|
CICLO ANUAL DE MADURACIÓN Y
REPRODUCCIÓN DEL "CHACAL" MACROBRACHIUM TENELLUM Y SU RELACIÓN
CON FACTORES AMBIENTALES EN LAS LAGUNAS COSTERAS DE MITLA Y TRES PALOS,
GUERRERO, MÉXICO. (DECAPODA: PALAEMONIDAE)
Trabajo recibido el 13
de noviembre de 1980 y aceptado para su publicación el 2 de julio de
1981.
MANUEL GUZMÁN ARROYO
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología.
Contribución 275 del Instituto de Ciencias del Mar y Limnología,
UNAM.
JOSÉ LUIS ROJAS GALAVIZ
Universidad Nacional Autónoma de
México, Instituto de Ciencias del Mar y Limnología.
Contribución 275 del Instituto de Ciencias del Mar y Limnología,
UNAM.
LUIS DANIEL GONZÁLEZ GONZÁLEZ
Universidad Nacional
Autónoma de México, Instituto de Ciencias del Mar y
Limnología. Contribución 275 del Instituto de Ciencias del Mar y
Limnología, UNAM.
El presente trabajo se
desarrolló dentro del periodo mayo 1975 a abril 1976, realizando
muestreos mensualmente en las dos lagunas. Se colectaron un total de 3,732
ejemplares durante el estudio. Se encontró que la proporción
sexual se mantiene en una relación de 1:1 solamente en una
estación de la Laguna de Mitla (Camalote). Se apreció que aumenta
para las hembras durante la temporada reproductiva y la de machos aumenta fuera
de ella. En el resto de las estaciones existen diferencias significativas no
manteniéndose esta proporción. La longitud total media a la cual
las hembras alcanzan la madurez sexual es de 74 mm en la Laguna de Tres Palos
(Arenal) y de 93 y 97 mm en la Laguna de Mitla (Camalote y Papayo). La
temporada de reproducción se presenta en ambas lagunas de junio a
noviembre, encontrando un pico máximo de reproducción en octubre
siendo igual la proporción de hembras maduras y ovígeras, para
ambas lagunas. Al determinar y analizar los ciclos anuales de fotoperiodo,
temperatura, precipitación pluvial y salinidad, se estableció una
correlación entre los distintos estadios de madurez sexual de las
hembras y los diferentes parámetros ambientales.
Se
establece mediante un diagrama temperatura/ salinidad, las condiciones
óptimas donde se presenta la mayor concentración de hembras
maduras y ovígeras.
Por último se encuentra
que es posible establecer un pronóstico de la proporción esperada
de hembras maduras y ovígeras, bajo condiciones diferentes de
temperatura y salinidad.
The present work was developed
from May 1975 to April 1976, monthly sampling was performed in both lagoons. In
this period of sampling there were obtained 3,732 individuals of freswater
prawn. The sexual composition analysis of the population, showed that in the
Mitla Lagoon (Camalote sampling spot), the sexual ratio founded was 1:1, the
female ratio increased during the reproductive period, on the other hand, the
male ratio decreases in this period, reaching the highest ratio in the
non-reproductive period. The total median lenght of females at which sexual
maturity was reached is 74 mm for the Tres Palos Lagoon and of 93 and 97 mm for
the Mitla Lagoon. The reproductive period is well defined in both lagoons, it
begins in June; reaching it's highest values during October being then equal,
the proportion of mature and egg-bearing and ending in
November.
By means of data analysis of the environmental
parameters (photoperiod, temperature, rainfall and salinity) and distinct
sexual maturity stages of females, it has been found that there is a high
significant correlation between the number of females in each stage and
rainfall. It has been found in a temperature/salinity diagram, the optimum.
conditions of maximum concentrations, of mature and eggbearing females.
Finally, it is establish a pronostic of the expected ratio of mature and
egg-bearing females, under distinct temperature and salinity
conditions.
La reproducción en su sentido más amplio, implica la aparición de nuevos individuos en la población. El proceso de la reproducción suele ser en general un proceso discreto, es decir discontinuo en el tiempo. En efecto la mayor parte de la producción de la progenie en una población suele limitarse a un intervalo generalmente corto del periodo de vida de los animales, encontrándose desde luego en el mundo animal un rango bastante amplío sobre lo que puede representar el intervalo de reproducción (Rabinovich, 1980). Aún considerándose el proceso discontinuo de la reproducción, se puede observar que en la mayor parte de los grupos animales, la intensidad, duración y la modalidad en que se lleva a cabo dicho proceso suele ser sumamente variable, incluso para la misma especie. El proceso de reproducción si bien está sujeto también a una cantidad de variables extrínsecas, presenta modalidades muy ligadas a las características de las especies. En general, los organismos deben responder a los cambios medioambientales, utilizando la estrategia reproductiva más adecuada para asegurar la supervivencia de la progenie. La estrategia reproductiva que mantiene en equilibrio a la población, depende fundamentalmente de aspectos tales como, supervivencia, fecundidad, densidad de la población, estructura sexual y tiempo de vida (Pianka, 1974). El objetivo del trabajo, es conocer el comportamiento del ciclo reproductor de la especieMacrobrachium tenellum, comparativamente en ambas lagunas, y su relación con los parámetros ambientales. ÁREA DE ESTUDIOLas lagunas costeras de Mitla y Tres Palos en el Estado de Guerrero, se encuentran dentro de la planicie costera, área delimitada al sur por el Océano Pacífico y hacia el norte por la Sierra Madre del Sur, y dentro de los 16° 43' y los 17° 05' de Latitud Norte, y los 99° 39' y los 100° 25' de Longitud Oeste. El clima es tropical húmedo, con una temporada de lluvias en verano, un invierno seco con temperaturas relativamente altas, mayores a 18°C, corresponde al Aw, el más seco de los tropicales húmedos (García, 1973). A lo largo de la costa del Pacífico Mexicano, existe un flujo de aire marítimo del SE durante la primavera y el verano, este flujo se invierte en invierno proviniendo en forma dominante del NE, los vientos marítimos del verano llevan humedad a la costa, en invierno son vientos secos hacia el mar, los vientos convectivos costeros son muy efectivos como agentes de evaporación local durante la época seca y son probablemente los responsables del abatimiento del nivel de agua en las lagunas costeras y consecuentemente del incremento de su salinidad (Lankford, 1974). La Laguna de Mitla se encuentra a 75 Km al noroeste de Acapulco, con una longitud de 45 kilómetros en su eje mayor y 8 Km en su parte más ancha Tres Palos, aproximadamente a 20 Km al sureste de Acapulco, con 30 Km en su eje mayor y 15 Km en la parte más ancha. Mitla ha perdido su comunicación con el mar, en cambio Tres Palos la mantiene a través de un estrecho y tortuoso canal al sur de la laguna (Fig. 1). MATERIAL Y MÉTODOSEl material fue colectado durante el periodo comprendido entre el mes de mayo de 1975 y el mes de abril de 1976 durante dicho periodo se colectaron un total de 3,732 ejemplares de "chacal" (Macrobrachium tenellum) de los cuales 2,125 corresponden a la Laguna de Tres Palos y 1,607 a la Laguna de Mitla. Fueron seleccionadas cuatro estaciones de muestreo por la importancia que tienen dentro de las pesquerías regionales, para la Laguna de Mitla fueron Camalote y Papayo, para la Laguna de Tres Palos, El Arenal y San Pedro de las Playas. Las dos primeras en ambas lagunas se tomaron como estaciones principales y las segundas como auxiliares. 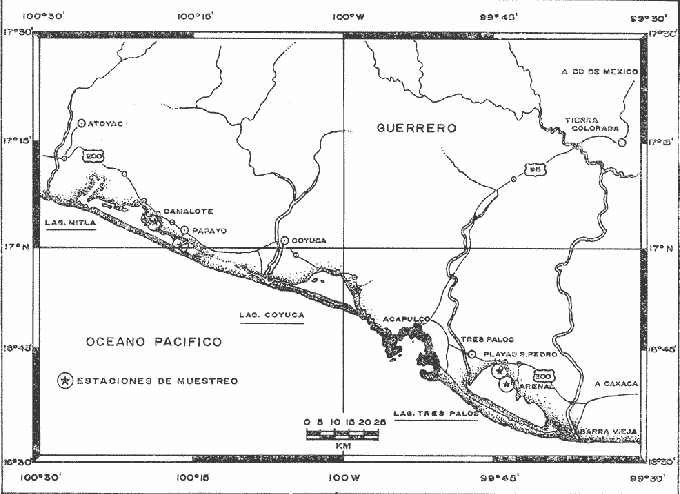 Fig. 1. Localización del área de estudio y estaciones de muestreo. El Arte de Pesca utilizado fue la Atarraya, de 5 m de largo y con una malla de 1 a 1.5 cm, ocasionalmente se utilizó un chinchorro playero de 25 m de largo por dos de alto. Simultáneamente se obtuvieron los siguientes parámetros: temperatura, salinidad, oxígeno malla de 1 cm. disuelto y profundidad. De los registros meteorológicos de obtuvo temperatura ambiente, precipitación y número de horas diarias de luz (Fotoperiodo). Fueron obtenidos los siguientes datos biomédicos: longitud total, longitud del cefalotórax en mm, peso en gramos, estado sexual y sexo. Se desarrollaron pruebas estadísticas, las más utilizadas fueron Chi cuadrado, la t de Student para diferencia de proporciones y análisis de varianza de dos vías, así como también correlaciones simples y múltiples para encontrar la posible relación entre los fenómenos de reproducción y los parámetros ambientales, utilizando también la prueba de bondad de ajuste para las distribuciones normales. RESULTADOS
COMPOSICIÓN SEXUALEn la Tabla 1 se muestra la proporción sexual de las hembras para cada uno de los meses de estudio y para cada estación de muestreo. A partir de la información sobre la composición por sexos de la población (Fig. 2), se elaboró un análisis estadístico para probar la diferencia en las proporciones sexuales en cada estación de muestreo, utilizando la distribución teórica de "Z", para ensayo de una cola, con un nivel de significación (Alfa) de 0.10, y se encontró que solamente en la estación Camalote de la Laguna de Mitla, la proporción sexual es de 1: 1, mientras que en las demás estaciones existen diferencias significativas, no manteniéndose esta proporción (Fig. 3). Al buscar diferencias en las proporciones sexuales de las dos lagunas, mediante una prueba de T de Student, se encontró que, a un nivel de significación de 0.05, sí existen diferencias en la proporción, tanto entre estaciones como entre las dos lagunas. La proporción total anual de hembras es la siguiente en cada estación de muestreo:  Las hembras dominaron en el mes de noviembre en Camalote, y en el mes de agosto en Arenal, en agosto y octubre en Papayo, en cambio en San Pedro de las Playas, nunca rebasaron el 40%, y de septiembre a marzo prácticamente no se encontraron. Dado que en Papayo se seguía el ciclo de pesca comercial éste se suspendió a partir de noviembre por lo cual no hay datos para esos meses. MADUREZ SEXUALPara la determinación de la talla del 50% o mediana (L50) de madurez sexual, se utilizó una curva acumulativa de la distribución de frecuencias expresadas en porciento (Fig. 4) tomándose aquella longitud como la talla de maduración sexual correspondiente al estado III (Hembras ovígeras). 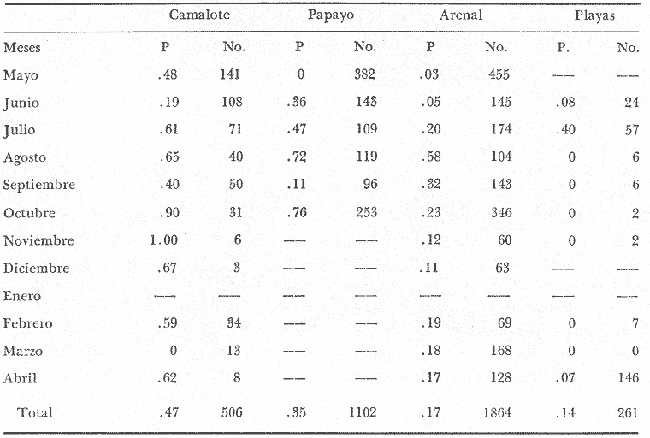 TABLA 1 PROPORCIÓN MENSUAL DE HEMBRAS EN LAS CUATRO ESTACIONES DE MUESTREO 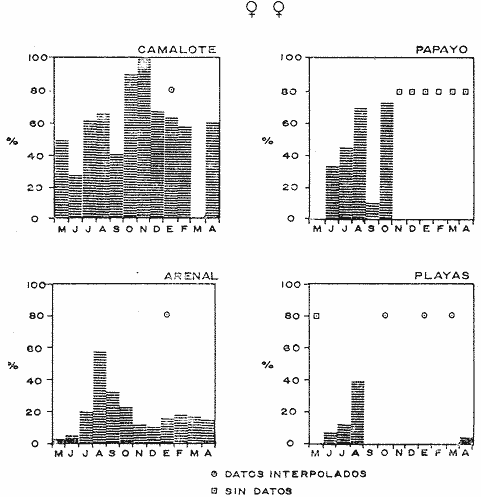 Fig. 2. Porcentaje de hembras en las cuatro estaciones de muestreo. 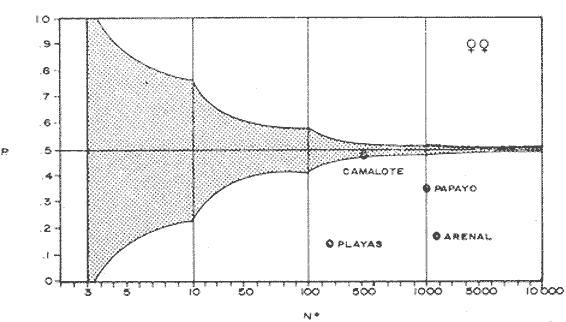 Fig. 3. Proporción de hembras en las cuatro estaciones de muestreo. Encontrando los siguientes valores en cada estación:  Se observa que los valores de la mediana y de la media son cercanos, la máxima varianza fue encontrada para Camalote y la mínima para Papayo, pero esto es debido a que en esta última estación se muestrearon las capturas comerciales con un evidente sesgo hacia los tamaños mayores. El coeficiente de variación son similares entre Camalote y Arenal, encontrándose más bajo en Papayo por las mismas razones aludidas anteriormente. El tamaño de L50 varía notoriamente entre la Laguna de Mitla (Camalote y Papayo) respecto al de Tres Palos (Arenal), asimismo, se encontró un valor muy bajo en el intervalo de 40 a 45 mm en la estación Camalote, esto debe considerarse como un registro extraordinario. SECUENCIA REPRODUCTIVA.Si se grafica la proporción de hembras en cada uno de los estadios II y III es posible observar como en cada uno de los estadios se forman distribuciones normales. Los datos observados se ajustaron bien a una distribución normal, a un nivel de significación (Alfa) de 0.05, e ajuste se realizó para dos estaciones y para cuatro estadios de madurez sexual y se observa cómo se sucede una a la otra incrementándose, llegando a un máximo y luego disminuyendo en la misma proporción que el siguiente se incrementa, esto se observa en hembras II y III (Fig. 5) siendo similares para ambas lagunas. 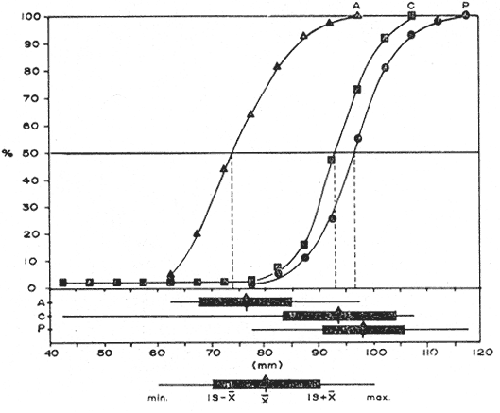 Fig. 4. Curva acumulativa de la distribución de frecuencia de longitud total de hembras. (A: Arenal, C: Camalote, P: Papayo). TEMPORADA DE REPRODUCCIÓNAnalizando la misma figura 5, se observa en la parte inferior de la misma los límites en la escala de tiempo mensual que indica la duración de cada estadio, las líneas cruzadas indican el tiempo medio y los cuadros enmarcan una desviación estandard a cada lado de la media. 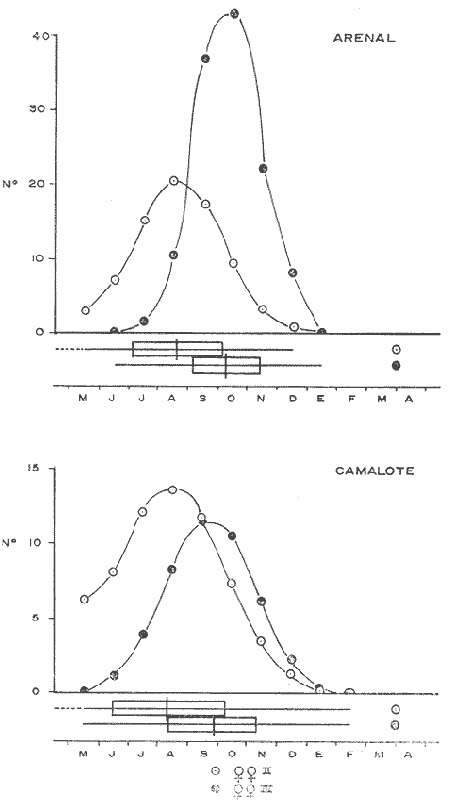 Fig. 5. Distribución normal de la proporción de hembras maduras (II) y ovígeras (III). En el siguiente cuadro se ven las principales características estadísticas de la temporada de reproducción. Se ha considerado como tiempo cero el primero de mayo, siendo el primer mes del estudio, y los datos se presentan como días a partir de dicha fecha, esto es Tiempos Reales. 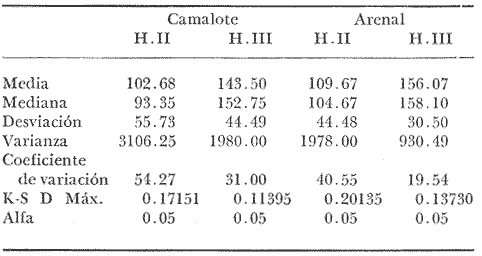 De acuerdo a lo anterior, el punto de la. temporada para hembras maduras (H II) corresponde al mes de agosto en ambas estaciones, y para hembras ovígeras (H III) a finales de septiembre en Camalote (Mitla) y principios de octubre en Arenal (Tres Palos) este desfasamiento es debido al ajuste, ya que las modas (picos) corresponden en ambos casos al mes de octubre tal y como corresponden a los datos observados. Trusdale y Mermilliod (1979) mencionan para M. ohione en ríos costeros de Louisiana, que la mayor proporción de hembras ovígeras se presenta en abril y mayo. Los resultados de las frecuencias observadas y teóricas para los estadios II y III son: 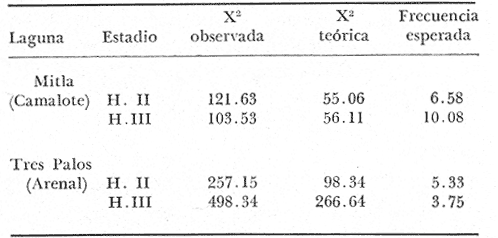 Para todos los casos se tiene que la X² observada es mucho mayor que la esperada teóricamente, que es igual para todos los casos X²0.05 = 18.307 para 10 grados de libertad rechazándose la hipótesis nula de que las frecuencias observadas son iguales en todos los meses del año, a un nivel de significación (Alfa) de 0.05. Lackey y Hubert (1977) utilizando el coeficiente de correlación de Pearson, determinan la similitud entre dos poblaciones, con el uso de alguna variable (peso, edad, etcétera) utilizando el mismo concepto se ha correlacionado la proporción de hembras para cada estadio y para cada laguna, encontrándose los siguientes resultados:  El ajuste se hizo con una ecuación tipo potencial de la forma: Log. Y = In a + b In X., previa transformación de los valores de X e Y (X + 1 0 Y + 1). Para una mayor certidumbre sobre la igualdad de los ciclos reproductivos en ambas lagunas se desarrollaron otros métodos de análisis estadísticos, uno, de ellos fue mediante un análisis de varianza (Anova) de dos vías solamente para hembras maduras y para hembras ovígeras, y para cada mes del estudio. Para hembras maduras se obtuvo la siguiente tabla de Anova; con una transformación de (X + l).  Para hembras ovígeras se obtuvo: 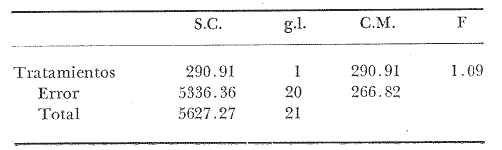 Siendo para ambos casos la F teórica igual a: F0.05 (1) 20, 21 = 2.10 Siendo mayor que las F observadas (0.09 y 1.09) por lo tanto las muestras comparadas no son significativamente diferentes a un nivel de significación (Alfa) de 0.05. Como un complemento de estos análisis se utilizó un test de igualdad de dos porcentajes (Sokal y Rohlf, 1969) únicamente para comparar los máximos (picos) de las distribuciones tanto de hembras maduras como ovígeras, de acuerdo a la siguiente ecuación: 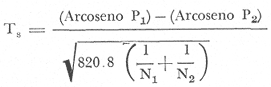 Donde P1 y P2 son las proporciones de las muestras comparadas, 820.8 es una constante que representa la varianza paramétrica de una distribución de proporciones con transformación. Encontrándose:  Siendo para hembras II la T teórica de: T0.05 (86) = 1.991 Y para las hembras III: T0.05 (108) = 1.984 Dado que la T observada es menor que la teórica esperada, se puede afirmar a un nivel de significación de 0.05, que las muestras son significativamente iguales. PARÁMETROS AMBIENTALESFotoperiodo. El fotoperiodo se considera como el número de horas de iluminación diaria, que se reciben sobre una área dada, las diferencias a lo largo del año en el número de horas de iluminación se debe a la declinación solar, siendo mayor durante el solsticio de verano y menor durante el de invierno, dada la cercanía de las lagunas este fenómeno se considera similar para ambas, de acuerdo a Granados (1980) es en mayo cuando se presenta el mayor fotoperiodo con 13.17 hrs. al día y en diciembre el menor con 10.48 hrs. (Figs. 6 y 7). Temperatura. La temperatura del agua, al igual que el fotoperiodo varía de acuerdo a las estaciones del año en íntima relación con la temperatura ambiente para Camalote se encontró la máxima temperatura del agua en el mes de agosto con 32.9°C y la mínima para el mes de diciembre con 29.5°C. Viendo las gráficas de las figuras 6 y 7 se observa que en general se mantienen más altas las temperaturas que en Arenal (Figs. 6 y 7). 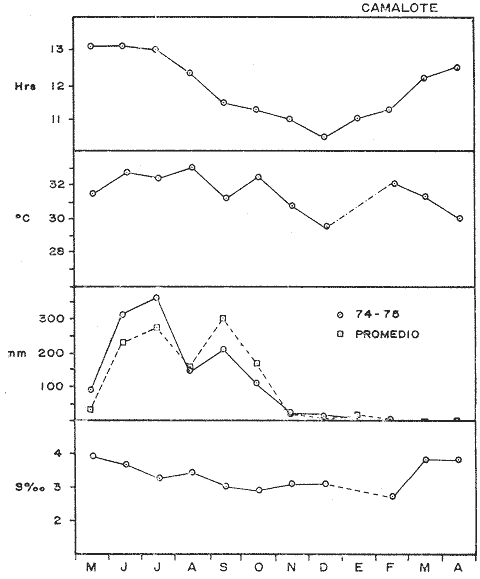 Fig. 6.Fotoperiodo, temperatura del agua, precipitación pluvial y salinidad en la Laguna de Mitla. Estación Camalote. 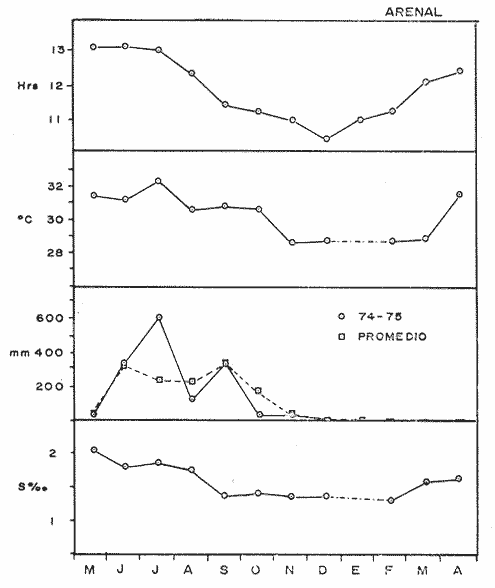 Fig. 7. Fotoperiodo, temperatura del agua, precipitación pluvial y salinidad en la Laguna de Tres Palos. Estación Arenal. Precipitación. Para el análisis de este parámetro se procedió en dos formas; la primera con los datos de precipitación que corresponden a la época del estudio señalado como 75-76 y la segunda con el promedio registrado para los años de recopilación de datos de cada una de las estaciones meteorológicas del área (García, 1973). Para Camalote y Arenal la diferencia entre estas dos series de datos no es muy marcada manteniéndose el mismo patrón, lluvias en verano con un mes intermedio (agosto) de menor precipitación (Canícula). Respecto a la precipitación entre estaciones correspondientes al periodo del estudio, no hay diferencia en la época, ya que la máxima precipitación ocurrió en Camalote en el mes de julio (363 mm) y en Arenal también (613 mm), en el promedio de 37 años para Arenal y de 8 para Camalote, se observa que la máxima precipitación ocurre en el mes de septiembre con 300.9 mm para Camalote y 353.9mm para Arenal (Figs. 6 y 7). Salinidad. Los valores máximos de salinidad en ambas lagunas no sobrepasan a las 5%, por lo que de acuerdo a la clasificación de Venecia, se deben considerar como oligohalinas. En Camalote la máxima salinidad fue de 3.93 para el mes de mayo y la mínima de 2.95 para el mes de septiembre, para Arenal fue de 2.04 para el mes de mayo como máxima y como mínima 1.32 para el mes de febrero (Figs. 6 y 7). En general, se observa una interacción negativa de la salinidad respecto a la precipitación, esto es que a mayor precipitación menor salinidad, y con la temperatura es positivo, a mayor temperatura mayor salinidad, la Laguna de Mitla mantiene más altos niveles de salinidad que la de Tres Palos a pesar de no mantener comunicación con el mar tal y como lo hace Tres Palos, pero en cambio no recibe ríos caudalosos a diferencia de Tres Palos (Río de la Sábana) (Fig. 1). Diagrama T/S.Analizando los datos de temperatura y salinidad a través de un diagrama T/S (Fig. 8) se observa que en ambos parámetros la Laguna de Mitla mantiene valores más altos que la de Tres Palos, el ciclo se mantiene en forma más regular en Mitla, dado que en Tres Palos se ven dos grupos relativamente separados: uno que va desde noviembre a marzo y otro los meses restantes, en el primero los valores son muy bajos de ambos parámetros y poco diferentes entre sí, por el contrario en el otro grupo es más variable y de valores más elevados. 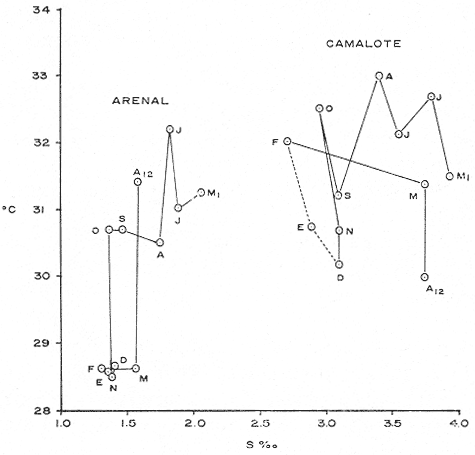 Fig. 8. Diagrama temperatura / salinidad, mensual en las dos lagunas. RELACIÓN ENTRE LOS PARÁMETROS AMBIENTALES Y EL CICLO DE MADURACIÓN Y REPRODUCCIÓN DEL CHACALSe desarrollaron coeficientes de correlación entre las variables ambientales (Fotoperiodo, Temperatura Salinidad y Precipitación), y la proporción de hembras maduras y ovígeras para cada una de las estaciones, con los siguientes resultados: Coeficiente de correlación de Pearson (r)  Observándose en general que las mayores correlaciones se encontraron para las hembras maduras con la temperatura y con la precipitación, esto es que: mayor temperatura y precipitación mayor grado de madurez. En la proporción de hembras también se encontró que dentro de la precipitación, la correlación fue mayor con la precipitación promedio, que con la registrada durante el estudio. Para las hembras ovígeras se encontró una mayor correlación con la salinidad pero negativa, para el caso del fotoperiodo no es clara la relación, ya que la mayor relación se registra con las hembras maduras de Camalote, pero también con las hembras ovígeras de Arenal. La regresión fue ajustada a una curva potencial del tipo: Lny = Lna + bln X. Con una transformación de N + 1. Substituyendo los puntos de la distribución del diagrama T/S por los valores encontrados del número de hembras para los estadios II y III, se desarrollaron las figuras 9 y 10. Respecto a las hembras II, se observa que el mayor número gravita en la estación Arenal entre los 30 y 31°C de temperatura y entre los 1.6 y 1.9 de salinidad, en cambio en la estación Camalote, es entre los 32 y 33°C y respecto a salinidad es alrededor de los 3.5%, En relación a las hembras III estos centros se desplazan, para Arenal quedan dentro del mismo rango de temperatura, pero para salinidad se reduce a 1.5%, respecto a Camalote queda dentro de 32.5°C y sobre los 3.0%. Estos diagramas nos dan una clara idea de la dispersión de los valores en número de dos dimensiones para las hembras en los estadios mencionados, dada la distribución bivariante observada. Considerando que las únicas variables ambientales que puede manejar el hombre prácticamente respecto al cultivo de esta especie, son la temperatura y la salinidad, se desarrolló un modelo de correlación lineal múltiple del tipo: Z = a + b X + c Y 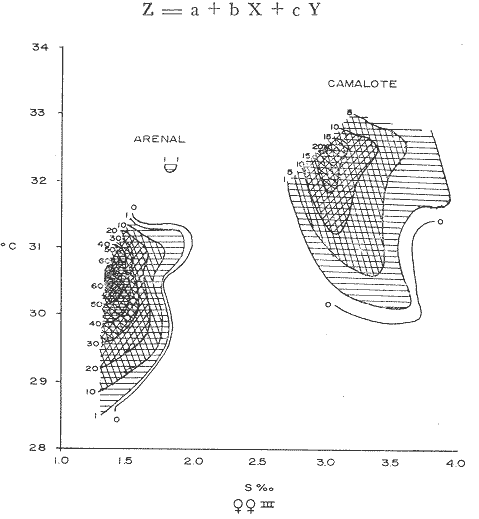 Fig. 9. Número de hembras maduras en las dos lagunas, representado en un diagrama temperatura/salinidad. 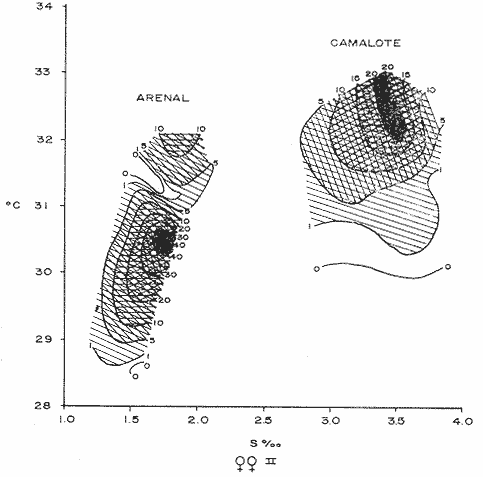 Fig. 10. Número de hembras ovígeras en las dos lagunas, representado en un diagrama temperatura/salinidad. Donde Z es la proporción de hembras ovígeras pronosticadas, X es la salinidad e Y la temperatura. Se encontró los siguientes resultados: Camalote: Z = 80.609 - 35.546 X + 1.811 Y, r = 0.544 Arenal: Z = -166.651 - 123.952 X + 12.970 Y, r = 0.652 Para ambas lagunas la r teórica es de: r (9), 0.05 = 0.521, como las r observadas son mayores que las teóricas se encuentra que los dos coeficientes son significativos a un nivel de confianza de 95% Manteniendo una mayor correlación en Arenal. Sustituyendo los valores de X e Y por los observados de salinidad y temperatura respectivamente, es posible saber o pronosticar la proporción de hembras esperadas, dentro del rango de valores observados. Sin embargo, debe considerarse el papel que juega el factor tiempo en los ciclos de maduración y desove, esto quiere decir que la temporada de desove y/o maduración tienden a ser cíclicas a pesar de variaciones de temperatura y salinidad que alteren el modelo desarrollado. Por otra parte al revisar bibliográficamente la distribución de M. tenellum se observó la estrecha relación que guarda su distribución respecto a la temperatura ambiente (Fig. 11), en especial respecto a la isoterma del mes más frío (18°C) como límite de los climas de latitudes medias encaja adecuadamente y va desde el sur del estado de Sonora y Baja California, México hasta el norte del Perú. DISCUSIÓNRespecto a la composición sexual fue observado que no se mantuvo la relación 1 : 1 en todas las estaciones de muestreo como era de esperarse y de acuerdo a Krebs y Davies (1978), en una población reproductora la relación numérica entre los sexos evoluciona hacia una estabilidad representada por la proporción 1 : 1, una desigualdad puede deberse a un mayor costo de una especie, en parte puede deberse a un dimorfismo sexual donde un sexo es mayor que otro. En nuestro caso interpretando los resultados se observa que sólo la estación Camalote se mantiene dicha proporción (a un nivel de confianza del 90%) y en las demás es manifiesta la desproporción, suponemos que estas diferencias están dadas por condiciones ecológicas, por ejemplo la estación Camalote es una ensenada de la Laguna de Mitla, en cuyos extremos las pendientes rocosas impiden la salida o entrada hacia esta ensenada, evitando así migraciones o intercambio de individuos, en la estación Papayo tal y como se menciona, se tomaron los datos de capturas comerciales con un evidente sesgo hacia los individuos de mayor tamaño, descriminándose los pequeños. En la estación Playas de Tres Palos, frente a la población del mismo nombre prácticamente alterada en cuanto a la destrucción del manglar, nunca hubo hembras ovígeras al no existir condiciones adecuadas para ellas, la estación Arenal manifestó un mayor número de hembras ovígeras que maduras, en condiciones de una población estable, dada la mortalidad es de esperarse un número menor de ovígeras, sugiriéndose una inmigración al área de hembras alterando la proporción esperada. Analizando el tamaño medio de hembras ovígeras, se encontró una diferencia marcada entre las estaciones de ambas lagunas, siendo semejantes los valores para Mitla y diferentes respecto a Arenal (Tres Palos), esto también difiere entre el número de individuos hembras presente en cada estadio, en general el número es mayor en Tres Palos que en Mitla pero en cuanto a tamaño es menor Tres Palos que Mitla. Se considera a esto como un fenómeno de densodependencia, esto es una mayor densidad de individuos en Tres Palos pero de menor tamaño, contra otra de menos individuos pero de tamaño más grande en Mitla, básicamente esto es una respuesta al medio ambiente local, disponibilidad de espacio y de alimento, manteniendo las poblaciones dos estrategias diferentes. Su secuencia reproductiva, de acuerdo a los análisis estadísticos comparativos, es similar en ambas lagunas, básicamente en las estaciones Camalote y Arenal, tanto en la proporción de hembras como en el periodo óptimo para cada estadio, lo que nos sugiere que las poblaciones responden a factores propios de la especie y a factores ambientales comunes a ambas lagunas, esto es factores regionales. Se considera parámetros ambientales regionales y por ende comunes a ambas lagunas el fotoperiodo y la precipitación y como parámetros locales o propios de cada laguna la salinidad y la temperatura. Analizando mediante correlaciones la variabilidad de estos parámetros respecto de los ciclos de maduración y reproducción, se encontró que la mayor relación es Con la precipitación y las hembras maduras (II) siendo mayor con la precipitación promedio que con la precipitación durante el estudio. También fue mayor la relación entre la temperatura y las hembras maduras, por una parte y luego la salinidad con las hembras ovígeras. Respecto al fotoperiodo la relación no es directa, apreciándose correlaciones bajas, las cuales sin embargo nos indican en forma general las relaciones existentes, entre las variables ambientales y los ciclos de maduración y reproducción. Esto nos hace pensar en análisis más extensos y de un mayor número de localidades, incluso análisis en series de tiempo más amplios que un año. 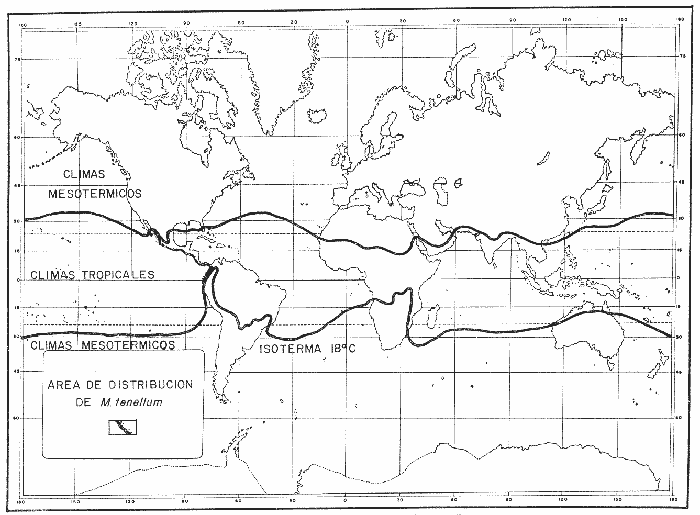 Fig. 11 Distribución Geográfica de Macrobrachium tenellum. El fotoperiodo no fue el factor más significativo, dado que muchas veces las respuestas al fotoperiodo son sutiles, pero en general son del tipo que permite poner en fase el ciclo de vida y sus actividades con las estaciones del año. La luz a través del fenómeno de la fotoperiodicidad es el factor responsable de mantener la sincronia fisiológica de los diferentes individuos entre sí, con los ciclos estacionales, de manera que los individuos alcancen su estado reproductor en la estación apropiada del año. En ocasiones el fotoperiodo es el detonante, que como indicador muy preciso tornará comandos sobre otros estímulos (Ej., la precipitación) que es un factor más notorio (Rabinovich, 1980). ConclusionesLa distribución geográfica del chacal M. tenellum se encuentra delimitada por la isoterma de los 18°C, que limita los climas de latitudes medias o mesotérmicas tanto boreales como australes de los climas sin invierno o tropicales. El ciclo de maduración va del mes de mayo a diciembre en Arenal y de mayo a febrero en Camalote, con modas en julio-agosto para ambas estaciones. El ciclo de reproducción va de junio a enero en Arenal y de mayo a febrero en Camalote, ambas con modas en septiembre-octubre. La respuesta de los ciclos de maduración del chacal van más en concordancia con las variables ambientales regionales que con las locales. Es mayor con la precipitación y dentro de ésta con los promedios de varios años que con la precipitación del año de estudio, lo que nos indica que responde a ciclos circanuales mediante mecanismos propios de la especie, en íntima concordancia con la ecología regional. Las diferencias en número o tamaño son dados por la ecología local (denso/dependencia o sea disponibilidad de alimento y espacio). AgradecimientosLos autores hacen patente su agradecimiento a las siguientes personas. Alfredo Martínez como coordinador del Programa Uso de la Zona Costera de Guerrero y Michoacán, bajo el patrocinio del cual fue realizado este proyecto. Sergio Cortés y Gilberto Díaz por su colaboración en el trabajo de campo, Héctor jardines por su ayuda en el proceso del material, Arturo Ocegada, Cecilia Quiroga, Armando Ortega y Craig B. Kensler por la revisión crítica del manuscrito. LITERATURABATSCHELET, E.,Introduction to Mathematics for Life Scientists.Springer-Verlag.Berlín1975. 495 p. EMLEN, J. M., Ecology: An Evolutionary approach. Addison-Wesley Pub. Co. 1977. 493 p. GARCÍA E., Pub. Esp. Inst. Geogr. Univ. Nal. Autón. México.Modificaciones al Sistema de Clasificación Climática de Koppen.1973.246 p. GRANADOS, B. A., Tesis profesional. Fac. Ciencias, Univ, Nal. Autón. México. Biología y aspectos poblacionales del langostino de río Macrobachium americanum (Bate, 1808) (Decápoda: Palaemonidae) en algunas áreas de los estados de Michoacán y Guerrero, México. 1980.110 p. GUZMÁN, A. M.,In: Informe Final. 3ra. Etapa Programa Uso de la Zona Costera de Michoacán y Guerrero, Convenio Comisión del Río Balsas, S.R.H., y Centro de Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Contrato de Estudios No. OC-E-03-75Biología, ecología y pesca del chacal Macrobachium tenellum en las lagunas costeras de Guerrero. Inf. Téc. 3. Centro Cienc. del Mar y Limnol. Univ. Nac. Autón. México. 149 p.1975. 759 p. GUZMÁN, A. M. y C. KENSLER Los langostinos del género Macrobrachium en México. II Simp. Asoc. Latinoam. Acuic.,México.1978. KREBS, J. R. y N. B. DAVIESBehavioural Ecology.Blackwell Scientific Pub.1978. 494 p. LACKEY, P. T. y W. A. HUBERTAnalysis of Exploited fish populationsVirginia Polythecnic and State University.Virginia1977.172 p. LANKFORD, R. L., Centro Cienc. del Mar y Limnol. Univ. Nac. Autón. de MéxicoInforme final de la segunda etapa de estudio sobre el uso de la zona costera de los estados de Michoacán y Guerrero. Subprograma de Geología.1975. 64 p. PIANKA, E. R.,Evolutionary Ecology. Harper and Row.Nueva York1974.356 p. RABINOVICH, J. E.,Introducción a la Ecología de Poblaciones animales. Editorial C.E.C.S.A.,México1980.313 p. SOKAL, R. R. y F. J. ROHLFBiometry. W. H. Freeman and Co. San Francisco1969. 776 p. TRUESDALE, F. M. y W. J. MERMILLIOD CrustaceanaThe river shrimp Macrobachium ohione (Smith) (Decapoda: Palaemonidae): its abundance, reproduction and growth in the Atchafalaya river basin of Louisiana, U.S.A.1979.61-73.36 (1)
|