|
ASPECTOS
ECOLÓGICOS DE LA CONTAMINACIÓN ORGÁNICA SOBRE EL
MACROBENTOS DE LAS CUENCAS DE SEDIMENTACIÓN EN LA BAHIA DE MARSELLA
(FRANCIA)
Trabajo recibido el 11 de septiembre de 1980 y aceptado para su
publicación el 4 de mayo de 1981.
VIVIANNE SOLÍS-WEISS
Universidad Nacional Autónoma de México, Instituto
de Ciencias del Mar y Limnología. Contribución 273 del Instituto
de Ciencias del Mar y Limnología, UNAM.
Se estudió
la macrofauna béntica de las cuencas de sedimentación de la
bahía y alre. dedores de Marsella. Se analizaron los grupos
faunísticos relacionando sus necesidades ecológicas conocidas con
las condiciones imperantes en estos medios. Se determinó que los tres
factores más importantes responsables de las variaciones observadas en
la composición de la fauna son: 1) la proporción creciente de
lodo, 2) el aporte e incorporación al sedimento de fibras
dePosidonia oceanica y 3) el aumento progresivo de la
cantidad de materia orgánica de origen humano proveniente de la Ciudad
de Marsella y sus puertos.
Las variaciones observadas en la
diversidad y abundancia de la macrofauna béntica de las cuencas de
sedimentación, permitió separar los efectos propios de cada
factor de variación y su importancia en la evolución de estos
ambientes. Se encontró que las fibras de P.
oceanicaincorporadas al sedimento, crean una equivalencia
edáfica con la grava fina, la cual es muy perceptible para la fauna. Se
indicaron también los efectos potencialmente destructivos de agentes
contaminantes difícilmente biodegradables que se adsorben a los
sedimentos finos y afectan de manera significativa la distribución
ecológica de la fauna. Finalmente se definen estos fondos no como
biocenosis diferenciadas, sino como una mezcla de varias biocenosis en
proporciones variables con características de inestabilidad y de
variación en el tiempo. A estas características se agregan, de
manera típica en estos ambientes, las perturbaciones más o menos
severas provacadas por los aportes directa o indirectamente ligados a la
contaminación.
The benthic macrofauna from: the sedimentation bassins of the Bay of
Marseille and its surroundings was studied. The faunistic groups were analized
correlating their known ecological needs with the dominant physical conditions
in these environments. The three factors that were found to be mainly
responsible for the changes observed in the composition of the fauna were: 1)
the increasing proportion of mud, 2) the enrichment and incorporation to the
sediment of the fibers ofPosidonia oceanica, 3) the
increasing amount of organic matter of human origin coming from the City of
Marseille and its harbor.
The variations observed in the
diversity and abundance of the bentic macrofauna of these sedimentation
bassins, allowed us to separate the effects of every variation factor and its
importance in the evolution of these environments. The P.
oceanica fibers incorporated to the sediment were found to create an
edaphic equivalence with fine gravel, this being very sensitive to the fauna.
The potentially destructive effects of polluting agents (poorly biodegradable)
adsorbed to fine sediments were indicated as well as their effects on the
fauna. Finally, these bassins were defined not as differentiated biocenoses but
as a mixture of different biocenoses in variable proportions, with instability
and time variation characteristics, adding to these the perturbations more or
less severe due to the contributions directly or indirectly tied to
pollution.
La ciudad portuaria de Marsella, cuya expansión ha sido muy intensa en los últimos años, ha venido a constituir un foco de contaminación importante para sus alrededores. En la Bahía de Marsella, la calidad de las aguas se ha visto seriamente afectada, dañando a la fauna marina que ahí habita. Esto es muy perceptible en el bentos, particularmente en los denominados "fondos blandos inestables" (Pérés y Picard, 1964). Los más lodosos de ellos a los que se ha dado en llamar comúnmente "cuencas de sedimentación", constituyen, por su posición geográfica o su estructura morfológica (hondonadas o depresiones) lugares privilegiados para acumulación de partículas de toda índole que llegan a sedimentarse ahí. En la Bahía de Marsella, objeto de este estudio, estas cuencas reciben cantidades importantes de materia orgánica de origen humano provenientes de desechos urbanos, y fibras semidescompuestas de la fanerógama marina Posidonia oceanica originarias de los ceibadales vecinos. Cuando se iniciaron los primeros estudios de la bahía (Pérès, 1954; Bellan, 1964) el aporte de fibras de P. oceanica, muy abundantes, era la principal característica fisionómica de estas cuencas y constituía la fuente principal de materia orgánica importada a estos fondos. Sin embargo, desde hace unos diez a quince años, el auge de la Ciudad de Marsella y sus puertos ha provocado aumento de los desechos del desagüe de Cortiou y degradación de las aguas portuarias. Además, en verano, para aminorar en algo.la contaminación en la playa del Prado, las aguas del Río Huveaune se desvían hacia el desagüe de Cortiou. Este descarga entonces, además de los desechos usuales a mayor concentración, agua dulce y sedimentos muy finos de origen fluviatil. Todo esto ha provocado la desaparición casi total de las extensas praderas de P. oceánica de esa región y por consiguiente, contribuido al desarrollo de las cuencas de sedimentación por dos causas: 1) por la fuerte producción en poco tiempo de fibras liberadas, debido a la destrucción de los ceibadales de P. oceánica y 2) por los aportes de materia orgánica y otros contaminantes producto de la actividad humana (contaminación doméstica, portuaria e industrial que se vierte al mar sin tratamiento previo). En la actualidad, estas cuencas a las que ya no se puede calificar de naturales, se están expandiendo con rapidez, debido al grave y constante aumento de la contaminación. Las biocenosis implicadas en las poblaciones de estos fondos de sedimentación son aún reconocibles, pero ya se pueden observar a menudo perturbaciones marcadas que dan lugar a estados que se pueden calificar de "Subcontaminación". La finalidad de este trabajo es la de tratar de analizar las poblaciones de las principales cuencas de sedimentación de la bahía y de los alrededores de Marsella y de observar sus particularidades en el espaci o para tratar de definir alguna sucesión entre las poblaciones típicas y las de la llamada "zona subnormal". Esto se hará tomando en cuenta el origen de las contaminaciones antes mencionadas y también, desde luego, el sentido preponderante de las corrientes marinas locales. ÁREA DE ESTUDIOLa zona de estudio se encuentra localizada dentro de la Bahía de Marsella (Francia). 43°18' lat. N 5°24' long. E con dos estaciones fuera de la bahía: 1) frente al desagüe de Cortiou (estación 6) y 2) una milla al este de dicho desagüe (estación 2). Son en total 10 estaciones que abarcan todas las cuencas de sedimentación importantes de los alrededores de Marsella. La localización de las estaciones se puede observar en la figura 1 y las características de éstas en la tabla 1. 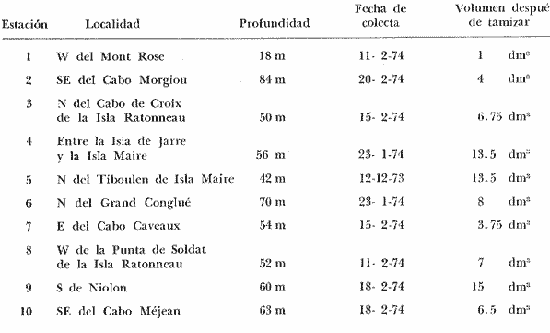 TABLA 1 CARACTERISTICAS DE LAS ESTACIONES DE MUESTREO 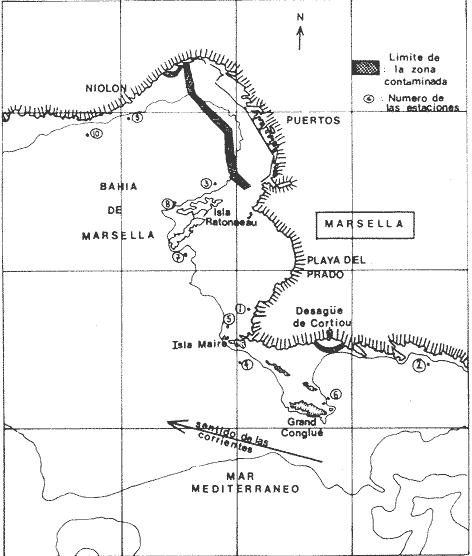 Fig. 1. Mapa de localización de las estaciones de muestreo en la Bahía de Marsella. MATERIAL Y MÉTODOSLa metodología empleada en este trabajo es la que ha sido definida y perfeccionada por J. Picard (1965). Resumiendo, se trata de recolectar con draga Charcot modificada un "volumen mínimo homogéneo" de 50 dm³ el cual se tamiza directamente en tamiz de malla de 0.8 mm para conservar únicamente a los especímenes pertenecientes a la macrofauna (diámetro > 1 mm). Las muestras tamizadas se fijan con formol al 10%. Posteriormente, las especies que poseen alguna fracción calcárea se fijan con alcohol a 70%. Se utilizó el mismo concepto de biocenosis y la misma nomenclatura empleada por Pérès y J. Picard (1964). La biocenosis implicadas en este estudio (por las afinidades con la fauna de las cuencas de sedimentación) son las siguientes que se enlistan a continuación con las abreviaciones con las que se utilizarán en los resultados y en las listas de especies. AF = Biocenosis de Arenas linas DC = Biocenosis de Fondos Detríticos Costeros DCT = Biocenosis de Fondos Detríticos Costeros Típicos LD = Biocenosis de Lodos Detríticos CP = Biocenosis de los ceibadales dePosidonia oceanica AFNS = Biocenosis de Arenas Finas en Niveles Someros AGCF = Biocenosis de Arenas Gruesas y Grava Fina bajo influencia de Corrientes de Fondo LP = Biocenosis de Lodos Profundos LTC = Biocenosis de Lodos Terrígenos Costeros. Se calculó la dominancia de las especies presentes en los muestreos y sus afinidades por las biocenosis AF, DC, LD y LTC según el método de J. Picard (1965) para medios de transición. Se tomaron en cuenta, además, las modificaciones aportadas al conjunto de especies características exclusivas de los LTC por C. Picard (1972) y la corrección del valor de la suma de dominancias medias de las especies características exclusivas de esta biocenosis que pasa así de 65 a 56. La cantidad de fibras presentes en los diferentes muestreos se calculó de manera indirecta. En efecto, no se la puede deducir directamente del volumen del sedimento después de tamizar, puesto que éste se compone, además, de grava y concherío más o menos grueso. Se comparó pues el volumen total de sedimento de cada muestra con el volumen que ocupan en esa muestra las fibras y se llegó a la seriación progresiva indicada en la figura 2. De igual manera, la comparación de estos datos (cantidad de fibras y cantidad de grava, cuya suma es igual al volumen de muestra después de tamizar) con los datos granulométricos indicando la cantidad del limo presente, permite deducir la cantidad de arena presente. Los resultados de estos cálculos procentuales se pueden ver en la tabla 2 y en la figura 3. 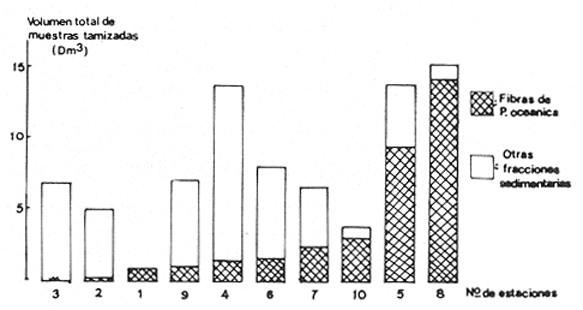 Fig. 2. Volumen ocupado por las fibras en las diez estaciones. 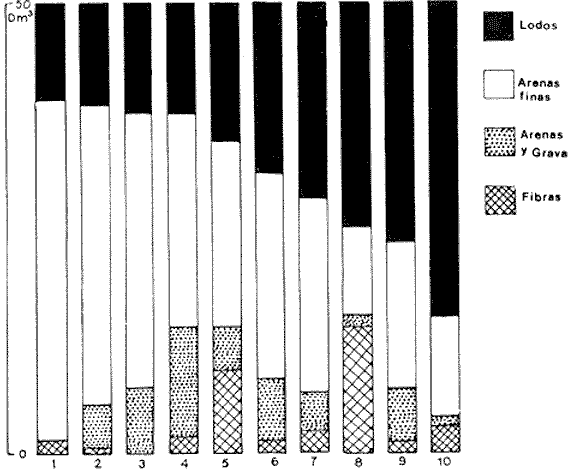 Fig. 3. Distribución porcentual de las diferentes fracciones sedimentarias en las estaciones de estudio. 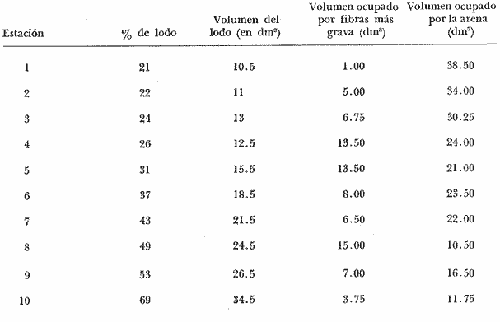 TABLA 2 VOLUMEN OCUPADO POR LAS DIFERENTES FRACCIONES SEDIMENTARIAS EN LAS DIEZ ESTACIONES En este análisis de la macrofauna de las 10 estaciones no se tomó en cuenta a la fauna Sésil cuya presencia en estos fondos no se encontraba ligada al sedimento sino a fenómenos de transporte y sedimentación propios de estos ambientes y que se encontraban ahí más o menos arrastrados como desechos sin posibilidad de sobrevivencia. Se trata de algunos serpúlidos grandes y de algunos ejemplares de esponjas. Sin embargo, sí se tomó en cuenta a la fauna de substrato sólido que aún en estos fondos blandos vive relativamente bien gracias a microsubstratos duros constituídos por fragmentos de conchas o rocas más o menos gruesos depositados sobre el fondo lodoso. La separación de la fauna en el laboratorio y su identificación posterior permitió ordenarlos en categorías ecológicas. Se conservaron las categorías utilizadas por J. Picard (1965) con los mismos criterios, agregándose además a la lista de la fauna "asociada a la grava" la mayoría de los individuos clasificados como indicadores de AGCF-DCT" por M. Ledoyer (1967). Además, en lo que se refiere a especies "Mixtícolas," (asociadas a sedimentos mixtos) se incluyó en esta categoría a los Anfípodos y Poliquetos definidos por Bellan (1964) y Bellan-Santini y Ledoyer (1973) como "detritícolas" con el fin de respetar el criterio de base de J. Picard, es decir la clasificación basada en el tamaño del grano sedimentológico y no en su origen, ya que una fracción de grava compuesta por fragmentos de conchas y otra compuesta por rocas muy fragmentadas pueden presentar una fauna idéntica. Se verá más adelante que, cuando por alguna razón existe en el medio algún agente que modifica la textura del sedimento (por ejemplo las fibras de Posidonia oceánica), las poblaciones que se instalan ahí lo hacen en función de las condiciones "equivalentes" creadas por el agente modificador sin que la composición u origen de éste jueguen ningún papel: sólo importa la textura resultante del sedimento. Se conservaron, además de las categorías definidas por J. Picard, tres categorías definidas por Ledoyer (1967) "indicadores DC-DCT", "indicadores de tendencias coralígenas" entre los cuales se incluye igualmente a los "indicadores de substrato duro" y finalmente a las especies "exclusivas de las praderas dePosidonia". Estas categorías de animales figuran aquí únicamente de manera accidental ya que la nomenclatura indica claramente que se trata de individuos de substrato duro o de ceibadales activos de Posidonia, pero es interesante observar la relativa variedad de estas especies (19 en total) que a pesar de Gas condiciones tan diferentes de las de su medio de origen llegan a sobrevivir en las cuencas de sedimentación. Finalmente se entiende aquí por "Especies de amplia distribución ecológica" a especies cuya repartición se extiende a todo tipo de substratos blandos excepto en el caso del anfípodo Phtisica marina que se encuentra comúnmente tanto en fondos duros como blandos. Hay que agregar, a este respecto, que la ecología de los Crustáceos y en particular de los Anfípodos se conoce bastante mal y que hasta que estudios posteriores determinen sus verdaderas exigencias, en especial en fondos blandos, se tiende a clasificar en categorías que parecen, la priori demasiado amplias o demasiado estrechas según los autores. RESULTADOS Y DISCUSIÓN
CARACTERÍSTICAS DEL SEDIMENTOSe numeraron las estaciones en función de la cantidad progresiva de lodo que contenía el sedimento tal y como lo indican los estudios granulométricos del sedimento que se resumen en la tabla 3. 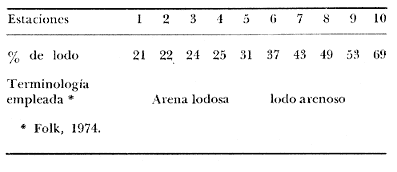 TABLA 3 PORCENTAJE DE LODO ( d 50 my) EN EL SEDIMENTO DE LAS ESTACIONES DE ESTUDIO Por consiguiente, habrá que considerar durante el desarrollo de la discusión, que cuando se habla de las estaciones, mientras su número sea más elevado su sedimento posee mayor cantidad de sedimentos finos. GRUPOS FAUNÍSTICOSSe identificaron aproximadamente 10,000 individuos de sección mayor a 1 mm pertenecientes a 330 especies de (los siguientes grupos taxonómicos: (Celenterados, Turbelarios, Nemertinos, Anélidos Poliquetos, Sipuncúlidos Moluscos, Equinodermos y Cordados. Se anexa lista completa de las especies encontradas, así como de la categoría ecológica a la que pertenecen. En el grupo de los Poliquetos, que domina en estos fondos lodosos, se encontraron varias especies que es de interés mencionar, ya que es la primera vez que se registran en el Mediterráneo y eso amplía su distribución conocida son: Ninoë armoricana, Glémarec, 1968 (Anélido Poliqueto): Descrita y observada anteriormente sólo en el Atlántico. Caulleriella cf. gracilis, Hartman, 1961 (Anélido Poliqueto): Como habitante de las costas de California. Los especímenes recolectados aquí son idénticos a los que describe Hartman, excepto por la forma de los ganchos parapodiales posteriores que en la descripción de Hartman presentan dentículos y en los especímenes del Mediterráneo son lisos. La especie se encuentra actualmente en estudio para determinar si esta variación representa alguna forma "vicaria" deC. gracilis propia al Mediterráneo o si se encuentran diferencias más profundas que permitan separarlas en dos subespecies o especies distintas. Eupanthalis sp. (Anélido Poliqueto): Especie actualmente en estudio, ya que los especímenes recolectados (en buen estado) hacen suponer que se trata de una especie nueva para la ciencia. Pista cf. cristata (Anélido Poliqueto): Se recolectaron individuos del género Pista afines a la especie P. cristata pero con diferencias morfológicas bastante marcadas como para pensar que se trata también de una especie nueva para la ciencia; la identificación final de estos individuos se encuentra actualmente en estudio. También entre los Sipuncúlidos se encontraron individuos del géneroOnchnesoma sp. que se recolectan por primera vez en el Mediterráneo. Su tamaño es muy pequeño (ø < 1 mm) aunque se les recolectó en estado maduro. A pesar de que las 10 estaciones de colecta constituían ambientes muy similares (cuencas de sedimentación), la gran diversidad encontrada en la distribución de las especies recolectadas indicaba claramente que dentro de las mismas biocenosis ciertos factores de variación del medio eran responsables de tal distribución. Se pudo determinar que los factores susceptibles de imponer modificaciones notables en la naturaleza de las poblaciones de las cuencas de sedimentación son los siguientes: A) La proporción de lodo (fracciones sedimentarias muy finas ø < 50 µ) existente en el substrato (ver tabla 3 que indica la seriación de las estaciones del 1 al 10). B) La proporción de fibras dePosidonia oceánica que, como se indicó más arriba, invaden progresivamente estos fondos. C) La cantidad de materia orgánica de origen humano. Estos tres factores son comunes a las 10 estaciones, en diferentes proporciones, de tal manera que, considerando las particularidades de cada estación tanto por el examen de (la parte componente del sedimento (grava, restos de conchas, lodo, fibras) como por un examen faunístico se pueden separar los efectos propios de cada uno. Para esto, se analizaron los efectos de dichos factores de variación sobre la fauna: A) Proporción de lodo En la tabla 3 se puede ver el porcentaje preciso de la cantidad de lodo presente en cada estación. Teóricamente, al progresar de la estación 1 a la 10 se debería encontrar cada vez mayor proporción de especies afines a las biocenosis típicas del lodo, como son las biocenosis LD (Lodos Detríticos) y LTC (Lodos Terrígenos Costeros). Sin embargo, no sucede así. Al observar las variaciones existentes en la dominancia media de las especies características de las biocenosis típicas (J. Picard, 1965) se hace necesario introducir factores de corrección a los porcentajes observados aquí para las biocenosis AF (Arenas Finas), DC (Detrítico Costero), LD (Lodos Detríticos) y LTC (Lodos Terrígenos Costeros) que compiten en estos fondos. 1) Afinidades relativas de la fauna para las biocenosis AF, DC, LD y LTC. Los resultados de cálculos de afinidades de las diferentes especies, se pueden apreciar en la tabla 4. 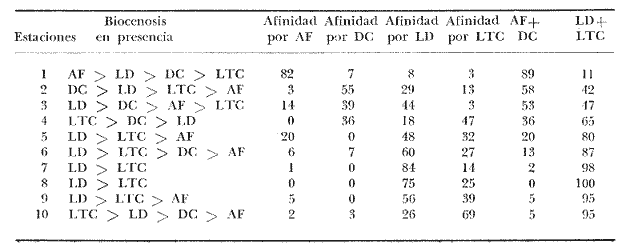 TABLA 4 AFINIDADES RESPECTIVAS DE LAS POBLACIONES DE LAS 10 ESTACIONES DE MUESTREO POR LAS BIOCENOSIS AF, DC, LD Y LTC Se observa que las afinidades de cada estación por las diferentes biocenosis no sigue ninguna secuencia determinada ni se rige por la proporción real de lodo. En cambio, al sumar las afinidades encontradas en cada estación por las biocenosis no lodosas (AF y DC) y por las biocenosis lodosas (LD y LTC) se observa una progresión lógica de afinidad creciente con creciente proporción de lodo (Fig. 4). Sin embargo, a partir de la estación 8, a pesar de que la proporción de lodo aumenta de manera significativa (pasando de 49% a 53% y a 69%) la afinidad por las biocenosis lodosas disminuye progresivamente. Esta "obstaculización" al desarrollo de la fauna típica de las biocenosis lodosas no se puede atribuir a las biocenosis LD o LTC transgrediendo a las 2 estaciones, ya que la estación 9 tiene mucho más afinidad con la biocenosis LD y la estación 10 más afinidad por los LTC (Tabla 4). De igual manera, la dominancia de especies características exclusivas de las biocenosis LD y LTC en los muestreos es sorprendentemente menor de la que se podría esperar: así, en la estación 9 sólo 3.40% de la población está representada por especies exclusivas de la biocenosis LD (Solís Wolfowitz, 1975), aún cuando su afinidad por esta biocenosis es de 56% y según J. Picard (1965) el porcentaje de especies características debería estar cerca de 7%. En la estación 10, el porcentaje de especímenes característicos exclusivos de los LTC llega apenas a 7.33% siendo que la afinidad de dicha estación por el lodo, al ser de 69 % hace suponer un porcentaje cercano a 2017, (J. Picard, 1965). Esto muestra que existe ahí algún agente perturbador claramente perceptible al pasar de 50% la proporción de lodo en el sedimento. Se cree que se puede tratar de un aspecto especial de la contaminación ligado a los sedimentos finos como sería adsorción en el limo de contaminantes químicos, como por ejemplo, detergentes. El equipo de Geoquímica de la Estación Marina de Endoume está actualmente estudiando este problema, ya que desde 1974 se ha podido notar la alarmante proporción de detergentes vertidos en el Golfo de Marsella (Arnoux y Chamley, 1974). 2) Comparación de especies características de sedimentos finos y gruesos. En la tabla 5 se pueden ver los porcentajes registrados en cada estación de especies ligadas al lodo, es decir las especies que se conocen como características exclusivas de los lodos detríticos (LD) y de los lodos terrígenos costeros (LTC), así como las especies definidas como limícolas estrictas y de las limcolas tolerantes. Al observar esta tabla, se puede notar que las variaciones de las 10 estaciones en los porcentajes registrados de estas especies limícolassensu lato son muy amplias (de 6.77% a 66.15%). Entre las estaciones 9 y 10 sólo hay 16 % de diferencia en la proporción de lodo y sin embargo hay en la estación 10 más del doble (en porcentaje) de especies exclusivas de los lodos terrígenos costeros y casi el doble de limícolas estrictas mientras que las limícolas tolerantes varían apenas y en cuanto a las características exclusivas de los LD se encuentran bastante disminuídas. 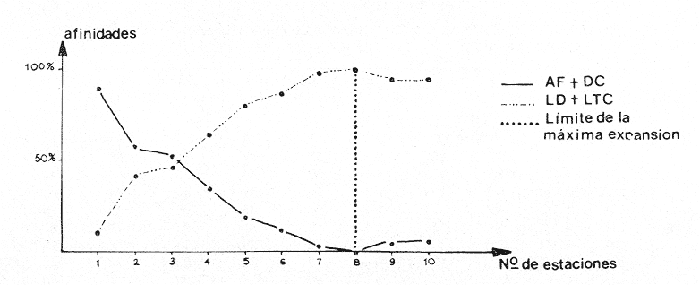 Fig. 4. Representación gráfica de las afinidades de las diez estaciones por las biocenosis más lodosas (LD+LTC) y las biocenosis menos lodosas (AF+DC). 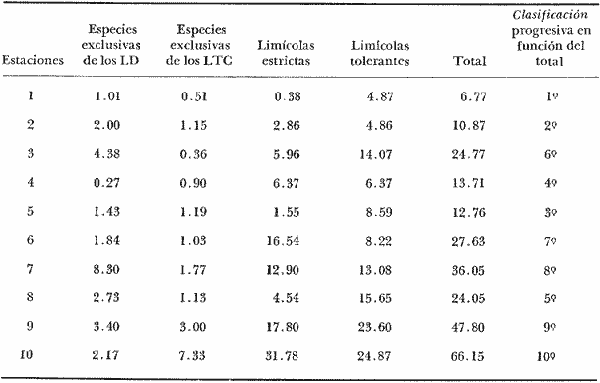 TABLA 5 ESPECIES CARACTERÍSTICAS DE FONDOS LODOSOS (EN PORCENTAJE POR ESTACIÓN) La estación 8 posee pocas especies limícolas en comparación con su proporción granulométrica de lodo (49%) y se ve que su posición en este sentido es equivalente a la de la estación 3 que posee una proporción de lodo mucho menor (24%). Estas aparentes anomalías indican una vez más la acción de otro agente perturbador que se discutirá más adelante. A continuación se muestra en la tabla 6 como se presentan las especies que necesitan de fracciones gruesas en el sedimento para desarrollarse (son las características exclusivas del DC, las especies que viven en la grava y las Mixtícolas, es decir que necesitan sedimentos mezclados para su desarrollo. Al observar esta tabla se puede ver que en la estación 2, casi 40% del total de las especies son especies características de la grava, porcentaje muy superior a cualquier estación. La estación 5, con 0% de afinidad para el DC (Tabla 6) presenta el mayor porcentaje de "especies ligadas a la grava" (2.15%) y un buen porcentaje de Mixtícolas (9.42%). Las estaciones 1 y 9 tan diferentes en su afinidad por las diferentes biocenosis y por su proporción de lodo (21% y 53%) presentan porcentajes iguales de Mixtícolas (9.13% y 9.2%) y número igual de dichas especies (en total 7). Así pues, la observación de las tablas 5 y 6 y los comentarios arriba expuestos nos llevan a considerar el segundo factor de variación (cantidad de fibras de Posidonia oceanica en el sedimento) para explicar las aparentes anomalías en la distribución de la fauna de las diez estaciones. Proporción de fibras de Posidonia oceanicaen el sedimento Ya se indicó más arriba el origen de las fibras dePosidonia oceanica que llegan a sedimentar en proporción cada vez mayor a las cuencas de sedimentación, ya que debido a la contaminación proveniente de Marsella, los ceíbadales han desaparecido casi totalmente. Estas fibras, al sedimentar en esas cuencas ya no sufren cambios ni transporte ulterior, excepto en el caso de la estación 1, cuyo sedimento se ve expuesto, debido a su escasa profundidad (tabla 1) a la acción de las olas de fondo durante las tormentas. La "malla" que van formando estas fibras dentro del sedimento se percibe manualmente al intentar fragmentar un "terrón" de sedimento originario de esas cuencas. 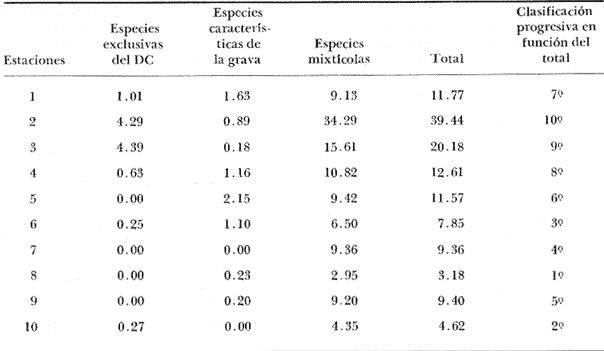 TABLA 6. ESPECIES CARACTERÍSTICAS DE LA GRAVA (EN PORCENTAJE POR ESTACIÓN) Los efectos esperados de estas fibras en la fauna que puebla estos fondos son teóricamente los siguientes: a) Dificultar la circulación de los organismos que se mueven en el sedimento. b) Favorecer a las especies excavadoras que necesitan de sedimento más compacto. c) Favorecer a las especies que se mueven sobre el sedimento. d) Enriquecer el sedimento en materia orgánica vegetal (ésto tiene importancia solo a nivel de microorganismos, ya que se ha visto que el macrobentos usualmente no aprovecha para su alimentación estas fibras). Las fibras, por lo tanto deben ser consideradas principalmente como elemento modificador edáfico más o menos inerte, ya que además no se degradan de manera notable al menos durante varios años. Al observar la figura 3 que muestra el volumen aproximado de fibras y grava presentes en cada estación, se puede ver que: a) Las estaciones 5 y 8 poseen una cantidad mucho mayor de fibras que las demás. b) Las estaciones 2 y 3 prácticamente no poseen fibras. c) La estación 1 sólo tiene fibras y nada de grava. En la figura 3 se encuentran representadas las proporciones de las diferentes fracciones sedimentarias medidas en las diez estaciones (lodo, arena, fibras y grava). Al comparar estos datos edáficos de la figura 3 con las tablas faunísticas (Tablas 5 y 6) se, puede ver como las fibras de P. oceanica constituyen un elemento de compensación comparable al efecto que darían arenas gruesas o grava fina. Esto se nota claramente al comparar las estaciones 4 y 5: en efecto las 2 estaciones se parecen desde el punto de vista edáfico (cantidades de lodo y arena iguales y mismo volumen rechazado por el tamiz). La gran diferencia entre estas dos estaciones reside en la cantidad de fibras deP. oceanica, mucho mayor en la estación 5 que en la 4. Las poblaciones bentónicas de sustratos blandos, según se sabe, reaccionan ante la textura del sedimento en el que se instalan. Así pues, el hecho que en la estación 5 las fibras tomen el lugar de la grava de la estación 4 se debe reflejar en la fauna característica de la grava. Las especies exclusivas del DC sí fueron afectadas por la falta de grava ya que desaparecieron totalmente en la estación 5 (en la estación 4 se encontraron solo 5 especies que representación tan 0.63% de la población). Esto es lógico, ya que su clasificación de "exclusivas" de una biocenosis hace que sean las primeras en sufrir por cambios edáficos en su habitat (J. Picard, 1965). Sin embargo, las especies características de la grava y las mixtícolas se ven favorecidas por este arreglo del sedimento, así que al agregar los porcentajes de especies representadas ahí de estas dos categorías ecológicas (sin las exclusivas DC), resulta que los porcentajes encontrados son prácticamente iguales (11.57% y 11.98%). Ahora, al observar qué tipo de especies características de la grava son las que se encuentran en la estación 5, se ve que son 3 especies de Anfípodos:Orchomene humilis Urothoë elegans yMetaphoxus pectinatus a las cuales Ledoyer (1968) atribuye una preferencia por arenas gruesas. En la categoría de Mixtícolas, se puede observar que la estación 5 presenta la mayor dominancia de las 10 estaciones de la especie Ampelisca diadema (2.26%) y de Aporrhais pespelicani(3.10%). Estas especies son, en general, bastante móviles y se desplazan sobre y nodentro del sedimento A. pespelicani al que no se puede considerar muy móvil, se ve favorecido, en este caso, por su etología alimenticia, ya que se coloca sobre su parte dorsal para alimentarse; por lo tanto, mayor soporte por parte del sedimento lo ayuda. Las especies "excavadoras" como las del géneroUrothoë también se ven favorecidas. En la estación 1, que no posee prácticamente nada de grava y solo cierta cantidad de fibras no muy elevada (ver Fig. 3), se puede notar que se encuentran dos especies de Crustáceos exclusivas del DC:Conilera cylindracea(0.13%) y Anapagurus leavis (0.88%) y entre las especies características de la grava 5 especies de CrustáceosEocuma sarsi, Leucothoë oboa, Urothoë elegans, Metaphoxus pectinatus y Cerapopsis longipes. En la categoría de Mixtícolas, 4 especies de Crustáceos:Eurydice, truncata, Macropipus parvulus, Ampelisca spinipes y Ampelisca sp., 2 de Poliquetos: Eteone longa y Nematonereis unicornisy 1 de Moluscos Dentalium inaequicostatum(la mayor dominancia: 6% de la población). La presencia de las especies exclusivas del DC es explicable por la poca cantidad de lodo presente en esa estación agregada a la equivalencia edafica de mayor soporte, que crean las fibras con las arenas gruesas o 1 grava fina. La gran mayoría de estas especies son móvilessobre el sedimento. Para terminar este análisis global, se observan los resultados de la estación 8 (que tiene la mayor cantidad de fibras de P. oceanica): La fauna de especies características de la grava de esta estación es muy pobre (1 individuo, de Venus ovata que más bien parece accidental aquí); es la estación con el menor porcentaje de mixtícolas representadas por Thyone inermis, Nucula nucleus, Syllis cornuta., Stylarioides eruca: Hyalinoecia tubicola y Dentalium inaequicostatum sin ningún Crustáceo de esta categoría. Aparentemente, la equivalencia edáfica creada por las fibras de Posidonia oceanicacon la grava fina o las arenas gruesas se refleja de manera clara en la fauna característica de la grava mientras la proporción de lodo no sea demasiado grande. Estas especies, sensibles al tamaño del sedimento y a las características que de ello, se derivan (porosidad, soporte, etc.), se ven eliminadas cuando la proporción de lodo no se compensa con las otras fracciones sedimentadas. La observación de la tabla 2 y de la tabla 6 muestran que cuando la proporción de lodo llega a 50%, o valores superiores, las especies ligadas a la grava desaparecen, aún cuando haya grava (estaciones 7 y 9) o elementos compensadores (fibras de la estación 8). Las especies mixtícolas también presentan esta tendencia, en las estaciones más lodosas sobre todo si se observan los grupos taxonómicos, ya que los primeros en ser eliminados son los Crustáceos y en menor grado los Moluscos; los Poliquetos son los que soportan mejor las condiciones de aumento de lodo. Veamos ahora como la presencia de materia orgánica, tercer factor de variación considerado en estos fondos se combina con los dos anteriores. C) Presencia de materia orgánica de origen humano Se trata de la materia orgánica originaria de los puertos y desagües de la ciudad, materia que se va a sedimentar preferentemente a estas cuencas. Para separar los efectos de la cantidad de materia orgánica de los de las fibras que también llegan a degradarse ahí, se ordenaron a las estaciones en 2 grupos: 1) "estaciones presumiblemente más contaminadas" y 2) "estaciones presumiblemente menos contaminadas", en base a su posición geográfica (Fig. 1) y teniendo en cuenta que las fuentes de materia orgánica más importante se encuentran en los puertos y en el desagüe de Cortiou, considerándose también el sentido de las corrientes y vientos locales. A la categoría 1) pertenecen las estaciones 6, 4, 3 y 9; a la categoría 2) pertenecen las estaciones 2, 5, 8 y 10; las estaciones 1 y 7 no se pueden definir "a priori" dentro de ninguna de estas dos categorías. Al analizar la fauna de estas estaciones se podrá ver si hay especies en el grupo de estaciones más contaminadas que varían en abundancia o con respecto a otras al pasar al grupo de las estaciones menos contaminadas. Se tienen pues tres seriaciones de las 10 estaciones sin relación con la fauna que son: a) Granulométrica (la más exacta). b) Cantidad creciente de fibras de Posidonia oceanica en el sedimento. c) Cantidad creciente de materia orgánica. Al comparar estos datos con las listas faunísticas (tablas 5 y 6) se podrá ver qué organismos se encuentran favorecidos u obstaculizados por los diferentes factores de variación o su combinación. Observemos primero el comportamiento de las especies limícolas estrictas. Al ser éstas, por definición, muy sensibles a la cantidad de lodo van a indicar los efectos de los otros factores si su desarrollo no sigue una progresión con la cantidad creciente de lodo. 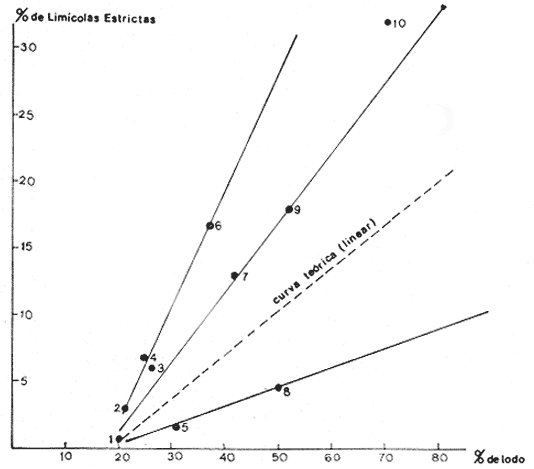 Fig. 5. Variación la dominancia de las poblaciones en función especies limícolas estrictas de las diez estaciones de la cantidad creciente de lodo. En la figura 5 se puede ver la variación del conjunto de las limícolas estrictas en función de la cantidad progresiva de sedimentos finos. Se puede observar de inmediato que, comparado con una curva teórica de desarrollo proporcional (y = ax + b), las estaciones 5 y 8 parecen estar "inhibidas" en su desarrollo con respecto a las demás, cuyo desarrollo se ve más acentuado que el de la curva teórica proporcional haciendo suponer un efecto sinergético con algún otro factor. El factor de inhibición aparente en las estaciones 5 y 8 se relacionó al comparar las diferentes seriaciones establecidas, con la cantidad de fibras presentes, ya que estas estaciones tienen la mayor cantidad de fibras de todas. Además, se deduce que tal efecto de inhibición a la expansión de especies limícolas estrictas es proporcional a la cantidad de fibras presentes, ya que es más acentuado en la estación 8 que en la estación 5 (esto se ve al trazar la normal a la curva teórica de desarrollo desde los puntos 5 y 8). En estos dos casos, al examinar con más detalle los grupos faunísticos de la categoría limícolas estrictas, se ve que las especies más afectadas son los Poliquetos Chaetazone setosa y Asychis gotoi y el MoluscoThyasira flexuosa. NOTA: Aún en el caso en que en la naturaleza el desarrollo de las especies limícolas estrictas variara de manera no linear a partir de cierto punto, las conclusiones generales que se deducen de aquí no varían. Esto parece confirmar la hipótesis según la cual las fibras presentes en el sedimento dificultan la progresión de los organismos móviles EN el sedimento como los Poliquetos. El caso de T. flexuosa, aparentemente más condicionado a otros factores se analiza en Solís Wolfowitz (1982a). Exceptuando a las estaciones 5, 8 y 1 se ve que todas las demás se ven favorecidas por algún factor que se une de manera sinergética a la cantidad de lodo presente. Ya que se construyó la figura 5 sobre la base de la cantidad de lodo y que se analizó el efecto del factor fibras de Posidonia oceanica en el párrafo precedente, se puede deducir que la cantidad de materia orgánica es la que favorece a las poblaciones de limícolas estrictas. Al trazar la normal a la curva teórica desde los puntos correspondientes a las estaciones, se puede ver qué estaciones se vieron más favorecidas por este factor: la estación 6 es la que se encuentra más favorecida seguida de las estaciones 9, 7, 4, 3 y 2. Se vio más arriba que las estaciones 6, 9, 4 y 3 pertenecían al grupo de las estaciones más ricas en materia orgánica de origen humano por su posición geográfica cercana a las fuentes de ésta. Las estaciones 7 y 1 quedaban sin significancia precisa para esta seriación. La estación 2 presenta un caso interesante ya que su posición parece favorecida en comparación con la cantidad de materia orgánica que posee, en principio escasa, por su posición geográfica (Fig. 1). Sin embargo, todo se aclara al observar con detalle su población de limícolas estrictas: el porcentaje total de limícolas estrictas se debe a la presencia del Poliqueto Chaetozone setosa que constituye 2% de la población de dicha estación. Aunque este porcentaje parezca elevado, se da uno cuenta al observar las estaciones 6 y 4 (más cercanas a zonas contaminadas) que los porcentajes que alcanza ahí C. setosa son mucho más elevados. Por ejemplo, la estación 6 con menos del doble de cantidad de lodo posee 4 veces más individuos de C. setosa que la estación 2. Así pues, la dominancia de C. setosa que todavía se observa en la estación 2 es más bien un efecto atenuado de lo que sucede en las estaciones 4 y sobre todo 6. La estación 10 también parece sobresalir de manera inusitada, ya que parece la estación más favorecida, en lo que respecta al desarrollo de limícolas estrictas de todas las estaciones. Sin embargo, la cantidad de materia orgánica que teóricamente posee es menor que la de la estación 9, por su posición (Fig. 1). Para explicar esto, se propone tomar en cuenta la NOTA de la página 30 que contempla la posibilidad de que el desarrollo teórico de las poblaciones de limícolas estrictas no siga una línea recta, sino una sigmoide como sucede casi siempre con todo tipo de desarrollo biológico natural (curvas de Michaelis-Menten). La sigmoide pasaría más cerca de la estación 10 y regularizaría su situación. Como no se conoce hasta ahora la cantidad óptima de lodo que necesitan las especies limícolas estrictas, no es posible deducir en qué momento se modifica la curva de desarrollo lineal a sigmoide, sobre todo si no se sabe qué efectos de competencia inter- e intraespecífico se pueden venir a sobreponer. Por esta razón se tomó como base de referencia en la figura 5 el desarrollo lineal. Hay que recordar para finalizar, que la figura 5 se hizo en base a la respuesta de las especies limícolas estrictas en conjunto y que representa pues las influencias inhibidoras o benéficas de los tres factores de variación para elconjunto de las poblaciones pertenecientes a esta categoría en las estaciones. Efecto de la materia orgánica y de las fibras en las poblacionesDespués de analizar las perturbaciones ocasionadas por estos factores en la fauna bentónica analizada a nivel de categorías ecológicas (limícolas, mixtícolas, etcétera) se pueden ver los efectos que se observaron a nivel de grupos taxonómicos. La tabla 7 muestra las diferentes categorías taxonómicas con sus abundancias y dominancias respectivas en cada estación. 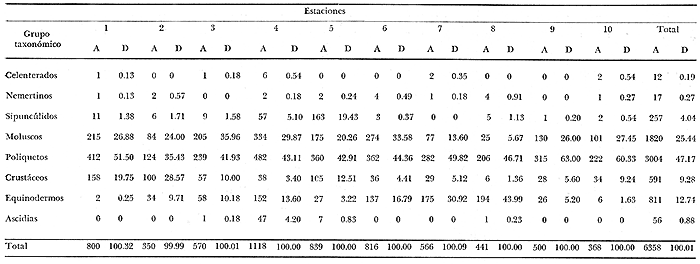 TABLA 7 ABUNDANCIAS (A) Y DOMINANCIAS (D) DE LOS GRANDES GRUPOS TAXONÓMICOS ESTUDIADOS EN LAS DIEZ ESTACIONES Se puede ver aquí que existe gran variación no solo en la abundancia de la fauna presente que varía de 350 a 1,118 individuos sino también en las dominancias relativas de cada grupo. La fauna de estos fondos se ve, en general, favorecida por la cantidad de materia orgánica en el sedimento, ya que al alejarse de las fuentes de contaminación humana se ve que la abundancia de organismos baja. Se tabularon únicamente los grupos cuya abundancia es significativa para el análisis de estos efectos dejándose fuera a los Celenterados Nemertinos, Ascidias y Sipuncúlidos. El examen de la tabla 7 y las consideraciones generales de los efectos de la materia orgánica cantidad de lodo y de fibras de Posidonia oceanica expuestas más arriba, nos llevan a la conclusión que los Moluscos se ven, en general, favorecidos por la cantidad de materia orgánica. En efecto, su abundancia es mayor, significativamente, en las estaciones más contaminadas y baja de manera perceptible en los grupos de estaciones menos contaminadas. De igual manera los Poliquetos y los Equinodermos son grupos que se ven favorecidos por la cantidad de materia orgánica presente en el sedimento. En cambio, con los Crustáceos, sucede lo contrario como se puede observar también en la tabla 7. ConclusionesEl análisis faunístico de las poblaciones de las cuencas de sedimentación de la Bahía de Marsella permitió demostrar que estos fondos no pueden ser considerados como entidades biocenóticas caracterizadas sino como una mezcla en proporciones variables de varias biocenosis con ciertas características de inestabilidad y variación en el tiempo, debidas precisamente al carácter de estos ambientes. Se definieron los principales factores de variación y evolución de estas cuencas: la proporción creciente de lodo, el aporte de fibras de Posidonia oceanica, el aumento de la cantidad de materia orgánica de origen humano y la intervención de elementos contaminantes difícilmente biodegradables, que pueden ser adsorbidos por los sedimentos finos. Se pudo observar cómo las fibras de P. oceanica incorporadas al sedimento crean una equivalencia edáfica con sedimentos más gruesos. Esto se puso en evidencia al analizar la respuesta de la fauna pobladora de estas cuencas a la mayor o menor cantidad de fibras del sedimento. La gran heterogeneidad de las poblaciones de las cuencas de sedimentación es muy notable y parece estar ligada directamente a la cantidad de materia orgánica que va en progresión de aumento paralelo con el desarrollo constante de la Ciudad de Marsella y sus puertos. Este estudio constituye una etapa más en el conocimiento del bentos de la región y su respuesta selectiva a un problema de gran actualidad: el aumento inquietante de la, contaminación en los fondos costeros. AgradecimientosEl autor desea expresar su agradecimiento a J. Picard por haber dirigido esta investigación y dado su apoyo y valiosos consejos, a J. M. Pérès, director de la Station Marine d'Endoume por haber dado todas las facilidades para realizar en dicha institución este proyecto, a M. Ledoyer, D. Bellan, D. Bellan-Santini, C. Picard, P. Vasseur, R. El Kaim y F. Picard por su ayuda en la revisión del trabajo y en los problemas taxonómicos encontrados. Al Consejo Nacional de Ciencia y Tecnología (CONACyT) cuyo financiamiento fue fundamental para la realización de este proyecto. LITERATURAARNOUX, A. y CHAMLEY, H., Minéraux des argiles et détergents des eaux interstitielles dans les sédiments superficiels du Golfe du Lion. (série D) C. R. Acad. Sc.Paris1974.999-1002.278 BELLANI-SANTINI, D. y M. LEDOYERThéthysInventaire des Amphipodes Gammariens récoltés dans la région de Marseille.1973.899-934.4 (4) BELLAN, G., Rec. Trav. St. Mar. EndoumeContribution á l'etude systématique bionomique et écologique des Annélides Polychètes de la Méditerrannée.1964.49-371.31 FAUVEL, P., Faune de France (5) Polychètes errantes. Le Chevalier Ed.París1923. 488 p. ---, Faune de France (16) Polychètes sédentaire. Le Chevalier Ed.París1927.494 p. FOLK, R. L., Petrology of Sedimentary Rocks.Hemphíll Publ. Co.Austin Texas1974. 182 p. GLEMAREC, M., Vie et MilieuNinoë armoricana n. sp. Polychaete lumbrinereidae from "Grande Vasière". (Gulf de Cascony).1968.315-322.19 (2A) HARTMAN, O., Atlas of the sedentariate polychaetous Annelids from California. Allan Hancock Found. Univ. of Southern CaliforniaLos Angeles1969.812p. LEDOYER, M., Rec. Trav. St. Mar. EndoumeEcologie de la faune vagile des biotopes méditerranéens accesibles en scaphandre autonome (III). Données analytiques sur les biotopes de substrat meuble. 1967.165-186.41(57) ---, Rec. Trav. St. Mar. Endoume Ecologie de la faune vagile des biotopes méditerranéens accesibles en scaphandre autonome. IV. Synthèse de l'étude écologique.1968. 125-295. 44(60) PERES, J. M., Rec. Trav. St. Mar. EudoumeContribution a l´étude des Annélides Polichètes de la Méditerranée occidentale.1954.83-154.8 (13) PERES, J. M., Y PICARD, J., Rec. Trav. St. Mar. Endoume Manuel de Bionomie benthique de la Mer Méditerranée.1964.7-122.31 (47) PICARD. C., Tethys Les peuplements de vase au large du golfe de Fos.1971.569-618.3 (3)(1972) ---, Rec. Trav. St. Mar. EndoumeRecherches qualitatives sur les biocenoses marines des substrats meubles dragables de la région marseillaise.1965.1-6036(52) SOLÍS WOLFOWITZ, V.,Etude des peuplements benthiques des fonds de décantation des parages de Marseille.These Fac. Sci.Marseille1975. 123 p. APÉNDICE
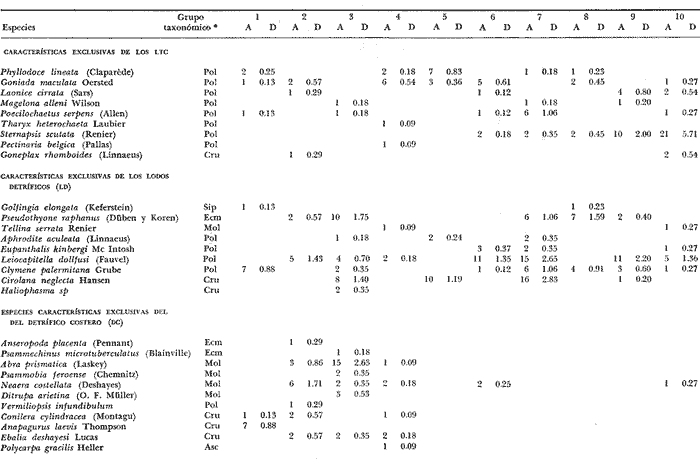 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 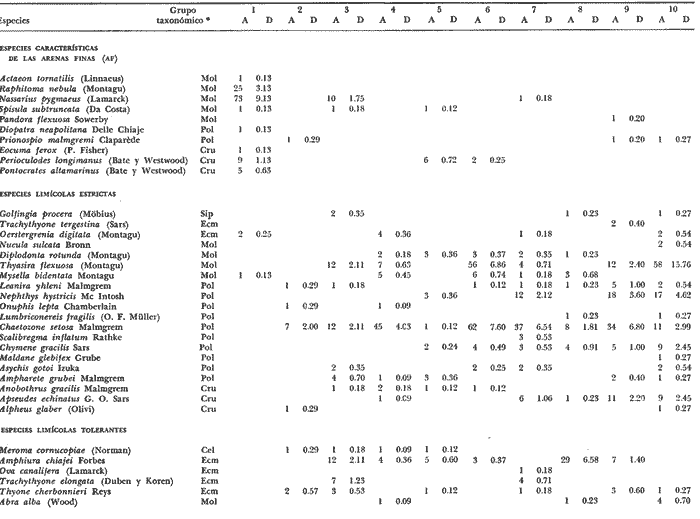 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 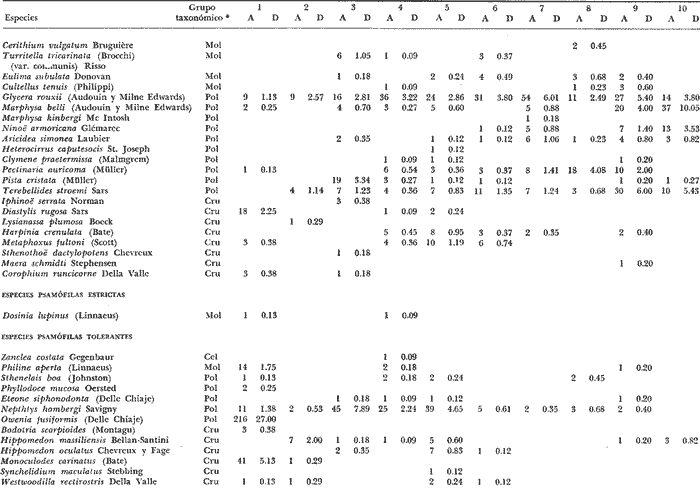 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 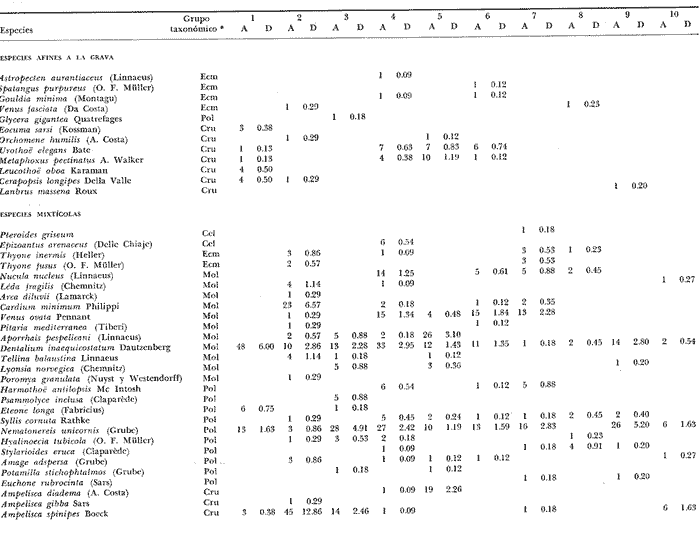 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 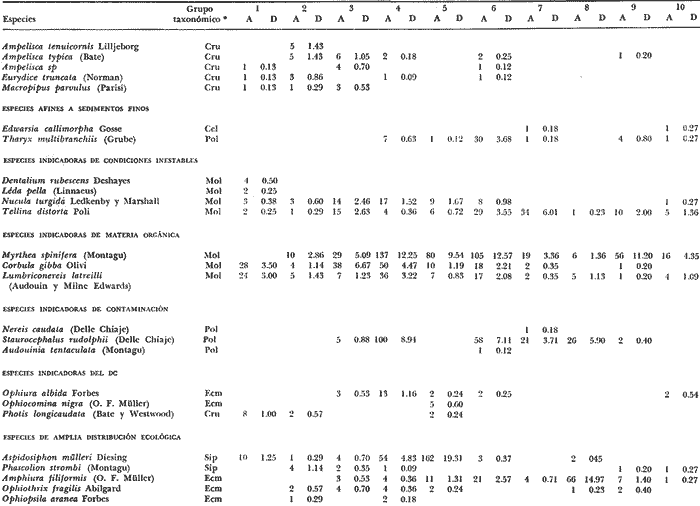 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 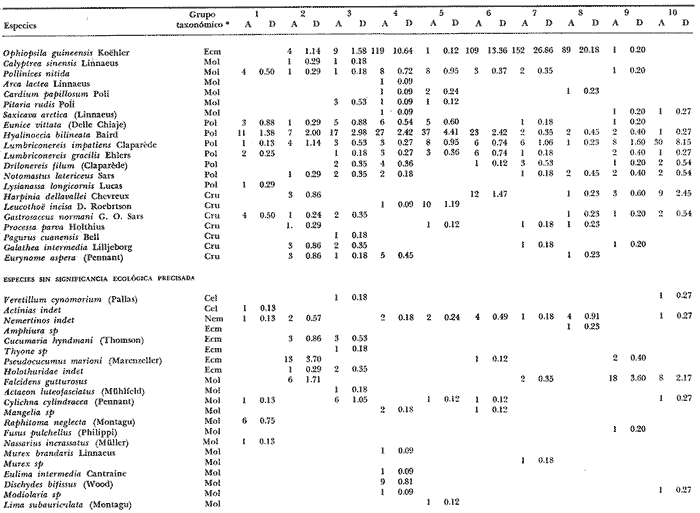 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 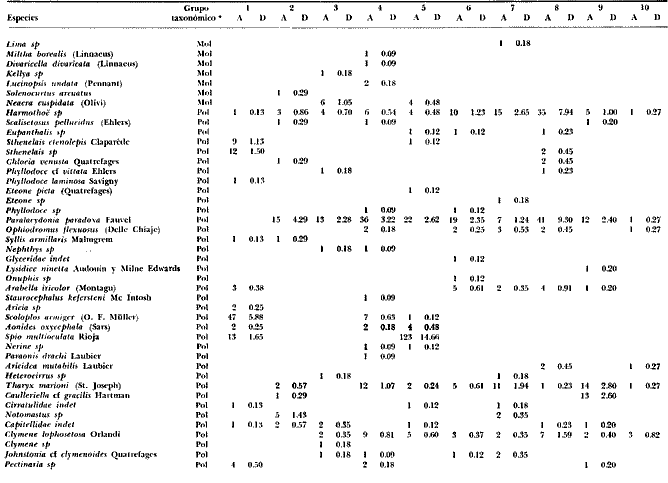 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 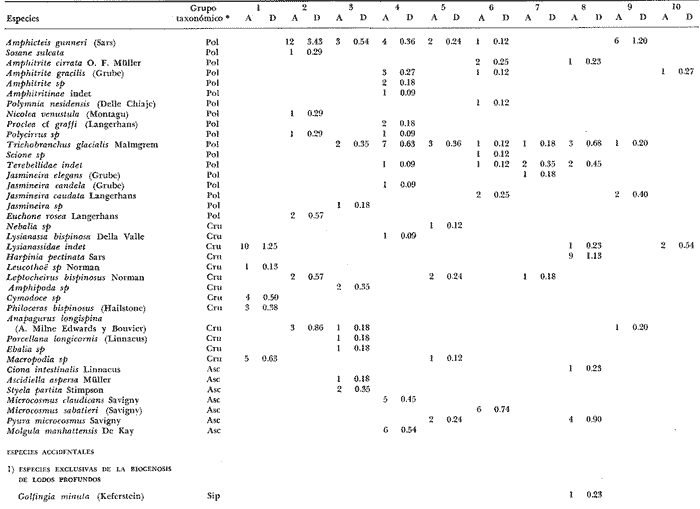 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA 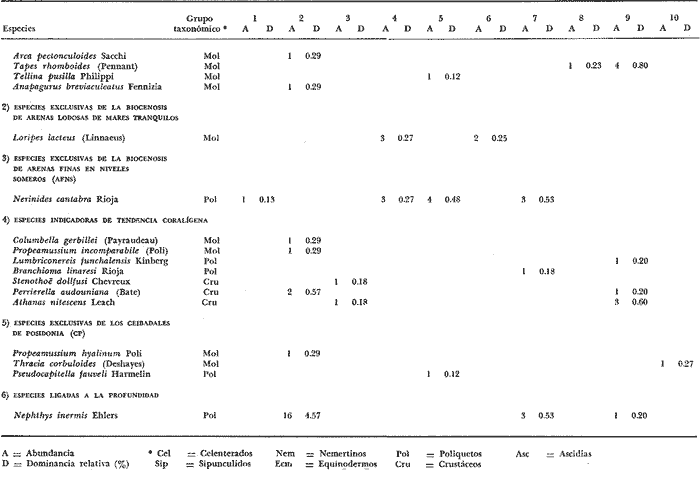 LISTA DE LAS ESPECIES RECOLECTADAS EN CADA ESTACIÓN SEGUN SU CATEGORÍA ECOLÓGICA
|