|
TAXONOMÍA,
BIOLOGÍA Y ECOLOGÍA DE LOS TETRAODONTIDOS DE LA LAGUNA DE
TÉRMINOS, SUR DEL GOLFO DE MÉXICO (PISCES:
TETRAODONTIDAE)
Recibido el 12 de diciembre de 1980 y aceptado para su
publicación el 15 de abril de 1981.
LÍA MALLARD COLMENERO
Departamento de
Pesca, Dirección General de Acuacultura, Alvaro Obregón No.
269-7° piso, México 7, D. F.
ALEJANDRO YÁÑEZ-ARANCIBIA
Universidad Nacional Autónoma de México, Instituto
de Ciencias del Mar y Limnología, Laboratorio de Ictiología y
Ecología Estuarina. Contribución 281 del Instituto de Ciencias
del Mar y Limnología, UNAM.
FELIPE AMEZCUA LINARES
Universidad Nacional Autónoma de México, Instituto
de Ciencias del Mar y Limnología, Laboratorio de Ictiología y
Ecología Estuarina. Contribución 281 del Instituto de Ciencias
del Mar y Limnología, UNAM.
Actividades de
campo entre julio de 1976 y marzo de 1979 permitieron obtener una importante
colección de peces de la familia Tetraodontidae de la Laguna de
Términos, Campeche. Se entrega una diagnosis taxonómica de la
familia y una clave artificial para determinar los géneros y especies.
Se realiza una revisión taxonómica de las mismas que se refuerza
con información merística y morfométrica,
coloración y comparación taxonómica con especies afines
del Golfo de México.
Cinco especies existen de esta
familia en el área,i.e., Sphoeroides
testudineus(Linnaeus), S. nephelus (Goode y Bean),
S. greeleyi (Gilbert),S. spengleri
(Bloch) yLagocephalus laevigatus (Linnaeus). Las dos
primeras representaron más del 95% de la captura total y dominan en
biomasa. Se discute la distribución y abundancia para cada especie, y se
entrega y discute información biológica sobre relaciones de
longitud/peso, tamaño de las poblaciones, proporción de sexos,
madurez y relaciones tróficas. La época del año y los
distintos habitat que existen dentro de la Laguna de Términos, rigen la
ecología de las poblaciones de estas especies. Todas presentan patrones
de distribución y abundancia definidos, destacando S.
testudineus (Linnaeus) yS. Nephelus (Goode y Bean)
como especies típicas de la estructura de las comunidades de peces de la
Laguna de Términos.
Field work carried out between
July, 1976, and March, 1979, resulted in an important collection of fish
beloging to the Tetraodontidae family from, the Terminos Lagoon in Campeche,
Mexico. This paper presents a taxonomic diagnosis of the family and an
artificial key to determine genus and species. It provides a taxonomic review
of these species reinforced with meristic and morphometric information,
coloration data and taxonomic comparison with similar species of the Gulf of
Mexico.
Five species of this family are found in the
area, i.e., Sphoeroides testudineus (Linnaeus),S.
nephelus(Goode and Bean), S. greeleyi(Gilbert),
S. spengleri (Bloch) andLagocephalus
laevigatus(Linnaeus). The first two comprise more than 95 per cent of
the total catch and predominate in biomass. Distribution and abundance of each
species are discussed and biological information is presented concerning
length/weight relations, populations size, proportion of the sexes, maturity
and trophic relations.
The time of year and the different
habitat which exist within the Terminos Lagoon determine the ecology of the
populations of these species. All of them show defined distribution and
abundance patterns;S. testudineus (Linnaeus) and S.
nephelus(Goode y Bean) stand out as typical species within the fish
community structure of the Terminos Lagoon.
El conocimiento de la ictiología lagunar estuarina, es uno de los aspectos importantes dentro de los estudios bioecológicos tendientes a evaluar y discutir los recursos bióticos de un área costera. El laboratorio de Ictiología y Ecología Estuarina del Instituto de Ciencias del Mar y Limnología (UNAM), ha realizado durante varios años estudios sobre ecología de comunidades de peces en la Laguna de Términos, Campeche, para obtener una interpretación integral de su ecología y ciclo natural así como conocer aspectos de la estructura de las comunidades ictiológicas. Los primeros resultados de estas investigaciones se han reportado, en los trabajos de Bravo-Núñez y Yáñez-Arancibia (1979), Amezcua Linares y Yáñez-Arancibia (1980), Yáñez-Arancibiaet al. (1980), Vargas Maldonado et al. (1981), Yáñez-Arancibia y Day (1982). La familia Tetraodontidae es considerada, por tales autores, como un componente comunitario muy importante en la ecología de la Laguna de Términos, por su diversidad, distribución frecuencia y abundancia. Distintas asociaciones de peces han sido definidas y caracterizadas en la laguna, cuya distribución ha sido altamente correlacionada con los diferentes ambientes lagunares que se presentan. La transferencia de energía entre estas comunidades distintas es muy importante especialmente en las especies comunes a todas ellas. La familia Tetraodontidae presenta especies con estas características. Si se considera además, que algunas de estas especies constituyen un recurso económico potencial de gran magnitud, que presentan heterogeneidad espacial y amplitud de sus espectros trópicos, se justifica ampliamente la necesidad de los estudios de esta familia para obtener información que contribuya a la interpretación ecológica del área y a la evaluación de los recursos. OBJETIVOSCon base en la importancia ecológica de la familia Tetraodontidae y de acuerdo a los antecedentes mencionados, el presente trabajo tiene los siguientes objetivos: 1. Determinar taxonómicamente las especies de Tetraodontidae de la Laguna de Términos. Confeccionar claves y diagnosis para su identificación, estructurando un catálogo descriptivo e ilustrado. 2. Caracterizar la distribución y abundancia de las especies en el área. 3. Obtener información biológica de cada una de las especies (distribución, abundancia, relación longitud/peso, madurez gonádica, hábitos alimenticios y espectro trópico). 4. Discutir para las especies estudiadas: los patrones de distribución y abundancia, la relación longitud/peso en un modelo de crecimiento, la madurez gonádica, el espectro trópico y la ecología trofodinámica en base a su alimentación y hábitos alimenticios. ANTECEDENTESLos estudios particulares sobre la familia Tetraodontidae son escasos. Hasta la fecha se han registrado 5 géneros, 1 subgénero y 21 especies a nivel mundial. Pero la información bibliográfica general, permite establecer que existe un total de 4 géneros, i.e., Colomesus Gill, 1844;Lagocephalus Swainson, 1839;OvoidesCuvier, 1886; SphoeroidesLacépède, 1798; y 14 especies reportadas para las costas del Atlántico occidental, de las cuales sólo 2 géneros y 5 especies se localizan en el Golfo de México, i.e., Lagocephalus laevigatus (Linnaeus, 1766); Sphoeroides spengleri(Bloch, 1782);Sphoeroides testudineus (Linnaeus, 1758);Sphoeroides nephelus (Goode y Bean, 1882) ySphoeroides dorsalis Longley, 1934. Para el área de estudio, previo a este trabajo se han reportado 2 géneros y 3 especies, i.e., Lagocephalus laevigatus (Linnaeus, 1766);Sphoeroides testudineus (Linnaeus 1758);Shpoeroides nephelus (Goode y Bean, 1882). |Aspectos generales y particulares sobre la ecología de la familia en ambientes estuarinos y costeros del Golfo de México y Mar Caribe, se han reportado en los trabajos de Linton (1905), Hildebrand y Schroeder (1928), Reid (1954 y 1955), Springer y Bullis (1956), Boschung (1957), Springer y Woodburn (1960), Tabb et al. (1862), Walker y Nelson (1964), Cervigón (1966), Randall (1967), Shipp y Yerger (1969), Swingle (197l), Carr y Adams (1973), Day et al. (1973), Cerame Vivas (1974), Perry y Perry (1974), Reséndez (1975), Goodson (1976), Smith (1976), Castro-Aguirre (1978) y Jordán et al. (1978). Publicaciones recientes sobre estudios ecológicos ictiofaunísticos en la Laguna de Términos y áreas adyacentes también incluyen en sus registros especies de Tetraodontidos; cabe citar los trabajos de: Bravo-Núñez y Yáñez-Arancibia (1979) quienes describen la estructura de las comunidades nectónicas en la Boca de Puerto Real; Amezcua Linares y Yáñez-Arancibia (1980) que estudian la ecología de las comunidades de peces e interacciones del habitat en los Sistemas Fluvio-Lagunares; Yáñez-Arancibia et al. (1980) integran una discusión del conocimiento ecológico de la laguna y de la incidencia de la estructura y función de las comunidades de peces; Vargas Maldonado et al. (1981) describen las comunidades nectónicas en áreas de Rizophora mangle y Thalassia testudinum, finalmente Sánchez-Gil et al. (1981) en un trabajo sobre la plataforma continental frente a la Laguna de Términos incluyen en sus registros 3 especies de tetraodontidos, de las cuales Sphoeroides dorsalis Longley 1934, no se encuentra en la laguna. Todos estos trabajos representan un valioso marco de referencia para discutir los objetivos aquí planteados, así como también permiten establecer la importancia que tiene esta familia de peces en los ecosistemas costeros. ÁREA DE ESTUDIOLa Laguna de Términos (Fig. 1), en el Sur del Golfo de México, es un excelente sitio para estudiar integración de procesos físicos y biológicos. No existen cambios estacionales fuertes de temperatura o luz, y el área tiene marcados gradientes físicos semipermanentes y alta diversidad de habitat estuarinos. Los vientos prevalecientes causan un flujo neto hacia dentro en la boca este y un flujo neto hacia afuera por la boca oeste, lo cual crea alta salinidad y condiciones de aguas claras en el sector oriental de la laguna. La mayor descarga de los ríos se da dentro de la parte oeste de la laguna, creando turbidez, y aguas ricas en nutrientes con baja salinidad. Un delta interior se forma en la laguna en la boca este. La mayoría de los procesos biológicos están fuertemente influenciados por estos gradientes. Extensas praderas de pastos marinos se presentan en la parte norte y oriental de la laguna, existiendo sucesión de pastos a bosques de manglar en esa área. Extensas áreas de arrecifes de ostión se localizan en la parte oeste y sur de la laguna, cerca de las bocas de los ríos. La producción de fitoplancton, niveles de clorofila y defoliación de manglares es mayor en las áreas de influencia fluvial. Las larvas y juveniles del necton ingresan a la laguna a través de la boca del este, reflejando las corrientes prevalecientes. La máxima descarga de los ríos es en octubre, durante la cual se alcanza la más alta productividad primaria y ocurre la entrada de juveniles a la laguna. Climática y meteorológicamente se caracteriza para el área un período de lluvias (junio-septiembre), un período de nortes (octubre-febrero) y un período de secas (marzo-mayo). Amplios detalles de esta síntesis pueden encontrarse en los trabajos de Phleger y Ayala-Castañares (1971), Gierloff-Emden (1977), Bravo-Núñez y Yáñez-Arancibia (1979), Amezcua Linares y Yáñez-Arancibia (1980), Vargas Maldonado et al. (1981) y Yáñez-Arancibia y Day (1982). 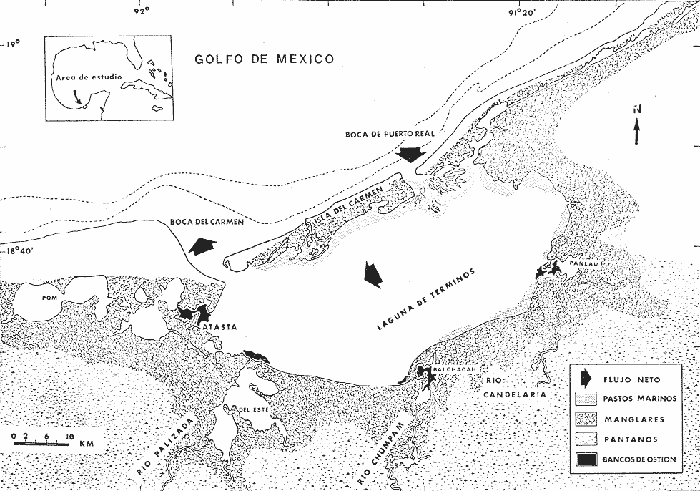 Fig. 1. Localización de la Laguna de Términos, Campeche. Se señala los rasgos geográficos y toponímicos principales y la distribución de los diferentes habitat ecológicos del sistema. Se destaca la dirección del flujo neto de las aguas y la zona de pastos marinos que indican áreas de mayor influencia marina. Se muestran los cuatro sistemas Fluvio-Lagunares principales asociados a la Laguna. (Tomada de Yáñez-Arancibia et al. 1980). MATERIAL Y MÉTODOS
ACTIVIDADES DE CAMPOLas capturas de peces se efectuaron desde el mes de julio de 1976 a marzo de 1979 de la siguiente forma: 1976 muestreo mensual de julio a diciembre 1977 capturas en los meses de enero, marzo, mayo, julio y septiembre 1978 captura en los meses de febrero, abril, junio, agosto y octubre 1979 capturas del mes de marzo 1980 muestreos mensuales durante este periodo, todavía no analizados cuantitativamente sirven además para estructurar los mapas de distribución de las especies. Todas las capturas fueron diurnas, en profundidades que no excedieron a los 4 m. Se empleó una embarcación de fibra de vidrio de 7 m de eslora, equipada con un motor de 48 HP, el arte de pesca fue una red camaronera de 10 m de largo y 9 de m de boca (abertura de trabajo de 5 m), puertas de 0.8 por 0.5 m y luz de malla de 3/4 de pulgada. Los arrastres fueron de 15 a 30 min de duración. 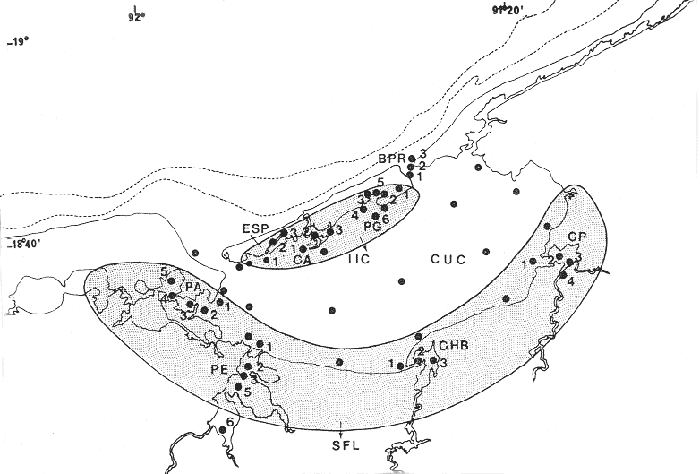 Fig. 2. Area de estudio. Localización de las zonas de muestreo. IIC = Interior de la Isla del Carmen. SFL = Sistemas Fluvio-Lagunares. CUC = Cuenca Central. En cada estación (Fig. 2) se efectuaron mediciones de: temperatura, salinidad, oxígeno, profundidad y transparencia. También se realizaron observaciones de: tipo de sustrato, vegetación sumergida y circundante, macrofauna béntica, mareas y datos climáticos. El material colectado se fijó en formalina al 10% neutralizada con borato de sodio. A los ejemplares capturados se les realizó una incisión ventral para la fijación del contenido estomacal y gónadas. MATERIAL DE ESTUDIODe los 22 cruceros realizados (173 colectas), se obtuvieron un total de 636 ejemplares, correspondientes a 6 especies de Tetraodontidae, distribuidas como se muestra en la tabla 1. 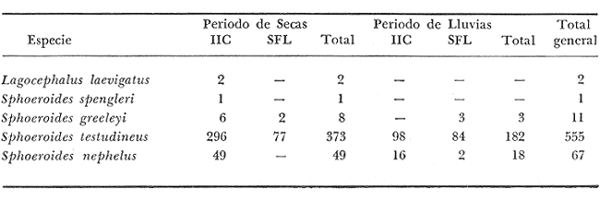 TABLA 1 RELACIÓN DEL NÚMERO DE EJEMPLARES CAPTURADOS, POR ESPECIE, ÉPOCA CLIMÀTICA Y LOCALIDAD. ACTIVIDADES DE LABORATORIOLos ejemplares fueron lavados, separados, reetiquetados y envasados en frascos de vidrio con alcohol metílico al 70%. La posición taxonómica de las especies fue determinada por los métodos convencionales y el empleo de literatura básica, principalmente los trabajos de Jordan y Evermann (1896-1900), Meek e Hildebrand (1923-1928), Cervigón (1966), Randall (1967), Shipp y Yerger (1969), Shipp (1972), Gallaway et al. (1972), Goodson (1976), Dickson y Moore (1977), Guitart (1977), Fischer (1978) y Tyler (1980). Las mediciones de longitud fueron hechas con un ictiómetro convencional de 50 cm y las de peso sobre material fijado, en una balanza Ohaus Triple Beam de 0. 1 g de precisión y 2,610 g de capacidad. RELACIÓN LONGITUD/PESOPara el análisis de relación longitud/peso, se consideró la relación entre el peso en gramos y la longitud total en milímetros. La función resultante es una exponencial, por ser la relación que existe entre una dimensión lineal y otra cúbica. Se utilizó el análisis de regresión de potencia, para determinar las constantes a y b de la ecuación: P = a Lb (1) donde: P = peso en gramos L = longitud en milímetros a y b = constantes a determinar en la que se calcula el coeficiente de regresión R. DETERMINACIÓN DE SEXO Y ANÁLISIS GONÁDICOEn general para muchos peces no existen caracteres sexuales externos evidentes por lo que es necesario proceder a disectarlos para reconocer el sexo. En las hembras las gónadas son de mayor tamaño y de forma de saco, presentando una luz central. En los machos las gónadas son consistentes, es decir no presentan luz. Para determinar fases de madurez gonádica considerada en el presente trabajo, se siguió el criterio de Nikolsky (1963). RELACIONES TRÓFICASSe examinaron un total de 87 ejemplares de todas las especies, distribuidos como se muestra en la tabla 2. 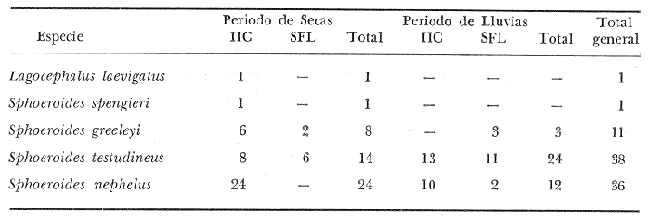 TABLA 2 RELACION DEL NÚMERO DE ESTOMAGOS ANALIZADOS, POR ESPECIE, ÉPOCA CLIMÁTICA Y LOCALIDAD. El examen de los estómagos y el estado del pez se hizo siguiendo el criterio de Laevastu (1971). Para el análisis de las fases de digestión, del contenido estomacal se siguieron los criterios de Carranza (1969) y Yáñez-Arancibia (1975). De acuerdo al tamaño de los estómagos, al tamaño y tipo de alimento y el grado de digestión, es necesario combinar entre sí los diferentes métodos para obtener una información más clara sobre la preferencia alimenticia en determinadas áreas. Se eligieron los métodos volumétricos, numérico, de frecuencia, el índice de importancia relativa de Pinkas et al. (1971), así como el índice de importancia relativa de Yáñez-Arancibia et al. (1976). La descripción detallada de todos los métodos se encuentran en la referencia de Yáñez-Arancibia et al. (1976) La descripción detalla del índice de Pinkas et al. (1971) y de Yáñez-Arancibia et al. (1976) se encuentran en el trabajo de Lara-Domínguezet al. (1981). ABREVIATURAS UTILIZADAS EN TABLAS Y GRÁFICASAn = Anfípodos BP = Boca de Puerto Real CA = Bajos del Cayo CB = Colonias de briozoarios CHB = Sistema Chumpán-Balchacah Ci = Cirrípedos Cn = Conchilla Cp = Sistema Candelaria-Panlau CUC = Cuenca Central Eq = Equinodermos ESP = Estero Pargo Fo = Foraminíferos Ga = Gasterópodos HI = Huevos de invertebrados Is = Isópodos IIC = Interior de la Isla del Carmen LM = Larvas de moluscos MNG = Puntilla Manigua MOND = Materia orgánica no determinable Ne = Nemátodos Ol = Oligoquetos Os = Ostrácodos PA = Sistema Pom-Atasta Pe = Pelecípodos PE = Sistema Palizada-del Este PG = Punta Gorda Pi = Picnogónidos PEC = Sistema Palizada-del Este. Boca del Carmen RC = Restos de crustáceos RV = Restos de vegetales SFL = Sistemas Fluvio-Lagunares Ta = Tanaidáceos RESULTADOS
FAMILIA TETRAODONTIDAE LINNAEUS
"Peces globo"Peces pequeños de talla moderada. Cuerpo oblongo o alargado generalmente comprimidos, capaz de inflarse tanto con aire como con agua hinchando el vientre. Piel áspera sin escamas típicas, generalmente cubierta con numerosas espinitas a menudo presentes en el dorso, vientre y flancos; algunas especies con apéndices dérmicos en el dorso, en los flancos y ocasionalmente en la barba. Línea lateral cuando se presenta es indiferenciada, forma un patrón complejo unido por debajo de los flancos cerca de la cabeza. Rostro largo, delgado y romo. Perfil cóncavo o convexo según la especie, hocico amplio, boca pequeña terminal, mandíbulas con dientes fusionados, modificados en forma de placa con una sutura en su parte media, 4 en total, 2 inferiores y 2 superiores. Cabeza larga y roma. Ojos en posición anterior de la cabeza. Nostrilos presentes y funcionales. Aberturas branquiales pequeñas sin una cubierta distinta, semejantes a una ranura, anteriores a las aletas pectorales con pequeñas branquiespinas. Sin aletas pélvicas. Dorsal y anal cortas insertadas posteriormente, sin espinas, con 7 a 15 radios suaves. Pectorales cortas o anchas. Caudal usualmente truncada o ligeramente redondeada con un margen posterior o con extensiones pequeñas en los lóbulos superior o inferior. Vejiga gaseosa presente. Las especies pertenecientes a esta familia presentan el cuerpo plateado o pigmentado, dorso oscuro, a menudo con patrones de reticulaciones o marcas vermiculadas claras. Flancos con puntos negros redondeados. Vientre pálido de color amarillo a blanco. Aletas pálidas algunas especies con la caudal bandeada. Su capacidad de inflarse es un mecanismo de defensa que los previene de ser ingeridos por predadores, se impulsan a través del agua con un aleteo, como ventilador, con sus aletas dorsal y anal. Todas las especies son preferentemente carnívoras. Existen reportes para el área oeste del Atlántico y especialmente en el Japón de que la carne de filete de muchas especies es de buen sabor y a menudo se consume localmente. No obstante muchas especies son tóxicas y su consumo ha causado envenenamientos y en algunas ocasiones muertes. La toxina se localiza principalmente en ciertos órganos viscerales, especialmente en el hígado y la carne se contamina por contacto con tales órganos. La cantidad de toxina presente y su efecto varía según la especie. La "ciguatera" es una de las manifestaciones causadas por las toxinas. Ninguno de los Tetraodóntidos son peces de arrecife, pero algunos se presentan ocasionalmente en esos ecosistemas y se les puede encontrar en áreas adyacentes. Son habitantes de mares cálidos y tropicales, la mayoría habita aguas someras de la costa, penetran en aguas salobres y algunas veces llegan hasta las desembocaduras de los ríos. Las gónadas de la familia Tetraodontidae son simétricas, el color varía según el estadio sexual en el que se encuentren, se diferencian de los riñones por el color, ya que éstos son rojo-café de menor tamaño, y se encuentran pegados al dorso o ligeramente desplazados, pero nunca presentan luz; se diferencian de los ciegos, por ser éstos más pequeños y desembocar siempre en el estómago. Existe una clara diferenciación en las gónadas de hembras y machos. Siendo las gónadas de machos triangulares y sin luz, y las de las hembras redondas y con luz central. Como anteriormente se menciona, el color cambia según el estadio en que se encuentren y éste se conserva independiente de que se trate de machos o hembras. En los individuos inmaduros, el color es amarillo, de tamaño reducido, los huevecillos no se distinguen a simple vista, en las hembras forman sinuosidades. En los individuos maduros las gónadas están desarrolladas ocupan gran parte de la cavidad desplazando organelos, de consistencia dura, son amarillas pálidas ligeramente rosadas, en los machos presentan además pigmentaciones en los extremos; con huevecillos distinguibles a simple vista de color blanco y miden de 1 a 5 mm. En los individuos desovados se observan diferencias notorias en machos y hembras, las gónadas de los machos son blancas y pigmentadas o no, de consistencia flácida, sumamente alargadas, mientras que en las hembras son rosadas, con mucha luz y dan el aspecto de globos desinflados que pueden o no presentar sinuosidades, donde sólo se observan residuos. CLAVE ARTIFICIAL PARA DETERMINAR LOS GÉNEROS Y ESPECIES DE TETRAODONTIDAE PRESENTES EN LA LAGUNA DE TÉRMINOSOtra especie del sur del Golfo de México reportada en el trabajo de Sánchez-Gil et al. 1981, esSphoeroides dorsalis Longley, no penetra a la Laguna de Términos. Se diferencia de las especies referidas en esta clave por los siguientes caracteres. Dorso sin reticulaciones, flancos de la cabeza con reticulaciones pálidos de forma irregular. Con un par de aletillas oscuras enfrente de la dorsal. Flancos con una línea regular de 1 a 5 puntos negros redondos. 1. Cuerpo oblongo. Aletas dorsal y anal pequeñas, compuestas de 6 a 8 radios suaves. Caudal redondeada o truncada de márgenes planos, algunas especies con radios ligeramente alargados, pero nunca cóncava o alunada. Lados bajos del pedúnculo sin pliegues cutáneos. Orificio nasal con una sola abertura pequeña............2 1' Cuerpo comparativamente alargado. Aletas dorsal y anal ligeramente largas, compuestas de 12 a 15 radios suaves. Caudal definitivamente cóncava o lunada. Lados bajos del pedúnculo con pliegues cutáneos. Orificio nasal con dos aberturas a cada lado..................... Lagocephalus laevigatus (Linnaeus) 2. Dorso generalmente con reticulaciones pálidas.............................3 2' Dorso sin reticulaciones. Con aletillas oscuras en la parte posterior. Aletas pectorales con 13 radios suaves. Espinitas cubriendo un área pequeña del dorso. Flancos con una línea regular de 11 a 14 puntos negros redondos................. Sphoeroides spengleri(Bloch) 2'' Sin aletillas oscuras en la parte posterior. Aletas pectorales con 12 radios suaves. Espinintas cubriendo la mitad del dorso, la otra mitad con escamas blandas no ocificadas. Flancos son un patrón de puntos negros uniformes......... Sphoeroides greeleyi (Gilbert) 3. Con reticulaciones pálidas de forma regular, ocupando una posición transversa en el dorso. Flancos totalmente cubiertos por grandes manchas oscuras redondeadas, ligeramente separadas unas de otras. Espinitas cubriendo la mayor parte del dorso, pero usualmente hundidas y no sensibles al tacto. Aletas pectorales con 15 radios........................ Sphoeroides testudineus(Linnaeus) 3' Con reticulaciones pálidas de forma irregular y variable en el dorso y a los lados. Flancos con una hilera irregular de manchas oscuras redondeadas. Espinitas cubriendo una porción variable del dorso, ocasionalmente ausentes pero a veces presentes como escamas blandas. Aletas pectorales con 14 radios y una mancha oscura notoria en la axila.......................... Sphoeroides nephelus (Goode y Bean) Género: Lagocephalus Swainson, 1839 Lagocephalus laevigatus (Linnaeus, 1766) N.v.: "Pez globo", "Tambor", "Xpu"
Tetraodon laevigatus Linnaeus, Syst. Nat., Ed. 12, 1766: 411 (Charleston, Carolina del Sur). Diagnosis. Cuerpo alargado, delgado. Profundidad 3.7 a 16.8 en la LP. Cabeza larga 2.6 a 12.9 en la LP; hocico romo 1.7 a 2.0 en la cabeza y los ojos grandes 2.7 a 4.8; interorbital muy amplio 2.4 en la longitud cefálica. Perfil convexo. Piel del dorso y flancos lisa, vientre con espinitas depresibles. D 12-15; A 11-13. Dorsal y anal largas y falcadas, el último radio muy corto. Pectorales cortas 1.6 a 2.0 con 17 radios. Caudal profundamente lunada. Margen inferior del pedúnculo caudal con pliegues cutáneos. Color. Dorso verde grisáceo, sin reticulaciones. Flancos plateado metálico, sin manchas redondeadas oscuras. Vientre pálido. Caudal oscura. Dorsal y pectorales con una banda ligeramente oscura en la base. Los juveniles presentan tres bandas anchas y oscuras transversales en el dorso, dispuestas sobre el pedúnculo caudal, en la base de la dorsal y a la altura de las pectorales; otras tres bandas similares pero mucho más delgadas sobre la cabeza. Distribución. En el Atlántico desde Massachussetts EUA hasta Argentina incluyendo Golfo de México. En el área de estudio: localidades BPR y PG (Fig. 3). 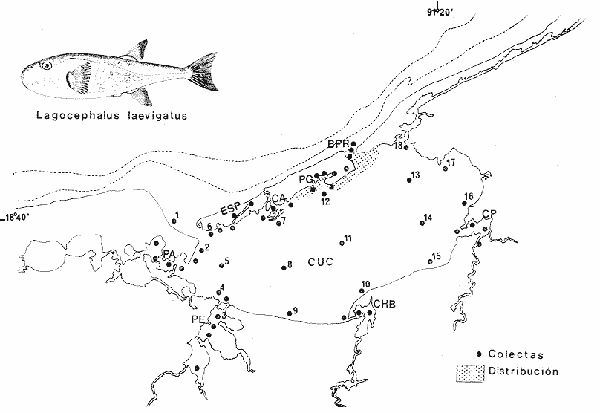 Fig. 3. Mapa de distribución de Lagocephalus laevigatus en la Laguna de Términos, en relación a las estaciones de colecta en las distintas localidades. Material examinado. Dos ejemplares de 89 y 442 mm de longitud total. Colección de referencia depositada en el Laboratorio de Ictiología y Ecología Estuarina del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. Catálogo ICMI, P/F 0.001928/1929 (Camp.) Observaciones taxonómicas. Algunas consideraciones taxonómicas sobre Lagocephalus laevigatus (Linnaeus) en la costa Atlántica y Golfo de México han sido hechas por: Jordan y Evermann (1896), Meek e Hildebrand (1925), Cervigón (1966), Randall (1967), Gallaway et al. (1972), SIC (1976), Dickson y Moore (1977), Guitart (1978) Shipp y Yerger (1969) y Tyler (1980). Esta especie se distingue de otros tetraodóntidos principalmente por no presentar manchas redondas oscuras en los flancos, por que tiene la piel del dorso y flancos completamente lisa y carece de espinitas, escamas, reticulaciones y apéndices dérmicos en el dorso; no obstante, el vientre está cubierto de espinitas a menudo desarrolladas. Difiere de las especies del géneroSphoeroides (Lacépède) por las características antes mencionadas, además de tener la aleta dorsal con 13 a 14 radios, la cual es cóncava o lunada y el color que es característico. Se cuenta con un reporte previo de Bravo-Núñez y Yáñez-Arancibia (1979) para la Laguna de Términos, Campeche. Observaciones biológicas y ecológicas.Abundancia. Existe un solo registro y consiste de dos ejemplares colectados en la Boca de Puerto Real y Punta Gorda, durante la época de secas. La especie es rara y poco frecuente en el ecosistema lagunar estudiado y por lo tanto no, es un componente estructural importante de las comunidades de peces en el área. Relación longitud/peso y frecuencia de tallas. No se efectuó la relación debido a que solo se colectaron dos ejemplares de 89 y 442 mm en el área. Madurez. Se analizó el ejemplar colectado en el área de Punta Gorda, resultando una hembra en estadio VII. Alimentación. Se analizó el estómago del ejemplar colectado en el área de Punta Gorda, el cual estaba totalmente destruido y los restos del contenido dispersos, por lo que no, se analizó el contenido por los métodos convencionales. Se procedió a diferenciar los restos por medio de una lupa, encontrándose: restos de vegetales, restos de peces y materia orgánica en gran proporción. Género:SphoeroidesLacépéde 1798Spheroides nephelus (Goode y Bean, 1882) N.v.: "Pez globo", "Tambor", "Xpu"
Tetraodon nephelus Goode y Bean, Proc. U. S. Nat. Mus., 1882, 5:412 (Río Indio y Penzacola, Florida). Diagnosis. Cuerpo elongado. Profundidad 3.5 a 5.6 en la LP, Cabeza grande 2.4 a 2.8 en la LP; hocico 1.6 a 2.2 en la cabeza y los ojos 2.6 a 4.8; interorbital pequeño con una banda oscura 6.2 a 11.0 en la longitud cefálica. Piel del dorso y vientre con espinitas notorias al tacto pero pueden faltar, flancos suaves. Pectorales 2.0 a 3.2 en la longitud cefálica, 14 radios suaves. Color. Dorso verde y café pizarra en adultos, con reticulaciones pálidas de forma irregular, más notorias a las flancos de la cabeza. Flancos con hileras irregulares de manchas redondeadas oscuras que decrecen en tamaño hacia la caudal, dos puntos oscuros muy conspicuos uno a cada lado de las axilas de las pectorales. Vientre pálido. Caudal con dos bandas ligeramente grises en la parte distal y basal. Otras aletas pálidas. Distribución. En el Atlántico, desde Florida, EUA y Bahamas hasta Brasil, incluyendo Golfo de México En el área de estudio: localidades BPR, PG, CA, ESP y PA (Fig. 4). Material examinado. 67 ejemplares de 46 a 150 mm de longitud total. Colección de referencia depositada en el Laboratorio de Ictiología y Ecología Estuarina del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. Catálogo ICML P/F 0.001926/1927 (Camp.). 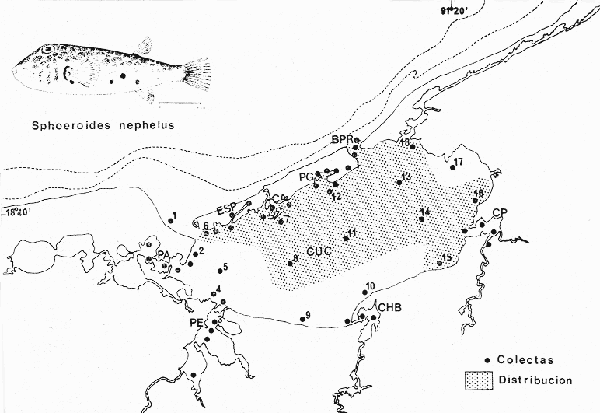 Fig. 4. Mapa de distribución de Sphoeroides nephelus en la Laguna de Términos, en relación a las estaciones de colecta en las distintas localidades. Observaciones taxonómicas. Algunas consideraciones taxonómicas sobre Sphoeroides nephelus (Goode y Bean) en la costa del Atlántico y Golfo de México han sido hechas por: Bohlke y Chaplin (1968), Goodson (1976), Dickson y Moore (1977), Greenberg (1977), Shipp y Yerger (1969) y Tyler (1980). Es la especie más coloreada del género. Las áreas oscuras del dorso son verdes en los juveniles y café oscuro en los adultos. Las partes bajas de los flancos de la cabeza hasta antes de las aberturas branquiales, muestran un color amarillo pálido combinado con tonos azul claro. A lo largo se marcan reticulaciones de color más pálido que se continúan en el dorso en forma irregular. Las manchas oscuras redondeadas que separan el vientre del dorso, decrecen en tamaño hacia la caudal, generalmente por debajo de las axilas de las pectorales y por detrás de éstas tiene dos manchas oscuras casi negras. Se distingue de S. testudineus (Linnaeus) y S. greeleyi (Gilbert) en las reticulaciones de forma irregular y las manchas oscuras, la presencia de espinitas cubriendo la mayor parte del dorso (pero ausentes en el material analizado) y con escamitas blandas. Difiere de S. spengleri(Bloch) y S. dorsalis Longley en que carece del par de apéndices dérmicos en el dorso. A diferencia deS. parvus (Shipp y Yerger) presenta las manchas oscuras de las axilas de las pectorales. Tyler (1980) menciona que S. nephelus ha sido reportado como S. marmoratus(Ranzani) en el Atlántico de América (Jordan y Evermann 18961900, Meek e Hildebrand 1925, Love-McConnell 1962). Por esto fue citado S. marmoratus para la Laguna de Términos por Bravo-Núñez y Yáñez-Arancibia (1979), Vargas Maldonado et al. (1981), Yáñez-Arancibiaet al. (1980, 1981, 1982). Por lo tanto debe entenderse que cuando esos autores mencionan a S. marmoratus (Ranzani) se trata de S. nephelus(Goode y Bean). La especie ha sido reportada previamente para la Laguna de Términos por: Yáñez-Arancibiaet al. (1980 y 1982) y Vargas Maldonado et al. (1981). 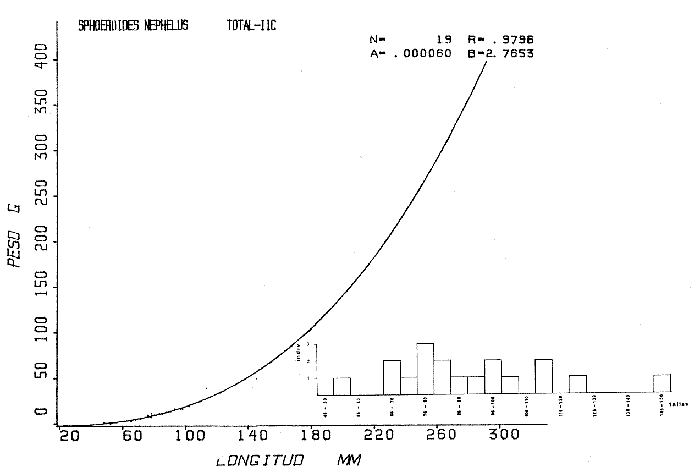 Fig. 5. Curva de regresión de la relación longitud/peso de Sphoeroides nephelus. Se señala la distribución de frecuencia de tallas. Total del Interior de la Isla del Carmen. 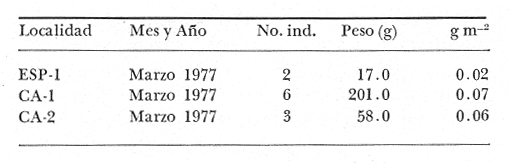 TABLA 3 SPHOEROIDES NEPHELUS ABUNDANCIA ESTIMADA DURANTE LA ÉPOCA DE SECAS (1977) EN EL INTERIOR DE LA ISLA DEL CARMEN (IIC) 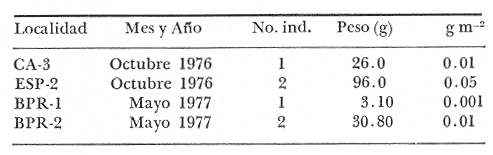 TABLA 3 ABUNDANCIA ESTIMADA DURANTE LA ÉPOCA DE LLUVIAS (1976-1977) EN EL INTERIOR DE LA ISLA DEL CARMEN (IIC) Observaciones biológicas y ecológicas. Abundancia. Se observa que para esta especie la biomasa cambia dependiendo de la época climática (Tabla 3). Observándose el valor más alto de 0.07 gm-² con 6 ejemplares para la época de secas y el valor más bajo 0.001 gm-² con un ejemplar para el periodo de lluvias. La especie es poco abundante, aun cuando es la segunda importante de los tetraodóntidos de la laguna. 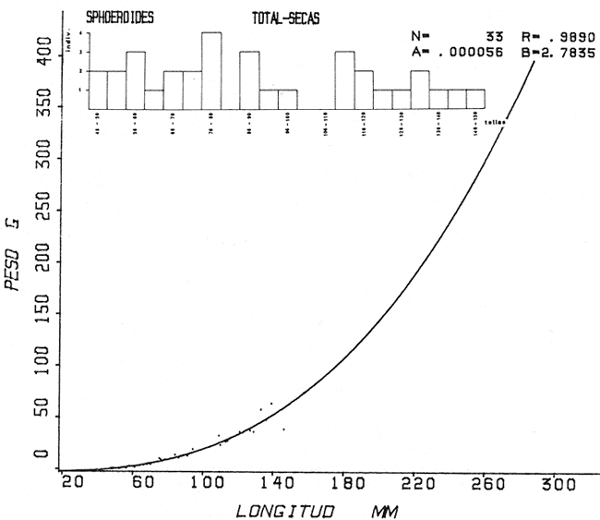 Fig. 6. Curva de regresión de la relación longitud/peso de Sphoeroides nephelus. Se señala la distribución de frecuencia de tallas. Total del Periodo de Secas. Relación longitud/peso y frecuencia de tallas. De acuerdo a la ecuación los valores calculados para las constantes a y b son: 0.000060 y 2.7653 respectivamente y el coeficiente de regresión R tiene un valor de 0.9796. El rango de tallas para el total de los ejemplares de una población del litoral interno de la Isla del Carmen es de 51 a 151 con una moda a los 80 mm. Estos resultados se muestra graficados en la figura 5. De acuerdo a la ecuación, los valores calculados para las constantes a y b y el coeficiente de regresión R se comparan para el total de las dos épocas climáticas así como el análisis de tallas en las figuras 6 y 7. Madurez. En la distribución de las fases de maduración sexual para esta especie, se encontró que para el total de la colección nunca aparecen individuos en reproducción. Se encontraron 4 ejemplares indiferenciados en sexo y en maduración, en el interior de la Isla del Carmen para la época de lluvias, cuyos rangos de longitud van de 46 a 62 mm. Dependiendo de la localidad se observa que solo se encuentran dos hembras en los estadios I y II para los Sistemas Fluvio-Lagunares en la época de lluvias, y el resto de los ejemplares analizados corresponden al área del Interior de la Isla del Carmen (Figs. 8 y 9). La figura 9 muestra la relación de la madurez gonádica y la talla de los individuos, es importante hacer notar que sólo se registraron hembras maduras con tallas de 95 a 147 mm. Comparando las dos épocas climáticas, se observa que la mayor frecuencia corresponde a la época de secas. Con respecto a la fase de maduración, se tiene que: la fase I es la mejor representada para el total de los ejemplares (juveniles); en la fase II no se, encontraron machos; en la fase III las hembras ocupan la mayor frecuencia y en la fase IV sólo se encontraron hembras (Fig. 10). 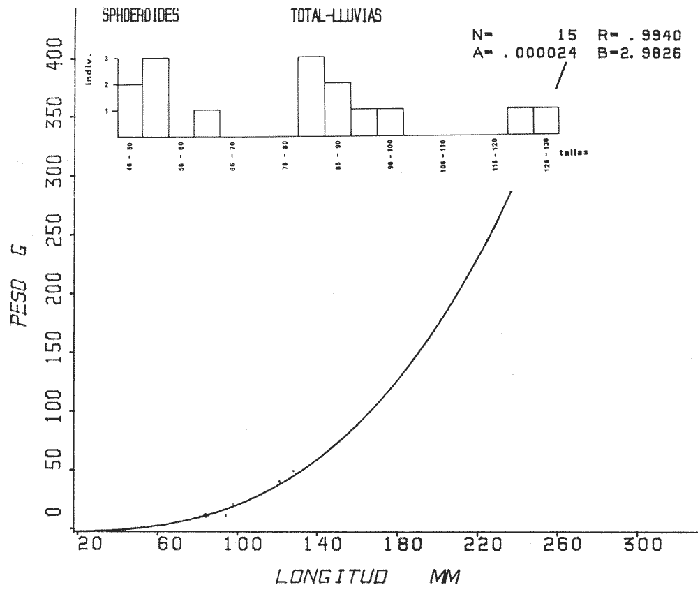 Fig. 7. Curva de regresión de la relación longitud/peso de Sphoeroides nephelus. Se señala la distribución de frecuencia de tallas. Total del Periodo de Lluvias. 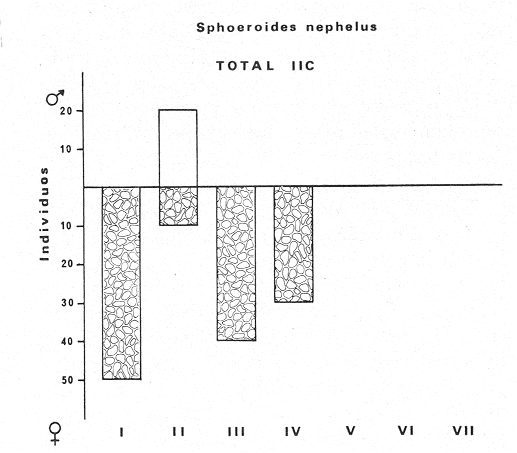 Fig. 8. Fases de desarrollo sexual y proporción de sexos de Sphoeroides nephelus para el Total del Interior de la Isla del Carmen.  Fig. 9. Relación de la longitud total y las fases de desarrollo sexual de Sphoeroides nephelus. Total del Interior de la Isla del Carmen. 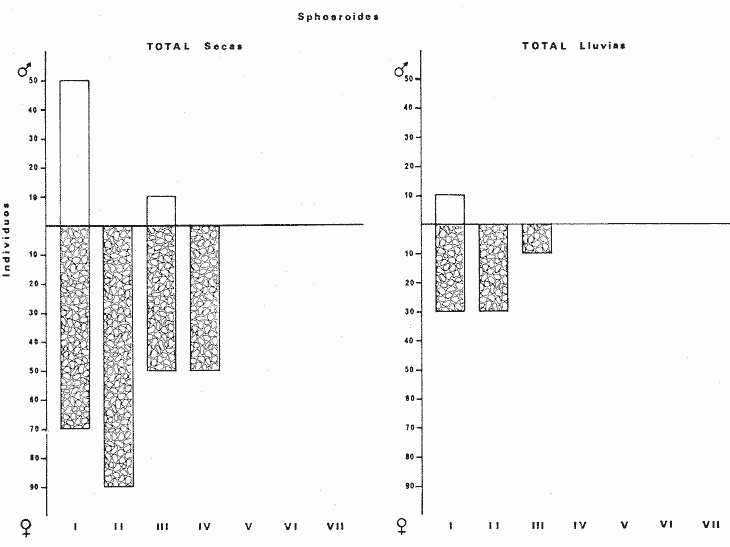 Fig. 10. Fases de desarrollo sexual y proporción de sexos de Sphoeroides nephelus para ambos periodos climáticos, independiente de la localidad. Se observa que el total de ejemplares sólo alcanzó la fase IV (en maduración). De la relación macho: hembra se observa que el total de la población guarda una proporción de 1:4.1 que permanece constante en la época de secas y disminuye a 1: 3.5 en la época de lluvias (Fig. 10). La figura 11 muestra la relación de la madurez gonádica y la talla de los individuos. Este análisis es necesario para determinar el tamaño al cual los ejemplares alcanzan la madurez gonádica y son aptos para reproducirse. En este caso las hembras empiezan a madurar a los 83 mm y los machos a los 85 mm. La figura 11 muestra la relación total del desarrollo sexual de la población, independientemente del sexo. Alimentación. El estudio de las relaciones tróficas se basó en 36 estómagos. 13 estómagos de peces del Interior de la Isla del Carmen y 23 en función de las épocas climáticas. El espectro trópico (Fig. 12) señala que numéricamente el principal alimento lo constituyen Oligoquetos, Huevos de Invertebrados, e Isópodos y en menor proporción Pelecípodos y Anfípodos. Por la frecuencia, materia orgánica, Restos de Vegetales y Pelecípodos constituyen el alimento importante. Volumétricamente, materia orgánica, Oligoquetos y Pelecípodos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestran que la materia orgánica, Oligoquetos, Pelecípodos, Huevos de Invertebrados e Isópodos presentan los índices más elevados. En relación a la figura 12 se puede establecer que el alimento principal es materia orgánica; el alimento secundario Pelecípodos, Restos de Vegetales y Restos de Crustáceos; el alimento circunstancial está representado por los otros grupos trópicos encontrados. 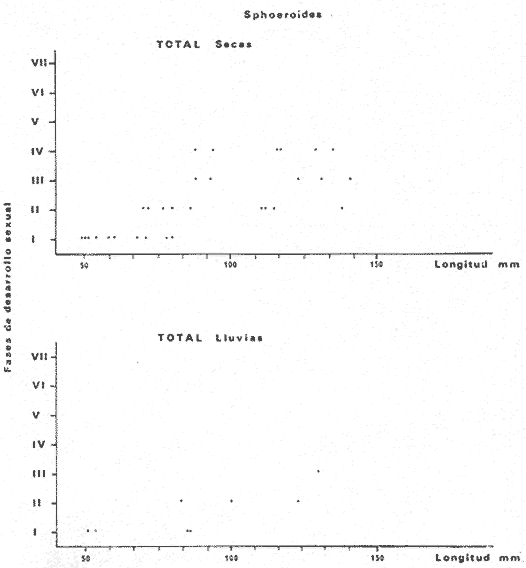 Fig. 11. Relación de la longitud total y las fases de desarrollo de Sphoeroides nephelus para ambos periodos climáticos, independiente de la localidad. Total del periodo de secas. El espectro trópico (Fig. 13) señala que numéricamente el principal alimento lo constituye los Restos de Crustáceos y en menor proporción Pelecípodos, Gasterópodos, Picnogónidos e Isópodos. Por la frecuencia, materia orgánica, Pelecípodos, Restos de Crustáceos y Restos de Vegetales constituyen el alimento importante. Volumétricamente, materia orgánica y Restos de Crustáceos son los grupos trópicos destacados. El análisis combinado, de estos datos para cuantificar los índices de importancia relativa, muestran que la materia orgánica y los Restos de Crustáceos presentan los índices más elevados. En relación a la figura 9 se puede establecer que el alimento principal es la materia orgánica; el alimento secundario lo constituyen Restos de Crustáceos, Pelecípodos y Restos de Vegetales; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. Total del periodo de lluvias. El espectro trópico (Fig. 14) señala que numéricamente el principal alimento lo constituyen los Foraminíferos y en menor proporción Gasterópodos y Restos de Crustáceos. Por la frecuencia, materia orgánica, Pelecípodos y Restos de Crustáceos constituyen el alimento importante. Volumétricamente, materia orgánica, Restos de Crustáceos y Equinodermos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestra que materia orgánica, Restos de Crustáceos, Foraminíferos, Gasterópodos y Pelecípodos presentan los índices más elevados. En relación a la figura 14, se puede establecer que el alimento principal es la materia orgánica y los Restos de Crustáceos; el alimento secundario lo constituyen Pelecípodos, Gasterópodos y Restos de Vegetales; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. 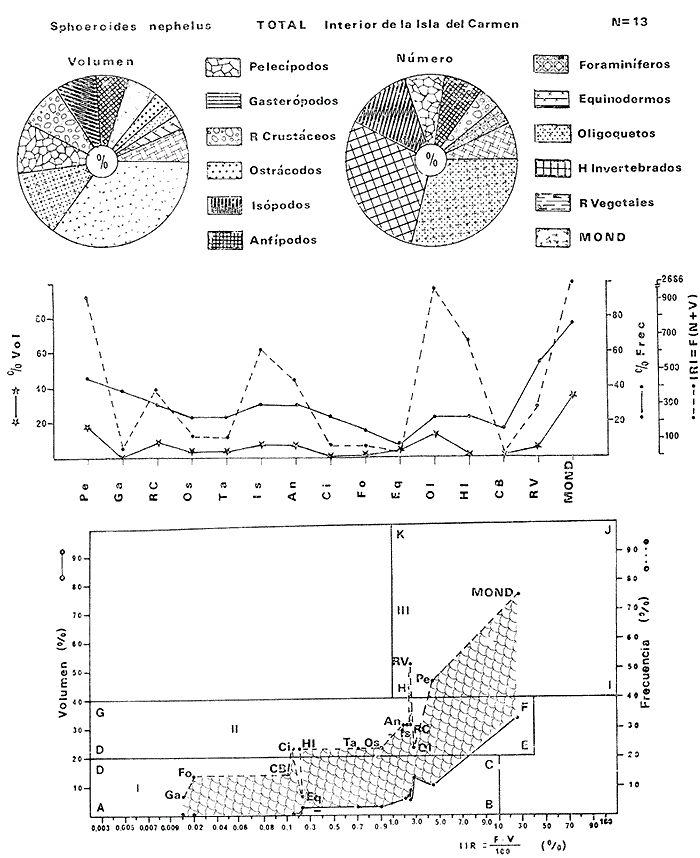 Fig. 12. Espectro tráfico de Sphoeroides nephelus. Total del interior de la Isla del Carmen. 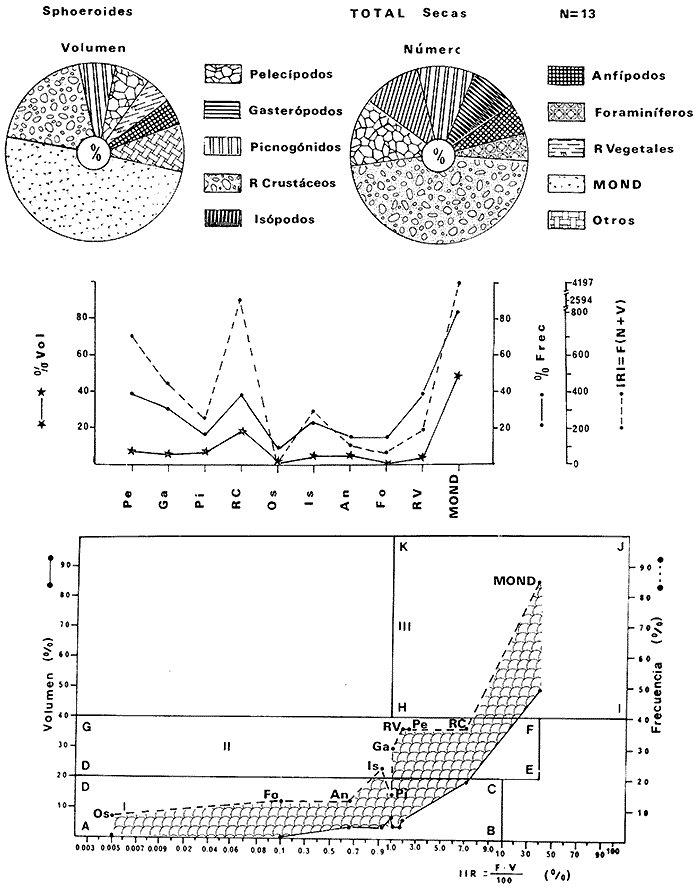 Fig. 13. Espectro trófico de Sphoeroides nephelus en el Total del Periodo de Secas. 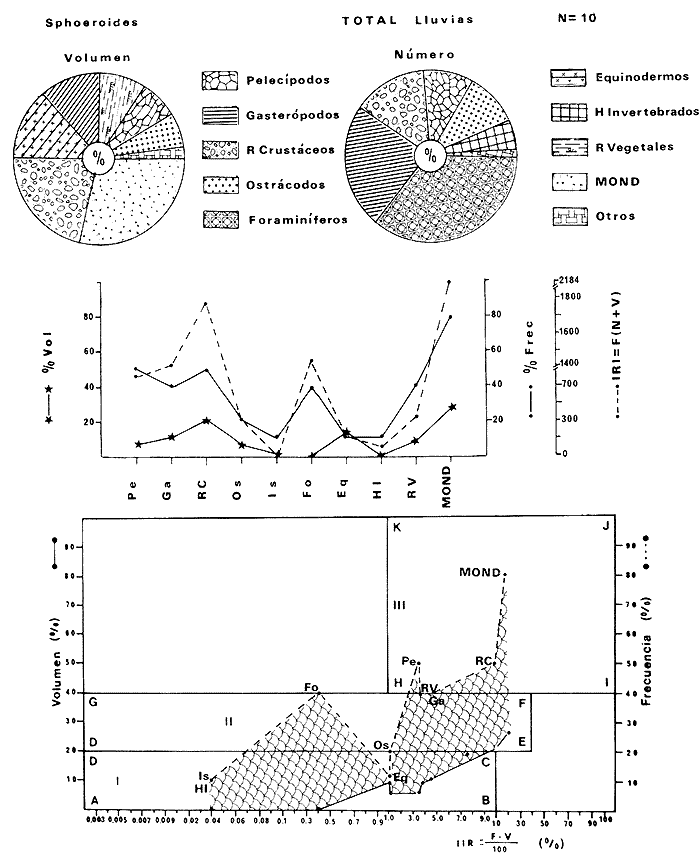 Fig. 14. Espectro trófico de Sphoeroides nephelus en el Total del Periodo de Lluvias. El análisis estadístico del estudio de la relación gravimétrica del alimento ingerido porSphoeroides nepehlus (Fig. 15), indica que el índice es notablemente grande tanto en juveniles como en adultos, es decir que el rango de variación es muy amplio, mostrando que no existe correlación entre el índice y el peso total del ejemplar comportándose con altas necesidades alimenticias durante toda su vida. La figura 15 muestra esto para una población del litoral interno de la Isla del Carmen. 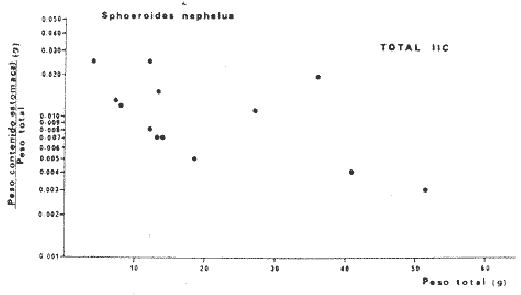 Fig. 15. Dispersión gravimétrica del alimento en relación al tamaño en peso de Sphoeroides nephelus. Total del Interior de la Isla del Carmen. La relación gravimétrica del alimento ingerido porSphoeroides nephelus se muestra en la figura 16. El análisis estadístico, indica un comportamiento diferente comparando los dos periodos climáticos, donde se observa que para el periodo de secas se comporta con la misma intensidad alimenticia en diferentes tallas, ya que el índice es notablemente grande tanto para juveniles como para adultos. El rango de variación es muy amplio y muestra que no existe correlación entre el índice y el peso total de los ejemplares. Por otra parte, para el periodo de lluvias el índice es notablemente grande en los estadios juveniles y éste decrece linealmente conforme aumenta de tamaño, la especie. El requerimiento alimenticio durante la época de lluvias es notablemente mayor en los juveniles. 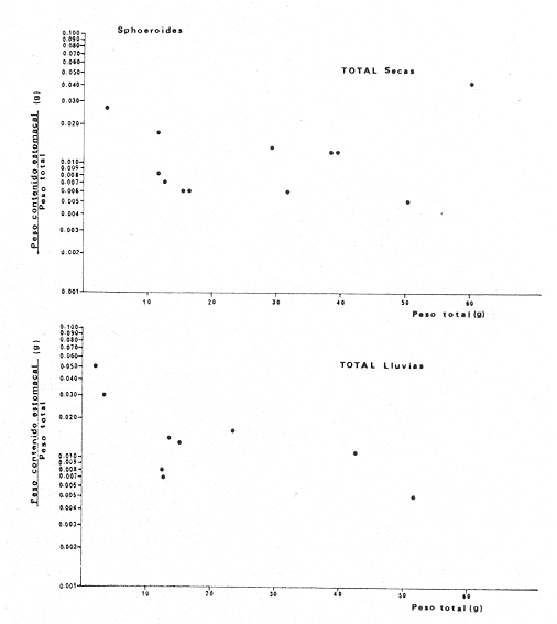 Fig. 16. Dispersión gravimétrica del alimento en relación al tamaño en peso de Sphoeroides nephelus. Periodo de Secas N = 13. Periodo de Lluvias N = 10. Sphoeroides spengleri (Bloch, 1782) N.v.: "Pez globo", "Tambor", "Xpu"
Tetraodon spengleri Bloch, Icth., 1782, 1:135, lám. 144 (Indias Orientales). Diagnosis. Cuerpo alargado, no comprimido. Profundidad 3.6 a 6.2 en la LP. Cabeza comprimida 2.2 a 3.1 en la LP; hocico romo proyectante 1.6 a 2.7 en la cabeza y los ojos 4.0 a 5.5; interorbital cóncavo varía con la edad 5.3 a 6.0 en la longitud cefálica. Piel del dorso, flancos y vientre cubierta con espinitas. Flancos con cirros dermales, hocico y cola desnudos; con un par de apéndices dérmicos en la parte posterior del dorso, localizados cerca del origen de la dorsal a cada lado. D 7 A 6. Pectorales 2.7 a 3.2 con 13 radios. Color. Dorso verde oscuro, sin reticulaciones, con áreas pálidas. Flancos con una hilera regular de 11 a 14 manchas negras redondeadas, separadas de un área oscura del dorso. Vientre pálido. Caudal con dos bandas oscuras transversales, una peduncular y otra distal. Otras aletas pálidas. Distribución. Ambos lados del Atlántico en el margen occidental: desde Massachusetts, EUA, hasta Santos, Brasil, incluyendo Golfo de México. En el área de estudio: localidad BPR (Fig. 17). Material examinado. Un ejemplar de 110 mm de longitud total. Colección de referencia depositada en el Laboratorio de Ictiología y Ecología Estuarina del Inst. de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. Catálogo ICML-P/F 0.001955/1955 (Camp.). Observaciones taxonómicas. Algunas consideraciones taxonómicas sobreSphoeroides spengleri (Bloch) en la costa del Atlántico y Golfo de México han sido hechas por: Jordan y Evermann (1896), Meek e Hildebrand (1925), Cervigón (1966), Randall (1968), Bohlke y Chaplin (1970), Dahl (1971), Gallaway et al. (1972), Goodson (1976), Dickson y Moore (1977), Guitart (1977), Greenberg (1977), Shipp y Yerger (1969) y Tyler (1980). Un carácter distintivo de esta especie que la diferencia de las otras es que presenta varios pares de apéndices dérmicos en el dorso, que varían en número de un individuo a otro. Difiere de S. testudineus (Linnaeus) y S. nephelus (Goode y Bean) en la característica mencionada y por carecer del patrón de reticulaciones en el dorso. De S. dorsalis Longley se diferencia por carecer de reticulaciones a los lados de la cabeza, por presentar una hilera definida de 5 a 8 manchas oscuras redondeadas en los flancos y más de un par de apéndices dérmicos en la porción posterior del dorso. Éste es el primer registro de Sphoeroides spengleri (Bloch) dentro de la Laguna de Términos Campeche, basado en un ejemplar colectado en el área de la Boca de Puerto Real. Observaciones biológicas y ecológicas. Abundancia. La especie es rara y poco frecuente. No juega un papel importante en la estructura de las comunidades de peces en el área. 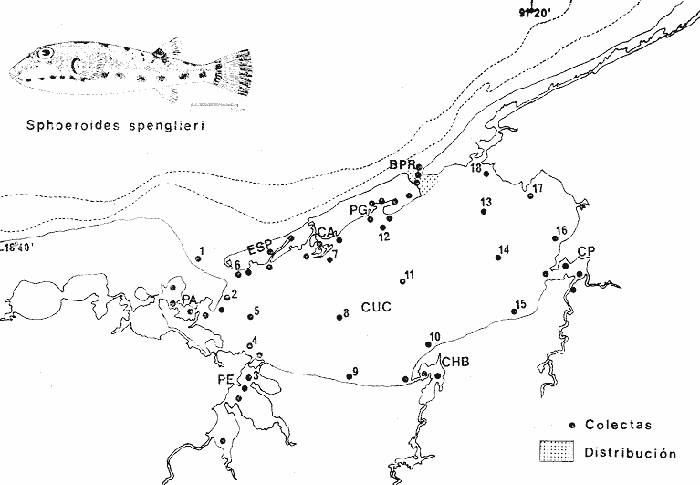 Fig. 17. Mapa de distribución de Sphoeroides spengleri en la Laguna de Términos, en relación a las estaciones de colecta en las distintas localidades. Relación longitud/peso y frecuencia de tallas. Sólo se colectó un ejemplar de 110 mm en el área de Puerto Real. Madurez. Se analizó un ejemplar capturado en la Boca de Puerto Real que resultó hembra en estadio II. Alimentación. Se analizó cualitativamente el contenido del estómago, encontrándose Restos de Equinodermos en gran proporción; y en menor proporción materia orgánica, Gasterópodos, Anfípodos y Restos de Vegetales. Sphoeroides greeleyi (Gilbert, 1900) N.v.: "Pez globo", "Tambor", "Xpu"
Tetraodon greeleyiGilbert, 1900; In: Randall, Caribbean-Reef Fishes 1968: 280. Diagnosis. Cuerpo alargado. Profundidad 3.1 a 4.6 en la LP. Cabeza 2.0 a 3.1 en la LP; hocico 1.8 a 2.2 en la cabeza y los ojos 3.0 a 4.5, interorbital varía con la edad 4.3 a 9.0 en la longitud cefálica. Piel del dorso cubierta con espinitas sensibles al tacto, dispuestas desde la nuca hasta antes de la dorsal. Vientre cubierto con espinitas desde el mentón hasta antes de la anal. Flancos y partes posteriores del dorso y vientre cubiertos con escamas dérmicas de márgenes libres y arregladas en series de líneas más o menos definidas, pero no osificadas. D 7-8; A 6-7. Pectorales 1.6 a 3.0 con 12 radios. Color. Dorso verde grisáceo sin reticulaciones con manchas oscuras redondeadas y pequeñas, rodeando la dorsal y pectorales formando una elipse en el dorso. Flancos generalmente sin un patrón de manchas redondeadas oscuras. Vientre pálido. Aletas pálidas. Distribución. En el Atlántico desde el Caribe a Brasil, incluyendo Golfo de México. En el área de estudio: localidades BPR, PG, CA, ESP, PA, PE (18). Material examinado. 11 ejemplares de 21 a 85 mm de longitud total. Colección de referencia depositada en el Laboratorio de Ictiología y Ecología Estuarina del Inst. de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. Catálogo ICMLT/F 0.001956/1962 (Camp.). 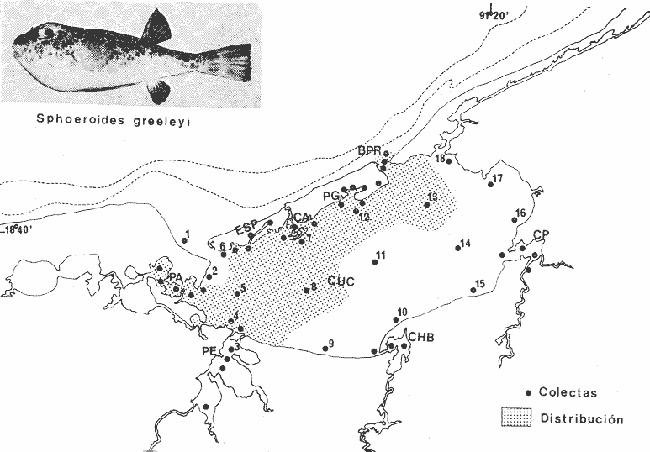 Fig. 18. Mapa de distribución de Sphoeroides greeleyi en la Laguna de Términos, en relación a las estaciones de colecta en las distintas localidades. Observaciones taxonómicas. Algunas consideraciones taxonómicas sobreSphoeroides greeleyi(Gilbert) en la costa Atlántica y Golfo de México han sido hechas por: Randall (1968) y Tyler (1980). La presencia de espinitas que cubren la mitad del dorso y vientre, combinadas con escamas dérmicas en la porción posterior es característica de esta especie. Difiere de S. testudineus (Linnaeus) yS. nephelus (Goode y Bean) en que carece de reticulaciones en el dorso y en que no tiene escamas membranosas en todo el cuerpo. DeS. spengleri (Bloch) que no presenta los apéndices dérmicos en el dorso. Este es el primer registro deSphoeroides greeleyi (Gilbert) dentro de la Laguna de Términos, Campeche, basado en 11 ejemplares colectados en la parte del Interior de la Isla y en los Sistemas Fluvio-Lagunares. Observaciones biológicas y ecológicas. Abundancia. Para el periodo de secas, se cuenta con dos registros en el Interior de la Isla del Carmen en las localidades ESP-1 y CA-2, ambos con un ejemplar y cuyos valores son 0.01 g m-2 y 0.00 1 g m-² respectivamente. Relación longitud/peso y frecuencia de tallas. De acuerdo a la ecuación, los valores calculados para las constantes a y b y el coeficiente de regresión R, se muestran graficados en las figuras 19 y 20. El rango de tallas para el total de ejemplares es de 21 a 85 mm con una moda a los 50 mm, encontrándose mejor representados en el Interior de la Isla del Carmen en el periodo de secas. En los Sistemas Fluvio-Lagunares sólo se registraron 3 ejemplares en la época de lluvias. Madurez. En la distribución de fases sexuales para Sphoeroides greeleyi, se registran solo fases I y II. Comparando las épocas climáticas se observa que en el periodo de secas existen más organismos en el Interior de la Isla del Carmen. 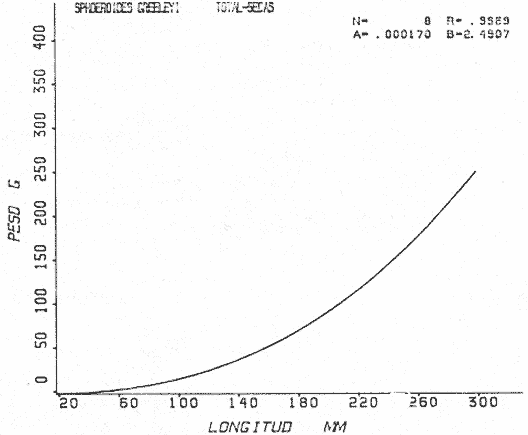 Fig. 19. Curva de regresión de la relación longitud/peso de Sphoeroides greeleyi. Total del Periodo de Secas. La relación macho: hembra para el total de los ejemplares es de 1:1.2 permaneciendo constante para la época de secas y aumentando a 1:2 en la época de lluvias en los Sistemas Fluvio-Lagunares. No se realizó el análisis de madurez gonádica en relación a la talla, debido que solo se encontraron fases I y II. Alimentación. El estudio de las relaciones tróficas se basó en el análisis del contenido de 11 estómagos en función de las épocas climáticas. 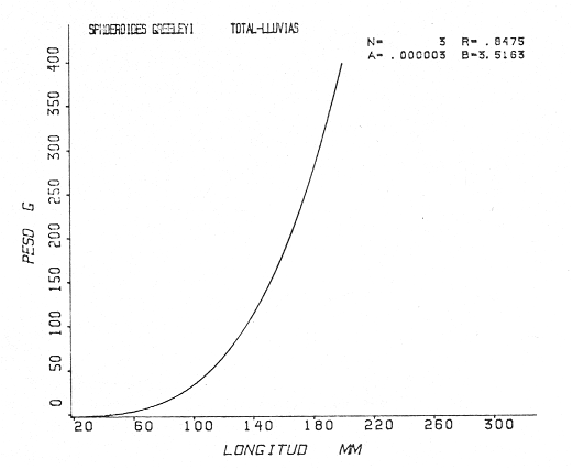 Fig. 20. Curva de regresión de la relación longitud/peso de Sphoeroides greeleyi. Total del Periodo de Lluvias. 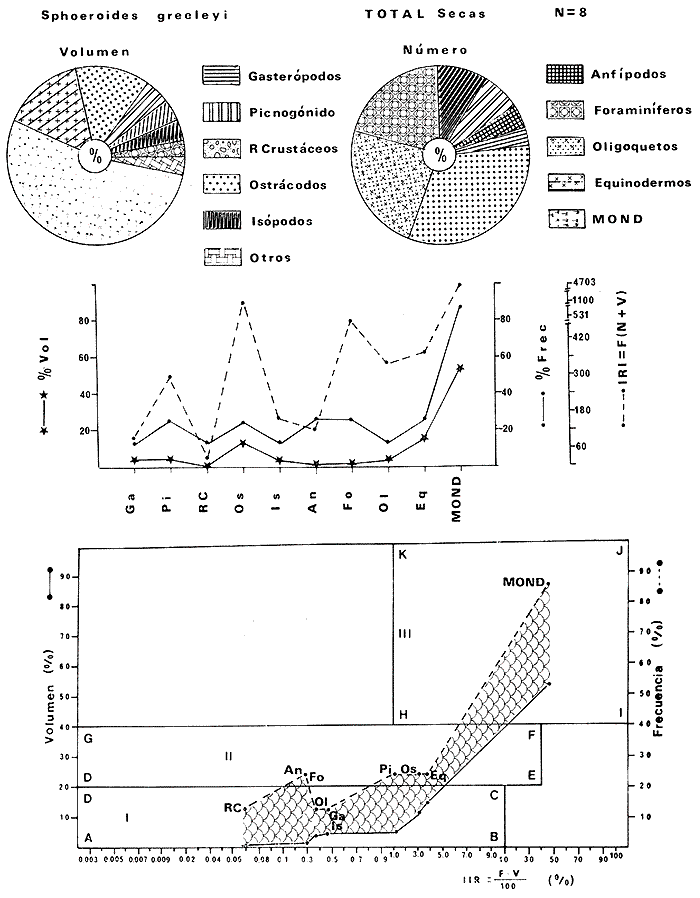 Fig. 21. Espectro trófico de Sphoeroides greeleyi en el Total del Periodo de Secas. 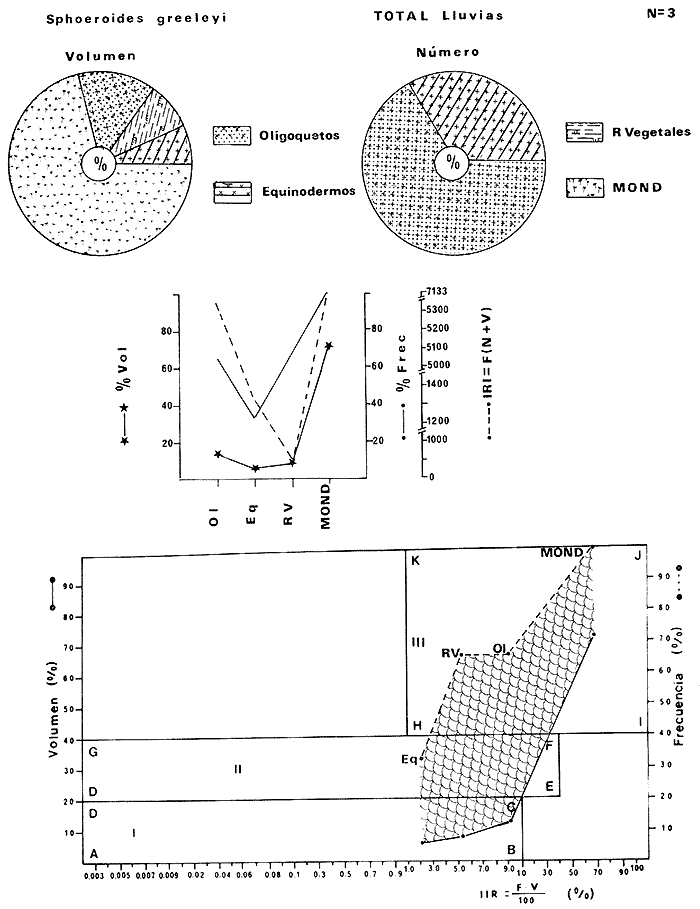 Fig. 22. Espectro trófico de Sphoeroides greeleyi en el Total del Periodo de Lluvias. Total del Periodo de secas. El espectro trópico (Fig. 21) señala que numéricamente el principal alimento lo constituyen Ostrácodos y en menor proporción Oligoquetos y Foraminíferos. Por la frecuencia, materia orgánica, Picnogónidos, Ostrácodos, Anfípodos, Foraminíferos y Equinodermos constituyen el alimento importante. Volumétricamente, materia orgánica, Equinodermos y Ostrácodos son los grupos trópicos destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestra que la materia orgánica, Ostrácodos, Foraminíferos, Equinodermos, Oligoquetos y Picnogónidos presentan los índices más elevados. En relación a la figura 21, se puede establecer que el alimento principal es la materia orgánica; el alimento secundario lo constituyen Equinodermos y Ostrácodos; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. Total del periodo de lluvias. El espectro trópico (Fig. 22) señala que numéricamente el principal alimento lo constituyen Oligoquetos y en menor proporción Equinodermos. Por la frecuencia, materia orgánica, Oligoquetos y Restos vegetales constituyen el alimento importante. Volumétricamente, materia orgánica y Oligoquetos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestran que materia orgánica, Oligoquetos y Equinodermos presentan los índices más elevados. En relación a la figura 22 se puede establecer que el alimento principal es la materia orgánica; el alimento secundario lo constituyen los otros grupos trópicos encontrados. 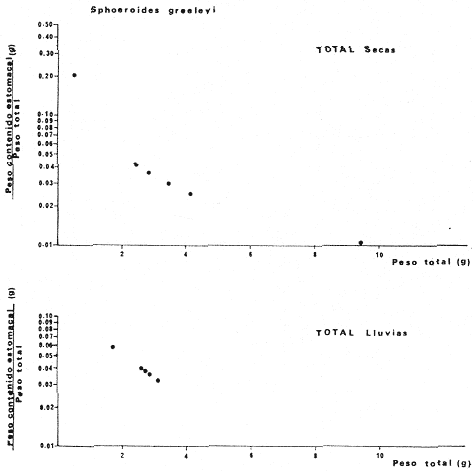 Fig. 23. Dispersión gravimétrica del alimento en relación al tamaño en peso de Sphoeroides greeleyi. Periodo de Secas N = 8. Periodo de Lluvias N = 11 La relación gravimétrica del alimento ingerido porSphoeroides greeleyi se muestra en la figura 23. El análisis estadístico indica un comportamiento semejante comparando los dos periodos climáticos, donde se observa que el índice es notablemente grande en los estadios juveniles y éste decrece linealmente conforme aumenta de tamaño la especie. Sphoeroides testudineus (Linnaeus, 1758) N.v.: "Pez globo", "Tambor", "Xpu"
Tetraodon testudineusLinnaeus, Syst. Nat., Ed. 10, 1758: 332 (Charleston, Carolina del Sur). Diagnosis. Cuerpo robusto, moderadamente alargado. Profundidad 2.9 a 6.3 en la LP. Cabeza amplia 2.5 a 3.7 en la LP; hocico largo 1.4 a 2.5 en la cabeza y los ojos pequeños 2.6 a 7.5; interorbital cóncavo ampliándose con la edad 3.1 a 7.5 en la longitud cefálica. Piel del dorso, flancos y vientre cubierta con espinitas generalmente hundidas no sensibles al tacto, hocico y cola suaves sin cirros. D 7-8; A 6-7. Pectorales cortas 1.2 a 3.1 en la longitud cefálica, 15 radios. Color. Dorso olivo oscuro, con líneas delgadas blanquecinas que se engrosan proporcionalmente con la edad, formando un patrón de reticulaciones arregladas en posición transversa de forma elíptica y circular; con una o dos barras entre los ojos. Flancos más claros con manchas redondeadas de diferente tamaño, oscuras casi negras dispuestas irregularmente, incrementándose en número con la edad. Vientre pálido. Caudal oscura hacia la parte distal. Pectorales con una banda oscura en la base. Otras aletas pálidas. Distribución. En el Atlántico. desde Massachussetts, EUA, hasta Brasil, incluyendo Golfo de México y las Antillas. En el área de estudio: localidades BPR, PG, CA, ESP, PA, PE, CHB, CP (Fig. 24). Material examinado. 557 ejemplares de 46 a 250 mm de longitud total. Colección de referencia depositada en el Laboratorio de Ictiología y Ecología Estuarina del Inst. de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. Catálogo ICMLP/F 0.001919/1925 (Camp.). Observaciones taxonómicas. Algunas consideraciones taxonómicas sobreSphoeroides testudineus (Linnaeus) en la costa Atlántica y Golfo de México han sido hechas por: Jordan y Evermann (1896), Meek e Hildebrand (1925), Cervigón (1966), Randall (1968), Bohlke y Chaplin (1968), Gallaway et al. (1972), Chaplin (1972), Perry y Perry (1974), Goodson (1976), SIC (1976), Dickson y Moore (1977), Greenberg (1977), Guitart (1978), Shipp y Yerger (1969), Castro Aguirre (1978) y Tyler (1980). Los caracteres taxonómicos tomados en cuenta en este estudio, para diferenciar Sphoeroides testudineus(Linnaeus) de S. nephelus (Goode y Bean) son determinantes, no presentándose confusión alguna en esclarecer diferencias básicas para la identificación correcta de la misma. La diferenciación se basa principalmente en la coloración del dorso, en la forma y tamaño del interorbital el número de radios en las pectorales y la distribución de las espinitas en el dorso. PresentandoS. testudineus (Linnaeus) las reticulaciones no tan pálidas y arregladas en un patrón de forma -regular, el interorbital cóncavo y más delgado, aunque es un carácter variado en individuos adultos del mismo tamaño y el dorso completamente cubierto con espinitas. De las otras especies estudiadas pertenecientes al género, se diferencía por presentar el patrón de reticulaciones pálidas en el dorso. A diferencia de S. eulepidotus (Metzelaar) y S. spengleri (Bloch) carece de aletillas dérmicas a los lados del cuerpo. Difiere enS. parvus (Shipp y Yerger) por presentar el patrón de reticulaciones en el dorso. Se separa de S. annulatus(Jenyns) y S. lobatus (Steindachner) por tener una distribución geográfica distinta y por la coloración del dorso. 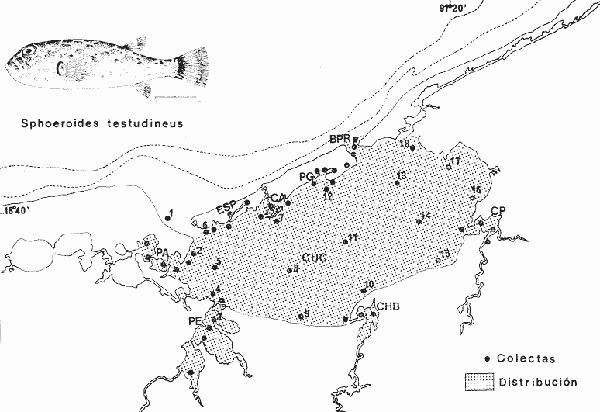 Fig. 24. Mapa de distribución de Sphoeroides testudineus en la Laguna de Términos, en relación a las estaciones de colecta en las distintas localidades. Es la única especie del género reportada en otra localidad mexicana, costa de Tampico, Tamp., y en el sistema estuarino Champotón, Camp. Así mismo ha sido reportada previamente para la Laguna de Términos por: Castro-Aguirre (1978), Bravo-Núñez y Yáñez-Arancibia (1979), Amezcua Linares y Yáñez-Arancibia (1980), Yáñez-Arancibia et al. (1980 y 1982), Yáñez-Arancibia (1981) y Vargas Maldonado et al. (1981). Observaciones biológicas y ecológicas. Abundancia. La biomasa de Sphoeroides testudineus en la Laguna de Términos fue muy variable durante 1976-1977, dependiendo de la localidad y la época climática (Tablas 4 y 5). En la Boca de Puerto Real para la época de secas, el máximo valor corresponde a la localidad BPR-3 de 0.20 gm-² con 4 ejemplares; mientras que el mínimo se registra en BPR-1 con un valor de 0.03 gm-² con un ejemplar. En esta última localidad en la época de lluvias la estación BPR-3 registra el valor más alto de biomasa 1.45 gm-² con 34 individuos; y la localidad BPR-1 registra el valor mínimo que corresponde a 0.03 g m-² con 1 individuo. Vargas Maldonado et al. (1981) reportan para el Interior de la Isla del Carmen en el periodo de secas en la localidad ESP-1 el máximo valor de 2.14 gm-² con 17 ejemplares; mientras que el mínimo valor de biomasa se registró en la estación ESP-2 siendo de 0.17 gm-² con 2 ejemplares. Durante la época de lluvias la estación ESP-1 registra el valor más grande de biomasa 0. 14 gm-² con 3 ejemplares y la estación CA-1 registra el menor valor 0.01 con 1 ejemplar. Para los Sistemas Fluvio-Lagunares, Amezcua Linares y Yáñez-Arancibia (1980) reportan en la época de secas, para los sistemas de Pom-Atasta y Palizada-del Este, valores muy cercanos de biomasa que van de 0.05 a 0.08 gm-² sólo la estación PA-5 registró un valor de 0. 17 gm-². En esta misma localidad, durante la época de lluvias el máximo valor es de 0.53 gm-² y lo registran las estaciones CP-1 con 15 ejemplares y CHB-2 con 5 ejemplares; mientras que el mínimo valor de biomasa, 0.008 gm-² corresponde a la estación CP-2 con un ejemplar. Los demás valores registrados para esta localidad van de 0.01 a 0.09 gm-² 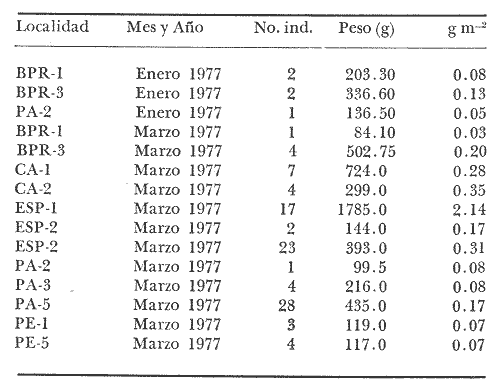 TABLA 4 SPHOEROIDES TESTIDINEUS ABUNDANCIA ESTIMADA DURANTE LA ÉPOCA DE SECAS (1977) EN LAS DOS ÁREAS ESTUDIADAS 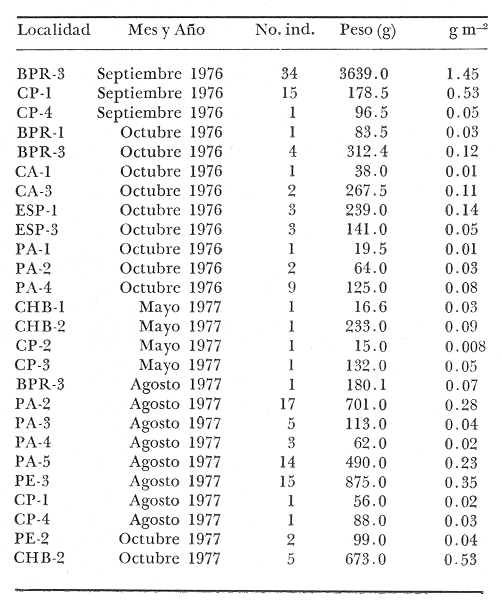 TABLA 5 SPHOEROIDES TESTIDINEUS ABUNDANCIA ESTIMADA DURANTE LA ÉPOCA DE LLUVIAS (1977) EN LAS DOS ÁREAS ESTUDIADAS 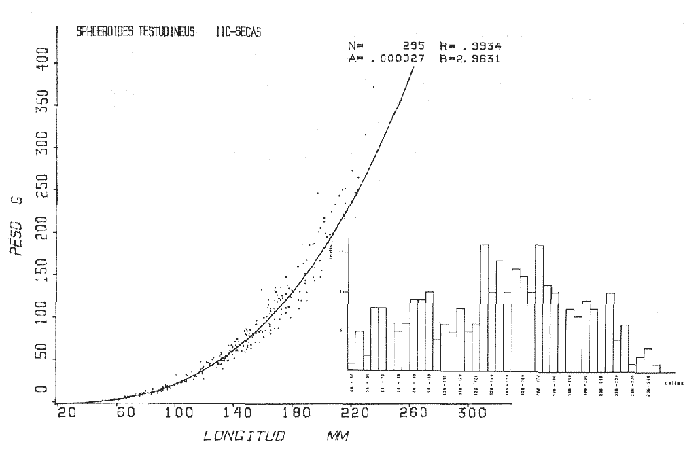 Fig. 25. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Interior de la Isla del Carmen. Época de Secas. 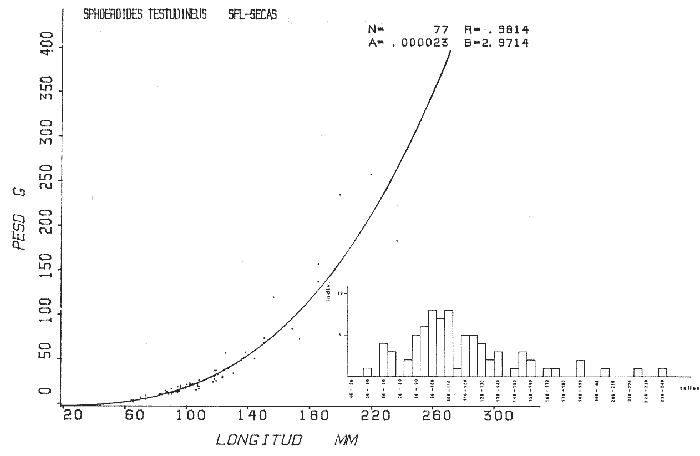 Fig. 26. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Sistemas Fluvio-Lagunares. Época de Secas. 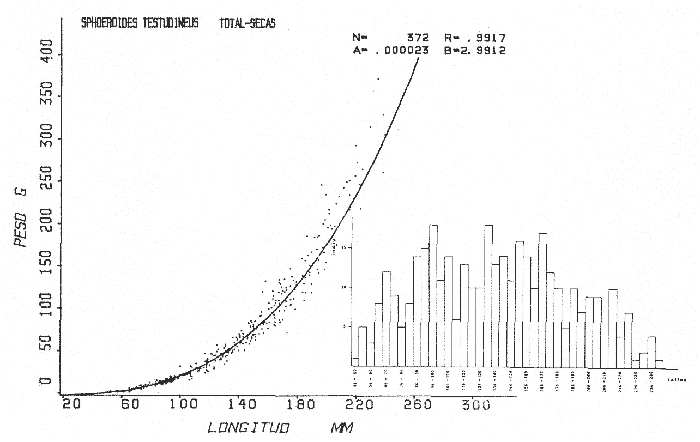 Fig. 27. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Total del Periodo de Secas. 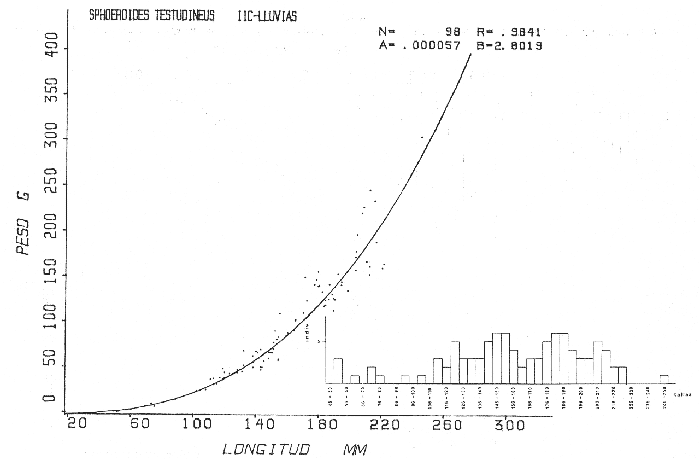 Fig. 28. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Interior de la Isla del Carmen. Época de Lluvias. 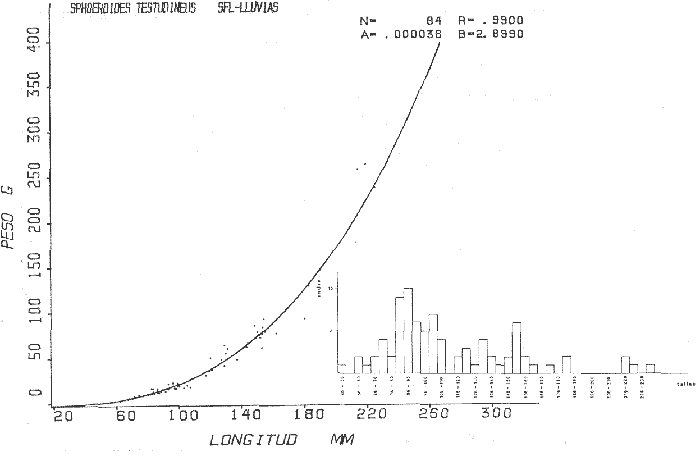 Fig. 29. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Sistema Fluvio-Lagunares. Época de Lluvias. 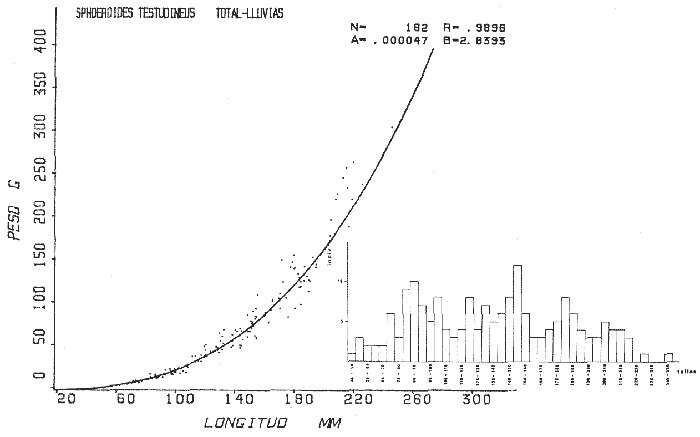 Fig. 30. Curva de regresión de la relación longitud/peso de Sphoeroides testudineus. Se señala la distribución de frecuencia de tallas. Total del Periodo de Lluvias. Relación longitud/peso y frecuencia de tallas. En la relación longitud/peso, de acuerdo a la ecuación los valores calculados para las constantes a y b y el coeficiente de regresión R se comparan para el total de los ejemplares en las dos épocas climáticas en las figuras 25 a 30. En la distribución de rangos de longitud para la especie se comparan las dos épocas climáticas y se observa una mayor frecuencia en el periodo de secas, para este mismo periodo comparando las dos localidades, se nota que el área del Interior de la Isla del Carmen se encuentran mejor representados los organismos con tallas más grandes; mientras que para los Sistemas Fluvio-Lagunares los ejemplares presentan tallas más pequeñas, como se observa en las modas. Para el periodo de lluvias el número de individuos en cada área es relativamente semejante, sin embargo existen diferencias al observar que las tallas más grandes son más frecuentes en organismos del Interior de la Isla del Carmen, donde se observa que la distribución es más homogénea, y las tallas más pequeñas las presentan ejemplares de los Sistemas Fluvio-Lagunares. El análisis de tallas y de relación longitud/peso se indica en las figuras 25 a 30. Madurez. La distribución de las fases sexuales en las dos épocas climáticas para las diferentes localidades, se muestran graficadas en la figura 31. Las figuras 32 y 33 muestran la relación de la madurez gonádica y la talla de los individuos, lo cual es fundamental para determinar el tamaño al cual la población alcanza la madurez y es apta para reproducirse, se nota que en el periodo de secas la población alcanza la madurez sexual (fase III) después de los 135 mm de longitud total. Para el periodo de lluvias se observan ejemplares maduros desde los 125 mm. El análisis de la proporción de sexos se sintetiza en la tabla 6. Alimentación. El estudio de las relaciones tróficas de Sphoeroides testudineus se basó en el análisis del contenido de 38 estómagos, en función de las localidades y las épocas climáticas. 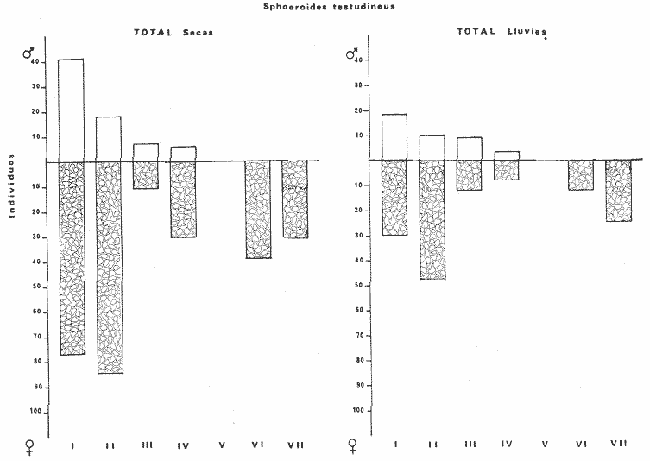 Fig. 31. Fases de desarrollo sexual y proporción de sexos de Sphoeroides testudineus para ambos periodos climáticos, independiente de la localidad. 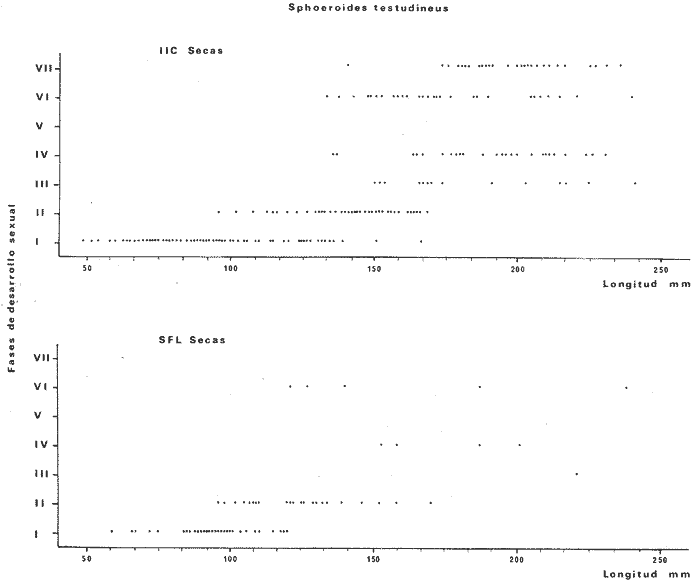 Fig. 32. Relación de la longitud total y las fases de desarrollo sexual de Sphoeroides testudineus en el Periodo de Secas de ambas localidades. Interior de la Isla del Carmen (secas). El espectro trópico (Fig. 34) señala que numéricamente el principal alimento lo constituyen Restos de Crustáceos y Pelecípodos y en menor proporción Foraminíferos, Huevos de Invertebrados y Gasterópodos. Por la frecuencia, materia orgánica, Pelecípodos y Gasterópodos constituyen el alimento importante. Volumétricamente, materia orgánica y Pelecípodos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestran que materia orgánica, Pelecípodos y Gasterópodos presentan los índices más elevados. En relación a la figura 34 se puede establecer que el alimento principal es la materia orgánica y Pelecípodos; el alimento secundario. lo constituyen Gasterópodos y Huevos de Invertebrados; el alimento circunstancial está representado por los otros grupos trópicos encontrados.  TABLA 6 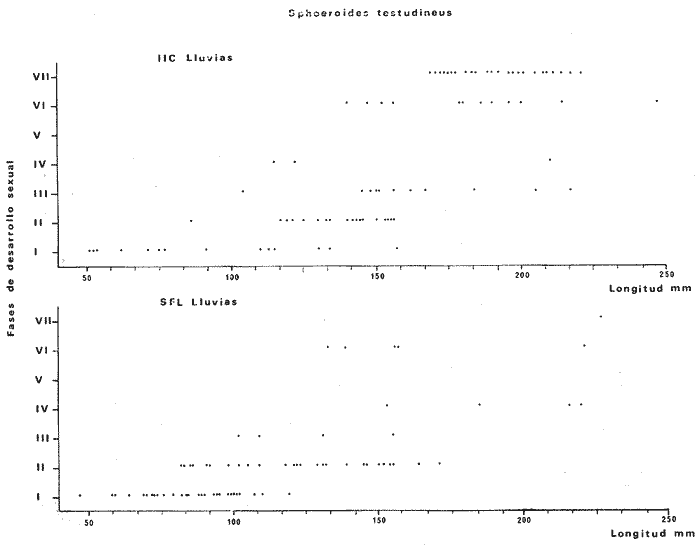 Fig. 33. Relación de la longitud total y las fases de desarrollo sexual de Sphoeroides testudineus en el Periodo de Lluvias de ambas localidades. Sistemas Fluvio-Lagunares (secas). El espectro trópico (Fig. 35) señala que numéricamente el principal alimento lo constituyen Tanaidáceos y en menor proporción Foraminíferos y Pelecípodos. Por la frecuencia, materia orgánica y Pelecípodos constituyen el alimento importante. Volumétricamente, materia orgánica, Pelecípodos y Tanaidáceos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestra que materia orgánica, Pelecípodos, Tanaidáceos y Foraminíferos presentan los índices más elevados. En relación a la figura 35 se puede establecer que el alimento principal es la materia orgánica y los Pelecípodos; el alimento secundario lo constituyen Tanaidáceos, Restos de Crustáceos y Restos de Vegetales; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. Total del periodo de secas. La combinación de los datos cuantitativos de las dos localidades, Interior de la Isla del Carmen y Sistemas Fluvio-Lagunares, permite describir la alimentación de la población para esta época. El espectro trópico (Fig. 36) señala que numéricamente el principal alimento lo constituyen Pelecípodos, Foraminíferos y Tanaidáceos. Por la frecuencia, materia orgánica, Pelecípodos y Foraminíferos constituyen el alimento importante. Volumétricamente, materia orgánica y Pelecípodos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestra que la materia orgánica, Pelecípodos, Foraminíferos, Restos de Crustáceos y Huevos de Invertebrados presentan los índices más elevados. En relación a la figura 36 se puede establecer que el alimento principal es materia orgánica y Pelecípodos; el alimento secundario lo constituyen Restos de Crustáceos, Gasterópodos y Huevos de Invertebrados; el alimento circunstancial está representado por los otros grupos trópicos encontrados. Interior de la Isla del Carmen (lluvias). El espectro trópico (Fig. 37) señala que numéricamente el principal alimento lo constituyen Pelecípodos y en menor proporción Gasterópodos. Por la frecuencia, materia orgánica, Pelecípodos y Gasterópodos constituyen el alimento importante. Volumétricamente, materia orgánica y Pelecípodos son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestra que la materia orgánica, Pelecípodos y Gasterópodos presentan los índices más elevados. En relación a la figura 37 se puede establecer que el alimento principal es la materia orgánica; el alimento secundario lo constituyen Pelecípodos; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. Sistemas Fluvio-Lagunares(lluvias). El espectro trópico (Fig. 38) señala que numéricamente el principal alimento lo constituyen Pelecípodos y Foraminíferos y en menor proporción Huevos de Invertebrados, Ostrácodos y Restos de Crustáceos. Por la frecuencia, materia orgánica, Restos de Crustáceos y Gasterópodos constituyen el alimento importante. Volumétricamente materia orgánica, Restos de Crustáceos y Pelecípodos, son los grupos trópicos más destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestran que la materia orgánica, Pelecípodos, Restos de Crustáceos, Foraminíferos y Huevos de Invertebrados presentan índices más elevados. En relación a la figura 38 se puede establecer que el alimento principal es la materia orgánica; el alimento secundario lo constituyen Restos de Crustáceos y Pelecípodos; y el alimento circunstancial está representado por los otros grupos tróficos encontrados. Total del periodo de lluvias. La combinación de los datos cuantitativos de las dos localidades, Interior de la Isla del Carmen y Sistemas Fluvio-Lagunares, permite describir la alimentación de la población para esta época. El espectro trópico (Fig. 39) señala que numéricamente el principal alimento lo constituyen Pelecípodos y Foraminíferos y en menor proporción Huevos de Invertebrados y Ostrácodos. Por la frecuencia, materia orgánica, Pelecípodos, Gasterópodos y Restos de Crustáceos constituyen el alimento importante. Volumétricamente, materia orgánica, Pelecípodos y Restos de Crustáceos son los grupos trópicos destacados. El análisis combinado de estos datos para cuantificar los índices de importancia relativa, muestran que materia orgánica, Pelecípodos, Restos de Crustáceos y Foraminíferos presentan los índices más elevados. En relación a la figura 39 se puede establecer que el alimento principal es materia orgánica; el alimento secundario lo constituyen Pelecípodos y Restos de Crustáceos; y el alimento circunstancial está representado por los otros grupos trópicos encontrados. El estudio de la relación gravimétrica del alimento ingerido por Sphoeroides testudineus se muestra en la figura 40. El análisis estadístico indica un comportamiento similar para los dos periodos climáticos, como se observa el índice es notablemente grande tanto en juveniles como en adultos, es decir que el rango de variación es muy amplio y muestra que no existe correlación entre el índice y el peso total de las necesidades ejemplar, comportándose con altas n alimenticias durante toda su vida. Esto representa a un consumidor de Segundo Orden. 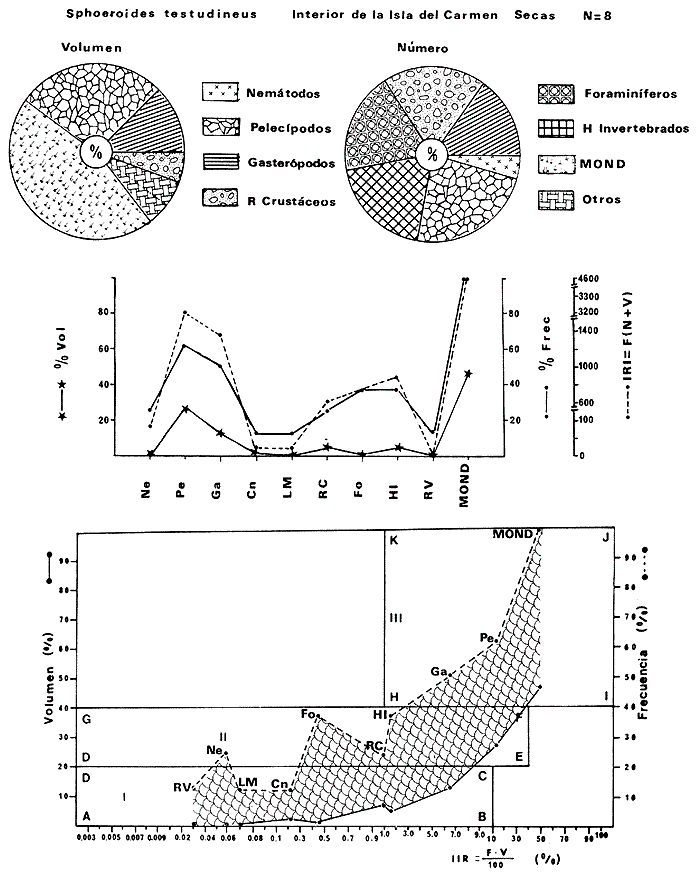 Fig. 34. Espectro trópico de Sphoeroides testudineus en el Periodo de Secas del Interior de la Isla del Carmen. 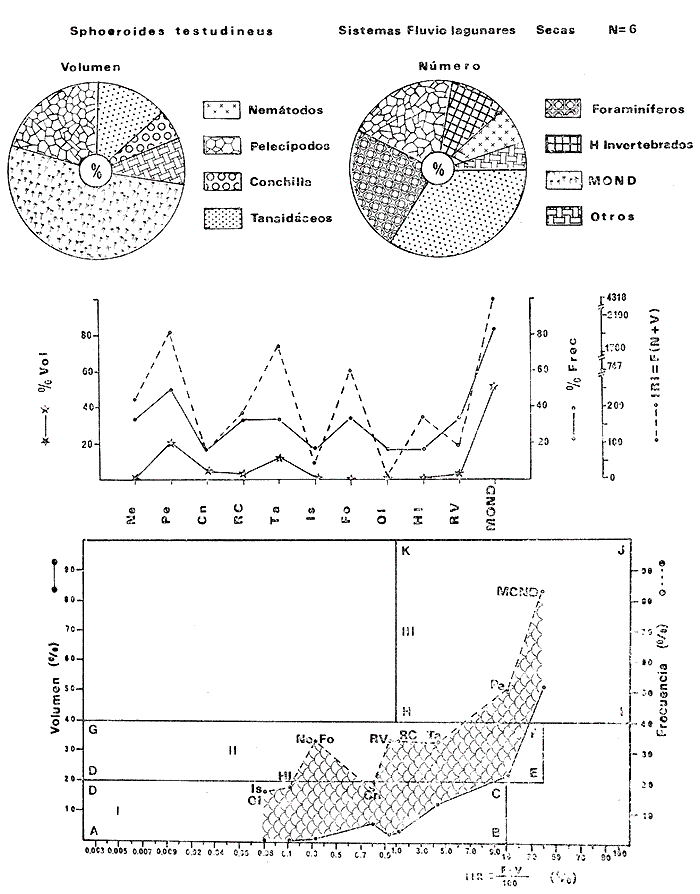 Fig. 35. Espectro trópico de Sphoeroides testudineus en el Periodo de Secas de los Sistemas Fluvio-Lagunares. 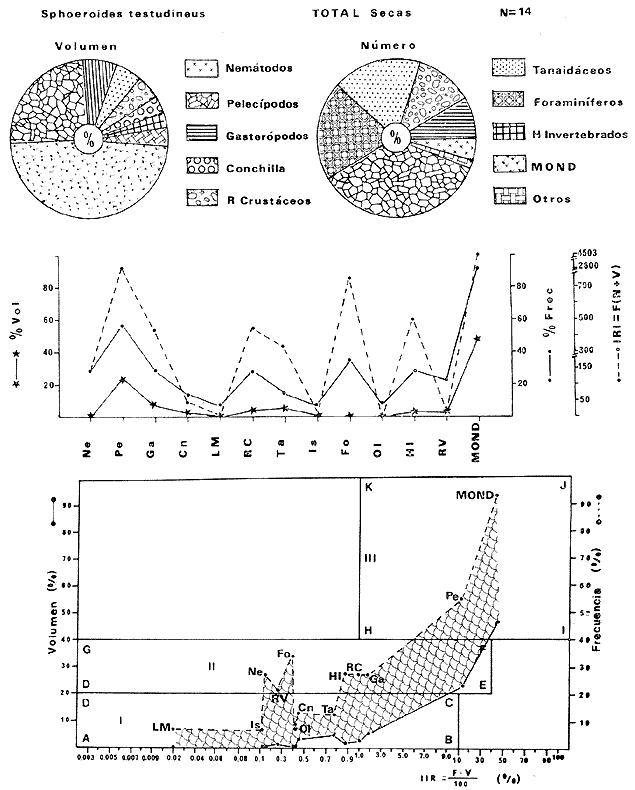 Fig. 36. Espectro trópico de Sphoeroides testudineus del Total del Periodo de Secas. 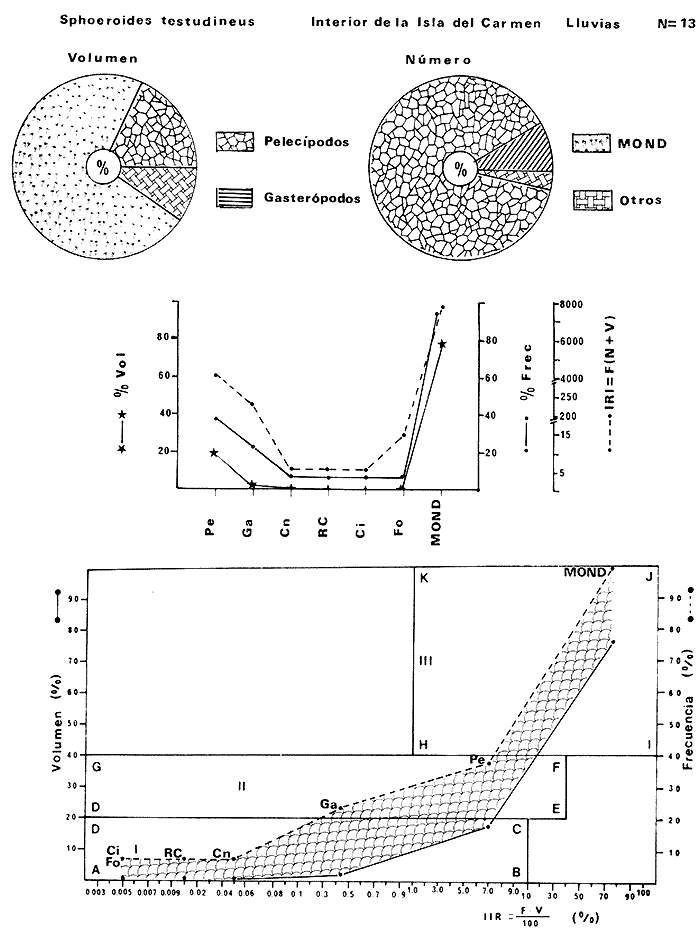 Fig. 37. Espectro trópico de Sphoeroides testudineus en el Periodo de Lluvias del Interior de la Isla del Carmen, 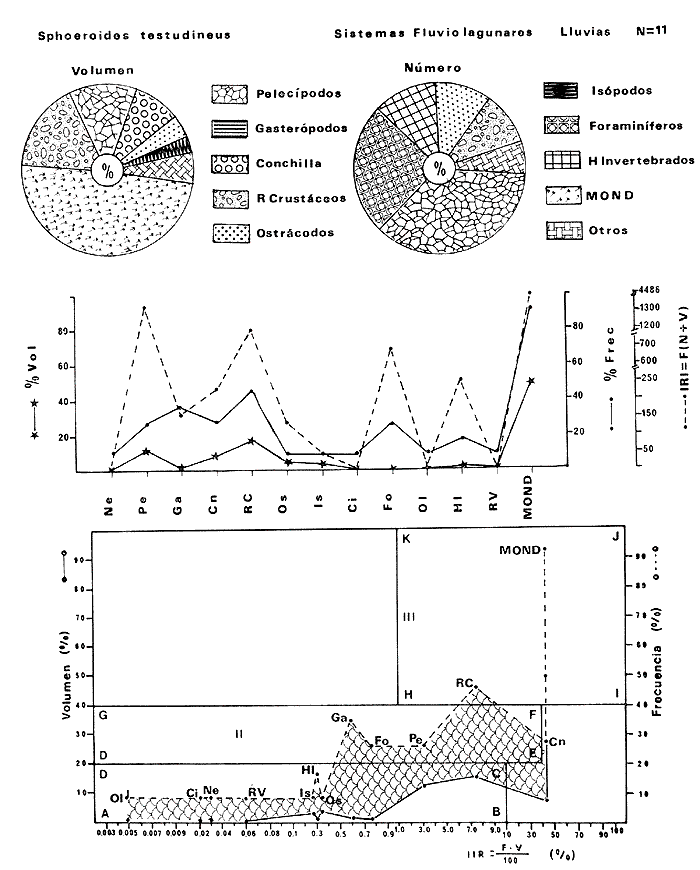 Fig. 38. Espectro trópico de Sphoeroides testudineus en el Periodo de Lluvias de los Sistemas Fluvio-Lagunares, 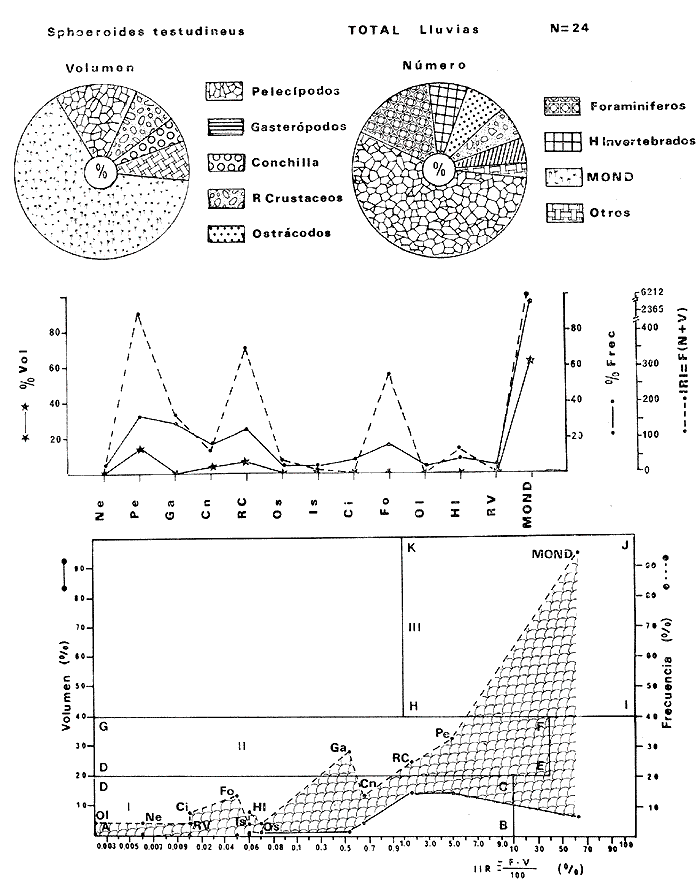 Fig. 39. Espectro trópico de Sphoeroides testudineus del Total del Periodo de Lluvias. DISCUSIÓN
DISTRIBUCIÓNDebido a la gran diversidad de subsistemas dentro de la Laguna de Términos y al reflejo de los gradientes causados por el patrón de circulación de las aguas, existen factores fundamentales como: salinidad, temperatura, nutrientes, sedimentación y turbidez, entre otros, que determinan la existencia de diferentes comunidades y la distribución de las especies. Con base en la información obtenida, se aprecia que la familia Tetraodontidae presenta diferentes patrones de distribución para cada una de las especies registradas, que pueden ser resumidos como sigue: 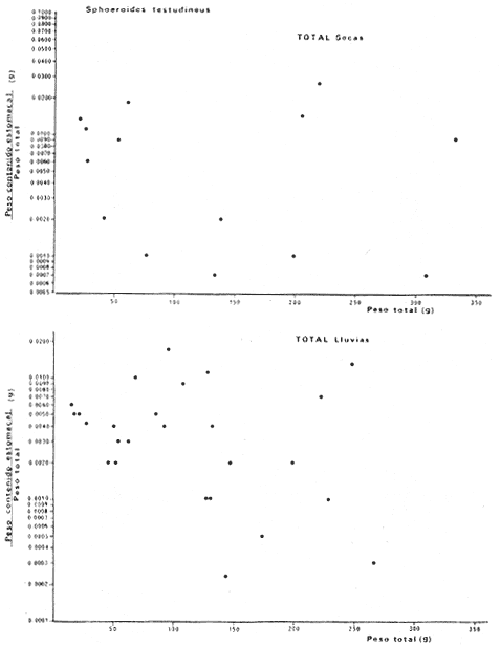 Fig. 40. Dispersión gravimétrica del alimento en relación al tamaño en peso de Sphoeroides testudineus. Periodo de Secas N = 14. Periodo de lluvias N = 38. Lagocephalus laevigatus y Sphoeroides spengleri son especies raras y poco frecuentes en la Laguna de Términos. Sólo habitan áreas con salinidades marinas de 27 a 39 ppm, temperaturas de 22 y 29°C, alta transparencia (80%) y sedimentos arenosos con vegetación sumergida de pastos marinos y macroalgas (Figs. 3 y 17). Sphoeroides nephelus es una especie cuya distribución queda restringida en áreas poco profundas y salinidades de 25 a 39 ppm, temperaturas de 19 y 32°C, alta transparencia (80%), en praderas de Thalassia testudinum. Todos los ejemplares corresponden al área del Interior de la Isla del Carmen y con mayor frecuencia en el periodo de secas, sólo dos ejemplares de Sphoeroides nephelus se colectaron en el Sistema Fluvio-Lagunar de Pom-Atasta (Fig. 4). Sphoeroides greeleyi es una especie poco frecuente en el área, pero con una distribución amplia y homogénea (Fig. 18) tanto en el Interior de la Isla del Carmen como en las localidades Pom-Atasta y Palizada-del Este de los Sistemas Fluvio-Lagunares. Puede decirse que se trata de una especie euritermohalina ya que se encuentra en áreas con salinidades de 21 a 39 ppm; temperaturas de 19 a 35°C; y transparencias de 10 a 80%. Se observa que habita áreas con pastos marinos, así como áreas con bancos de ostión. Sphoeroides testudineus es una de las especies mejor representadas y de amplia distribución, es por tanto característica del área y se le encuentra en todos los subsistemas (Fig. 24). Se trata de una especie euritermohalina con gran capacidad adaptativa que vive tanto en mar abierto -se ha colectado a 24 brazas en el Golfo de México (Bohlke y Chaplin, 1970)-, como en aguas someras con fondos fangosos y/o arenosos, cercanos a la costa o en arrecifes, común en habitat de pastos marinos, en áreas pobladas por mangle y marismas. Sin embargo, su mayor abundancia es en aguas claras sobre praderas de Thalassia testudinum. Perry y Perry (1974) y Castro-Aguirre (1978) reportan que aparentemente penetra a la desembocadura de los ríos, habiendo sido registrada con anterioridad en la Laguna de Términos por Yáñez-Arancibia et al. (1980 y 1982). ABUNDANCIAComo se menciona anteriormenteLagocephalus laevigatus y Sphoeroides spengleri son especies raras, la primera con un registro de biomasa muy bajo de 0.005 gm², en el área de Boca de Puerto Real. La segunda no cuenta con registros de abundancia. Sphoeroides nephelus es la segunda especie importante en abundancia de los Tetraodontidae, dentro de la Laguna de Términos. Los registros que existen son para la Boca de Puerto Real y para el Interior de la Isla del Carmen. En esta última en la época de secas existen valores más altos de biomasa, que en la época de lluvias, donde el máximo valor es de 0.07 gm-² y el mínimo es de 0.00 1 gm-². El máximo valor de biomasa paraSphoeroides greeleyi corresponde a 0.01 gm-² y el mínimo a 0.001 gm-², ambos para el Interior de la Isla del Carmen en el periodo de secas. Diversos autores han reportado aSphoeroides testudineus como un componente comunitario importante en todos los subsistemas de la Laguna de Términos, por su alta frecuencia y abundancia, que varía dependiendo de la localidad y la época climática. Bravo-Núñez y Yáñez-Arancibia (1979) reportan para la Boca de Puerto Real el máximo valor de biomasa con 1.45 gm-² y el mínimo con 0.03 gm-². Vargas Maldonado et al. (1981) reportan para el Interior de la Isla del Carmen, el máximo valor de biomasa con 2.14 gm-² y el mínimo con 0.01 gm-². Amezcua Linares y Yañez-Arancibia (1980) reportan para los Sistemas Fluvio-Lagunares el valor más alto de biomasa con 0.53 gm-² y el más bajo con 0.008 gm-². El máximo valor de biomasa para esta especie, corresponde a la época de secas en el área del Interior de la Isla del Carmen con 2.14 gm-² y el mínimo se registra en la época de lluvias en los Sistemas Fluvio-Lagunares con 0.008 gm². Del análisis anterior, se observa en general que para la familia Tetraodontidae, los valores más altos de biomasa corresponden al área del Interior de la Isla del Carmen en el periodo de secas. RELACIÓN LONGITUD/PESOLas diferencias obtenidas para la constante b en cada una de las especies, reflejan pequeñas variaciones. Se observa que los valores registrados para las especiesSphoeroides testudineus y Sphoeroides nephelus varían independientemente de la época climática de 2.7653 a 2.9912 con rangos de talla de 46 a 250 mm.Sphoeroides greeleyi tiene el valor más alto de la constante b de 3.5163 y el más bajo de 2.4907, en la época de lluvias y secas respectivamente; con rangos de talla de 21 a 85 mm. De lo que se puede inferir que los cambios de la constante b son debidos principalmente a las diferencias de rangos de talla que presenta cada especie, a la relación alométrica que van mostrando los individuos a medida que crecen y a la época climática. MADUREZDe las, especiesLagocephalus laevigatus y Sphoeroides spengleri sólo se analizó un ejemplar, de cada una, resultando hembras en fase VII y II respectivamente. De la distribución de fases sexuales para Sphoeroides nephelus se reporta que el total de ejemplares sólo alcanzó la fase de maduración sexual. IV. En la figura 8, se observa que la mayor frecuencia corresponde a las hembras, los machos sólo se encuentran en fase II siendo menos frecuentes. Sólo se registraron hembras maduras con tallas que van de 95 a 147 mm. Se encontraron dos ejemplares indeterminados en las dos épocas climáticas con rangos de longitud de 50 a 73 mm. De la distribución de fases sexuales para Sphoeroides nephelus se reporta que el total de ejemplares sólo alcanzó la fase de maduración sexual IV. En la figura 10 se nota que la mayor frecuencia corresponde a las hembras, los machos sólo se encuentran en fase I y III, siendo menos frecuentes. La figura 11, muestra la relación de la longitud total y las fases de desarrollo, donde se observa que a los 100 mm se encuentra con frecuencia ejemplares en fase III. Se registraron 4 ejemplares indiferenciados en maduración, en la época de lluvias con rangos de longitud de 46 a 62 mm. De la distribución de fases sexuales para Sphoeroides greeleyi se reporta que el total de los ejemplares son juveniles ya que solo se encontraron fases de maduración I y II; comparando las dos épocas climáticas se observa que el periodo de secas se encuentra mejor representado. Del análisis se observa que Sphoeroides testudineus es la única especie que presenta todas las fases de desarrollo gonádico. En la figura 31 se observa que la mayor frecuencia corresponde a las hembras, los machos se encuentran en las fases I a IV. Asimismo, se observa que la época de secas se encuentra mejor representada. La talla a la cual se estima que empiezan a madurar los, ejemplares es después de los 125 mm en la época de lluvias y después de los 135 mm en la época de secas. En las figuras 32 y 33 no aparece claramente la talla a la cual maduran los organismos, pero tentativamente puede proponerse la de 125 mm como talla promedio de maduración gonádica de los individuos. Se registraron 3 ejemplares indiferenciados en maduración en la época de secas con rangos de longitud de 51, 54 y 62 mm. Los estadios de fase VII de Lagocephalus laevigatus, aparentemente no tiene asociación alguna con sus hábitos reproductivos. Mientras que Sphoeroides testudineus puede ser explicado tomando en consideración que es una especie que pasa la mayor parte de su ciclo de vida dentro de la Laguna de Términos, como lo demuestra su alta frecuencia, amplia distribución, además de presentar una población heterogénea en tamaño. Álvarez Cadena y Flores-Coto (1981) reportan huevos y larvas de esta especie en gran proporción en el área de la Cuenca Central de la laguna; sin embargo, no se considera razón suficiente como para suponer que pueda llevar a cabo la reproducción dentro de ésta ya que podría ser el resultado de inmigración de áreas costeras inmediatas a ésta. De cualquier forma son necesarios más estudios para poder tener un conocimiento más preciso. ALIMENTACIÓNEn la especieLagocephalus laevigatus no se analizó cuantitativamente el espectro trópico. El estudio cualitativo efectuado indica que puede alimentarse de materia orgánica de origen diverso (detritus) en gran proporción, Restos de Vegetales y Peces. El espectro trópico de Sphoeroides nepheluses amplio. Del análisis efectuado, se puede establecer que la especie es un consumidor de Segundo Orden. El alimento principal queda constituido por materia orgánica de origen diverso (detritus); el alimento secundario lo constituyen Pelecípodos, Restos de Vegetales y Restos de Crustáceos; y diversos grupos menores del meiobentos constituyen el alimento circunstancial (Fig. 12). Del análisis hecho en función de distintos periodos climáticos, se puede establecer que la especie es un consumidor de Segundo Orden. El alimento principal lo constituyen materia orgánica de origen diverso (detritus) y Restos de Crustáceos; el alimento secundario lo constituyen Pelecípodos, Gasterópodos y Restos de Vegetales; y diversos grupos menores del meiobentos constituyen el alimento circunstancial. Este patrón de alimentación sufre ligeras modificaciones en función del periodo climático (Figs. 13 y 14). En la especie Sphoeroides spengleri no se analizó cuantitativamente el espectro trópico. El estudio cualitativo efectuado indica que puede alimentarse de Equinodermos en gran proporción, materia orgánica de origen diverso (detritus), Gasterópodos, Anfípodos y Restos de Vegetales. Randall (1967) analiza la alimentación deSphoeroides spengleri, encontrándose que es un depredador de macro-invertebrados. Por esos antecedentes y los de este estudio, se puede establecer que la especie puede comportarse también como un consumidor de Tercer Orden, dependiendo de la localidad y la disponibilidad del alimento. El espectro trópico deSphoeroides greeleyi es menos amplio. Del análisis efectuado, se puede establecer que la especie es un consumidor de Segundo Orden. El alimento principal queda constituido por materia orgánica de origen diverso (detritus), el alimento secundario lo constituyen Equinodermos, Ostrácodos, Restos de Vegetales, Oligoquetos; y otros grupos menores del meiobentos constituyen el alimento circunstancial (Figs. 21 y 22). El espectro trópico de Sphoeroides testudineus es muy amplio. Del análisis hecho en función de distintas localidades y periodos climáticos, se puede establecer que la especie es un consumidor de Segundo Orden. El alimento principal lo constituyen materia orgánica de origen diverso (detritus) y Pelecípodos; el alimento' secundario lo constituyen Gasterópodos, Restos de Crustáceos, Huevos de Invertebrados, Tanaidáceos y Restos de Vegetales; y diversos grupos menores del meiobentos constituyen el alimento circunstancial. Este patrón de alimentación sufre modificaciones pequeñas en función del periodo climático y la localidad dentro de la laguna (Figs. 34 a 39). Del examen del espectro trópico, se encuentra que éste no presenta diferencias fundamentales en relación a la época del año. La materia orgánica de origen animal, constituye el alimento principal, encontrándose en gran proporción para todas las especies. Sin embargo, la materia orgánica encontrada en los estómagos puede ser producto de la digestión de otros grupos trópicos, o ingestión directa de ella desde el medio. Esto último parece menos probable. Conclusiones1. La familia Tetraodontidae está muy bien representada en la Laguna de Términos, por su diversidad, distribución y abundancia. Cinco especies juegan un importante papel en la estructura y función de las comunidades ictiofaunísticas del área. 2. Se precisa el estatus taxonómico de las especies, discutiendo las diferencias y afinidades entre ellas, esto es:Lagocephalus laevigatus (Linnaeus), Sphoeroides spengleri(Bloch), Sphoeroides greeleyi(Gilbert),Sphoeroides testudineus (Linnaeus) ySphoeroides nephelus (Goode y Bean). 3.Lagocephalus laevigatus y Sphoeroides spengleri son especies cuya distribución queda restringida en áreas de mayor y persistente influencia marina. Sphoeroides nephelus cuya distribución está mejor representada en praderas deThalassia testudinum. Sphoeroides greeleyiySphoeroides testudineus son especies con una distribución amplia y homogénea, la segunda es más frecuente y se considera una especie típica en la estructura de las comunidades del área de estudio. 4. El orden de importancia de la abundancia de las especies y sus valores máximos es el siguiente: Sphoeroides testudineus: 2.14 gm-² Sphoeroides nephelus: 0.22 gm-² Sphoeroides greeleyi: 0.01 gm-² Lagocephalus laevigatus: 0.005 gm-² Estos datos, medidos en gramos por metro cuadrado, multiplicados por diez dan kg. Ha-¹. 5. La especie Sphoeroides testudineus presenta las tallas mayores de entre todas las especies, así como también estados larvarios y huevecillos en el ictioplancton. En todas las especies la constante b de alometría refleja variaciones pequeñas indicando un crecimiento alométrico de las especies. 6. La única especie que, aparentemente, utiliza en forma directa la Laguna de Términos para su ciclo reproductor es Sphoeroides testudineus; Sphoeroides nephelus utiliza la Laguna de Términos en alguna etapa de su ciclo de vida para protección, crianza y alimentación, al igual queSphoeroides greeleyi. Lagocephalus laevigatus y Sphoeroides spengleri son especies visitantes ocasionales en el área. 7.Sphoeroides nephelus madura sexualmente después de los 100 mm de longitud total. Sphoeroides testudineus tiene una talla promedio de maduración gonádica a los 215 mm de longitud. 8. El espectro trópico de las especies es amplio, considerándose a todas ellas como consumidores de Segundo Orden. Con variaciones individuales, de acuerdo a la época climática y a la localidad dentro de la Laguna de Términos, el alimentó principal es la materia orgánica de origen diverso (detritus) y Pelecípodos; el alimento secundario son Crustáceos, Restos de Vegetales y Gasterópodos; y otros grupos bénticos diversos complementa el régimen alimenticio de las especies. AgradecimientosLos autores agradecen a Agustín Ayala-Castañares, José Luis Rojas Galavíz, Antonio García-Cubas y Alfredo Laguarda Figueras, la cuidadosa revisión del manuscrito, haciendo valiosos comentarios y sugerencias. A los compañeros del Laboratorio de Ictiología y Ecología Estuarina, por sus valiosas críticas y comentarios a lo largo de la realización de este estudio, especialmente Ana Laura Lara-Domínguez, Patricia Sánchez-Gil, Irma Vargas Maldonado, Arturo Aguirre León y Silvia Díaz Ruiz. El apoyo institucional y el financiamiento para la realización del presente trabajo fue brindado por el Instituto de Ciencias del Mar y Limnología de la UNAM y parcialmente por el Programa Regional de Desarrollo Científico y Tecnológico de la Organización de los Estados Americanos (OEA) a través de su Oficina de Proyectos Especiales en Washington. LITERATURAÁLVAREZ CADENA, J. y C. FLORES-COTO An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México Clave para identificación de familias de larvas de peces de la Laguna de Términos, Campeche, México.1981. 199-208.8 (1:) AMEZCUA LINARES, F. y A. YÁÑEZ-ARANCIBIA An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México.,Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces.1980.70-118.7 (1) BOHLKE, J. E. y CH. C. G. CHAPLIN Fishes of the Bahamas and Adjacent Tropical Waters. Livingston Publ. Co.,Pennsylvania1968.771 p. BOSCHUNG, H. T., Ph. D. Dissertation The fishes of the Mobile Bay and the Gulf coast of Alabama. Univ. Alabama1957. 626 p. BRAVO-NÚÑEZ, E. y A. YÁÑEZ-ARANCIBIAAn. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoEcología de la Boca de Puerto Real, Laguna de Términos. I. Descripción del área y análisis estructural de las comunidades de peces.1979.125-182.6(1) CARR, W. E. S. y C. A. ADAMSTrans. Amer. Fish. Soc.,Food habits of juvenile marine fishes occupying seagrass beds in the estuarine zone near Crystal River, Florida.1973. 51 540.102 CARRANZA, I., Informe preliminar sobre alimentación y hábitos alimenticios de las principales especies de peces de las zonas de los planes pilotos Yavaros y Escuinapa.3er. Informe Secretaría de Recursos Hidráulicos e Instituto de Biología, Univ. Nal. Autón. México.1969. 50 p., 7 figuras. Contrato de Estudios No. EI-69-51 CASTRO-AGUIRRE, J. L.,Dir. Gral. Inst. Nal. Pesca, México, Serie Científica Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México con aspectos zoogeográficos y ecológicos.1978.1-298.19 CHAPLIN, C.,Fishwatcher's Guide to West Atlantic Coral Reefs.Livingston, Publ. Co.Pennsylvania1972. 64 p. CERAME-VIVAS, M. J., P. F. Z. Properties, Inc., Mangroves of Puerto Rico. 1974. 64 p. CERVIGON, M. F., Los Peces Marinos de Venezuela. Estación de Investigaciones Marinas de Margarita, Fundación La Salle de Ciencias Naturales, Caracas, Monogr.1966.1-951.11 y 12 DAHL, G., Los Peces del Norte de Colombia. INDERENA Bogotá1972. 391 p. DAY, J. W., W. G. SMITH y CH. S. HOPKINSON Proc. Second Coastal Marsh and Estuary Management Symposium. Some trophic relationship of marsh and estuarine areas. In: Chabreck, R. H. (Ed.) Louisiana State University Press1973.115-135. DICKSON, H. H. y R. H. MOOREFishes of the Gulf of Mexico, Texas. Louisiana and Adjacent Waters. Texas A & M University Press1977.327 p. FISCHER, W. FAO Species Identification Sheets for Fishery Purposes. Western Central Atlantic (fishing area 31). FISCHER, W. (Ed.) FAO.Roma.1978.5. GALLAWAY, B. J., J. C. PARKER y D. MOORE Key to the Estuarine and Marine Fishes of Texas.Texas A & M University Press1972. 177 p. GIERLOFF-EMDEN, H. G.,In: Orbital remote sensing of coastal and offshore environments: a manual of interpretation. Laguna de Términos and Campeche Bay, Gulf of México.Berlin1977.78-89. GOODSON, G., Fishes of the Atlantic Coast.Marquest Colorguide Books Palos Verdes Estates, California1976.203 p. GREENBERG, J., Guide to Corals and Fishes of Florida, the Bahamas and the Caribbean. Pub. Seahawk PressFlorida1977. 64 p. GUITART, J. D., Acad. Cienc. Cuba, Serie OceanológicaSinopsis de los peces marinos de Cuba.1978.762-767.4 HILDEBRAND, S. F. y W. C. SCHRODER Bull. U.S. Bureau Fish. Comm., Fishes of Chesapeake Bay.1928.1-366.43 (1) JORDAN, D. S. y B. W. EVERMANNBull. U.S. Nat. Mus.,The Fishes of North and Middle America.1896-1900.1-3313. 398 láms. 985 figs.14(47) JORDAN, E., M. ANGOT y R. DE LA TORRE An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoProspección biológica de la Laguna de Nichupté, Cancún, Quintana Roo. Nota Científica.1978.179-188.5 (1) LAEVASTU, T., Manual de Métodos de Biología Pesquera. Publicación. FAO. Ed.AcribiaEspaña1971.243 páginas. LARA-DOMÍNGUEZ, A. L., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoBiología y ecología del bagre Arius melanopus Günther en Laguna de Términos, Sur del Golfo de México. (Pisces: Ariidae).1981.267-3048 (1) LINTON, E., Bull. U.S. Bureau of Fisheries Comm., Parasites of fish of Beaufort. 1905.321-428.24 LOWE-MCCONNELL, R. H.,J. Linnean Soc. London The fishes of the British Guiana continental shelf, Atlantic coast of South America, with notes on their natural history.1962.669-700.44 (30) MEEK, E. S. y S. F. HILDEBRAND Field Mus. Nat. Hist. Publ., Zool. Ser., The marine fishes of Panama.1923-1928.1-1045.15(215, 226, 249) NIKOLSKY, G. V., The Ecology of Fishes.Academic Press Inc., New York 1963.352 p. PERRY, J. A. y S. D. PERRYSer. Cienc. Nat., Los peces comunes de la costa Atlántica de Costa Rica. Facultad de Ciencias y Letras, Departamento de Biología.1974.17.1 (7): PINKAS, L., M. S. OLIPHANT, y 1. K. L. IVERSON Dept. Fish. Game Cal. Fish. Bull.Food habits of albacore, bluefin tuna and bonito in California waters. 19711-105.15 RANDALL, J. E., Stud. Trop. Oceanogr., Univ. Miami Food habits of reef fishes of the West Indies.1967.665-847.5 ---, Caribbean Reef Fishes. T. F. H. Publications. Inc. New York1968.278 p. REID, G. K., Bull. Mar. Sci. Gulf Caribb., An ecological study of the Gulf of Mexico fishes in the vicinity of Cedar Key Florida.1954.1-94.4(1) ---, Texas J. Sci., A summer study of the biology and ecology of east Bay Texas. Part II. The fish fauna, and summary.1955.429-453.7 (4) RESÉNDEZ, M. A.,An. Inst. Biol. Univ. Nal. Autón. México. Ser. Zoología Lista preliminar de peces colectados en las lagunas de Nichupté y Bojorquez, Cancún, Quintana Roo, México.1975.87-100.46 (1) RICKER, W. E.,Computation and Interpretation of Biological Statistics of fish population. J. Fish. Res. Bd.Canada1975.1-382.191 SÁNCHEZ-GIL P., A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARES An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoDiversidad, distribución y abundancia de las especies y poblaciones de peces demersales de la Sonda de Campeche (Verano, 1978).1981.209-240.8 (1) SHIPP, R. L., Proc. Acad. Nat. Sci. Philad.Three new pufferfishes (Tetraodontidae: Sphoeroides) from the southern Caribbean.1972.129-134.124 (6) SHIPP, R. L. y K. W. YERGER Copeia Status, characters, and distribution of the northern and southern. puffers of the genus Sphoeroides. 1969.425-433.3 S.I.C., Catálogo de Peces Marinos Mexicanos. Secretaría de Industria y Comercio. Subsecretaría de Pesca, Instituto Nacional de PescaMéxico1976.462 p. SMITH, G. B., Dept. Nat. Res., Mar. Res. Lab., Publ.,Ecology and distribution of eastern Gulf of Mexico Reef fishes. 1976148.19 SPRINGER, S. y H. R. BULLIS Fish. Wild. Serv. Spec. Sci. Rept., Collections by the "Oregon" in the Gulf of Mexico. U.S.1956.1-134.196 SPRINGER, V. G. y K. D. WOODBURNProp. Pap. Ser. Alar. Lab. Fla., An ecological studies of the fishes of the Tampa Bay area.1960.1-104.1 SWINGLE, H. A., Alabama Mar. Res. Bull., Biology of Alabama estuaries areas: Cooperative Gulf of Mexico estuarine inventory.1971.1-128.5 TABB, D. C., D. L. DUBROW y R. B. MANNING Fla. St. Bd. Conserv., The ecology of northern, Florida Bay and adjacent estuaries.1962.1-80.39 TYLER, J. C.,Osteology, phylogeny and higher classification of the fishes of the Order Plectognathi (Tetraodontiformes). CircularNOAA Tech. Rept. NMFS1980.1-422434 VARGAS MALDONADO, L, A. YÁÑEZ-ARANCIBIA y F. AMEZCUA LINARESAn. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México Ecología y estructura de las comunidades de peces en áreas de Rhizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos, Sur del Golfo de México. 1981.241-266.8(1) WALKER, T. H. y W. R. NELSONAlabama Dep. Conserv., Seafoods Div. Typed. File., Rep.: Occurrence of vertebrates in the 1963-64 estuarine survey. In: Preliminary Marine Ecological Study of Mobile Bay.1964.138-184. YÁNEZ-ARANCIBIA, A., In: Informe Final 2da. Etapa Programa Uso de la Zona Costera de Michoacán y Guerrero. Convenio Comisión del Río Balsas, S. R. H. y Centro de Cienc. del Mar y Limnol Univ. Nal. Autón. México. Contrato de Estudios No. OC-E-03-74Relaciones tróficas de la fauna ictiológica del sistema lagunar costero de Guerrero y aspectos parciales de dinámica de poblaciones de peces de importancia comercial. 230 p., 1975. 750 p. 71 figs. ---, Tech Pap. Mar. Sci., Ecological studies in Puerto Real Inlet, Laguna de Términos, México. Discussion on trophic structure of fish community in Thelassia testudinum banks. Proc. Sem. Duke Univ. Mar. Lab. Beaufort NC, USA, (UNESCO, IABO). UNESCO1981.191-232.33 Aug. 1978. YÁÑEZ-ARANCIBIA, A. y J. W. DAYOceanologica Acta (en prensa). Ecological characterization of Terminos Lagoon, a tropical lagoonestuarine system in the Southern Gulf of Mexico. ISCOL-UNESCO '81BordeauxFrance1982. Sept. 7-14, 1981. YÁÑEZ-ARANCIBIA, A., J. CURIEL-GÓMEZ Y V. LEYTON An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. MéxicoProspección biológica y ecológica del bagre marino Galeichthys caerulescens (Gunther) en el sistema lagunar costero de Guerrero, México (Pisces: Ariidae). 1976.125-180.3 (1) YÁÑEZ-ARANCIBIA, A., F. AMEZCUA LINARES y J. W. DAY Estuarine Perspectives.Fish community structure and function in Terminos Lagoon, a tropical estuary in the southern Gulf of Mexico. In: Kennedy, V. (Ed.) Academic Press Inc.,New York1980.465-482. YÁÑEZ-ARANCIBIA, A., A. L. LARA-DOMINGUEZ, D. SÁNCHEZ-GIL, J. VARGAS, P. CHAVANCE, F. AMEZCUA, A. AGUIRRE Y S. DÍAZ Intern. Symp. Coastal Lagoons France, Oceanologica Acta (en prensa). Ecosystem dynamics and nichthemeral and seasonal programming of fish community structure in a tropical estuarine inlet, Mexico.1982.
|