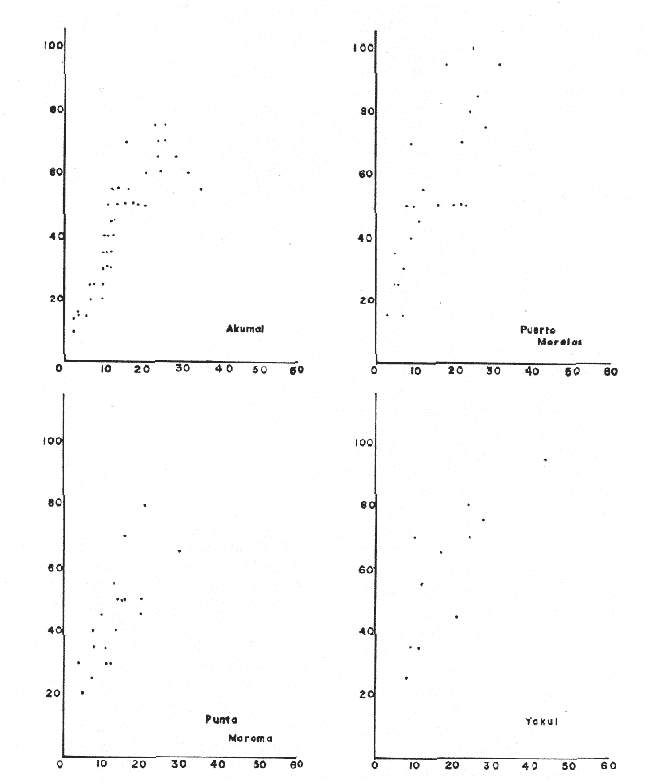 Fig. 4. Parejas de valores (anillos, altura) separadas por localidad. Por lo anterior, se aceptó la hipótesis nula (Ho), y todos los tratamientos posteriores se aplicaron a los valores registrados como una misma muestra, pudiéndose considerar que todas las inferencias derivadas de su análisis son válidas para las 4 localidades. Modelo de crecimiento La función de crecimiento que se utilizó para la elaboración del modelo fue una curva exponencial de tipo Von Bertalanffy, ya que se considera una de las expresiones de crecimiento más completas, su uso en pesquerías esta ampliamente difundido (Gulland, 1971), y su aplicación a problemas similares ha resultado adecuado (Grigg, 1974). La expresión general de esta ecuación es: Y = Y α (I -exp (-k e)) (1) donde Y= dimensión del espécimen a la edad e e = edad Y α = dimensión máxima promedio para la población k= valor constante para la población, que refleja el cambio en la velocidad de crecimiento, con la edad. Generalmente esta ecuación se utiliza en pesquerías considerando como "Y" la longitud de los espéciménes (Gulland, 1971). De esta manera la ecuación de Von Bertalanffy se expresa en este caso como: h = hα (1 - exp (-k e)) (2) en la cual h = altura de la colonia a la edad e h α= altura máxima promedio para la población e = edad de la colonia en años. Para llevar a cabo el ajuste de este modelo a los datos por medio del método de mínimos cuadrados, es necesario hacer una transformación logarítmica de la ecuación (2), que la convierte en: In (I - h/hα) = k e (3) Aplicándose así una regresión lineal a las parejas (In (1-hi/h*), ei), este procedimiento da como resultado una ecuación del tipo: In (I - h/hα) = k e + b (3') Sin embargo, este análisis presenta dos problemas: por un lado la necesidad de conocer de antemano el valor de hα (o contar con un método para calcularlo) y por otro la aparición del término b al aplicar una regresión lineal tradicional. Este término en la ecuación final de crecimiento (tipo (2)), implicaría que las colonias de 0 años (e = 0) tienen un tamaño exp(b) cm, lo cual carece de sentido para esta especie. Para solucionar estos dos puntos se utilizó el siguiente procedimiento: el valor de hoo se calculó por iteración, asignado un valor inicial arbitrario y minimizando el valor de b. El valor obtenido fue de 116.82 cm, el cual concuerda con las observaciones de campo efectuadas durante la evaluación poblacional de la especie a lo largo de más de 150 km de costa (Jordán, 1977). Con este valor de hoo se aplicó una regresión forzada al origen para calcular el valor de k, en base al proceso propuesto por Zar (1974). La ecuación resultante expresa el crecimiento en altura de las colonias en función de la edad para la población y es la siguiente: h = 116.82 cm (1 - exp (-0.0396 e)) (3) cuyo coeficiente de correlación, r² = 0.87, es altamente significativo considerando el número de datos (n = 120). En la figura 5 se observa la representación gráfica del modelo y los puntos que lo originan. En la literatura existen sólo otros dos estudios que proðorcionan información acerca del crecimiento dePlexaura homomalla. Uno de ellos es el de Opresko (1974), quien en base a la estructura de altura y estimaciones de la biomasa de una población de 4 años establece el carácter anual de los anillos de crecimiento de especímenes de la Florida. El otro lo realizó Kinzie (1974), en Gran Cayman, podando colonias de diversas alturas y midiendo la tasa de crecimiento en las puntas regeneradas (aún cuando la información reunida por Kinzie solo abarca 2 años, provee un marco de referencia de utilidad sobre el tema). 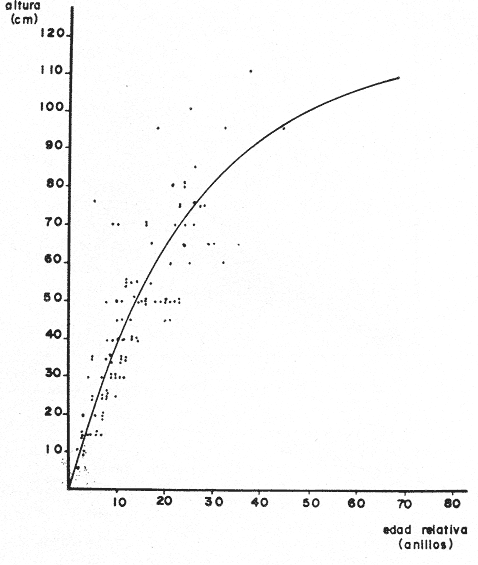 Fig. 5. Modelo de Crecimiento propuesto junto con los valores muestrales a partir de los cuales se obtuvo el mismo. En la figura 6 se presentan los modelos que resultan de los datos obtenidos en Florida, Gran Cayman y Quintana Roo. Suponiendo la periodicidad de los anillos como anual, puede observarse que el modelo obtenido para la especie en este trabajo se sitúa dentro del rango de los otros, lo cual refuerza su validez y ratifica el carácter anual de ¡los anillos de crecimiento. PREDICCIÓN DE LA EDAD Y VARIABILIDAD EN EL CRECIMIENTOCon el modelo de crecimiento elaborado, es posible predecir la edad de los individuos de una población. En este caso el modelo descrito permite hacer tal estimación de una manera indirecta, a partir de la altura de las colonias, por medio de una predicción inversa. Esto se lleva a cabo encontrando la edad correspondiente a una altura dada en la gráfica de la figura 5 o bien, despejando la ecuación (3) para e, que se expresa como: e = -1 ÷ 0.0396 In (1 - h/ 116.83) (4) y sustituyendo el valor conocido de la altura (h) lo cual permite resolver la ecuación (4) para conocer la edad correspondiente (e). 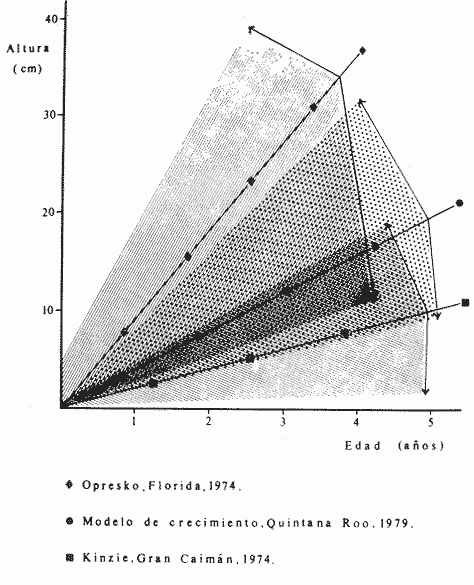 Fig. 6. Comparación entre los modelos de crecimiento para Plexaura homomalla reportados por Opresko, Kinzie y los autores. El rango de los valores muestrales utilizados en cada caso se indica con flechas. Los valores de la edad así obtenidos tienen cierta incertidumbre como consecuencia, entre otras cosas, de la variabilidad natural del fenómeno, los errores de medición, el muestreo y el procesamiento de la información. El orden de magnitud de esta incertidumbre sobre la predicción (es decir, la precisión de la misma) se estima calculando los intervalos de confianza que le corresponden al modelo. Aun cuando el cálculo de los intervalos de confianza, para modelos de predicción en general, supone una dispersión uniforme de los datos a lo largo de toda la curva, esta suposición no parece ser adecuada en este caso. En las gráficas de la figura 4 puede observarse que la dispersión de los datos se incrementa conforme aumenta la edad y la altura. Por esta razón, para obtener intervalos de confianza que representasen este fenómeno se procedió al análisis de la varianza de los datos. Para ello se estudió la variación de la dispersión de la edad en función de la altura, debido a que ésta es la variable sujeta a determinación directa y considerando además, que el muestreo agrupó a las colonias en clases de altura de 5 cm. Para todas aquellas clases que constan de un número suficiente de datos (n > 5) se calculó la desviación standard de e: y los valores obtenidos se graficaron contra el valor de altura correspondiente a cada grupo de datos 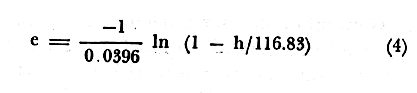 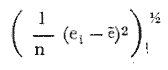 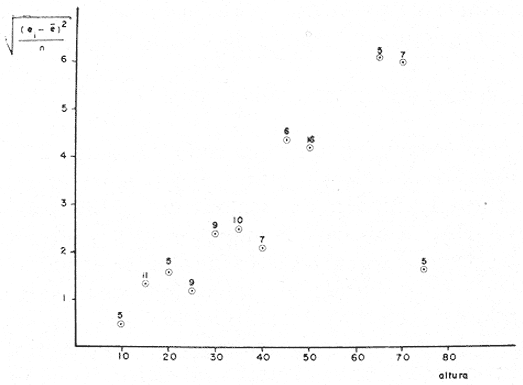 Fig. 7. Variación de la desviación standard de la edad con el incremento en altura. Analizando dicha representación gráfica (Fig. 7), se observa una tendencia lineal o de tipo cuadrático. Por ello se ensayaron los modelos correspondientes 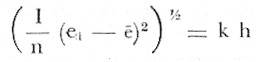 para la lineal y 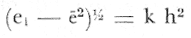 para la cuadrática (k es una constante). Haciendo las transformaciones necesarias, se aplicaron regresiones forzadas al origen para obtener los valores de k. (Para estos cálculos no se utilizó el punto de h = 75 cm debido a su comportamiento disparado del resto de los datos, lo cual es aceptable tomando en cuenta que dicho punto además era de los de menor número de valores (n = 5)). Los resultados de las regresiones son los siguientes:  Tomando en cuenta la sencillez de su manejo y el valor de los coeficientes de correlación de ambos modelos; se consideró al modelo lineal más adecuado en este caso. De aquí que la expresión de la varianza necesario para el cálculo de los intervalos de confianza sea: 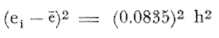 Substituyendo esta ecuación en la expresión tradicional para los intervalos de confianza (predición de e) se pueden obtener dichos intervalos sobre la expresión transformada log (1 (1-hi/*) = ke y aplicando la transformación inversa, obtener los intervalos de confianza para la predicción de la edad (e) en base a la altura de la colonia (h) y a la ecuación h = 116.83 (1 - exp (-0.0396 e)) (Fig. 8). Puede observarse que estos intervalos reflejan la dispersión creciente con la altura que muestran los datos. Por otro lado existe un factor más que influye en la precisión de la predicción de edad con este modelo. Este factor es la cantidad de individuos de la misma altura cuya edad se quiere predecir. Es decir, no se obtiene la misma incertidumbre sobre la predicción de la edad de una colonia, que sobre la de la edad promedio de un grupo de estas. Así mismo, es distinta la precisión en la edad promedio de un grupo de cinco colonias que en un grupo de cincuenta colonias de la misma altura. En la figura 9 se presentan los intervalos de confianza sobre la predicción de la edad media de diferentes números de colonias (n = 1, 5 y 50). Puede observarse que si bien la predicción de la edad de una colonia, o de un número reducido de éstas tiene una precisión demasiado baja; la precisión aumenta considerablemente al aumentar el número de colonias de la misma altura cuya edad promedio se quiere estimar. Por ejemplo, para una altura de 40 cm, la predicción de 10.6 años tendría una incertidumbre de ± 8.5 años para una colonia: ±1.9 años para n 20 y de solamente ± 0.89 años para n = 100. Esta situación determina que el modelo obtenido sea de gran utilidad en la estimación de la edad de grandes números de colonias, como es en el caso del estudio poblacional al cual va dirigido. Dentro del estudio P. homomalla que se ha estado llevando á cabo en el Caribe Mexicano, la evaluación poblacional de la especie se esta haciendo agrupando a las colonias en clases de altura (Jordán y Nugent, 1978) y de ahí que el modelo desarrollado es adecuado para conocer la estructura de edades de esta población. 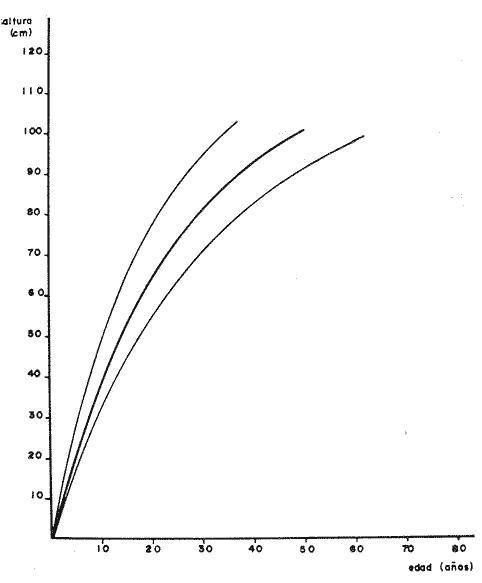 Fig. 8. Intervalos de confianza para la predicción de la edad en base a la altura utilizando el modelo propuesto. Se muestra el intervalo correspondiente a la predicción de la edad promedio para 10 colonias de la misma altura (n = 10). 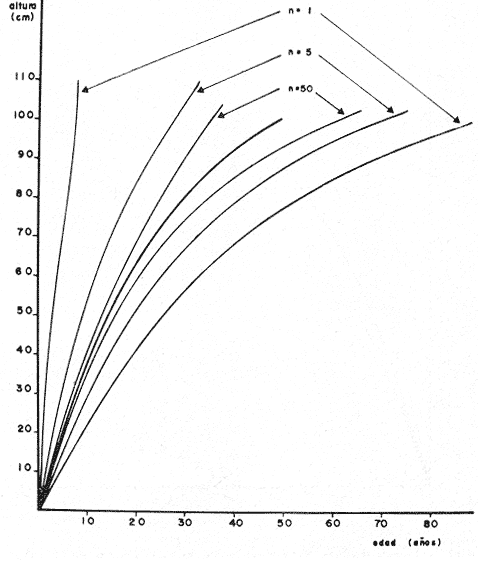 Fig. 9. Variación de la amplitud de los intervalos de confianza para la predicción de la edad promedio de un grupo de colonias de la misma altura en base al número de colonias (n). Variabilidad en el crecimiento La dispersión que muestran los datos obtenidos de la relación altura-anillos repercute en el uso del modelo de crecimiento, ya que influye sobre la forma y la amplitud de los intervalos de confianza para la predicción de edad. Por ello, se ha considerado importante analizar las posibles causas de esta dispersión, que en este caso pueden ser principalmente la imprecisión en la medición de alturas y en el conteo de los anillos, por lo que respecta al proceso experimental; y la variabilidad natural inherente a la población estudiada, en cuanto a la edad y la altura de las colonias se refiere. De los procedimientos experimentales cabe destacar que la imprecisión al medir las colonias fue de ± 2.5 cm, lo que implica un máximo de 3 años de error en el modelo para colonias de hasta 80 cm de altura. Por otro lado, la repetibilidad en el conteo de anillos es muy alta (98% de las muestras no reportaron desviaciones mayores a 2 anillos en los conteos repetidos), por lo que los errores de determinación originan solamente una pequeña parte de la dispersión observada. Como además, la incertidumbre se mantiene constante para todas las alturas, no es posible explicar el aumento de la dispersión con la altura y el comportamiento de los puntos únicamente en base a estos aspectos. Si la dispersión de los datos no parece ser consecuencia únicamente del manejo experimental, es de esperarse que exista de hecho en la población una fuerte variabilidad y que ésta se vea reflejada en la muestra con la cual se trabajó. Este. fenómeno es común a los estudios sobre crecimiento de octocorales independientemente de las técnicas experimentales utilizadas (Opresko, 1974; Kinzie, 1974) o las especies con las que se trabaja (Grigg, 1974). Esta variabilidad puede deberse tanto a factores externos como a características inherentes a las poblaciones estudiadas. Dentro de las primeras, y por lo que se refiere a las interacciones biológicas entre P. homomalla y los demás integrantes de la comunidad arrecifal, se podría considerar la influencia de depredadores como Hermodice carunculata(Glynn, 1973) o Cyphoma gibbossum(Kinzie, 1974). Estos últimos, además de alimentarse de las colonias dejan expuesto parcialmente el esqueleto protéico de las Mismas, lo que puede favorecer la implantación de epibiontes como algas filamentosas oMillepora alcicornis, retrasando el crecimiento de la colonia por la presencia de ellos o por el proceso de regeneración. Puede decirse, sin embargo, que debido a que se trata de depredadores no específicos, su influencia en la tasa de crecimiento deP. homomalla es poco significativa (no obstante, habría que definir cuantitativamente la intercalación para estimar su importancia relativa real respecto a la variabilidad). También, la competencia por luz o por espacio con otros corales (Connel, 1974) podría dar lugar a que las colonias crecieran inclinadas o deformadas, haciendo diferir la relación entre la edad y sus dimensiones con respecto a las colonias que crecieran libremente. Esta interacción es difícil de expresar cuantitativamente para integrarla al problema que aquí se trata, pero la distribución de parche y el tipo de medio ambiente local en el que se encuentra la especie en la zona de estudio minimizan el efecto de los scleractinios arborescentes y masivos sobre la población de P. homomalla. Otra influencia externa que parece ser mucho más importante que las anteriormente discutidas es el medio ambiente físico, principalmente olas y corrientes (Jordán y Nugent, 1978). Para comprender la relevancia que las condiciones ambientales llegan a tener sobre la forma de crecimiento dePlexaura homomalla, es necesario llamar la atención sobre una característica inherente a la especie (y a los octocorales en general) que es: la forma de organización de tipo colonial. Esto implica que las consideraciones acerca de la relación entre sus dimensiones y su edad se analicen y se utilicen con un criterio distinto al que se aplica en estudios poblacionales de organismos no coloniales (conclusión a la que también llegan Huges y Jackson (1980) al describir los procesos que intervienen en el crecimiento y mortalidad de corales escleractinios). Es así, que al comparar entre sí organismos de una misma especie cuando ésta no es colonial, se puede apreciar que las dimensiones de cada individuo mantienen ciertas proporciones características en su forma y estructura. En el caso de los octocorales, esto es válido únicamente para los pólipos, es decir, los individuos que integran la colonia. Sin embargo, cada colonia es el resultado de la agregación de un gran número de pólipos, los que al reproducirse asexualmente incrementan la biomasa y el tamaño de la colonia de acuerdo a un patrón básico, de tipo arborescente paraP. homomalla, como se puede apreciar en las colonias que se muestran en la figura 10. Siguiendo este patrón de crecimiento pueden presentarse múltiples variaciones en sus proporciones sin que se mantenga una relación estrecha de las dimensiones con la edad. Estas variaciones son apreciables no solo entre colonias sino inclusive a lo largo de la vida de un mismo ejemplar. Esto último se debe a que la biomasa de la colonia puede incrementarse indistintamente aumentando el tamaño y número de puntas o de ramas, en engrosamiento de la base o de otras porciones de las ramas, etc. (Kinzie, 1974). 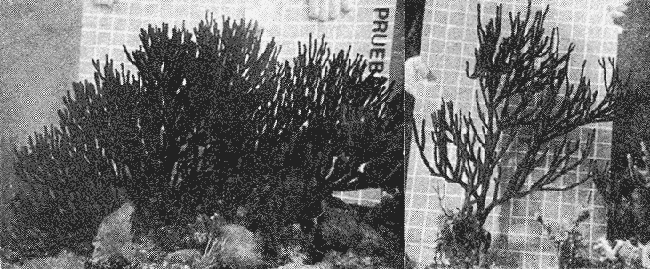 Fig. 10. Dos colonias de Plexaura homomalla. Puede observarse tanto el patrón arborescente de crecimiento de la especie, como las variaciones en forma y densidad que suelen presentarse. Esta variabilidad de los octocorales por lo que se refiere a su forma de crecimiento, se ve reflejada claramente en su respuesta morfológica a las condiciones ambientales, caracterizándolos como organismos ecomorfos. Esto se ha puesto en evidencia en múltiples estudios tendientes a estimar la influencia del medio ambiente sobre la orientación y la forma de gorgonáceos y otros octocorales, en función de algunas características del habitat (como las corrientes predominantes o el oleaje). Entre estos trabajos se encuentra el de Wainwright y Dillon (1969) con Gorgonia ventalinayG. flabellum; el de Grigg (1972) con Muricea californica y M. fructicosa; el de Velmirov (1976) con Eunicella cavollini y el de Muzik y Wainwright (1977) quienes estudian cinco especies de cinco familias y dos órdenes distintos. Por lo que respecta a Plexaura homomalla, Jordán y Nugent (1978) al iniciar la evaluación poblacional de la especie en el Caribe Mexicano, clasifican las colonias en "redondeadas" características de lugares protegidos y "aplanadas o de candelabro", que predominan en aguas agitadas. Esta cualidad ecomórfica y la naturaleza colonial de P. homomalla, son aparentemente los factores que más contribuyen a la gran variabilidad que puede apreciarse tanto en las formas de crecimiento como en la relación altura-edad, que se ha estudiado en la población de esta especie en el Caribe Mexicano y, posiblemente, en los otros estudios a los que se ha hecho referencia en este trabajo. Conclusiones1. Los anillos de crecimiento que presentan las colonias de P. homomalla de la costa de Quintana Roo, pueden considerarse marcas de crecimiento anuales. 2. La ecuación: h= 116.82 (l-exp (0.0396 e)) es el modelo de crecimiento más adecuado para describir la relación entre la altura de las colonias h (cm) y su edad e (años) 3. El modelo de crecimiento obtenido permite estimar indirectamente la edad de las colonias con buena precisión para estudios poblacionales. 4. Los datos experimentales muestran una fuerte dispersión cuyo origen más probable es la variabilidad en el crecimiento que se puede atribuir al carácter colonial de la especie, mismo que da lugar a que sean organismos ecomorfos. AgradecimientosLos autores agradecen a las siguientes personas la ayuda prestada para la elaboración de este estudio: A Ignacio Méndez y Joaquín Díaz del Instituto de Investigaciones en Matemáticas Aplica das y Sistemas, por su asistencia en los tratamientos estadísticos utilizados. A Richard W. Grigg de la Universidad de Hawaii, por sus comentarios. A los compañeros de trabajo: Eduardo Martín, Hilda León y Patricia Thomé; por su ayuda en el trabajo de campo y la confección de las gráficas. Este trabajo fue realizado con el apoyo del Consejo Nacional de Ciencia y Tecnología (CONACYT) como parte del Proyecto de Investigación ecológica correspondiente al programa: "Investigaciones Ecológicas y Químicas para la Extracción de Protaglandinas a partir de Corales Blandos en el Caribe Mexicano" (Convenio CONACYT-UNAM). LITERATURACONNELL, J. H., Biology and Geology of Coral.Population ecology of reef buildind corals. In: Jones, 0. R. y Endean, R. (Eds.). Reefs. V. II. Academie Press New York. 1974. 205-246. DANTAN, J. L.,Corallium rubrum. Lamarch. Soc. Zool. France. Bull.Recherches sur la croissance du corail rouge 1928.42-46.53 EVANS, J. W.,Clinocardium nuttalli, Science Tidal growth increments in the cockle1971.416-4171976 GLYNN, P. W., Biology and Geology of Coral Reefs Aspects of the ecology of coral reefs in the Western Atlantic region O. A. jones y R. Endean (Eds.). Biology I, Academic Press New York1973.271-324.Volume II GOLDBERG, W. M., Mar. Biol. Comparative study of the chemistry and structure of Gorgonian and Antipatharian Coral Skeletons.1976.253-267. (35): GRIGG, R. W., Limnol. and Oceanogr. Orientation and growth forms of sea fans. 1972.185-192.17 (2) ---, Ecology Growth Rings: annual periodicity in two gorgonian Corals. 1974.876-881.55 ---, Fishery management of precious and stony corals in HawaiiUNIHI-SEAGRANT-TR-7703Hawaii1976. 48 pp. GULLAND, J. A., Manual de Métodos para la Evaluación de Poblaciones de Peces. FAO, Ed. Acribia, Zaragoza1971. 307 p. HUGES, T. P., J. B. C. JACKSONScience Do corals lie about their age? some consequences of partial mortality, fission and fusion.1980.713-715. 209 JOHNSTON, G., A History of the British Zoophytes. John Von Voorst2nd Ed.London1847.434 p. Vol. I. JORDAN, E., Proyecto de investigación sobre "La Extracción de Prostaglandinas de Corales Blandos y su Exploración Racional con el fin de Preservar el Recurso". Programa celebrado entre la UNAM, a través del CCML y el CONACYT. Informe del Programa sobre "Ecología y Evaluación de las Poblaciones de Plexaura homomalla y su Explotación Racional con el fin de Preservar el Recurso". 1977. JORDAN, E., R. S. NUGENTAn Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México Evaluación poblacional de Plexaura homomalla (Esper.) en la costa noreste de la Península de Yucatán (Octocorallia). 1978.179-188.5 (1): KINZIE, R. A., Prostaglandins from Plexaura homomalla: Ecology, Utilization and Conservation of a Major Medical Marine Resource: A. Symposium Plexaura homomalla: The biology and ecology of a harvestable marine resource. In: Bayer, F. M. (Ed.) Stud. Tropic. Oceanogr. 12, Univ. of Miami Press197422-38. MARGALEFF, R.,Ecología. Ed. Omega. Barcelona.1974.951pp. MARTIN, E., Tesis profesional Reproducción del coral blando Plexatira homomalla (Esper) en el Mar Caribe Mexicano.Fac. de Ciencias, Universidad Nacional Autónoma de México.1980. MUZIK, K. and S. WAINWRIGHTBull. Mar. Scie. Morphology and habitat of five fijian sea fans.1977.308-337.27 (2): OPRESKO, D. M.,Prostaglandins from Plexaura homomalla: Ecology Utilization and Conservation of a Major Medical Marine Resource: a Symposium.Recolonization and regrowth of a population of the gorgonian Plexaura homomalla. In: Bayer, F. M. (Ed.) Stud. Trop. Oceanogr.,1974.101-110.12: VAS OOSTAN, J., The physiology of fishes. Skin and scales. In: M. E. Brown (Ed.) Acad. Press. Inc., New York1957.207-244. VERRIL, A. E., Trans. Com. Acad. Arts. and SciencesReview of the corals and polips of the West Coast of America.1968-70.377-570.1 (6, 7) WEINHEIMER, A. J., R. L. SPRAGGINS Plexaura homomalla (Esper.). Tetrahedron Lett. Two new protaglandins isolated from the gorgonian1969.5183. WINER, B. J., Statistical principles in experimental design Mc Graw Hill2nd. Ed.Londres1971.470 p. ZAR, J. H., Biostatistical Analysis. Prenticc-Hall Inc., New Jersey1974. 620 p.
|