|
DIVERSIDAD,
DISTRIBUCIÓN Y ABUNDANCIA DE LAS ESPECIES Y POBLACIONES DE PECES
DEMERSALES DE LA SONDA DE CAMPECHE (VERANO 1978)
Trabajo presentado
parcialmente en la I Reunión Nacional sobre Aprovechamiento de la Fauna
de Acompañamiento del Camarón, Guaymas Sonora, Mayo 29, 1979.
Recibido el 20 de agosto de 1980 y aceptado para su publicación el 28 de
noviembre de 1980.
PATRICIA SÁNCHEZ-GIL
ALEJANDRO YÁÑEZ-ARANCIBIA
FELIPE AMEZCUA LINARES
Universidad Nacional
Autónoma de México. Instituto de Ciencias del Mar y
Limnología. Laboratorio de Ictiología y Ecología
Estuarina. Contribución 221 del Instituto de Ciencias del Mar y
Limnología, UNAM.
Se discuten los patrones de
diversidad, distribución y abundancia de las especies y poblaciones de
peces demersales de la Sonda de Campeche, durante el verano de 1978.
Análisis de "cluster" para 17 parámetros ambientales de la zona
de estudió, permite definir dos subsistemas ecológicos del
área, integrados en dendrogramas de similaridad. Fueron analizados
13,872 individuos de 42 arrastres de "trawl" en 21 estaciones de colectas,
distribuidos en 46 familias, 80 géneros y 100 especies. Se efectuaron
estudios de diversidad (H', D, J' y H'w), abundancia (indiv. m-²
g.m-²), distribución, gradiente de biomasa y análisis de
"cluster" de las diferentes colectas para determinar el grado de afinidad
ecológica entre los componentes del ecosistema. Esto último
muestra la presencia de dos grupos de poblaciones cuyos dendrogramas de
afinidad están altamente correlacionados con el dendrograma del habitat.
La integración de un "cluster" total del ecosistema (habitat y
poblaciones de peces) muestra el mismo patrón de los dos subsistemas
definidos aisladamente. Los espectros de diversidad para ambas zonas muestran
poca variación en la equitatividad de las especies pero hay variaciones
en la riqueza de especies y el número de individuos, influenciados por
la heterogeneidad y la biomasa. A su vez la distribución del "standing
crop" y número de individuos se ve influenciado por la
batimetría, los sedimentos y la influencia de aguas epicontinentales de
la Laguna de Términos y estuarios adyacentes. El promedio de biomasa
total para el área de estudio es de 8.0 Kg
Ha-¹.
El 46% de las 100 especies se encuentran
también dentro del complejo lagunar-estuarino de Laguna de
Términos. Quince de las 100 son consideradas típicas de las
comunidades estudiadas: Arilus felis, Cetengraulis edentulus,
Cynoscion nothus, Diplectrum radiale, Priacanthus arenatus, Syacium gunteri,
Symphurus Plagiusa, Trachurus lathami, Synodus foetens, Cholorscombrus
chrysurus, Eucinostomus gula, Opisthonema oglinum, Harengula pensacolae,
Lagocephalus laevigatus yCynoscion arenarius. De
las cuales el 73% en algún momento de su ciclo de vida utilizan los
sistemas estuarinos adyacentes, lo que demuestra la trascendencia
ecológica de la Laguna de Términos en la estructura y
función de las ictiocomunidades demersales de la Sonda de
Campeche.
Demersal fish were collected during
summer, 1978, doing 42 traw1 collections at 21 stations in Campeche Sound to
determine patterns of diversity, distribution and abundance. Seventeen
environmental parameters were also analyzed at each station.
13,872 individual fish distributed in 46 families, 80 genera, and 100
species were analyzed for distribution, abundance (g M-2), and diversity (H',
D, J', H'w). Cluster analysis was used to determine affinity among fish
populations, environmental parameters (habitat), and the combination of the
two. The cluster analysis showed two groups of populations highly correlated
with two habitat cluster. The cluster of ecosystem (habitat plus fish
populations) showed the same pattern of both subsystem defined
índependently. Diversity analysis in both zones indicate little
variation in equitability of species but there are variations in species
richness and number of individuals, affected by heterogeneity and biomass.
Variation of standig crop and number of individuals are influenced by
batimetry, sediments, and epicontinental water influence of Terminos Lagoon and
adjacent estuaries.
Total biomass in the study area was 0.8
g wet weight m-², lower than that reported for Terminos Lagoon. 46% of
the total species collected are also found in Terminos Lagoon. 15 species made
up 82% of the total catch and were widely distributed:Arius felis,
Cetengraulis edentulus, Cynoscion nothus, Diplectrum radiale, Priacanthus
arenatus, Syacium gunteri, Symphurus plagiusa, Trachurus lathami, Synodus
foetens, Chloroscombrus chrysurus, Eucinostomus gula, Opisthonema oglinum,
Harengula pensacolae, Lagocephalus laevigatus,and Cynoscion
arenarius. 73% of these, utilize adjacent estuarine systems during
their life history. Because of this, Terminos Lagoon and other estuarine
systems play a very important role in the ecology of these species.
La Bahía de Campeche está considerada como una zona de particular importancia por su gran potencial biológico. Es una de las regiones que más interés presenta para la pesca en México y ofrece grandes posibilidades para la captura de especies de alto valor comercial entre crustáceos y peces. Se considera como Bahía de Campeche el área sur del Golfo de México entre Veracruz y el noroeste de la Península de Yucatán, y la Sonda o Banco de Campeche es el área sobre la plataforma continental de esta bahía. Ciudad del Carmen junto con Campeche se encuentran entre los puertos pesqueros más importantes siendo terminales pesqueras de la Sonda de Campeche. Allí se capturan comercialmente tres especies de camarón, información que ha sido analizada por Schultz-Ruiz y Chávez (1976), indicando que el Golfo de México contribuye a la producción camaronera nacional con un 45% del total. Por otro lado, la información de la FAO (Klima, 1977), señala que la producción pesquera total del Atlántico, centro-occidental es del orden de 2.8 millones de toneladas (1.4 producción comercial, 0.4 producción deportiva y 1.0 fauna de acompañamiento, la cual es desechada durante las capturas de camarón). Según FAO el rendimiento potencial estimado de todas las especies de la zona, exceptuando los peces pelágicos de altamar, es del orden de 5.369 millones de t. y el rendimiento potencial de peces demersales se calcula en 2.5 millones de t., de las cuales más del 50% se encuentra en el Golfo de México. En este contexto es razonable suponer que la fauna ictiológica que allí ocurre desarrolla un papel ecológico y económico trascendente. El programa OPLAC (Oceanografía de la Plataforma Continental de Campeche) fue iniciado en el Instituto de Ciencias del Mar y Limnología de la UNAM en junio de 1978, con el fin de llevar a cabo investigaciones de carácter multidisciplinario (química marina y contaminación, microbiología, geología, ecología béntica y ecología de peces). Dentro de esa línea el presente trabajo pretende fundamentalmente dar a conocer algunos aspectos ecológicos de la fauna ictiológica de la plataforma continental de Campeche, que además de ayudar a interpretar adecuadamente la ecología de la Laguna de Términos, resulta ser una información valiosa por sí misma puesto que se obtiene de un área poco conocida. OBJETIVOSEste estudio se propone los siguientes objetivos relativos al conocimiento ecológico de las poblaciones de peces demersales: 1. Determinar la composición de la fauna ictiológica en la plataforma continental frente a la Laguna de Términos, Campeche. 2. Precisar algunos de los patrones de distribución de la ictiofauna, en relación con la salinidad, temperatura, profundidad, sedimento, disponibilidad de alimento, otros. 3. Caracterizar las poblaciones y especies típicas de la comunidad, así como las áreas de distribución, diversidad y abundancia. 4. Discutir la estructura y función del sistema ecológico analizado. 5. Realizar un análisis complementario a los estudios ictiológicos existentes para otras áreas del Golfo de México. 6. Detectar líneas de investigación futuras y plantear hipótesis de investigación sobre ecología de recursos pesqueros demersales. ANTECEDENTESUn interesante análisis de la importancia biológica de la Bahía de Campeche hacen Licea-Durán (1971, 1972), Santoyo y Signoret (1972) y Villalobos y Zamora (1975, 1977), donde analizan las características hidrográficas más sobresalientes y su relación con el fitoplancton y la productividad primaria. Sobre la fauna ictiológica existe información de la FAO, el Instituto Nacional de Pesca de México, y de los informes de los Convenios Cubano-Soviéticos. Pero es fundamental destacar que estos estudios se refieren a la Sonda de Campeche frente a la Península de Yucatán (Carranza, 1969; Claro et al ., 1974; Sauskan y Olachea, 1974; Allen y Tashiro, 1976; Klima, 1976a, 1976b y 1977; Erdman, 1977), o hacia el norte del Golfo de México y costa del Atlántico Norteamericano (Merriman y Warfel, 1948; Clarke, 1959; Springer y Woodburn, 1960; Richards, 1963; Roithmayr, 1965; Beamish, 1966; Chávez, 1966; Gunter, 1967; Bullis y Carpenter, 1968; Moore et al ., 1970; Reséndez, 1971; Oviatt y Nixon, 1973; Smith, 1976; Chittenden y McEachran, 1976; Hoese y Moore, 1977). Por lo tanto se carece de información específica frente a la Laguna de Términos. Asimismo, existen estudios sobre la ecología de la fauna ictiológica de la propia Laguna de Términos que corresponden a los resultados de investigaciones desarrolladas en el área entre 1976 y 1979 (Amezcua-Linares y Yáñez-Arancibia, 1980; Bravo-Núñez y Yañez-Arancibia, 1979; Yáñez-Arancibia, 1978; Yáñez-Arancibia y Amezcua-Linares, 1979; Yáñez-Arancibia et al ., 1980; Yáñez-Arancibia y Day, 1981; Vargas Maldonado et al ., 1981). Por otra parte, en general, tampoco existen investigaciones sistemáticas sobre los recursos de peces demersales en la costa del Pacífico de México; pero algunos trabajos ya publicados (Ramírez-Hernández y Arvizu, 1963; Ramírez-Hernández y Páez, 1965; Ramírez-Hernández et al ., 1964 y 1965; Chávez y Arvizu, 1972), constituyen un importante antecedente para proyecciones futuras en esas investigaciones. ÁREA DE ESTUDIOLa Sonda de Campeche o Banco de Campeche forma parte de la plataforma continental al noroeste de la Península de Yucatán. Se extiende en un área de aproximadamente 50,000 millas cuadradas, desde los 200 metros hasta la línea de playa (Villalobos y Zamora, 1975). La zona donde se realizó este estudio se localiza en el extremo oriental de la plataforma continental mexicana frente a la Laguna de Términos, entre los 18º30' - 20º15' N y 91º00' - 93º00' O. Se realizaron 6 transectos y 26 estaciones en 21 de las cuales se efectuaron colectas de peces (Fig. 1). CLIMAPara la Laguna de Términos y áreas adyacentes de acuerdo a García (1973), el clima predominante es Amw, cálido sub-húmedo con lluvias en verano (de mayo a septiembre), la temperatura anual promedio supera los 26ºC y la precipitación anual varía entre los 1,100 y 2,000 mm. Los datos del Servicio Meteorológico Nacional de México (in:Gutiérrez-Estrada, 1977), indican que los vientos tienen una dirección dominante de E a SE durante todo el año con una intensidad máxima de 8 nudos, exceptuando el mes de octubre donde son de N a NO y van de 50 a 72 nudos. 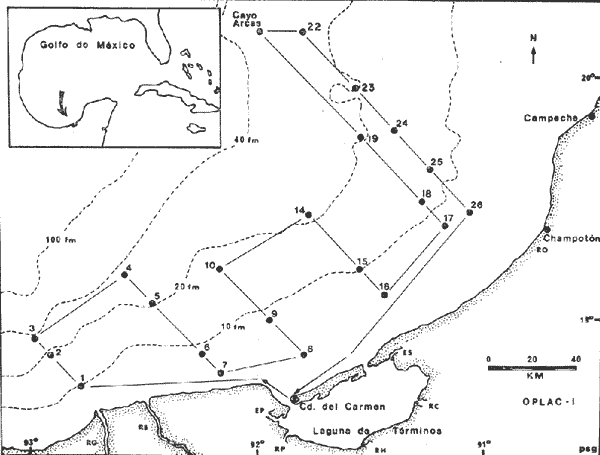 Fig. 1. Sonda de Campeche frente a la Laguna de Términos. Se señalan los principales rasgos fisiográficos y batimétricos, así como los transectos y estaciones del Crucero OPLAC-1. Las abreviaturas corresponden a: RG = Río Grijalva, RS = Río San Pedro, EP = Estero del Pom, RP = Río Palizada, RH = Río Chumpán, RC = Río Candelaria, ES = Estero Sabancuy y RO = Río Champotón. GEOMORFOLOGÍA Y SEDIMENTOSLa plataforma continental frente a la Laguna de Términos es muy amplia y se modifica conforme se incrementa la profundidad, pero la configuración general de la superficie es convexa (Gutiérrez-Estrada, 1977). El análisis de los sedimentos realizado por Gutiérrez-Estrada (1977), indica que la fuente principal de procedencia sedimentaria de la plataforma, es el sistema fluvial de los ríos Grijalva-Usumacinta y la plataforma carbonatada de Yucatán. Machado et al . (1979), encontraron que la porción oeste de la zona costera del Golfo de México adyacente a la Laguna de Términos, es de composición terrígena, mientras que la porción este es de origen calcáreo corroborando el extenso estudio de Gutiérrez-Estrada (1977) (Fig. 2). El contenido de materia orgánica es más abundante en la zona limo-arcillosa por el aporte fluvial y estuarino. que allí se presenta; sin embargo, el porcentaje de Ca CO ³. tiene una relación directa con los sedimentos más gruesos (Tabla l). 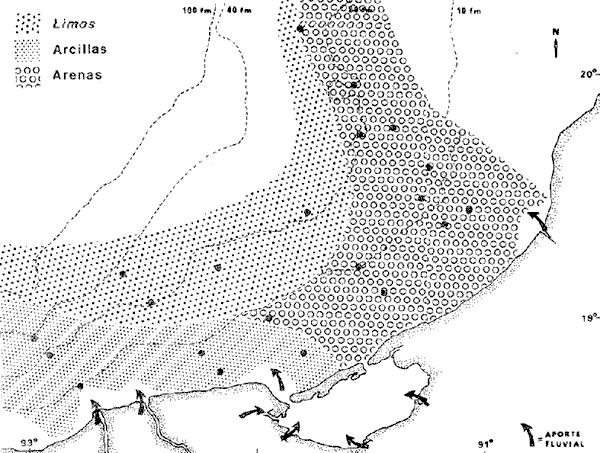 Fig. 2. Composición y distribución de los sedimentos. Basado en Gutiérrez-Estrada (1977) y Machado et al. (1979). Se señalan las principales zonas de aporte fluvial, hacia la plataforma continental. HIDROLOGÍALa dinámica de las aguas neríticas contribuye a la complejidad ambiental del área. En la figura 1 se señala la fisiografía de los aportes fluviales a la plataforma frente a la Laguna de Términos. Villalobos y Zamora (1975) destacan que en esa área existen características hidrológicas sobresalientes por el aporte de las aguas epicontinentales y estuarinas propias de la zona costera. Mancilla y Vargas (1980), han demostrado que a través de la Boca del Carmen se produce el flujo neto de la Laguna de Términos hacia la plataforma continental adyacente. Emi1sson (1976), reporta que durante el verano (época de este estudio) las corrientes litorales se dirigen al noroeste con una velocidad de 6 cm / seg. TEMPERATURA, SALINIDAD Y OXÍGENOLa variación térmica anual de toda el área es de 25 a 29ºC. Villalobos y Zamora (1977), reportan que a fines de primavera el rango de temperatura del agua es de 28 a 29ºC conservando el mismo patrón en la superficie que en el fondo (entre los 5 y 20 m). 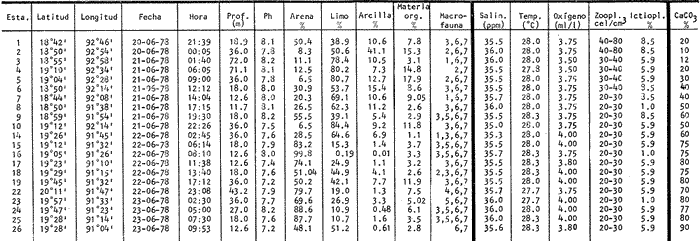 TABLA 1 ESTACIONES DE COLECTA DEL ÁREA DE ESTUDIO. PARÁMETROS AMBIENTALES Y CARACTERIZACIÓN DEL HABITAT La salinidad al igual que la temperatura varía muy poco anualmente, encontrándose los valores más altos en las masas superficiales con un promedio de 35 a 37.8%, Por otra parte en la zona de los ríos Grijalva, San Pedro y Boca del Carmen existe una masa de origen fluvial que influye notablemente en la distribución local de la salinidad. El oxígeno disuelto es relativamente variable. En áreas de influencia de los ríos, los valores más altos se encuentran en las aguas epicontinentales y los menores en las masas oceánicas y zonas costeras, en promedio van de 3.0 a 4.0 ml/l. ASPECTOS ECOLÓGICOS PRELIMINARESEl clima, la hidrología regional, la propia fisiografía costera y las corrientes litorales, determinan la existencia de dos zonas histológicas (Villalobos y Zamora, 1977): una cerca de las 120 millas de distancia de las bocas de la laguna y otra menos significativa paralela a la costa occidental de Yucatán, lo que trae como consecuencia una eutroficación favorable a la productividad del área. De ahí que en estudios cuantitativos, el zooplacton alcanza valores importantes precisamente frente a la Laguna de Términos (de 20 a 80 cel/cm³). Aun cuando este patrón no se presenta durante todo el año, muestra una producción significativa en la estructura del ecosistema. MATERIAL Y MÉTODOS
COLECTAS DE PECES Y OBTENCIÓN DE PARÁMETROS AMBIENTALESLas colectas de peces se efectuaron a bordo de un buque camaronero de gran tamaño denominado "La Nueva Ley de Pesca" perteneciente a la Naviera Rex, S. A. Las artes de pesca fueron dos redes camaroneras tipo comercial que trabajan con una abertura de 30 pies (9 m) y malla de 1 1/3". Estas capturas fueron diurnas y nocturnas a una velocidad promedio de 2 nudos, durante media hora cada una y a diferentes profundidades. La captura de ambas redes para cada estación de colecta fue reunida a bordo, fijada con formalina al 10% y procesada en su totalidad. Se obtuvieron 13,872 individuos, en las estaciones indicadas en la figura 1. Toda la ictiofauna después de ser reetiquetada y preservada en alcohol metílico al 70%, fue determinada taxonómicamente empleando la literatura básica de jordan y Evermann (1896-1900); Meek e Hildebrand (1923-1928); Norman (1934); Cervigón (1966) y Fisher (1978) El análisis biológico de las especies está en proceso. Adicionalmente fue reunida una extensa información de parámetros ambientales para cada estación de colecta (Tabla l), entre los cuales los datos de presencia de macrofauna béntica y de composición de los sedimentos (Machado et al ., 1979) fueron obtenidos simultáneamente con las colectas de peces. Análisis estadísticos determinaron la homología de la información de Villalobos y Zamora (1977) y Gutiérrez-Estrada (1977) para la época de este estudio, y fueron reunidos en la tabla 1. DIVERSIDAD Y ABUNDANCIALa diversidad fue considerada como el número de especies encontradas. Para cada estación de colecta fue calculado el índice de Shanon y Weaver (1963) de la siguiente manera: 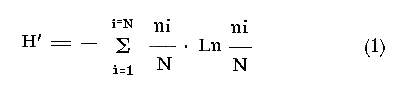 donde ni = al número de individuos de una sola especie y N = al número total de individuos de todas las especies en la colecta. Existen dos componentes principales de la diversidad, la riqueza (D) o variedad de las especies (Margalef, 1969) y la uniformidad (J') o equidad en la distribución de los individuos en las especies (Pielou, 1966). Por lo que fueron calculados estos índices complementarios al anterior. D = (S-1)/Ln (2) donde S = al número total de especies de una colecta y N = al total de individuos de todas las especies. J' = H' / Hmax = H'/Ln S (3) donde Ln de S es el valor máximo posible de H' y H' = H' máx. cuando las especies son igualmente abundantes. La abundancia fue tomada en términos de densidad (número de individuos por área) y de biomasa "standing crop" (g. peso húmedo m -²). El índice de biomasa calculado para cada una de las estaciones de colecta, se obtuvo de la relación modificada de Shanon y Weaver (Wilhm, 1968). 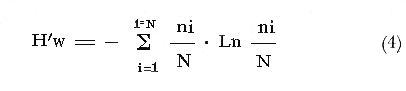 donde ni = a los gramos de una especie en una colecta y N = los gramos totales de todas las especies en la colecta. La relación entre H' y H'w no es necesariamente directa. El uso de estos índices es importante para interpretar la estructura de la comunidad, la afinidad ecológica entre las estaciones de colecta y el valor de importancia de los diferentes patrones en la distribución y abundancia de las poblaciones. Todos los cálculos de estos índices se efectuaron sobre la base de logaritmos naturales. ANÁLISIS DE " CLUSTER "Con la información obtenida, fue elaborado un programa de computación basado en los principios del "cluster analysis" o agrupamiento de datos para analizar la similaridad entre las colectas (Program 46: Single Linkage Cluster Analysis,in: Davies, 1971), consiste en el proceso de reunir en dos o más grupos a las unidades en base a sus atributos y/o características. Las características que son compartidas entre dos o más unidades (estaciones de colecta), proporcionan el grado de afinidad existente. El rango de similaridad va de 0 a 1. Por medio de las caracteristicas individuales cada estación fue comparada contra todas las demás en pasos de 0.01 (de significancia), de tal manera que las estaciones que presentan más características comunes, obtienen los valores más altos de similaridad (muy cercanos a l). Este tipo de programa ha sido utilizado por Horn y Allen (1976), Daniels (1979), Yáfiez-Arancibia et al. (1980) y Vargas Maldonado et. al. (1981), en la delimitación de subsistemas o habitat sobre la base de la afinidad ecológica de poblaciones de peces. La afinidad o similaridad ecológica, puede estar dada en términos cualitativos y/o cuantitativos. En este estudio se utilizaron ambos. Dicho programa permitió efectuar tres análisis de agrupamiento por afinidad de las estaciones de colecta con base a: A) Las especies presentes y su abundancia numérica relativa. B) Sus características ambientales (Tabla 1), y C) La integración ecológica o combinación de las características anteriores (A y B). Además, fue elaborado un diagrama de enrejado (Trellis), esto es, una matriz simétrica m · m con una diagonal principal de similaridades de 1 (Davies, 1971), confrontando entre sí todas las estaciones de colecta con sus valores o coeficientes de similaridad. RESULTADOSFueron analizados 13,872 individuos, distribuidos en 46 familias, 80 géneros y 100 especies. El ordenamiento sistemático sigue el criterio de Greenwood, et al. (1966 y 1967) para categorías supragenéricas. En la lista a continuación, las especies marcadas con: (T) = Se han encontrado dentro de la Laguna de Términos. (A) = Son exclusivas de la Zona A. (B) = Son exclusivas de la Zona B. LISTA SISTEMÁTICA DE LAS ESPECIESClase Chondreichthyes Orden Squaliformes Suborden Galeoidei Familia 1 Carcharhinidae Género 1.Carcharhinus Blainville, 1816 1)Carcharinus remotus (Dumeril, 1865) (B) Familia II Sphyrnidae Género 2. Sphyrna Rafinesque, 1810 2)Sphyrna tiburo (Linnaeus, 1758) (B) Orden Rajiformes Suborden Rhinobatoidei Familia III Rhinobatidae Género 3.Rhinobatus Bloch y Schneider, 1801 3) Rhinobatus lentiginosus(Garman, 1880) (B) Suborden Rajoidei Familia IV Rajidae Género 4. Raja Linnaeus, 1758 4) Raja texana Chandler, 1921 Familia V Dasyatidae Género 5.Dasyatis Rafinesque, 1810 5)Dasyatis sabina LeSueur, 1824 (T) Suborden Torpedinoidei Familia VI Torpedinidae Género 6.NarcineHenle, 1834 6) Narcine brasiliensis(Olfers, 1831) (A) Suborden Myliobatoidei Género 7.Urolophidae Familia VII Urolophus Müller y Henle, 1838 7) Urolophus jamaicensis (Cuvier, 1817) (B) (T) Clase Osteichthyes División I Taeniopaedia Superorden Elopomorpha Orden Anguilliformes Suborden Anguilloidei Familia VIII Congridae Género 8.Congrina Jordan y Hubbs, 1925 8)Congrina flava (Gonde y Bean, 1859) (A) Género 9. Hoplunnis Kaup, 1859 9) Hoplunnis diomedianus Goode y Bean,1895 (A) Superorden Clupeomorpha Orden Clupeiformes Suborden Clupeoidei Familia IX Clupeidae Género 10. Harengula Cuvier y Valenciennes, 1847 10) Harengula pensacolae Goode y Bean, 1879 Género 11.OpisthonemaGill, 1861 11)Opisthonema oglinum(LeSueur,1817) (T) Género 12.ClupanodonLacépéde, 1803 12)Clupanodon pseudohispanicus Poey, 1861 Familia X Engraulidae Género 13. Cetengraulis Günther,1868 13)Cetengraulis edentulus (Cuvier,1829) (T) Género 14. Anchoa Jordan y Evermann, 1927 14) Anchoa lamprotaeniaHildebrand, 1943 (T) 15)Anchoa pectoralis Jordan y Seale, 1926 División III Euteleostei Superorden Protacanthopterygii Orden Salmoniformes Suborden Myctophoidei Familia XI Synodontidae Género 15.Synodus Bloch y Schrieider, 1801 16)Synodus foetens (Linnaeus, 1766) (T) Género 16. Saurida Cuvier y Valenciennes, 1849 17) Saurida brasiliensis Normann (Regan, 1910) Superorden Ostariophysi Orden Siluriformes Familia XII Ariidae Género 17. Arius Cuvier y Valenciennes, 1840 18) Arius felis (Linnaeus, 1766) (T) Género 18.Bagre Oken, 1817 19) Bagre marinus(Mitchill, 1815) (A) (T) Superorden Paracanthopterygii Orden Batrachoidiformes Familia XIII Batrachoididae Género 19.Nautopaedium Jordan, 1919 20)Nautopaedium porosissimum (Cuvier y Valenciennes, 1837 (T) Orden Lophiformes Suborden Antennaroidei Familia XIV Antennarfidae Género 20.AntennariusLacépède, 1798 21)Antennarius ocellatus (Bloch y Schneider, 1780) (A) 22) Antennarius scaber(Cuvier, 1817) (A) Familia XV Ogcocephalidae Género 21.Ogcocephalus Fisher, 1813 23)Ogcocephalus raditans (Mitchill, 1818) Género 22.Halieutichthys Poey, 1863 24)Halieutichthys aculeatus(MitchilI, 1818) (A) Orden Gadiformes Suborden Ophidioidei Familia XVI Ophidiidae Género 23.Lepophidium Gill, 1863 25)Lepophidium brevibarbe (Cuvier, 1829) (A) 26) Lepophidium marmoratum (Goode y Bean, 1885) (A) Superorden Acanthopterygii Orden Gasterosteiformes Suborden Syngnathoidei Familia XVII Syngnathidae Género 24. Hippocampus Rafinesque, 1810 27)Hippocampus hudsonius (DeKay, 1842) (T) Orden Scorpaeniformes Suborden Scorpaenoidei Familia XVIII Scorpaenidae Género 25. Scorpaena Linnaeus, 1738 28) Scorpaena calcarata Goode y Bean, 1882 29) Scorpaena plumieri Bloch, 1789 (T) 30) Scorpaena dispar Longley e Hildebrand, 1940 (A) 31) Scorpaena brasiliensis Cuvier y Valenciennes, 1829 (B) Familia XIX Triglidae Género 26. Prionotus Lacépède, 1802 32) Prionotus punctatus Bloch, 1793 33) Prionotus stearnsi Jordan y Swain, 1844 (A) 34) Prionotus beanii Goode, 1896 (A) 35)Prionotus scitulus Jordan y Gilbert, 1882 (B) Orden Dactylopteriformes Familia XX Dactylopteridae Género 27.Dactylopterus Lacépède, 1802 36) Dactylopterus volitans Linnaeus, 1758 (T) Orden Perciformes Suborden Percoidei Familia XXI Centropomidae Género 28.Centropomus Lacépède, 1802 37)Centropomus undecimalis (Bloch, 1792) (A) (T) Familia XXII Serranidae Género 29. DiplectrumHolbrook, 1856 38) Diplectrum formosum(Linnaeus, 1766) 39) Diplectrum radiale(Quoy y Gaimard,1824) (T) Género 30.Serranus Cuvier, 1817 40)Serranus atrobranchus (Cuvier,1829) (A) Género 31 Epinephelus Bloch, 1793 41) Epinephelus guttatus (Linnaeus, 1758) (A) (T) Familia XXIII Priacanthidae Género 32. Priacanthus Cuvier, 1817 42)Priacanthus arenatus Cuvier y Valenciannes, 1829 Familia XXIV Echeneidae Género 33.Echeneis Linnaeus, 1758 43) Echeneis naucrates Linnaeus, 1758 (B) (T) Familia XXV Carangidae Género 34.Selar Bleeker, 1850 44) Selar crumenophthalmus (Bloch, 1793) Género 35.Trachurus Rafinesque, 1810 45)Trachurus lathami Nichols, 1920 Género 36 Vomer Cuvier y Valenciennes, 1833 46) Vomer setapinnis (Mitchill, 1815) Género 37.SeleneLacépède, 1803 47)Selene vomer (Linnaeus,1758) (A) (T) Género 38. ChoroscombrusGirard, 1858 48) Chloroscombrus chrysurus(Linnaeus, 1766) (T) Género 39.Caranx Lacépède, 1802 49)Caranx crysos (Mitchill, 1815) (B) Familia XXVI Lutjanidae Género 40. RhomboplitesGill, 1862 50)Rhomboplites aurorubens (Cuvier y Valenciennes, 1862) (A) Género 41. Pristipomoides Bleeker, 1852 51) Pristipomoides macrophthalmusMüller y Troschel, 1884 Género 42.Lutjanus Bloch, 1790 52)Lutjanus synagris (Linnaeus, 1758) (T) Familia XXVII Gerridae Género 43. Eucinostomus Baird y Girard, 1855 53) Eucinoslomus gula (Cuvier y Valenciennes,1830) (T) 54)Eucinostomus argenteus (Baird y Girard,1854) (T) Género 44.DiapterusRanzani, 1840 55)Diapterus rhombeus(Cuvier y Valenciennes, 1830) (A) Familia XXVIII Pomadasyiclae Género 45.Haemulon Cuvier, 1829 56)Haemulon aurolineatum Cuvier, 1829 (T) 57) Haemulon sp (A) (T) Género 46. OrthopristisGirard, 1859 58) Orthopristis chrysopterus (Linnaeus, 1766) (T) Familia XXIX Sparidae Género 47.Calamus Swaison, 1839 59)Calamus calamus (Cuvier y Valenciennes, 1830) (T) Género 48, Archosargus Gill, 1865 60) Archosargus unimaculatus(Bloch, 1792) (T) Género 49.Otrynter Jordan y Evermann, 1896 61)Otrynter caprinus (Bean, 1882) Familia XXX Sciaenidae Género 50. CynoscionGill, 1854 62) Cynoscion nothus (Holbrook, 1855) (A) (T) 63)Cynoscion arenariusGingsburg;1929 (T) Género 51 Menticirrhus Gill, 1861 64) Menticirrhus americanus (Linnaeus, 1758) Género 52. StelliferOken, 1817 65) Stellifer colonensisMeek e Hilde brand 1925 (T) Género 53.Eques Bloch, 1793 66) Eques lanceolatus (Linnaeus, 1758) (B) Familia XXXI Mullidae Género 54.Upeneus Cuvier, 1829 67)Upeneus Parvus Poey, 1853 Familia XXXII Ephippidae Género 55.Chaetodipterus Lacépède, 1802 68)Chaetodipterus faber (Broussonet,1782) (B) (T) Suborden Sphyraenoidei Familia XXXIII Sphyraenidae Género 56.Sphyraena Bloch y Schneider, 1801 69)Sphyraena guachancho (Cuvier y Valenciennes, 1883) Suborden Polynemoidei Familia XXXIV Polynemidae Género 57. PolynemusLinnaeus, 1754 70) Polynemus octonemusGirard,1858 (T) Suborden Labroidei Familia XXXV Scaridae Género 58. Nicholsina Fowler, 1915 71) Nicholsina ustus(Valenciennes,1839) (B) (T) Suborden Gobioidei Familia XXXVI Gobiidae Género 59. Bollmania Jordan, 1889 72) Bollmania boqueronensis Evermann y Marsh, 1892 Suborden Scombroidei Familia XXXVII Trichiuridae Género 60.TrichiurusLinnaeus, 1758 73)Trichiurus lepturus (Linnaeus, 1758) (A) (T) Familia XXXVIII Scombridae Género 61. Scomber Linnaeus, 1758 74)Scomber colias Gmelin, 1789 (A) Familia XXXIX Stromatidae Género 62. Peprilus Cuvier, 1800 75) Peprilus triacanthus (Peck, 1800) 76) Peprilus paru (Linnaeus, 1758) Orden Pleuronectiformes Suborden Pleuronectoidei Familia XL Bothidae Género 63. Syacium Ranzani, 1840 77) Syacium gunteri Gingsburg, 1933 78) Syacium micrurum Ranzani, 1840 (A) 79) Syacium papillosum(Linnaeus, 1758) Género 64.Engyophrys Jordan y Bollman, 1890 80)Engyophrys sentus Gingsburg,1933 (A) Género 65. Cyclopsetta Gill, 1889 81) Cyclopsetta fimbriata Goode y Bean, 1886 Género 66. TrichopsettaGill, 1889 82) Trichopsetta ventralis (Goode y Bean,1886) (A) Género 67.Ancylopsetta Gill, 1864 83)Ancylopsetta quadrocellata Gill, 1864 (A) (T) 84) Ancylopsetta dilecta (Goode y Bean, 1883) Género 68.Citharichthys Goode, 1881 85)Citharichthys macrops Drësel, 1885 86)Citharichthys spilopterus (Günther,1862) (T) Género 69. Etropus Jordan y Gilbert, 1882 87) Etropus crossotus Jordan y Gilbert, 1886 (T) Género 70.Bothus Rafinesque, 1810 88)Bothus ocellatus (Agassiz, 1831) Suborden Soleoidei Familia XLI Soleidae Género 71. Achirus Lacépède, 1803 89) Achirus fasciatusLacépède, 1766 (A) 90) Achirus lineatus (Linnaeus, 1758) (B) (T) Género 72.Gymnachirus Kaup, 1858 91) Gymnachirus nudus Kaup, 1858 (A) Familia XLIl Cynoglossidae Género 73.Symphurus Rafinesque, 1810 92)Symphurus plagiusa Linnacus, 1766 (T) Orden Tetraodontiformes Suborden Balistoidei Familia XLIII Balistidae Género 74.Balistes Linnaeus, 1758 93)Balistes capriscus (Gmelin, 1788) (T) Género 75. MonacanthusCuvier, 1817 94) Monacanthus hispidus (LeSueur, 1824) (B) (T) Género 76.Alutera Cuvier, 1817 95)Alutera schoepfi (Walbaum, 1792) Familia XLIV Ostraciidae Género 77.LactophrysSwainson, 1758 96)Lactophrys tricornis (Linnaeus,1758) (B) (T) Suborden Tetraodontoidei Familia XLV Tetraodontidae Género 78.Lagocephalus Swainson, 1839 97)Lagocephalus laevigatits (Linnaeus,1766) (T) Género 79. SphoeroidesLacépède, 1798 98)Sphoeroides dorsalis Longley, 1934 99)Sphoeroides greeleyi (Gilbert, 1900) (T) Familia XLVI Diodontidae Género 80.Chilomycterus Bibron, 1846 100)Chilomycterus schoepfi (Walbaum, 1792) (T) DIVERSIDAD Y ABUNDANCIA EN LAS ESTACIONES DECOLECTALa cuantificación está basada en el registro total de la captura para cada una de las estaciones de colecta. Con base a este registro individual se puede presentar el panorama que se describe a continuación. El número de especies colectadas en cada estación varía de 15 (Estaciones 14 y 22) a 28 (Estaciones 1 y 24). Los valores del índice H' van de 0.81 (Estación 15) a 2.49 (Estación 9). La densidad (indvm-²) varía de 0.001 (Estación 22) a 0.37 (Estación 4). El rango de "standing crop" (g m-²) va de 0.28 (Estación 22) a 1.65 (Estación 4). De las, 21 colectas de peces más del 90% de las especies se obtuvo en la colecta 14 (Estación 17). Presentándose una relación entre colectas y el número de especies, muy evidente hasta dicha estación; pero posteriormente el incremento de especies no es significativo (Fig. 3), con la aparición de especies raras y muy poco abundantes. AFINIDAD DE LAS ESTACIONES Y DELIMITACIÓN DEL HABITATEn base al agrupamiento por afinidad de las estaciones de colecta, fueron determinados dos grandes grupos, denominados en este trabajo Zona A y Zona B (Figs. 4, 5 y 6, Tabla 2). A) La afinidad por presencia y abundancia numérica de las especies tiene un valor de 0.96 para las estaciones de la Zona A, mientras que para las estaciones de la Zona B, la afinidad es de 0.95. El valor de similaridad existente entre ambos grupos es de 0.90 (representado por la estación 3). B) La afinidad por características del habitat (parámetros ambientales de la tabla 1) es de 0.97 en ambas zonas. La afinidad entre estos grupos es de 0.96. 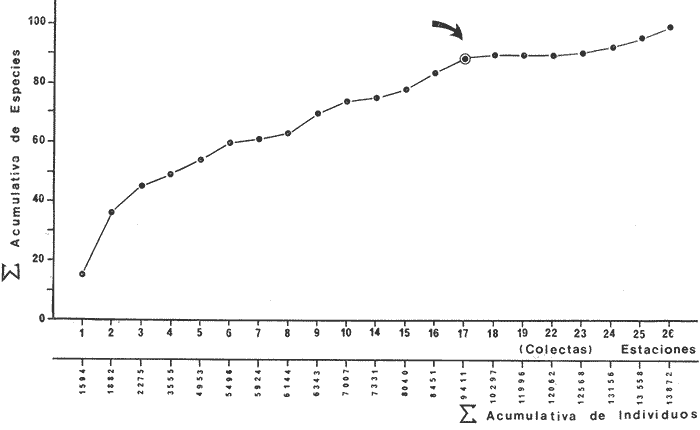 Fig. 3. Incremento acumulativo de las especies en relación a todas las estaciones de colecta y a la sumatoria acumulativa del número de individuos. En la colecta 14 (Estación 17) se obtiene el 90% del total de las especies.  TABLA 2 DELIMITACIÓN DEL HABITAT Y DEFINICIÓN DEL ECOSISTEMA 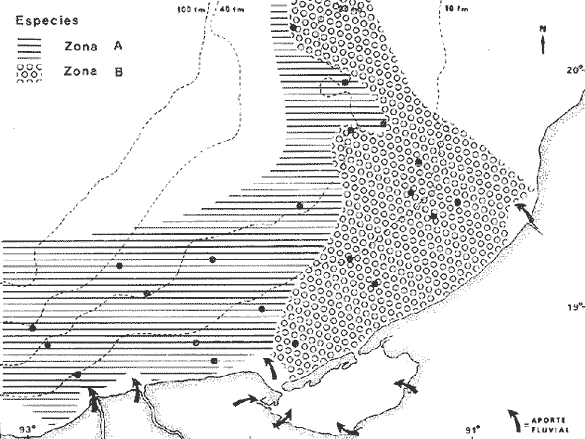 Fig. 4. Afinidad de las estaciones de colecta con base a la composición y abundancia numérica de las especies. Se muestra la agrupación de las estaciones en dos grandes zonas delimitadas por el análisis de "cluster". C) Por último, la afinidad resultante de la relación de los valores de abundancia numérica de las especies y los valores de las características ambientales tomadas de cada estación es de 0.96 para la Zona A, para las estaciones de la Zona B es de 0.97. La similaridad existente entre ambos grupos es de 0.95. El dendrograma analizado en la discusión detalla este panorama (Figs. 24 y 25). DIVERSIDAD DE LA COMUNIDADDoce familias constituyen el 88.56% en número de la captura total (Tabla 3). Para el total de la captura diez especies aportan el 75% en número (Tabla 4). Los índices calculados y su distribución en ambas zonas se concentran en la tabla 5. 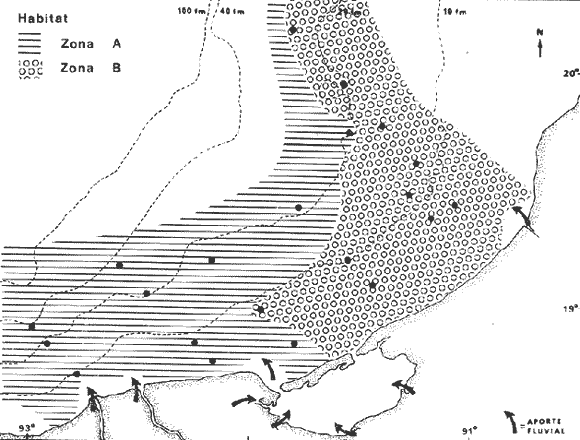 Fig. 5. Afinidad de las estaciones de colecta con base a los parámetros ambientales (Tabla 1). Se muestra la agrupación de las estaciones en dos grandes zonas delimitadas por el análisis de "cluster". 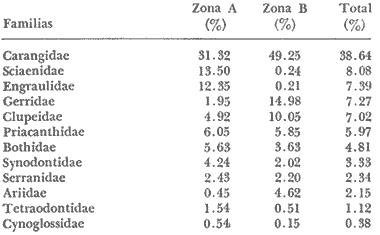 TABLA 3 DISTRIBUCIÓN PORCENTUAL DE LAS PRINCIPALES FAMILIAS 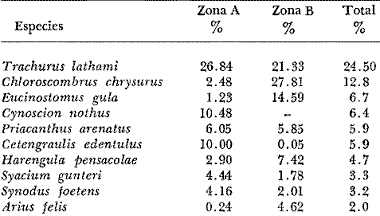 TABLA 4 DISTRIBUCIÓN PORCENTUAL DE LAS PRINCIPALES ESPECIES  Fig. 6. Afinidad de las estaciones de colecta con base a la integración ecológica de la composición y abundancia numérica de las especies y las características ambientales (Tabla 1). Se muestra la agrupación de las estaciones en dos grandes zonas delimitadas por el análisis de "cluster". DISTRIBUCIÓN DE LAS POBLACIONESPara el total de la captura, 15 especies resultaron ser las más importante s por su frecuencia y abundancia (Tabla 6). La distribución de estas especies en el área sigue, aparentemente, tres patrones generales, de los cuales dos son muy definidos.  TABLA 5 RANGOS DE LOS ÍNDICES ECOLÓGICOS BatimetríaDe acuerdo a la batimetría, en profundidades menores a 18 m, Arius felis se presenta a todo lo largo de la zona litoral (Fig. 7). A su vezCetengraulis edentulus y Cynoscion nothusse encuentran sólo en la zona A distribuidos hasta los 36 m (Figs. 8 y 9). Para profundidades siempre mayores a los 18 mDiplectrum radiale, Priacanthus arenatus ySyacium gunteri se encuentran abarcando estaciones tanto de la Zona A como de la B (Figs. 10, 11 y 12). Symphurus plagiusa aun cuando se presenta en algunas estaciones de la Zona B, es más abundante en la Zona A (Fig. 13). 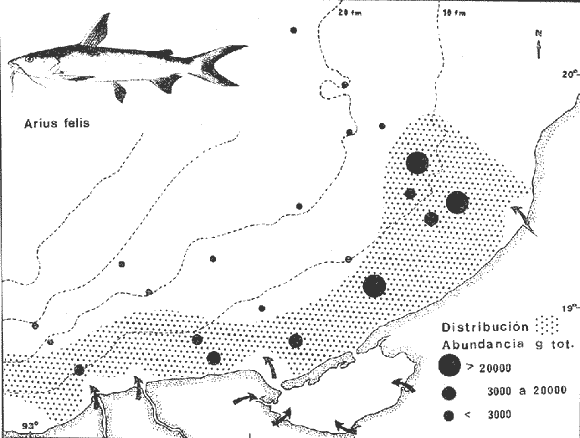 Fig. 7. Distribución y abundancia de: Arius felis en la Sonda de Campeche Crucero OPLAC-1, verano de 1978. 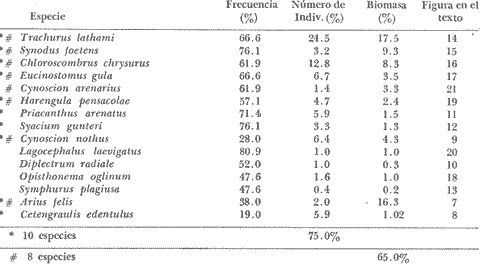 TABLA 6 DISTRIBUCIÓN DE FRECUENCIA Y ABUNDANCIA DE LAS ESPECIES TIPICAS DE LA COMUNIDAD EN LA CAPTURA TOTAL 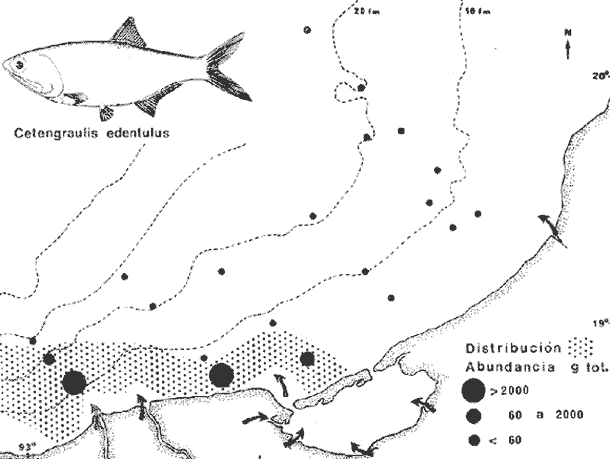 Fig. 8. Distribución y abundancia de: Cetengraulis edentulus en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Descarga de ríosOtro patrón está determinado para especies que, en mayor o menor grado, están pobremente representadas en las áreas de aporte fluvial de la región; como se observó con Trachurus lathami, Synodus foetens, Chloroscombrus chrysurus yEucinostomus gula (Figs. 14, 15, 16 y 17), aun cuandoS. foetens y E. gula son componentes típicos de las comunidades lagunares. OtrosPor último, las especiesOpisthonema oglinum, Harengula pensacolae, Lagocephalus laevigatus y Cynoscion arenarius, muestran una distribución cuyo patrón parece seguir un comportamiento distinto a los anteriores (Figs. 18, 19, 20 y 21). Al momento, esta distribución no puede ser explicada enteramente pero podría tener relación con los sedimentos. Los valores más altos de abundancia dentro de las estaciones de la Zona A, van de 200 a más de 14,000 g de captura total. Cetengraulis edentulus, Cynoscion nothus, Syacium gunteri, Symphurus plagiusa, Trachurus lathami, Synodus foetens, Opisthonema oglinum, y Cynoscion arenarius. 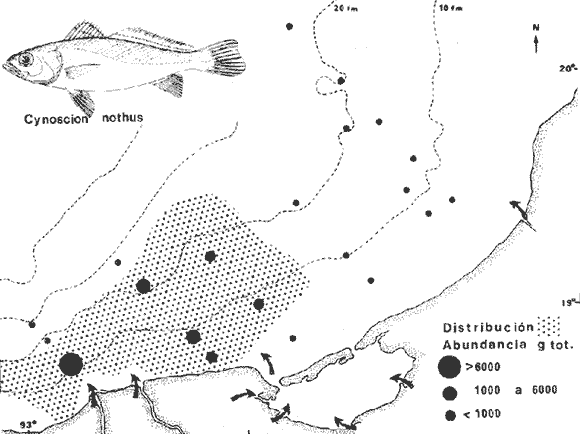 Fig. 9. Distribución y abundancia de: Cynoscion nothus en la Sonda de Campeche. Crucero OPLAC-I, verano de 1978. Para la Zona B, los valores más altos van de 200 a más de 40,000 g de captura total. Arius felis, Diplectrum radiale, Priacanthus arenatus, Chloroscombrus chrysurus, Eucinostomus gula, Harengula pensacolaeyLagocephalus laevigatus. ABUNDANCIA DE LAS ESPECIES Y POBLACIONESDoce familias constituyen el 81.51% del peso de la captura total (Tabla 7). Para el total de la captura ocho especies aportan el 65% del peso total (Tabla 8). DISCUSIÓN
HABITATEs posible establecer que las características ambientales con mayor implicación ecológica en la plataforma continental adyacente a la Laguna de Términos, dependen en gran medida de la circulación litoral y del flujo de los ríos Grijalva y San Pedro en el extremo occidental y del río Champotón al oriente; mientras que el área central depende principalmente de la descarga de la Laguna de Términos a través de la Boca del Carmen. Es decir, a la zona de estudio le corresponden características muy especiales debido al intercambio de aguas oceánicas y costeras, además de la transición de materiales terrígenos y sedimentos calcáreos de la región. 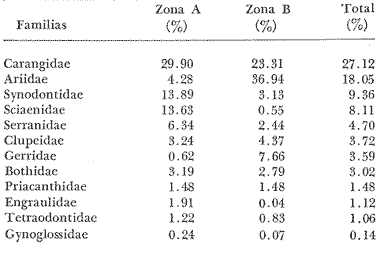 TABLA 7 DISTRIBUCIÓN PORCENTUAL DE LAS PRINCIPALES FAMILIAS  TABLA 8 DISTRIBUCIÓN PORCENTUAL DE LAS PRINCIPALES ESPECIES Los valores de salinidad y temperatura presentan un patrón casi uniforme, compuesto por salinidades que van de 35.0 a 36.0%, y temperaturas de 27.7 a 28.3ºC, por lo cual no parece que estos factores sean los determinantes en la distribución de las especies durante este estudio. Hacia el extremo noroeste del área, posiblemente como reflejo de la descarga de la laguna, se encontraron los valores más altos de pH (7.8-8.2) acompañados de los más bajos de oxígeno disuelto (3.75 ml/1) y en las estaciones más cercanas a la Boca del Carmen la concentración más alta de materia orgánica (10%), consecuencia de la variedad de productores primarios existente dentro de la laguna (Yáñez-Arancibia y Day, 1981). En la porción noreste del área de estudio se presenta la relación inversa de estos parámetros, con los valores más bajos de pH (7.2-7.9), los valores más altos de oxígeno disuelto (4.0 ml/l) y la concentración más baja de materia orgánica (3-10%). Estas diferencias ambientales dentro del área de colecta, se ven corroboradas por otros parámetros del habitat como son la composición y distribución de los sedimentos (Fig. 2), lo cual enmarca este planteamiento. La relación independiente de estos parámetros, plantea fluctuaciones poco marcadas en toda el área, reguladas por la meteorología, climatología y oceanografía locales (Tabla l). El análisis de esta discusión desde un punto de vista cuantitativo, muestra un alto grado de afinidad entre las estaciones de colecta donde se alcanzan valores muy significativos de similaridad ecológica en dos grupos bien definidos y en el área total (Figs. 5 y 24B). COMPOSICIÓN DE LA FAUNA ICTIOLÓGICAAsociada a las características ambientales, la fauna ictiológica de la plataforma continental está también muy relacionada con la Laguna de Términos. De las 100 especies capturadas, el 46% pueden ser encontradas en alguna etapa de su vida dentro de la laguna. Quince especies son consideradas típicas de la comunidad por su distribución, frecuencia y abundancia, por lo que han sido tratadas de manera particular en este trabajo (Tabla 6). De estas 15 especies, 11 (73%) se encuentran en la Laguna de Términos; sin embargo, la fauna que compone a cada una de las zonas descritas, como A y B, aparentemente presenta relaciones distintas con ella (lo que es analizado más adelante en Distribución). Se encontró que el 30% de las especies de la captura total, está compuesto por especies exclusivas de la Zona A, de las cuales 8 (27%) se han reportado en el interior de la laguna. Estas especies no son representativas, en cuanto al número de individuos y frecuencia, pero sí en cuanto a su biomasa. Por otra parte el 16%, de especies de la captura total, está compuesto por especies exclusivas de la Zona B, de las cuales también 8 (50%) se han reportado dentro de la laguna, pero en su mayoría son especies poco significativas en cuanto a sus valores de importancia. En cuanto a la composición por familias, cabe mencionar que la familia Carangidae, aun cuando no es típica de comunidades demersales, presenta 6 especies y los valores más altos en número de individuos (38.64%) y biomasa (27.12%) de la captura total. Esta familia es de hábitos pelágicos y su ocurrencia en un muestreo demersal puede deberse a los siguientes factores: a) sus hábitos alimenticios carnívoros predando sobre organismos demersales; b) lo somero de la Sonda de Campeche y la eventual relación estrecha entre el bentos y el pélagos; c) sus hábitos gregarios, lo que permite que por formar cardúmenes su captura sea abundante; d) el arte de pesca que baja y sube abierto y puede capturar fauna en toda la columna de agua. DIVERSIDADAun cuando a nivel de familia, Carangidae resulta ser la mejor representada en toda el área, aparentemente la estructura de la fauna para las dos zonas es diferente. La proporción de las familias dominantes es casi inversa en cuanto a cantidad de individuos en cada una de las dos zonas. A nivel de especie esta diferencia no es menos aparente, encontrándose que las especies mejor representadas en la Zona A no lo son para la B, con algunas excepciones (Tablas 3 y 4). Complementando esta información con los valores de los diferentes índices calculados para evaluar la diversidad, se observó que las zonas del área de estudio son muy semejantes en cuanto a su uniformidad, pero presentan diferencias en el número de individuos y riqueza de especies, siendo mayores los valores que corresponden a las estaciones de la Zona A (Tabla 5, Figs. 22 y 23). El análisis "cluster", muestra una afinidad muy grande entre las estaciones que forman la Zona A (0.96) y las que componen la B (0.95); sin embargo, la similaridad existente entre ambas es menor (0.90). A su vez el diagrama Trellis, presenta porcentajes muy altos de afinidad entre todas las estaciones de colecta (82-97%), destacando los valores más bajos en la estación 3 y los más altos en la estación 8. Esta última se encuentra en un área intermedia entre las dos zonas. Los datos reflejan una comunidad homogénea en cuanto a su estructura general (equitatividad), pero está representada en cada una de las zonas por especies distintas (Figs. 24A y 25). El 54% de las especies de toda la captura son comunes a las dos zonas, pero en la Zona A se encuentra el 84% de todas las especies compuesto por el 59% de los individuos de la captura total. A su vez, en la Zona B se presenta el 70% de las especies representadas por el 41% en número de individuos. Esta información establece una mayor diversidad en la Zona A, lo que probablemente está determinado por las características ambientales que en ella se encuentran, ya que existe una correlación muy alta entre estos parámetros. La integración de un "cluster" total (habitat, poblaciones de peces), confirma este planteamiento, agrupando con un valor muy alto de similaridad (0.95), a las estaciones de las dos zonas en discusión (Fig. 24C). La estructura general de la comunidad, está representada por valores semejantes a los reportados por otros autores para comunidades de peces demersales al norte del Golfo de México. De un total de 100 especies colectadas durante el muestreo, 10 resultaron ser las más abundantes numéricamente constituyendo el 75.4% de la captura total (Tabla 4). Un patrón similar de dominancia es señalado por Merriman y Warfel (1948) en Block Island Texas, donde 6 especies aportan el 91% del número total de individuos, Richards (1963) en Long Island Sound, donde 10 especies constituyen el 91%, Oviatt y Nixon (1973) en Narragansett Bay Rhode Island, 10 especies aportan el 91%. Aun cuando las especies representativas para cada una de estas áreas no son las mismas, estos datos permiten estimar la abundancia relativa de estas comunidades en las costas del Golfo; asimismo se encontró que la relación de los índices de diversidad y biomasa calculados en este estudio, son comparativamente mayores a los reportados en otras localidades (Tabla 9). 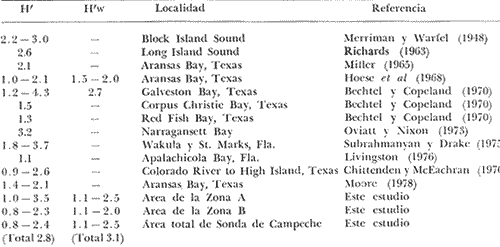 TABLA 9 VALORES DE DIVERSIDAD DE COMUNIDADES DE PECES EN DIFERENTES ÁREAS DEL GOLFO DE MÉXICO Y COSTA ATLÁNTICA DE NORTEAMÉRICA DISTRIBUCIÓNAparentemente la distribución de las especies de este estudio, está determinada principalmente por tres parámetros ambientales asociados. La composición y distribución de los sedimentos, la batimetría y la influencia fluvial y estuarina. Para algunas especies, el parámetro determinante es la batimetría. Estas especies son generalmente eurihalinas y la mayoría se han reportado en aguas estuarinas (Gunter, 1945; Miller, 1965; Thomas et al., 1971; Yáñez-Arancibia et al.,1980). Las especies que se encuentran ocupando profundidades menores a los 36 m, son especies características de zonas costeras muy comunes en fondos fangosos, asociadas a aguas estuarinas y bocas de ríos, especialmente Arius felis yCynoscion nothus (Chittenden y McEarchran, 1976; Fisher, 1978), las cuales se alimentan principalmente de crustáceos y peces, aunque Arius felis según Springer y Woodburn (1960), tiene un espectro trófico más amplio. Ambas especies son muy abundantes a lo largo de las costas de EUA (Anderson, 1968, in : Chittenden y McEachran, 1976). Cetengraulis edentulus presenta hábitos pelágicos y es esencialmente una especie filtradora (Fisher, 1978). Aún cuando Arius felisse ha reportado a profundidades mayores (91 m, Franks et al.,1972), es más abundante en aguas poco profundas, las otras dos especies no se han reportado a más de 30 m (Fisher, 1978). La mayor abundancia deArius felis , se encontró dentro de las estaciones de la Zona B (Fig. 7). Chittenden y McEachran (1976), mencionan que las especies que habitan sustratos blandos, no se les encuentra más allá de los 110 m. Algo similar fue observado con Cetengraulis edentulus y Cynoscion nothus quienes presentaron su mayor abundancia dentro de la Zona A (Figs. 8 y 9). Otras especies sólo fueron colectadas a profundidades mayores a 18 m, su distribución es muy amplia pero sus valores de abundancia en la captura total no son muy significativos y no parecen estar relacionados con la composición de los sedimentos del área de estudio. Aun cuando se ha reportado que Diplectrum radiale y Priacanthus arenatusson son comunes en fondos rocosos y coralinos (Smith, 1976), en este estudio sólo fueron ligeramente más abundantes en las estaciones de la Zona B (Figs. 10 y 11). Se dice que estas especies son frecuentes entre los 15 y 75 m, donde los ejemplares más grandes tienden a ocupar las profundidades mayores (Fisher, 1978). Syacium gunteri y Symphurus plagiusa son característicamente eurihalinas (Springer y Woodburn, 1960) y se les encuentra comúnmente asociadas a fondos fangosos (Cervigón, 1966; Fisher 1978). En este trabajo la mayor abundancia de estas especies se presentó en la Zona A (Figs. 12 y 13). Estas especies poseen hábitos alimenticios muy distintos, Diplectrum radiale y Priacanthus arenatus se alimentan principalmente de crustáceos y peces mientras que Syacium gunteri y Symphurus plagiusa de pequeños invertebrados, poliquetos, copépodos, anfípodos y ostrácodos (Cervigón, 1966; Randall, 1967; Fisher, 1978). 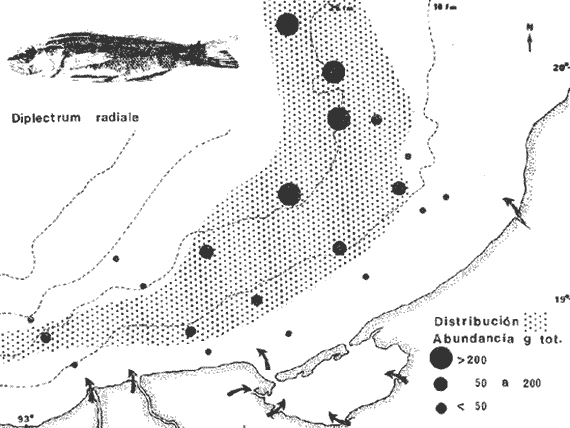 Fig. 10. Distribución y abundancia de: Diplectrurn radiale en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Un segundo patrón fue presentado por especies cuya distribución no parece estar relacionada con la profundidad. Estas especies se encontraron pobremente representadas en las áreas de aporte fluvial, todas son típicamente marinas y poseen hábitos gregarios.Trachurus lathami , según Chittenden y McEachran (1976), se encuentra ocupando fondos rocosos; sin embargo, en este estudio, T. lathami , se colectó con frecuencia en fondos fangosos y/o arcillosos.Synodus foetens se ha reportado comúnmente en fondos fangosos (Cervigón, 1966) y en sus formas adultas como visitante de estuarios (Thomas et al.,1971; Yáñez-Arancibiaet al., 1980). Ambas especies se alimentan principalmente de peces. La mayor abundancia de estas especies se registró en la Zona A (Figs. 14 y 15).Chioroscombrus chrysurus yEucinostomus gula, no se consideran específicas de algún sedimento, se les ha colectado de manera semejante a este estudio tanto en zonas fangosas como arenosas; sin embargo, se ha observado que los ejemplares adultos deEucinostomus gula, tienden a ocupar fondos arenosos y los ejemplares pequeños de ambas especies se encuentran en áreas protegidas, o muy cercanas a la costa como lagunas y manglares (Chávez 1972; Fisher, 1978; Yáñez-Arancibiaet al.,1980), su alimentación es muy variada incluyendo pequeños invertebrados, poliquetos, copépodos, anfípodos, ostrácodos. La abundancia de estas especies fue mayor dentro de la Zona B (Figs. 16 y 17). 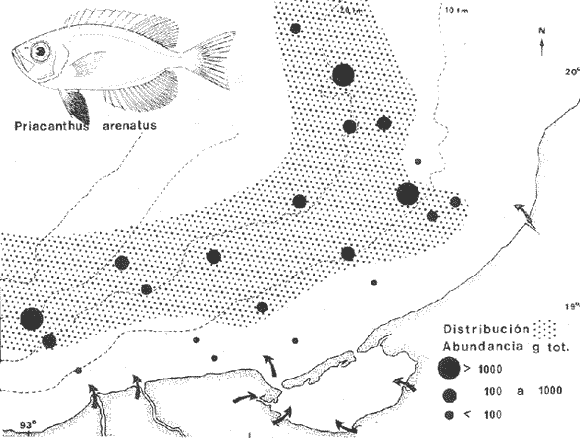 Fig. 11. Distribución y abundancia de: Priacanthus arenatus en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Otros patrones de distribución semejantes a éstos, han sido observados para comunidades de peces demersales en otras áreas del Golfo de México (Hildebrand, 1955; Topp y Hoff, 1972; Chittenden y McEachran, 1976). Por último, hubo especies que presentaron una distribución muy amplia y no parecen estar relacionadas terminantemente con ninguno de los parámetros anteriores, a todas se les ha colectado tanto en fondos arenosos como fangosos y en diferentes profundidades (Gunter, 1945; Springer y Woodburn, 1960; Moore et al., 1970; Livingston, 1977), considerándoseles muy abundantes en el Golfo de México. Estas especies tienen hábitos pelágico-costeros y en su mayoría son eurihalinas. Opisthonema oglinum yHarengula pensacolae se alimentan de pequeños crustáceos e ictioplancton, se les encuentra frecuentemente juntas (Fisher, 1978), lo que también ha sido observado en este estudio; ambas presentaron una distribución muy semejante, con la mayor abundancia dentro de las estaciones de la Zona A y B respectivamente (Figs. 18 y 19).Lagocephalus laevigatus y Cynoscion arenarius son muy frecuentes en aguas costeras; sin embargo, poseen hábitos distintos. Se ha reportado que Lagocephalus laevigatus es una especie no gregaria que consume un gran número de grupos tróficos, mientras que Cynoscion arenarius tiende a formar cardúmenes relacionados con la crianza y alimentación cerca de ríos y estuarios y se alimenta principalmente de peces y crustáceos (Fisher, 1978). En este estudio, ambas especies presentaron una frecuencia muy alta (Tabla 6) y fueron más abundantes dentro de la Zona A y B respectivamente (Figs. 20 y 2l). 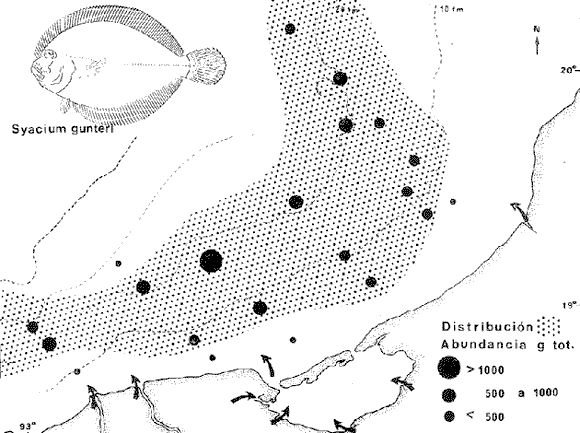 Fig. 12. Distribución y abundancia de: Syacium gunteri en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Como la distribución de todas estas especies está altamente relacionada con la dinámica ambiental del área de estudio, puede pensarse que el presente patrón de distribución varía en las diferentes épocas del año. ABUNDANCIADoce son las familias que se encuentran mejor representadas por sus valores de abundancia (biomasa) en las distintas zonas del área, y constituyen un alto porcentaje de la captura total (574 kg). Carangidae es la familia más abundante (29.9%) en la Zona A y conserva un alto porcentaje dentro de la Zona B (23.9%) donde la más representativa es la familia Ariidae (36.9%) (Tabla 7). A nivel de especie, el análisis de los valores de abundancia presenta un patrón muy semejante al de diversidad; sin embargo, el índice de biomasa (H'w) señala que no existe mucha diferencia entre las dos zonas (Tabla 5). Este índice otorga la misma importancia al número de gramos que al número de individuos, sin tomar en cuenta el tamaño de los mismos. Al relacionar estos parámetros, se encontró que la composición de las poblaciones, en términos de biomasa, también es distinta entre las dos zonas por la heterogeneidad de los individuos. 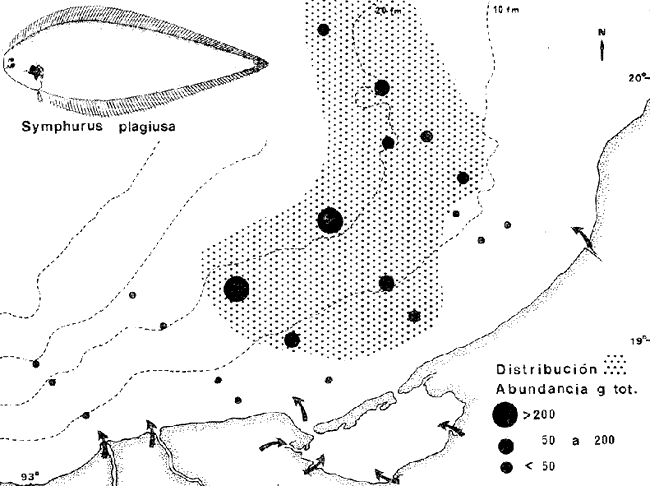 Fig. 13. Distribución y abundancia de: Symphurus plagiusa en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Los valores más altos de biomasa de la Zona A, los presentan las estaciones 3, 4, 5, 6 y 10. Todas estas estaciones excepto la 6 (18 m), se encuentran en el área de mayor profundidad de la Zona A (36-72 m). Los valores más bajos de biomasa se presentan en las estaciones 9, 14 y 23, que corresponden a las estaciones límite con la Zona B, independientemente de la profundidad (Figs. 22 y 26). Los valores más altos de biomasa de la Zona B, los presentan las estaciones 16, 17, 25 y 26. Estas estaciones con excepción de la 25 (18-36 m), es encuentran ocupando las profundidades más bajas de la Zona B y asociadas a comunidades de macroalgas y Thalassia testudinum. Los valores más bajos de esta zona, se encuentran en las estaciones 8 y 22, que resultan ser el límite entre las dos zonas, como sucede con los valores más bajos de la Zona A (Figs. 23 y 26). Estos datos indican que existe una relación importante entre la Profundidad del área de estudio y la distribución de la biomasa de las poblaciones, lo que ha sido observado por otros autores en comunidades de peces de otras localidades del Golfo de México (Moore et al ., 1964; Oviatt y Nixon, 1973; Smith, 1976). Moe y Martín (1965) in: Smith (1976) mencionan que la distribución de la biomasa relacionada con la profundidad, refleja el desplazamiento espacial de interacciones competitivas entre especies ecológicamente equivalentes. 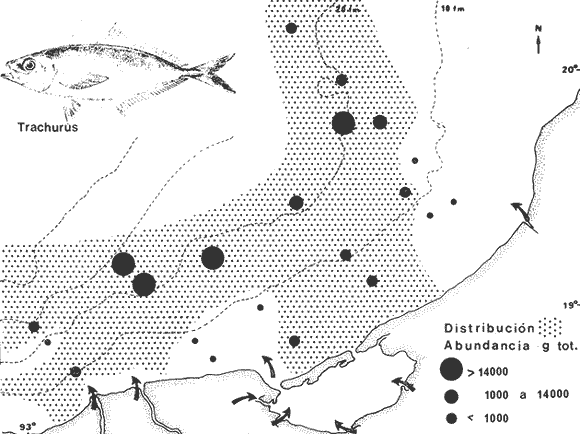 Fig. 14. Distribución y abundancia de: Trachurus lathami en la Sonda de Campeche. Crucero OPLAC-1, verano de 1978. Por otra parte, Chittenden y McEachran (1976), mencionan que los cambios sedimentarios en el Golfo de México, son un factor muy importante en la composición y estructura de las poblaciones de peces demersales, lo que en este estudio resulta ser muy claro, encontrándose que los valores más bajos de abundancia se presentan en las estaciones que unen a las dos provincias sedimentarias en el área de colecta. La distribución de la biomasa en la Zona A es directamente proporcional a la profundidad, pero en la Zona B es inversa. En ambos casos, los valores más bajos son más bien independientes de la profundidad (Fig. 26). El promedio de "standing crop" calculado para toda el área es de 0.8 g M-² (8.0 kg Ha-¹). Debe destacarse que las estaciones de menor profundidad de la Zona B presentan comunidades de Thalassia testudinum y esto está en relación directa con los valores más altos de biomasa de peces. INTERACCIONES DEL COMPLEJO ESTUARIO-PLATAFORMACon frecuencia se reporta la influencia del aporte fluvial como un factor determinante en la composición de la fauna y productividad de las áreas adyacentes. Gunter (1967) relaciona pesquerías en diferentes áreas del Golfo de México, con zonas estuarinas y de descarga de ríos. Mooreet al.(1970) reportan que la distribución de peces demersales frente a Louisiana y Texas está fuertemente relacionada con la descarga del río Mississippi, lo mismo que la captura de camarón y clupéidos (Stone, 1976). Sutcliffe (1972) muestra una correlación positiva entre los escurrimientos y la captura de cuatro especies de peces comercialmente importantes en el Golfo de St. Lawrence. Turner (1977) señala una correlación positiva para muchas localidades, entre las capturas comerciales de peneidos y las áreas de vegetación intermareal en zonas costeras protegidas. Stone et al.(1978) han presentado evidencias de que las pesquerías están relacionadas con áreas pantanosas intermareales. Yáñez-Arancibia et al. (1980) muestran que la producción de peces dentro de Laguna de Términos es mayor en los sistemas fluvio-lagunares que en las áreas de mayor influencia marina. Por los resultados de este estudio, parece que la influencia de la Laguna de Términos, es muy importante en la dinámica de la comunidad ictiológica de la plataforma adyacente aunque esto debe estar relacionado con los diferentes patrones de descarga. Esta información es la base para el desarrollo de nuevos estudios, que de manera cuantitativa al igual que este, describan los procesos que regulan esta interacción (Walne, 1972; Darnell y Soniat, 1979) y la dependencia biológica que significa la Laguna de Términos para muchas especies de la Sonda de Campeche. Conclusiones1. Las características ambientales de mayor importancia ecológica en la Sonda de Campeche (Tabla 1) dependen de la circulación litoral, los aportes fluviales y la transición de materiales terrígenos y sedimentos calcáreos de la región. Presentándose variaciones reguladas por la meteorología, climatología y oceanografía locales. 2. Los rangos estrechos de salinidades de 35 a 36%, y temperaturas de 27.7 y 28.3ºC, determina que estos parámetros no influyen determinantemente en la composición de las comunidades de peces demersales en el área, durante el periodo de verano. 3. Desde el punto de vista ambiental, la plataforma continental adyacente a Laguna de Términos presenta, en época de verano, dos habitat o subsistemas bien definidos en este estudio como Zonas A y B. 4. Existen no menos de 100 especies de peces demersales en el área de estudio. De las cuales el 46% pueden ser encontradas en alguna etapa de su vida dentro de la Laguna de Términos. 5. Se distinguen dos zonas bien marcadas de distribución de las poblaciones, altamente correlacionadas con las características de los habitat delimitados. Zonas A y B. 6. La familia Carangidae es la mejor representada en toda el área. Pero 15 especies de diversas familias son consideradas típicas de la comunidad por su distribución, frecuencia y abundancia. Éstas son: Arius felis, Cetengraulis edentulus, Cynoscion nothus., Diplectrum radiale, Priacanthus arenatus, Syacium gunteri, Symphurus plagiusa, Trachurus lathami, Synodus foetens, Chloroscombrus chrysurus, Eucinostomus gula, Opisthonema oglinum, Harengula pensacolae, Lagocephalus laevigatus yCynoscion arenarius. 7. La distribución y abundancia de las especies, está determinada por distintos patrones, siendo los más significativos, la composición y distribución de los sedimentos, la batimetría y la influencia de las aguas epicontinentales. 8. La diversidad (H') en las distintas colectas, varió de 0.8 a 2.4, promedio 2.8. La biomasa (H'w) varió de 1.1 a 2.5, promedio de 3.1. 9. El "standing crop" varió de 0.03 a 1.6, promedio de 0.8 g m-². Valores que son similares a los reportados en otras bahías someras del Golfo de México. El promedio de "standing crop" calculado para toda el área de 8.0 kg Ha-¹. 10. Los resultados de este estudio y la discusión planteada, permiten establecer que la Laguna de Términos y otros estuarios adyacentes tienen una influencia notable en la dinámica ambiental, y la estructura y función de las comunidades de peces de la plataforma. De las 15 especies que definen a las comunidades en cuestión, el 73% utilizan los sistemas estuarinos conectados a la Sonda de Campeche, en algún momento de su vida. AgradecimientosEste estudio fue auspiciado por el Instituto de Ciencias del Mar y Limnología de la UNAM, institución que proporcionó además el financiamiento y los medios necesarios para el análisis de laboratorio y el procesamiento de datos de computación. Se agradece especialmente a la Empresa Naviera Rex, S. A., de Ciudad del Carmen, Campeche, a través de su propietario Felice Re Sambruno, el facilitar el buque "La Nueva Ley de Pesca" para el trabajo de campo del Crucero OPLAC-I. Reconocimiento especial a Agustín Ayala-Castañares, Antonio García-Cubas, José Luis Rojas Galavíz y Alfredo Laguarda Figueras por la revisión crítica del manuscrito haciendo atinadas y valiosas sugerencias. Por esto último se le agradece también a John W. Day de Louisiana State University y C. B. Subrahmanyam de Florida A & M University la discusión de los resultados. Al personal del laboratorio de Ictiología y Ecología Estuarina, por sus discusiones a la interpretación de los resultados en distintas etapas de su desarrollo y la ayuda en algunos problemas taxonómicos, especialmente a Ana Laura Lara Domínguez, Irma Vargas Maldonado y Margarito Álvarez Rubio. A. Arturo Gamietea y Gilberto Montelongo por su valiosa cooperación en la elaboración del programa de computación para el procesamiento de la información obtenida. LITERATURAALLEN, D. M J. E. TASHIRO, Proceedings: Colloquium on Snaper-Grouper Fishery Resources of the Western Central Atlantic Ocean. Status of the commercial snaper grouper fishery. In: Bullis, H. R. y A. C. Jones (Eds.). Florida Sea Gran Program Rep., 1976. 41-76.17: AMEZCUA-LINARES, F A. YÁÑEZ-ARANCIBIA, An.Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de los sistemas fluvio-lagunares asociados a la Laguna de Términos. El habitat y estructura de las comunidades de peces. 1980.69-118. 7 (1): ANDERSON, W. W., Fishes taken during shrimp trawling along the south Atlantic coast of the United States, U.S. Fish Wildl. Serv., Spec. Sci. Rep. Fish., 1968. 1-60.570: 1931-1935. BEAMISH, F. W. H., Vertical migration by demersal fish in the north west Atlantic. J. Fish. Res. Bd. Can., 1966.109-139. 23: BECHTEL, T. J B. J. COPELAND, Fish species diversity indices as indicators of pollution in Galveston Bay, Texas. Contr. Mar. Sci., 1970.103-132. 15: BRAVO-NÚÑEZ, E. A. YÁÑEZ-ARANCIBIA, An. Centro Cienc. del Mar y Limnol.Univ. Nal. Autón. México,Ecología de la Boca de Puerto Real, Laguna de Términos. 1. Descripción del área y análisis estructural de las comunidades de peces.1979.125-182.6 (1): BULLIS, H. R. J. S. CARPFNTER, The Future of the Fishing Industry of the United States. Latent fishery resources of the central west Atlantic region. In: De Witt, Gilbert (Ed.). Unív. of Washington Publ. Fish. New Ser., 1.1968. CARRANZA, J., Los recursos naturales del sureste y su aprovechamiento. Inst. Mex. Rec. Nat. Renov., 1969. 1-151-238.3 (2): CERVIGON, F., Los Peces Marinos de Venezuela. Estación de investigaciones marinas de Margarita. Fundación La Salle de Ciencias Naturales.Monogr. Caracas, 1966. 1-951, 385 figs. 1 y 2: CHÁVEZ, E. A., Mem. IV Congr. Nac. Ocean. Notas acerca de la ictiofauna del estuario del Río Tuxpan y sus relaciones con la temperatura y la salinidad. In: Carranza, J. (Ed.). (México),1972. 177-199 Nov. 17-19, 1969: CHÁVEZ, H., Acta Zool. Mem. Pobl. Mus. His. Nat. Peces colectados en el arrecife Triángulo Oeste y en Cayo Arcas, Sonda de Campeche,Cd. México1966. CHÁVEZ, H. J. ARVIZU, Mem. IV Congr. Nac. Ocean. Estudio de los recursos pesqueros demersales del Golfo de California, 1968-1969. III. Fauna de acompañamiento del camarón (peces finos y "basura"). In: Carranza, J. (Ed.). (México), 1972.361-378. Nov. 17-19, 1969: CHITTENDEN, M. E. J. D. McEACHRAN Composition, ecology and dynamic of demersal fish communities in the Northwestern Gulf of Mexico. Texas A&M University Press. Sea Grant, 1976. 1-104.76 (208): CLARKE, J. R., International Oceanography Congress Seasonal changes in abundance within a community of demersal fishes. 1959. 331-332. CLARO, R., D. V. RADAKOV Y. S. RESHETNIKOV A. SILVA Algunas características de la ictiofauna de la Plataforma Cubana. Academia de Ciencias de Cuba, Serie Oceanológica, Instituto de Oceanologia, 1974.1-10. 20: DANIELS, K., Proc. Third Coastal Marsh and Estuary Management Symposium. Habitat designation based on cluster analysis of Ichtyofauna. In: Day, J. W., D. D. Culley, R. E. Turner y A. J. Murphrey (Eds.). Louisiana State University Division of Counting Education, Baton Rouge, La. 1979. 317-324. DARNELL, R. M. SONIAT, T. M Ecological Processes in Coastal and Marine Systems. Marine Science The estuary continental shelf as an interactive system. In: Robert J. Livingstone . (Ed.). Plenum Press, New York1979.489-525. 10: DAVIES, R. G., Computer Programing in Cuantitative Biology. Academic Press Inc., London,1971. 492 p. EMILSSON, I,Reunión Internacional de Trabajo COI/OAA/PNUMA Sobre Contaminación Marina en el Caribe y Regiones Adyacentes., La oceanografía regional con respecto a los problemas actuales y futuros de la contaminación y de los recursos vivos, Golfo de México. 1976.1-24. 13-18 Dic., 1976 ERDMAN, D. S., Symposium on Progress in Marine Research in the Caribbean and Adjacent Regions. Caracas, Venezuela, Papers on Fisheries, Aquaculture and Marine Biology. Spawing patterns of fish from the northeastern Caribbean. In: Stewart, H. B. (Ed.) FAO Fish. Rep., 1977. 145-169.200: 12-16 July 1976. FISHER, W. FAO species identification sheets for fishery purposes. Western Central Atlantic (fishing area 31). FISHER, W. (Ed.), FAO, Roma,1978. 1-7. FRANKS, J. S., J. CHRISTMAS, W. L. SILER R. COMBS R. WALLER C. BURNS A study of the nektonic and bentic faunas of the shallow Gulf of Mexico off the state of Mississippi. Gulf Res. Rep.,1972.1-148. 1: GARCÍA, E., Modificaciones al Sistema de Clasificación Cilmática de Köppen (para adaptarlo a las condiciones de la República Mexicana). Univ. Nal. Autón. México, Instituto de Geografía, 1973. 246 p. GUNTER, G., Publ. Inst. Mar. Sci. Univ. Texas, Studies on marine fishes of Texas. 1945.1-190. , 1: _____, Estuaries, Am. Ass. Adv. Sci., Publ, Spec., Some relationships of estuaries to the fisheries of the Gulf of Mexico. In: Lauff, G. A. (Ed.). 1967.621-638. 83: GUTIÉRREZ-ESTRADA, M., Tesis M. en C. Sedimentología del área de transición entre las provincias terrígena y carbonatada del suroeste del Golfo de México. Fac. Ciencias. Univ. Nal. Autón. México, 1977. 175 p. GREENWOOD, P. H. D. E. ROSEN S. H. WEITZMANN G. S. MYERS, Bull. Am. Mus. Nat. Hist., Phyletic studies of teleostean fishes, with a provisional classification of living forms. 1966. 339-456.131 (4): GREENWOOD, P. H. G. S. MYERS D. E. ROSEN S. H. WEITZMANN, Named main divisions of teleostean fishes. Proc. Biol. Soc. Wash., 1967.227-228. 80: HILDBRAND,H. H., Publ. Inst. Mar. Sci.,Texas,A study of the fauna of the pink shrimp (Penaeus duorarum Burkenroad) ground in the Gulf of Campeche. 1955. 169-232. 1: HOESE, H. D. B. J. COPELAND F. N. MOSELEY E. D. LANE, Fauna of the Aransas Pass Inlet, Texas. III. Diel and seasonal variations in trawlable organisms of the adjacent arca. Texas J. Sci., 1968. 20:33-60. HOESE, H. D. R. H. MOORE, Fishes of the Gulf of Mexico (Texas, Louisiana and Adjacent Waters). Texas A&M University Press. 1977. 376 p. HORN, M. H. L. G. ALLEN, Bull. South Cal. Acad. Sci., Numbers of species and faunal resemblance of marine fishes in California bays and estuaries. 1976. 159-170.75 (2): JORDAN, D. S. B. W. EVERMANN, Bull. U.S. Nat. Mus., The fishes of north and midle America. 1896-1900.1-3313, 398 láms., 958 figs.1-4 (47): KLIMA, E. F., CICAR II Symposium Progress in Marine Research in the Caribbean and Adiacent Regions , An assessment of the fish stocks and fisheries of the Campeche Bank. UNESCO-FAO. WECAF Stud.1976a. 1-24. 5: _____, Proceedings: Colloquium on Snaper-Grouper Fishery Resources ot fhe Western Central Atlantic Ocean.Snaper and grouper resources of the Western Central Atlantic Ocean. In: Bullis, H. R. y A. C. Jones (Eds.). Florida Sea Grant Program Rep., 1976b.540. 17: _____, Symposium on Progress in Marine Research in the Caribbean and Adjacent Regions. An overview of the fishery resources of the Western Central Atlantic Region. In: Stewart, H. B. (Ed.). Papers on Fisheries. Aquaculture and Marine Biology. FAO Fish. Rep., CaracasVenezuela,1977. 231-252. 200: 12-16 July 1976. LICEA-DURÁN, S., Symposium on Progress in Marine Research in the Caribbean and Adjacent Regions. Caracas Venezuela, Papers on Fisheries Aquaculture and Marine Biology. Variación estacional del fitoplancton de la Bahía de Campeche, México In: Stewart, H. B. (Ed.). FAO Fish. Rep., 1977.253-273. 200:12-16 July 1976.(1971-1972). LIVINGSTON, R. J. P. S. SHERIDAN B. G. MCLANE F. G. LEWIS, III ,G. G. KOBYLINSKI, Proceedings of the Conference on the Apalachicola Drainage System. The biota of Apalachicola Bay system functional relationships. In: Livingston, R. J. y E. A. Joyce, Jr. (Eds.) Fla Mar. Res. Publ., Gainesville Florida. 1977. 75-100. (26): 23-24 April 1976, MACHADO, A., F. LEY LOU, V. M. ALBA R. CRUZ, Características texturales, pH y porcentajes de materia orgánica, humedad y minerales ligeros de los sedimentos obtenidos durante el Crucero OPLAC-1 del CCML.Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México. (Rep. Téc. Inédito), 1979. 22 p. MANCILLA-PERAZA, M. M. VARGAS-FLORES, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Los primeros estudios sobre el flujo neto de agua a través de la Laguna de Términos, Campeche. 1980. 142.7 (2): MARGALEF, R., Perspectives in Ecological Theory. The University of Chicago Press,Chicago, 1969. 111 p. MEEK, E. S. S. F. HILDEBRAND, The marine fishes of Panama. Field. Mus. Nat. Hist. Publ. Zool. Ser., 1923-1928. 1-1045, 118 láms.15 (215, 226 y 249): MERRIMAN, D H. E. WARFEL, VII. Analysis of fish population. Bull.Studies on the marine resources of southern New England,Bingham Oceanogr. Coll., 1948.131-164.2 (4): MILLER, J. S., A trawl study of the shallow Gulf fishes near Port Aransas, Publ. Inst. Mar. Sci. Univ. Texas.,Texas. 1965. 80-107.10: MOE, M. A. G. T. MARTIN Fishes taken in northly trawl samples offshore of Pinellas County, Florida, with new additions to the fish fauna of Tampa Bay area. Tulane Stud. Zool., 1965.129-151. 12: MOORE, D H. A. BRUSHER L. TRENTRelative abundance seasonal distribution and species composition of demersal fishes off Louisiana and Texas, Contr. Mar. Sci., 1970. 45-70.15: 1962-1964. MOORE, H. R., Variatíon in the diversity of summer estuarine fish populations in Aransas Bay, Texas, Estuar. Coast. mar. Sci., 1978.495-501. 6:1966-1973. NORMAN, J. R. The trustees of the British Museum (Natural Hístory), A Systematic Monography of the Flatfishes (Heterosomata). Psettodidae, Bothidae, Pleuronectidae. London, 1934. 1-459, 317 figs. 1: OVIATT, C. A. S. W. NIXON, The demersal fish of Narragansett Bay: an analysis of community structure, distribution and abundance. Estuar. Coast. mar. Sci., 1973. 361-378. 1: PIELOU, E. C., J. Theoret. Biol., The measurement of diversity in different types of biological collections. 1966.131-144. 13: RAMÍREZ - HERNÁNDEZ, E. J. ARVIZU, An. Inst. Nal. Inv. Biol.-Pesq., Investigaciones ictiológicas en las costas de Baja California. I.México, 1965.293-324. 1: RAMÍREZ-HERNÁNDEZ, E. J. PÁEz, An. Inst. Nal. Inv. Biol.Pesq. Investigaciones ictiológicas en las costas de Guerrero. I. México 1965.327-358. 1: RAMÍREZ - HERNÄNDEZ, E. G. CARRILLO D. LLUCH, Investigaciones ictiológicas en las costas de Chiapas. Lista de peces colectados en las capturas camaroneras Secretaría de Industria y Comercio, Dir. Gral. Pes. Ind. Conn. México, Publ. Inst. Nal. Inv. Biol.-Pesq.,1964.147. 5:(agosto-septiembre, 1959, abril-mayo-junio, 1960). RAMÍREZ-HERNÁNDEZ, E. N. VÁZQUEZ, R. MÁRQUEZ C. GUERRA, Investigaciones ictiológicas en las costas de Sinaloa. I. Lista de peces colectados en las capturas camaroneras Secretaría de Industria y Comercio. Dir. Gral. Pes. Ind. Conn México, Publ. Inst. Nal. Inv. Biol.-Pesq., 1965.1-36. 12:(agosto, 1961; abril-octubre, 1962; mayo-septiembre, 1963). RANDALL, J. E., Food habits of reef fishes of the west indies. Stud. Trop. Oceanogr. Miami, 1967.1-847. 5: RESÉNDEZ, A., An. Inst. Biol. Univ. Nal. Autón. México. Ser. Cienc. del Mar y Limnol., Peces colectados en el arrecife La Blanquilla, Veracruz, México. 1971.7-30. 42 ( 1): RICHARDS, S. W., Bull. Bingham Oceanogr. Coll.,The demersal fish population of Long Island Sound. 1963. 5-101.18 (2): ROITMAYR, C. M., Industrial bottomfish fishery of the Northern Gulf of Mexico. Spec. Scient. Rep. U.S. Fish. Wildl. Serv., 1965.1-23. 518:1959-1963. SANTOYO, H. M. SIGNORET, Hidrología y fitoplancton en un transecto en la plataforma continental de la Bahía de Campeche, Rev. Lat-amer. Microbiol.,México.1972.207-215. 15:(Agosto, 1972), SAUSKAN, V. I. A. OLACHEA, Ictiofauna bentónica del banco de Campeche. Resum. Invest. Inst. Nal. Pesca Ant. Invest. Pesq. Cuba,1974.102-106. 1: SCHULTZ-RUIZ, L. E. E. A. CHÁVEZ, Mem. Simp. Sobre Biol. y Dinám. Pobl. Camarones. Guaymas, Son., Contribución al conocimiento de la biología pesquera del camarón blanco (Penaeus setiferus) del Golfo de Campeche, México. México. 1976.57-73. 8-13. Ago. 1976: SHANNON , E. C. W. WEAVER, The Mathematical Theory of Communication. University of Illinois Press. Urbana,1963. 119 p. SMITH, G. B., Ecology and distribution of eastern Gulf of Mexico reef fishes. Fía. Mar. Res. Publ., 1976.1-78. 19: SPRINGER, V. G. K. D. WOODBURN, An ecological study of the fishes of the Tampa Bay area. Fla. St. Bd. Prof. Pap. Serv., 1960. 1-104. 1: STONE, J. H., Environmental factors related to Louisiana menhaden harvest. Louisiana State Un¡versity, Center for Wetland Resources, Baton Rouge, La. Final Rept. U.S. Dept. Comm., NOAA, Nat. Mar. Fish. Serv., Fla. St. Petersburg1976. STONE, J. H. J. W. DAY Jr. L. M. BAHR Jr. R. A. MULLER, Estuarine Interactions. The impact of possible climatic changes on estuarine ecosystems. In: M. L. Wiley (Ed.). Academic Press Inc. New York1978.305-322. SUBRAHMANYAM, C. B. S. H. DRAKE, Bull. Mar. Sci., Studies on the animal communities, in two north Florida salt marshes. Part 1. Fish communities. 1975.445-465.25 (4): SUTCLIFFE, W. H., Some relations of land drainage nutrients, particulate material, and fish catch in two eastern Canadian bays. J. Fish. Res. Bd. Canada,. 1972. 357-362.29 (4): THOMAS, J. P. WAGNER H. LOESCH, Coast. Stud. Bull., Studies of the fishes of Barataria Bay, Louisiana, an estuarine community. 1971.1-68. 6: TOPP, R . W. F. H. HOFF Jr. Flatfishes (Pleuronectiformes). Fla. Dep. Nat. Resourc. Mem. Hourglass Cruises., 1972.1-135.4 (2): TURNER, R. E., Intertidal vegetation and commercial yields of Penaeid shrimps. Trans. Am., Fish. Soc., 1977.411-416.106 (5): VARGAS MALDONADO, I. A. YÁÑEZ-ARANCIBIA F. AMÉZCUA- LINARES An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología y estructura de las comunidades de peces en áreas de Rhizophora mangle y Thalassia testudinum de la Isla del Carmen, Laguna de Términos, Sur del Golfo de México. 1981.241-266.8 (1): VILLALOBOS, A. y M. E. ZAMORA, Importancia biológica de la Bahía de Campeche. Mem. I. Simp. Latinoam. Oceanogr., Biol. (México),1975.375-394. 25-29 Nov. 1974. _____, Importancia biológica de la Bahía de Campeac y de la Península de Yucatán (Segunda parte). Mem. II. Simp. Latinoam. Oceanogr. Biol. Cumaná Venezuela,Publ. Univ. Oriente. 1977. 79-117. Nov. 24-28, 1975. WALNE, P. R., The Estuarine Environment. The importance of estuaries to commercial fisheries. In: Barnes, R. S. K. y J. Green (Eds.) Applied Science Publishers Ltd., London. 1972.107-118. WILHM, J. L., Ecology, Use of biomass in Shannon's formula. 1968.153-156.49 (1): YÁÑEZ-ARANCIBIA, A., Ecology in the entrance of Puerto Real, Terminos Lagoon. II. Discusion on trophic structure of fish communities in banks of Thalassia testudinum. In: Lasserre, P., H. Postma, J. Costlow y M. Steyert (Eds.). Coastal Lagoon Research: Present and Future. II. Proceedings: UNESCO/ IABO Seminar, Duke Univ. Mar. Lab. Sept., 1978 (in press). 1978. YÁÑEZ-ARANCIBIA, A. F. AMÉZCUA - LINARES, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de la raya Urolophus jamaisensis (Cuvier). En Laguna de Términos, un sistema estuarino del Sur del Golfo de México (Pisces: Urolophidae).1979.123-136.6 (2): YÁÑEZ-ARANCIBIA, A. J. W. DAY, JR., Ecological characterization of Terminos Lagoon a tropical lagoon-estuarine system in the southern Gulf of Mexico. Proc. International Symposium on Coastal Lagoon, ISCOL-UNESCO, Bordeaux France, Oceanologica Acta.1981. Sept. 7-14, 1981. YÁÑEZ-ARANCIBIA, A. F. AMÉZCUA-LINARES J. W. DAY Estuarine Perspectives. Fish community structure and function in Terminos Lagoon, a tropical estuary in Southern Gulf of Mexico. In: Kennedy, V. (Ed.). Academic Press Inc., New York.1980.465-482.
|