|
VARIATIONS IN NATURAL
POPULATIONS OF NERITINA LATISSIMA BRODERIP IN COSTA RICA. (GASTROPODA:
NERITIDAE)
Trabajo recibido el 30 de enero de 1981 y aceptado para su
publicación el 25 de agosto de
1981.
MARTA F. VALDÉZ
Laboratoire de Zoologie, Ecole Normale Supéricure, 46 Rue
D'Ulm/75230 París. France.
CARLOS R. VILLALOBOS
Centro de
Ciencias del Mar y Limnología. Universidad de Costa Rica. Costa
Rica.
Se estudia la variación
temporal y geográfica de los morfos de Neritina
latissima, en relación con las características
físicas y químicas del ambiente, en la costa del Pacífico
de Costa Rica. Se analizan asimismo, en forma comparativa, la estructura de
población y la intensidad de depredación.
Los
resultados indican que la variación temporal, así como la
distribución espacial no parecen estar relacionadas con los factores de
tipo climático, físico o químico. La fluctuación en
las épocas de aumento de la fracción juvenil, así como las
diferencias en la composición de los grupos poblacionales, tanto dentro
como entre localidades, muestra que no existe sincronización en las
ciclos reproductivos de estas poblaciones.
Únicamente
mecanismos biológicos selectivos de depredación actúan
sobre la variación fenotípica en esta especie, en que los morfos
más llamativos o contrastantes, en relación con la
coloración del sustrato, son los más
depredados.
This work
analyses temporary and geographic variations exhibited by individuals of
Neritina latissima with respect to the physical and chemical
characteristics of the environment, on the Pacific coast of Costa Rica. The
population structure and predation intensity within individuals is also
analysed.
The results indicate that both variation in time
and geographical distribution do not appear to be direct1y related to
environmental factors.
No synchronization exists in the
reproductive cycles of the different population groups, as shown by the results
on the fluctuations during time at which juveniles become numerous, as well as
from the observed differences in the composition of the population groups, not
only within but also between locations.
It appears that only
selective biological mechanisms influence the phenotipic variation in this
species in wich, the more attractive or constrasting forms in regard to the
color of the substrate, show more intensive predation
rates.
The genusNeritina exhibits a tropical distribution (Keen, 1971). Common species include reclivata, virginea, punctulata and piratica, distributed from Louisiana in the Gulf of México, through the West Indies (Warmke and Abbott, 1962); layardiin India (Govindan and Natarajan, 1974) and latissima, from Acapulco, Mexico through Guayaquil, Ecuador (Keen, 1971). Most species are found inhabiting rivers and estuaries where they show a great variability in color pattern (Fig. 1). Very little work has been done on the ecology ofNeritina. Thus, Day, Smith, Wagner and Stowe (1973) studied the community structure of N. reclivata in a salt marsh in Louisiana (USA). They found the species to be an important functional component of the ecosystem. Govindan and Natarajan (1974) studied the ecology and life cycle of N. layardi. 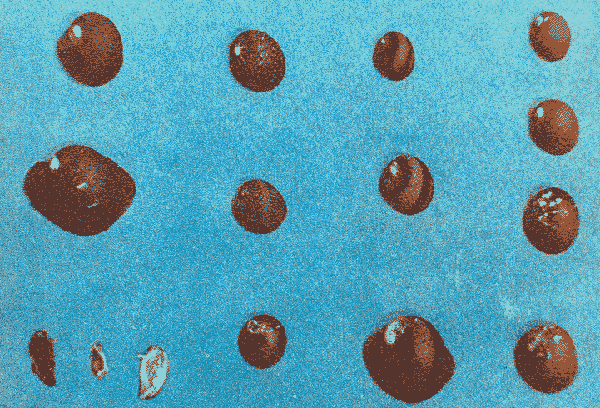 Regarding N. latissima Broderip (1833) described the species; Keen (1971) established its distribution and Little (1972) has presented some information on the problem of osmoregulation. Considering thatN. latissima is a species that inhabits estuarine systems and is actively colonizing fresh-water environments, exhibiting in addition, a great deal of phenotypic variation, it seems to be a group of particular interst. This work presents the results of the geographical and temporary variation of different phenotypic froms, their relation to the physical and chemical characteristics of the environment and the resulting population structure. AREA OF STUDY AND METHODSeveral localities in the Nicoya Península, Puntarenas Province, Costa Rica (9°40'N – 85°00'W), were selected for this study (Fig. 2). These included sites from Río Pánica, Bahía Ballena to Río Lajas, Cabuya. The mains study sites were Río Pánica and Quebrada Colorada. At these two areas, field work was conducted from April, 1977 through December, 1978. Remaining locations were sampled only in December, 1978. 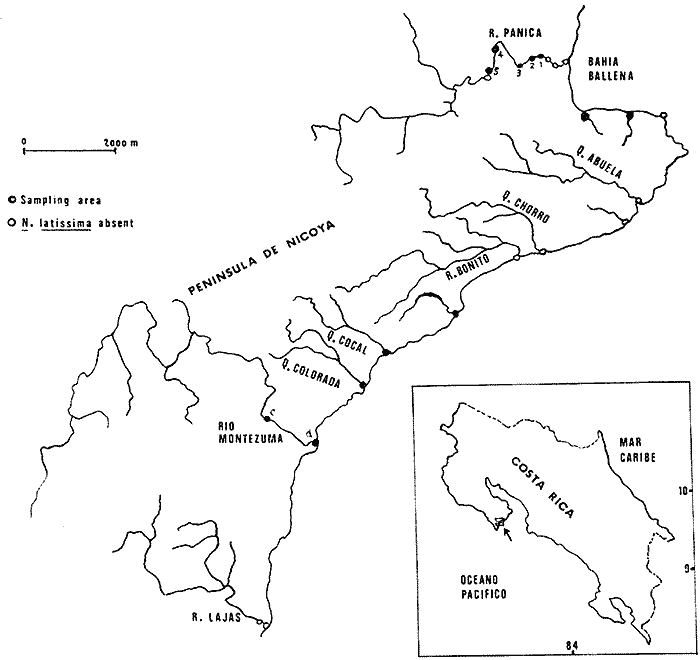 Fig. 2. Area of study and sampling localities. Puntarenas Province, Costa Rica. All locations, except Ro Pánica show an irregular rocky substrate with shallow pools scattered throughout and a rapid flow. Most rocks are of calcareous type and covered by microscopic algae and fine sediment. Río Pánica is a shallow body of water that exhibits daily fluctuations in depth due to the incoming tide. The substrate is primarily composed of pebbles which are also covered by algae. In addition, it shows a high content of Pirolusite (MnO2). Climatological data was obtained from. the Servicio Meteorológico Nacional and a field station located in Pochote, approximately 4 Km NE of Río Pánica. Water temperature was recorded every month at the main study sites. Similarly, using a modification of the Winkler method, water samples were analyzed to determine, Oxygen concentration. Clorinity was also determined using the Mohr technique. Occasionally, samples were taken to analyze Calcium concentration. This was done by means of an atomic absorption spectrophotometer. The structure of the radula from different morphos was compared using a scanning microscope Hitachi HHS-2R. In orden to study the morpholigical development ofN. latissima, individuals were collected in the field and placed in tanks in the laboratory. Counts of capsule were made and the different stages were determined. At Quebrada Colorada a random. sample that included all available sizes, was tagged using adhesive numbers. Each individual number was covered with a drop of "Loctite", a quick-set adhesive. The individuals were placed inside concrete pools built on the creek. Every month the length and width was recorded by means, of a Mitutoyo dial caliper (0.05 mm graduation). Frequency of females in the different population groups was indirectly determined from electrophoretic analysis of the locus of Glucose 6-Phosphate Dehydrogenase (Valdéz and Barrantes, 1980). The relative density of N. latissima was determined every three months at Quebrada Colorada and Río Pánica, using quadrats 0.25 m x 0.25 m, randomly distributed. At both sites, also every three months the frequency of the different morphos was recorded. This allowed to evaluate the role of predators, by looking at the kind of damage produced in the shell (Valdés and Barrantes, 1980). Quantitative analysis of the presence of Manganese was, also done and is reported elsewhere (Valdés and Briceño, 1980). RESULTSTable 1 shows climatological, physical and chemical characteristics of the two mains, study sites. As expected, no seasonal variations, were observed. However, when both sites are compared there are differences in the concentration of Oxygen and Chlorinity. Higher values of dissolved Oxygen at Quebrada Colorada are dueto, a more rapid flow there, as a consequence of the creek being considerably steep. At Río Pánica, higher Chlorinity values must be expected since it is subject to more intensive tidal effects. N. latissima seems to be a species with a wide range of tolerance to physical. and chemical parameters. Thus, Chlorinity values ranging from 1,982 mg/1 on intertidal pools down to 650 mg/l Río Pánica, were easily recorded. 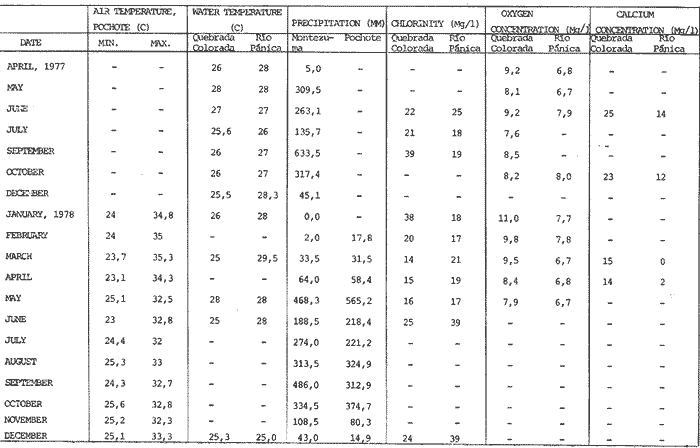 TABLE 1 CLIMATOLOGICAL, PHISICAL AND CHEMICAL CHARACTERISTICS OF THE STUDY AREAS 1977-1978 Higher values of Calcium wree always obtained in Quebrada Colorada. Figure 3 shows the structure of the radula of the common morphos as well as that of the individuals from Río Pánica (the so-called false, dark purple morpho). The radula must be clasified as ripidoglosadal according to Hyman (1967). It consists of a pair of central teeth bordered by two mandible-shaped teeth and, numerous marginal teeth. No significant differences were observed when the structure was compared within morphos. 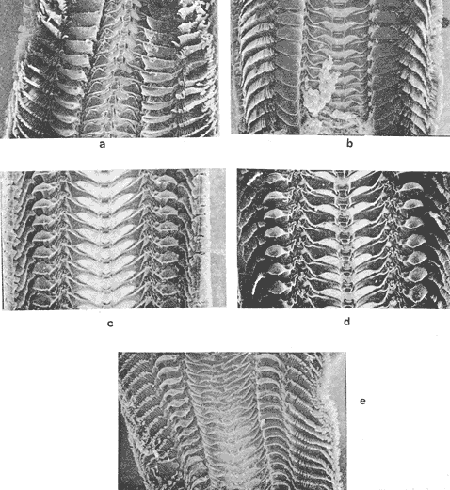 Fig. 3. Electron microscopy structure of the radula of differe with two black stripes, 40x; yellow with two black stripes, 75x; at morphos of N. latissima. (a) green morpho, 50x; (b) green (d) brown, 85x; (e) purple, 40x. Observations on the development of the embryos indicate that four blastomeres are produced within 144 hours after capsules are deposited. At 192 hours, 40 to 50 ciliated embryos similar to those described by Andrews (1935) for other species of Neritina are observed. These resemble the veliger larvae of other molluks. After 28 days several individuals are noted inside each capsule. Each individual show a well defined shell, operculum and several tentacles that are progressively reduced to two. After two months the individuales are born. Table 2 shows the results on the mean annual growth for snails tagged at Quebrada Colorada. As expected, the small individuals exhibited the highest growth rate. Small less than 2 mm grew up to 11 mm within a year, thus representing the Zero year class. The data also show that those individuals with an initial size of 11 mm reached a size of 22 mm in the same period. These represent the One year old class. Individuals larger than 20 mm were fairly uncommon and the growth rate difficult to determine Similar results have been oftained by Frank (1969) with several marine gastropods and Villalobos (1980) with tropical cirripeds. 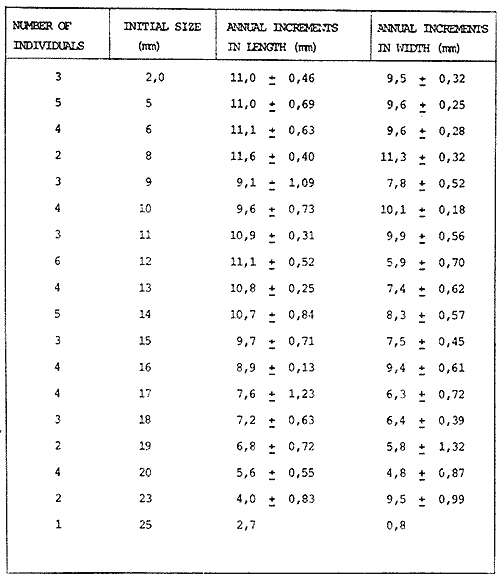 TABLE 2 MEAN ANNUAL GROWTH FOR DIFFERENT GROUP CATEGORIES OF INDIVIDUALS OF N. LATISSIMA. QUEBRADA COLORADA. 1977-1978 In general, the age structure of N. Latissima at Quebrada Colorada is characterized by a dominance of the individuals of one and two years of age (Fig. 4). Individuals with a length less than 5 mm showed very low frequencies, except in January and April of 1978 at wich, the group comprised between 10 and 15% of the population sample. However, these results do not appear to reflect the true size distribution. In fact, a certain percentage of the juveniles actually do exist, hidden in crevices, probably as response to the rapid flow of the creek. Hence, they are not recorded by common sampling methods. 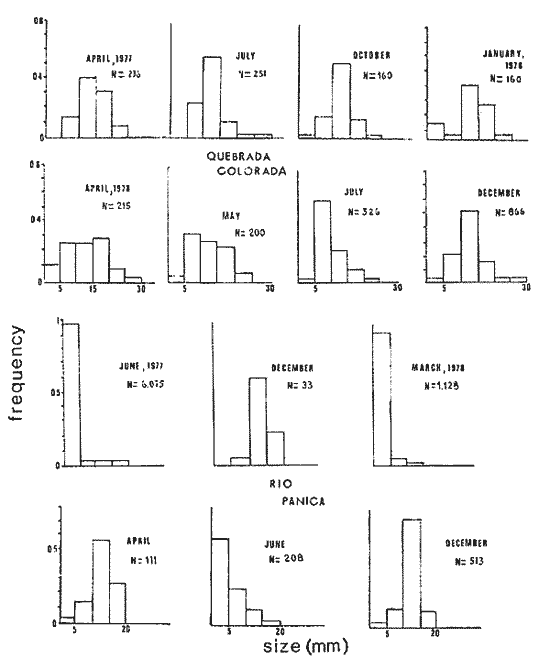 Fig. 4. Temporal variation in the frequency of different size categories in N. latissima. Quebrada Colorada and Río Pánica. 1977-1978. This contention is supported by the fact that at Río Pánica, with a very slow current and a different substrate, juveniles are found in percentages as high as 62 and 99% (Fig. 4). Observations in other localities were done during December, 1978. The results are presented in Figure 6. At Río Pánica, a decrease in group sizes less than 10 mm is observed as collecting stations move away from the mouth of the river. In Río Montezuma, at the continental site, the population in composed of individuals of 20 to 30, mm. These show morphological differences when they are compared with individuals from other localities. Thus, in addition to a greater size, the shell is more circular shaped and the operculum exhibit a light orange coloration. Sampling throughout al localities indicated that migration between population groups is fairly uncommon. In Quebrada Abuela, Quebrada Charro and Río Bonito no individuals were found. Table 3 shows the results on the relative density, Lloyd's Mean Crowding Indice (*m) and the dispersion coefficient for individuals at both, Quebrada Colorada and Río Pánica. At the former, the relative density and Lloyd's Indice increase during the month of April and July and decrease in October, December and January. The same pattern is observed in Río Pánica but the measurements do not appear to be related to any climatological, physical or chemical characteristics of the environment. During the period of study the population showed a contagious distribution. At Quebrada Colorada, spacing accordina to size was observed between individuals (Fig. 5). Thus, snails with a length around 5 mm as well as those over 20 mm, are mainly found near the month and rarely in the upper parts of the creek (t = 3,28; p < 0,01). On the other hand, for sizes ranging between 6 and 19 mm no significant differences were observed between the two, areas (t = 0,065; p < 0,05). 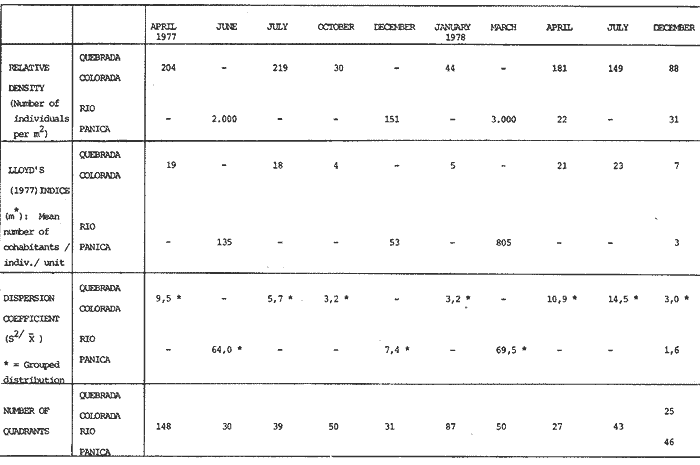 TABLE 3 RELATIVE DENSITY, LLOYD'S INDICE (m) AND DISPERSION COEFICIENT (S2/X) FOR INDIVIDUALS OF N. LATISSIMA AT QUEBRADA COLORADA AND RIO PANICA. 1977-1978. DISCUSSIONThe type of spatial distribution as well as the variation in time exhibited byN. latissima, during periods of increase in numbers, at both Quebrada Colorada and Río Pánica do not appear to be Telated to climatic, physical or chemical characteristics of the environment. The fluctuations observed during the time when juveniles become numerous, at a particular locality and between localities, as well as the observed differences in- the composition of the population groups (Figs. 4 and 6), indicate that no synchronization exists with regard to the reproductive cycles of the populations. A feasible explanation may be related to the Interaction of physiological mechanisms with fine local variations on certain environmental factors such as precipitation. Unfortunately, data of this kind is lacking for most of the tropical populations of mollusks. In intertidal arcas of the temperate regions, characteristics such as growth rate, size distributions, reproductive cycles and mortality are strongly influenced by such factors as tidal exposure and water temperature (Berry, 1961). 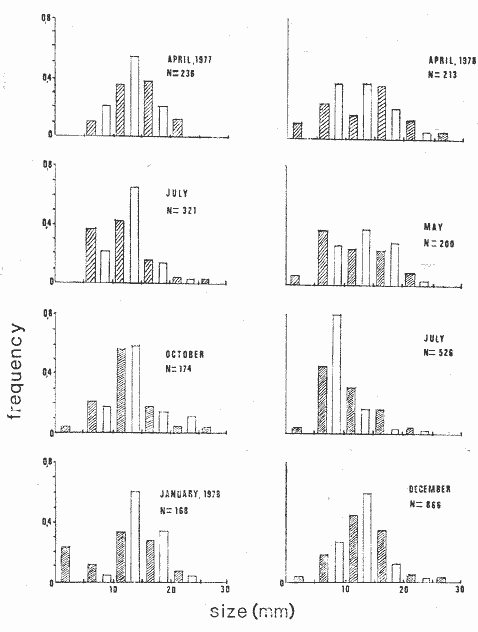 Fig. 5. Size distribution of individuals of N. latissima in relation to sampling area at Quebrada Colorada. 1977-1978. Shaded bars: mouth of creek. Unshaded bars: upper zone. 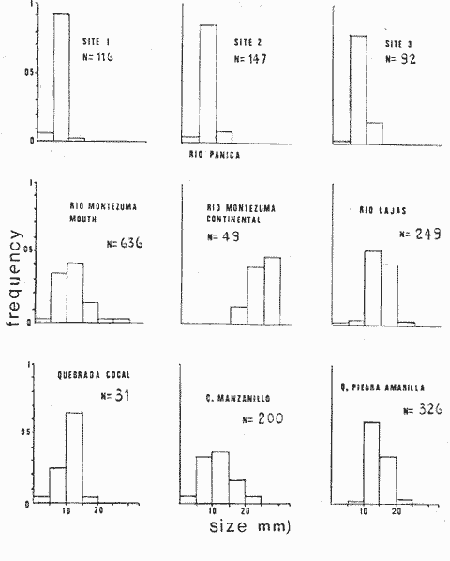 Fig. 6. Size distribution of individuals of N. latissima at several localities. December, 1978. Juveniles tend to concentrate at sites close to the mouth of the creeks (Fig. 5). At the continental site at Río Montezuma, however, neither juveniles nor egg capsules were observed, in spite of females being highly numerous. This may be dueto particular requirements in t terms of certain minerals and sand, which are necessary for the reinforcement of ,the egg capsule. Similar requirements have been defected by Andrews (1935) for other species ofNeritina. In spite of the difficulties in the evaluation of the frecuencies of juveniles, it may be noted that when present, they reach high frecuencies as observed in Río Pánica (Fig. 4). It is then suggested that the low number of individuals belonging to the largest size classes is the result of high predation rates, particularly by birds. As indicated, snails at the continental site at Río Montezuma exhibit external characteristics that make the individuals easy to separate from those of other geographical areas. Individuals are circularshaped, show a bigger size, have a light orange operculuin and exhibit a single coloration pattern. Considering that predominant sizes there are over 20 min and the fact that no egg capsules were found, it seems appropiate to assume that this group has recently colonized the area and therefore, neither regulatory mechanims at the system level, nor adaptative strategies have been developed to assure a successful colonization. In general, it may be considered that evolutionary changes in N. latissima are the Tesults of the interaction of several factors at the genetic and the environmental level. The study of such interactions becomes, then, of prime importancel in order to determine the relative contribution of each factor to the evolutionary biology of this species. AgradecimientosACKNOWLEDGMENTS We like to express our thanks to the staff of the Research Center on Cell and Mollecular Biology at the University of Costa Rica, as well as to the staff of the Electron Microscopy Unit at the same Institution. Thanks are also given to Miss Elizabeth Barrantes, Luis Cruz, Walter González, Jesús Rojas, Enrique Lahmann, Jorge Jiménez, Clemencia León and Fernando Cratz, for helping us in collecting some of the information on the field and in the laboratory. This work was partially supported. by the Costa Rica National Council for Science and Technology; the University of Costa Rica and the Organization of American States. LITERATURAANDREWS, E., The egg capsules of certain Neritidae. J. Morphol.1935.31-54.57 BERRY, A., J. Anim. Ecol., Some factors affecting the distribution of Littorina saxatilis.1961.27-45.30 BRODERIP, W. J., Proc. Zool. Soc. London.Characters of new species of Mollusca and Conchifera.1833.172-202. DAY, J. W., W. C. SMITH, P. R. WAGNER and W. C. STOWE Community structure and carbon budget of a salt marsh and shallow bay estuarine system in Louisiana. Pub. LSU-SG72-04.1973.80 p. FRANK, P.,Oecologia Growth rates and longevity of some gastropod mollusks on the coral reef at Heron Island.1969.232-250.2 GOVINDAN, K. and R. NATARAJANProc. Indian Nat. Sci. Acad. Part B, Biol. Sci.Studies in Neritidae (Neritacea: Prosobranchia) from Peninsular India.1974.225-239.38 HYMAN, L., The Invertebrates. Vol. VI. Mollusca I.Mc. Graw-Hill Book Co.1967. 792 p. KEEN, A. M., Sea Shelsl of Tropical West América. Stanford University Press.1971.1064 pp. LITTLE, G., J. Exp. Biol., The evolution of kidney function in the Neritacea (Gastropoda: Prosobranchia).1972.249-261.56 VALDEZ, M. F. and R. BARRANTESRev. Biol. Trop.,Variación geográfica y alozimica en Neritina latissima (Gastropoda: Neritidae) en Costa Rica.1980.181-192.28 VALDEZ, M. F. and J. BRICEÑO Rev. Biol. Trop.Incorporación de manganeso en Neritina latissima (Gastropoda: Neritidae) en Costa Rica.1980.175-180.28 VILLALOBOS, C. R., Rev. Biol. Trop.Variations in population structure in the genus Tetraclita (Crustacea: Cirripedia) between temperate and tropical populations. IV. The age structure of T. stalactifera and concluding remarks.1980. 353-359.28 WARMKE, G. L. and R. T. ABBOTTCaribbean Sheashells Dover Publications, Inc. N. Y.1962. 348 p.
|