|
FORAMINÍFEROS RECIENTES DE LA LAGUNA DE
TAMIAHUA, VERACRUZ, MÉXICO
Trabajo recibido el 8 de mayo de 1980 y
aceptado para su publicación el 5 de septiembre de
1980.
AGUSTÍN AYALA-CASTAÑARES
LUIS R. SEGURA
Instituto de Ciencias del Mar y
Limnología. Universidad Nacional Autónoma de México.
Contribución 223 del Instituto de Ciencias del Mar y Limnología,
UNAM.
La Laguna de Tamiahua está situada en el litoral del
Golfo de México, Estado de Veracruz, entre los ríos Pánuco
y Tuxpan. Presenta los siguientes rasgos fisiográficos: a) La existencia
de numerosos arroyos, en su mayoría de flujo estacional; b) varias
islas, las principales son la Del Toro, Juana Ramírez y Del Idolo; c)
una extensa barrera arenosa (Cabo Rojo); d) canales que la conectan con los
ríos Pánuco y Tuxpan; e) una boca angosta en la porción
sur; y f) rasgos morfológicos que indican la evidencia de antiguas
bocas.
El clima en la región es subhúmedo, con
evaporación moderada, fuertes lluvias y seco en el invierno, con
excepción de las tormentas con vientos del N: en verano prevalecen
vientos del E y en invierno del N y NE.
Su forma es alargada
y tiene una profundidad media de 2 a 3 m. La barrera arenosa de Cabo Rojo es de
forma angular y está constituida principalmente por arenas
cuarcíticas; en su porción norte hay grandes dunas activas que
disminuyen hacia el S. Existen dos sistemas de bermas truncadas entre
sí, y en sotavento un crecimiento local por los
manglares.
El agua es predominantemente salobre;
distinguiéndose dos facies, una ultrahalina con salinidad mayor de 30%,
y una polihalina con salinidad entre 16 y 30%. La primera restringida a la boca
y áreas vecinas, afectadas directamente por las mareas, en tanto que el
resto es polihalino.
Frente a la costa se encuentra una
cadena de arrecifes coralinos vivos; en la barrera de Cabo Rojo existe, en el
subsuelo, un arrecife coralino muerto y en el fondo de la laguna otro
más, también muerto. Estos arrecifes parecen tener especial
importancia en el origen de la barrera arenosa y en el desarrollo
geológico de la laguna.
Se colectaron 65 muestras
tanto para el estudio de los foraminíferos como de los sedimentos. Estos
en general son principalmente limo-arcillosos, habiéndose distinguido 5
grupos texturales: I arenas finas, entre bien y moderadamente clasificadas; II
arenas muy finas de transición pobremente clasificadas; III limos
pobremente clasificados; IV arcillas muy pobremente clasificadas; y V arcillas
pobremente clasificadas.
Las poblaciones totales de
foraminíferos varían entre 14 y 17,600 ejemplares por muestra.
Las poblaciones vivientes están representadas principalmente por los
géneros Ammonia y Elphidium. Se
determinaron 42 géneros y 70 especies.
En base a las
asociaciones de foraminíferos pudieron establecerse 3 biofacies: a) de
esteros y ríos; b) lagunar y c) de golfo abierto. En general reflejan
las condiciones hidrográficas y ecológicas de la región.
La productividad orgánica relativa de la laguna fue en el momento del
muestreo, más bien moderada o baja; la tasa de sedimentación de
la laguna parece moderada.
The "Laguna de
Tamiahua" is located in the littoral of the Gulf of Mexico between the Panuco
and Tuxpan rivers in the state of Veracruz. The main physiographic features of
the lagoon are:
a) The existance of various streams, the
majority only with seasonal flow; b) various islands, the principal of these
being Del Toro, Juana Ramirez and Del Idolo; c) an extensive sand barrier (Cabo
Rojo); d) connecting channels wiht the Panuco and Tuxpan rivers; e) a narrow
inlet in the southern portion; and f) morphological features indicating the
evidence of ancient inlets.
The climate in the region is
sub-humid with moderate evaporation, heavy rainfall and dry during the winter
with the excepction of storms associated with northerly winds. and 30%. The
first one is restricted on the inlet and surrounding areas directly affected
and north-east.
The lagoon is long and has an average depth
between 2 and 3 meters. The sand barrier of "Cabo Rojo" is of angular form and
is composed mainly of quartzitic sands; in the northern portion of the barrier
there are largo active dunes, decreasing in size towards the south. Two systems
of berms are found which truncate one another and on the seward side of the
barrier there is a local increase due to mangrove growth. The lagoon water is
predominantly brackish with two well defined water masses, one ultrahaline with
salinity above 30%, and the other polyhaline wtih salinity between 16 and 30%.
The first one is restricted to the inlet and surroundin areas directly affected
by the tides; the rest is pholyhaline.
Parallel to the coast
there is a chain of live coral reefs; in the barrier of "Cabo Rojo", in the
sursurface there is a dead coral reef and in the lagoon's bottom another dead
coral reef is located. These reefs seem to have special importance in the
origin of the sand barrier and the geological development of the
lagoon.
65 samples were collected for the study of
foraminifera and sediments. The majority of these samples are silty clays can
be five textural groups: I fine sands, between well and moderately sorted; II
very fine transition sands, poorly sorted; III poorly sorted silt; IV very
poorly sorted clays; and V poorly sorted clays.
The total
populations of foraminifera were between 14 and 17,600 specimens per sample.
The living populations were represented mainly by the
generaAmmonia and Elphidium. A total of
42 genera and 70 species was determined.
On the basis of the
associations of foraminifera, it was possible to establish 3 biofacies: a)
"esteros" and rivers; b) lagoonal; and c) open gulf. These biofacies generally
reflect the hydrographic and ecological conditions of the region. The relative
organic productivity of the lagoon was, during the time of the study, moderate
or low. The rate of sedimentation in the lagoon seems to be
moderate.
Este trabajo es otra contribución al conocimiento de la ecología y la distribución de la fauna de foraminíferos de las lagunas litorales del Golfo de México; con anterioridad se han pupublicado trabajos en áreas cercanas (Ayala-Castañares y Segura, 1968) y (Segura, 1963). Forma parte de un estudio más amplio del área, donde también se han estudiado la geología marina (Cruz, 1968) y los micromoluscos (García-Cubas Jr., 1969). Un resumen de todos estos estudios fue presentado en el Segundo Congreso Internacional de Oceanografía, en Moscú, 1966 por A. Ayala-Castañares y en el Simposio Internacional sobre Lagunas Costeras (Ayala-Castañares, Cruz, García-Cubas Jr. y Segura, 1969). El objetivo fundamental de este estudio fue determinar las poblaciones de foraminíferos en los sedimentos superficiales de la Laguna de Tamiahua, con un criterio sedimentológico. Se identifican las especies, su abundancia y los factores ecológicos de la laguna, para tratar de definir hasta que grado la distribución de los foraminíferos refleja las condiciones ecológicas y de sedimentación. La metodología aplicada es similar a la empleada en la Laguna de Términos, Camp. (Ayala-Castañares, 1963) y la Laguna Madre, Tamps., (Ayala-Castañares y Segura, 1968). Los foraminíferos fueron considerados como elementos constitutivos de los sedimentos y procesados cualitativa y cuantitativamente; los datos referentes a la distribución de los foraminíferos se correlacionaron de acuerdo con la información disponible sobre los parámetros ambientales. Los estudios de laboratorio se efectuaron inicialmente en el desaparecido Departamento de Micropaleontología y Ciencias Marinas del Instituto de Geología y se terminaron en el Laboratorio de Ecología de Foraminíferos y Micropaleontología del Centro de Ciencias del Mar y Linmología, ambos de la Universidad Nacional Autónoma de México. Las muestras empleadas para el estudio fueron colectadas por A. Yáñez y R. Cruz, en 1964. Ejemplares representativos de las especies se encuentran depositados en la colección micropaleontológica del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México (CCML- Mi.) ANTECEDENTESA pesar de las dimensiones y de su interés científico la Laguna de Tamiahua ha sido poco estudiada, pudiendo decirse que entre las primeras investigaciones realizadas en esa área se cuentan las de carácter geográfico-descriptivo, hechas en el siglo pasado por Núñez-Ortega (1869) y García Cubas (1891); en el presente siglo, con el mismo carácter, los trabajos de Muñoz-Lumbier (1919), Tamayo (1949-1962), aunque se ha hecho énfasis principalmente en las disciplinas de tipo biológico, siendo de mencionarse entre los trabajos hasta ahora publicados los estudios sobre moluscos hechos por De Buen (1957), Sevilla y Mondragón (1965) y García-Cubas, Jr (1969); sobre vegetación, tanto sumergida como circundante: Humm y Hildebrand (1962), Poggie (1962) y González Medrano (1966); sobre peces por Rosen (1960, Ramírez (1965), Álvarez (1970) y Reséndez (1970); las de carácter geológico, Muir (1936), Benavides (1956), Cruz (1968), Mac Intire y Ho (1969), Ayala-Castañares, Cruz, García-Cubas, Jr. y Segura (1969). Existen además trabajos realizados por A. Villalobos y colaboradores en 1968 (inédito), sobre los efectos de perforación de pozos petroleros de exploración en la ecología de la laguna, así como los de H. Bravo, S. Salazár, A. Vázquez Botello y E. Mandelli (1978) sobre la concentración de hidrocarburos aromáticos polinucleares en las aguas de la laguna. Muchas de estas investigaciones han sido realizadas por personal de los Institutos de Biología, Geología y del Instituto de Ciencias del Mar y Limnología, de la Universidad Nacional Autónoma de México. En la actualidad, otras instituciones tales como el Instituto Nacional de Pesca, del Departamento de Pesca, tienen programas permanentes en la región. DESCRIPCIÓN GENERAL DE LA LAGUNA DE TAMIAHUA, VERACRUZ
UBICACIÓNLa Laguna de Tamiahua se encuentra en el Estado de Veracruz, entre los ríos Pánuco al N y Tuxpan al S, con los que se comunica por canales (Fig. l). Está situada en la Cuenca Terciaria de Tampico-Misantla; en su margen continental existen lomeríos de sedimentos terciarios y cuaternarios, con excepción de la Sierra de Tantima, que es volcánica. FISIOGRAFÍALa laguna es de forma irregular, alargada en sentido norte-sur; en su borde continental desembocan varios ríos, en su mayor parte de flujo estacional, entre los que sobresalen La Laja, Cucharas, Tancochin y Tampache, que en épocas de lluvia aportan grandes cantidades de sedimentos especialmente limo-arcillosos producto de la erosión de rocas terciarias de la llanura costera; algunos de esos ríos forman pequeños deltas en su desembocadura. El borde oriental presenta numerosas entrantes y salientes, con algunos manglares. En el centro de la laguna, cerca de la Isla Del Toro, existe un arrecife coralino muerto. 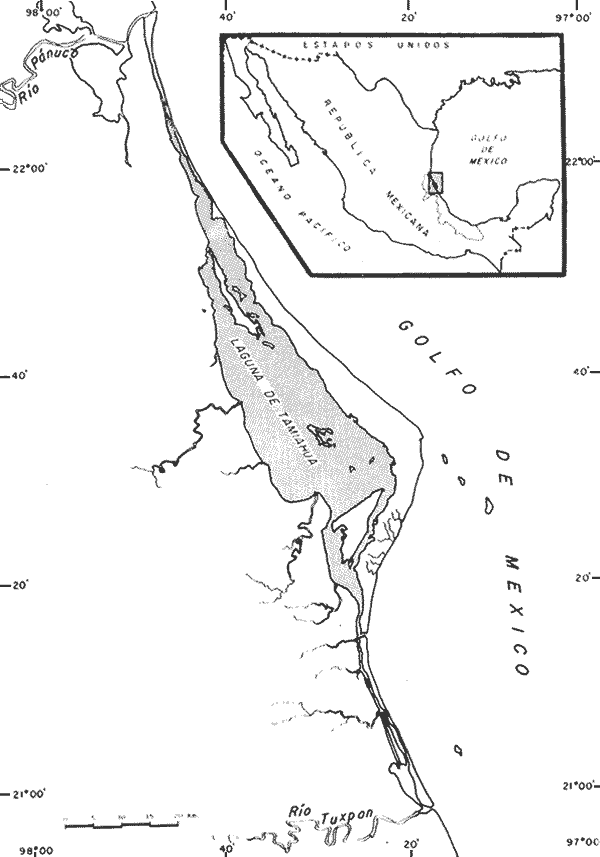 Fig. 1. Mapa de localización de la Laguna de Tamiahua, Ver. En el interior de la laguna (Fig. 2) se localizan varias islas, siendo las principales Juana Ramírez, Del Toro y Del Idolo. Está separada del Golfo de México por una barrera arenosa de forma angular denominada Cabo Rojo y presenta una boca en su extremo sur, llamada Boca de Corazones. En las islas Juana Ramírez y Del Toro hay antiguas dunas estabilizadas, que probablemente representan vestigios de una antigua barrera arenosa. En la isla Del Idolo, en cambio, sólo se observan cordones de playa y antiguos meandros de ríos. La barrera arenosa de Cabo Rojo es de forma angular con vértice hacia el E, longitud máxima de 130 km; anchura máxima de 6 km. y mínima de 500 m. Interrumpe la línea de costa, para proyectarse aproximadamente a 15 km. dentro del Gofo de México. Se distinguen dos porciones: a) una parte norte entre el río Pánuco y los 21º33'N que sigue una dirección NW-SE, con grandes dunas activas en barlovento, varias bermas paralelas a la playa y en sotavento dunas estabilizadoras; y b) una parte sur, entre los 21º33'N y la Boca de Corazones, con rumbo NE-SW, con dunas de menor altura y bermas paralelas a la costa que truncan a las de la porción N. Está compuesta por arenas cuarcíticas, salvo algunos lugares de sotavento, donde existen manglares en los que se encuentran sedimentos finos. En el subsuelo, a una profundidad de 12.5 m donde convergen las dos porciones, existe un antiguo arrecife coralino. En el Golfo de México, frente a la barrera arenosa, se halla una cadena de arrecifes coralinos vivos alineados con el del subsuelo de Cabo Rojo y con el del fondo de la laguna. Todos estos arrecifes parecen tener especial importancia en el origen y la formación de dos barreras arenosas: una antigua, representada actualmente por parte de las islas Juana Ramírez y Del Toro, y la actual (Cabo Rojo). Los arrecifes vivos podrían servir en el futuro de basamento a la formación de barreras arenosas. BOCA DE CORAZONESLa única comunicación con el mar, denominada Boca de Corazones es angosta e inestable y tiende a emigrar hacia el S; su canal más profundo es de aproximadamente 1.5 m. Hay evidencias morfológicas de la existencia de otras tres bocas, una en la porción norte de Cabo Rojo y las dos restantes al S de la Boca de Corazones. CLIMAEl clima es subhúmedo (García, 1970), lluvioso en verano y seco en invierno, modificado por las frecuentes tormentas o "nortes" que se presentan con fuertes vientos en esa misma dirección; durante el verano, prevalecen vientos del E. La evaporación es moderada. VEGETACIÓN CIRCUNDANTELos manglares están ampliamente distribuidos en los bordes de la laguna, observándose las siguientes especies: Rizophora mangle, Avicennia germinans, Laguncularia racemosa y Conocarpus erectus.Además, existen palmeras, selva mediana y una vegetación pionera a lo largo de las playas, representada por las especies Ipomoea littoralis, Croton cortesianus, Coccoloba uvifera, Randia echinocarpa (Poggie, 1962 y González Medrano, 1966). BATIMETRÍATiene una profundidad de 2 a 3 m (Cruz, 1968). El fondo de la parte occidental media tiene una pendiente más suave que el de la parte oriental, donde hay un canal más profundo, mantenido por la acción de las mareas y en parte por dragados (Fig. 3). MAREAS Y CIRCULACIÓNSe desconoce en detalle el régimen de mareas, pero comparándolo con la información sobre Tampico, Tamps. y Tuxpan, Ver., puede extrapolarse que es diurno y su rango muy pequeño. Asimismo, se desconocen las características de la circulación del agua, aunque los principales factores que la afectan son las mareas, el viento y las descargas de los ríos (Cruz, 1966). 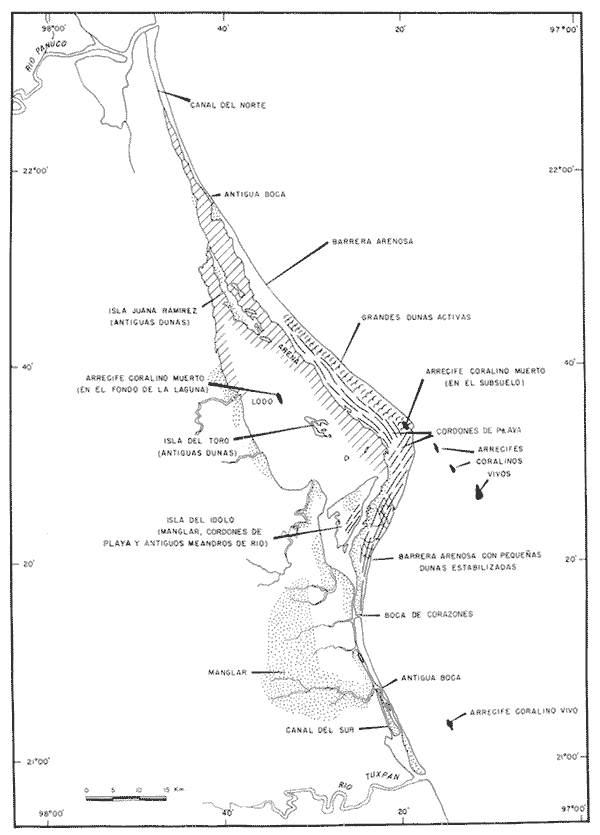 Fig. 2. Fisiografía generalizada (según Cruz, 1968). 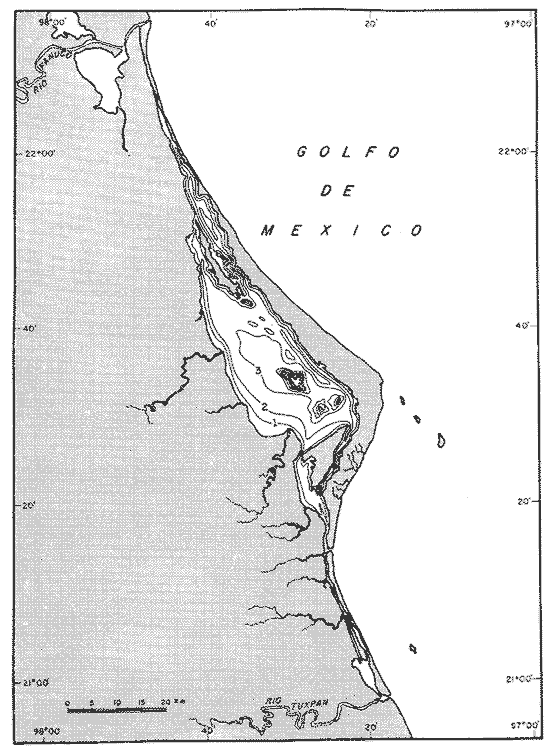 Fig. 3. Batimetría. Isóbatas en metros (según, Cruz, 1968). SALINIDADLa salinidad del agua de la laguna tanto en la superficie como en el fondo, indica dos facies: una polihalina entre 16 y 30% y otra, ultrahalina mayor de 30%, (Cruz, 1968). La facies ultrahalina se distribuye en la porción sur de la laguna, afectada por las mareas que penetran en la Boca de Corazones. La salinidad en el resto de las aguas de la laguna es polihalina (Fig. 4). TEMPERATURALa temperatura del agua de la superficie y el fondo; en términos generales se comportan de manera similar. Las temperaturas más bajas (23.1ºC), se encontraron en el área cercana a la boca; en el resto de la laguna oscilaron entre 29ºC y 31.4ºC; la máxima temperatura fue de 34ºC (Cruz, 1968). Las temperaturas más elevadas correspondieron con las áreas de marismas y manglares. TRANSPARENCIA DEL AGUAEn general, las aguas son poco transparentes y contienen abundantes terrígenos finos en suspensión. Según mediciones con el disco de Secchi, se diferenciaron las tres áreas indicadas en la figura 5: a) aguas claras con visibilidad mayor de 1 m; b) aguas de turbidez media con visibilidad entre 0.5 y 1 m; y c) aguas con visibilidad menor de 0.5 m. SEDIMENTOSSe distinguieron cinco grupos de sedimentos en función al diámetro de las partículas y su grado de clasificación según Cruz (1968). Grupo I: arenas finas, entre bien y moderadamente clasificadas, distribuidas en la parte norte de la laguna, en áreas cercanas al sotavento de la barrera arenosa y a las playas. Grupo II: arenas finas, de transición, pobremente clasificadas. Grupo III: limos pobremente clasificados, procedentes de los ríos y esteros del sur, distribuidos en las porciones occidental y sur de la laguna. Grupo IV: arcilla muy pobremente clasificada, que predomina en la parte subcentral de la laguna. Grupo V: arcillas pobremente clasificadas, en la porción central de la cuenca (Fig. 6). MATERIALES Y MÉTODOSSe estudiaron 65 muestras de sedimentos superficiales húmedo, distribuidas en toda el área (Fig. 7). Las dimensiones de las muestras que se utilizaron fueron aproximadamente de 10 cm² de superficie y 1 cm de espesor y corresponden a 10 cm³ de sedimentos. Los métodos de preparación y estudio empleados fueron iguales a los utilizados en la Laguna de Términos (Ayala Castañares, 1963) y la Laguna Madre (Ayala-Castañares y Segura, 1968), comprendieron el análisis cualitativo y cuantitativo de las poblaciones de foraminíferos tanto vivos como poblaciones totales. DISTRIBUCIONES DE LOS FORAMINÍFEROSComo ya se indicó, la fauna de foraminíferos contenida en los sedimentos, fue estudiada tanto cuantitativa como cualitativamente, identificándose en cada muestra, el número y el porcentaje de ejemplares vivos y muertos de cada especie, los resultados se concentraron en las tablas 1 a 8. Del resultado de esa cuantificación, se conocieron las poblaciones totales y vivientes de foraminíferos para cada muestra y se determinaron los porcentajes de todos los foraminíferos en la laguna. Se hicieron numerosos mapas que indican la distribución de los mismos en la laguna y se determinaron las tendencias de distribución. 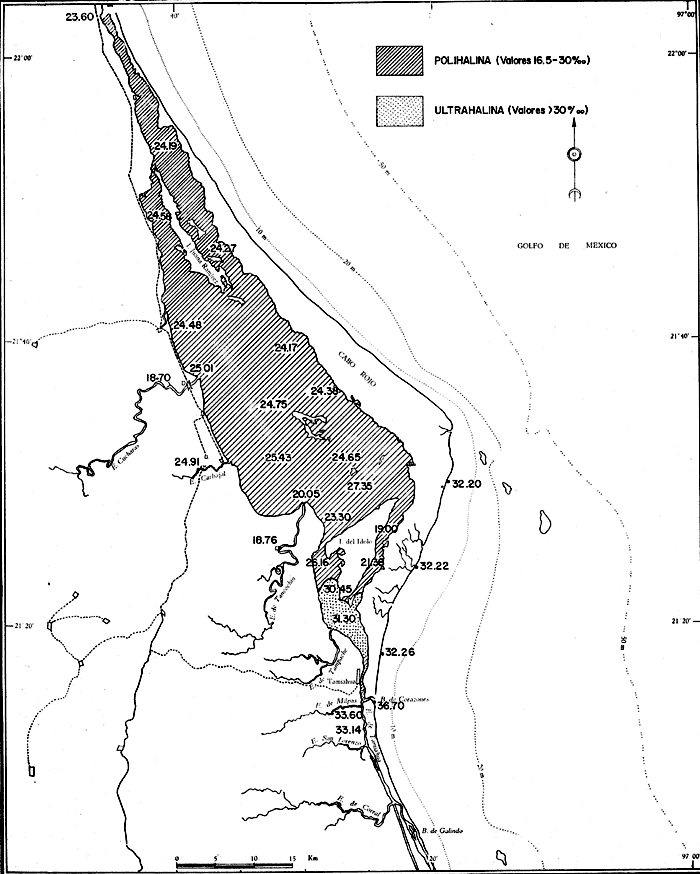 Fig. 4 Distribución de la salinidad del agua del fondo en partes por mil (según Cruz, 1968). 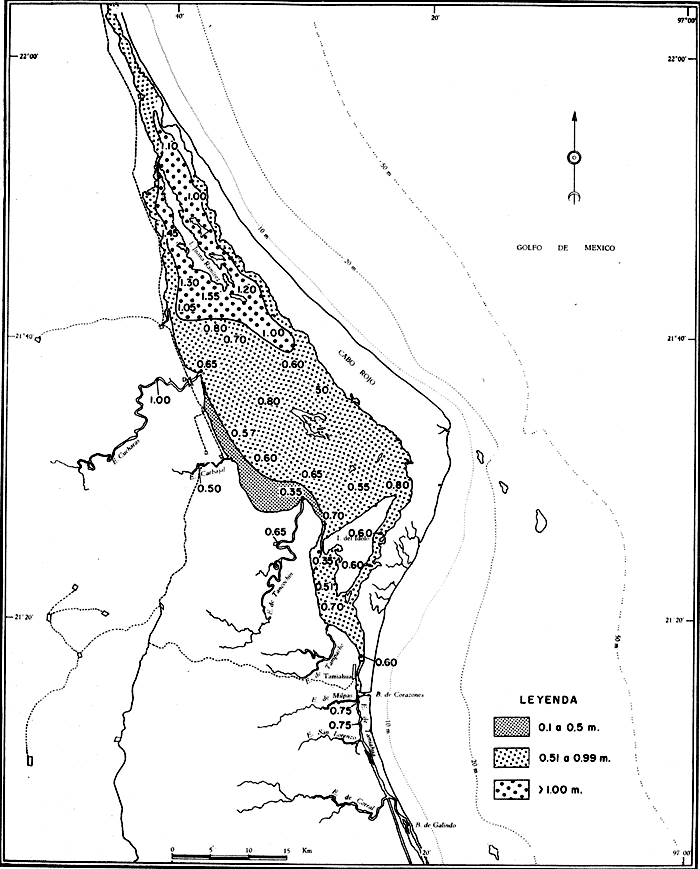 Fig. 5. Transparencia del agua. Los intervalos corresponden a lecturas del disco de Secchi (según Cruz, 1968). 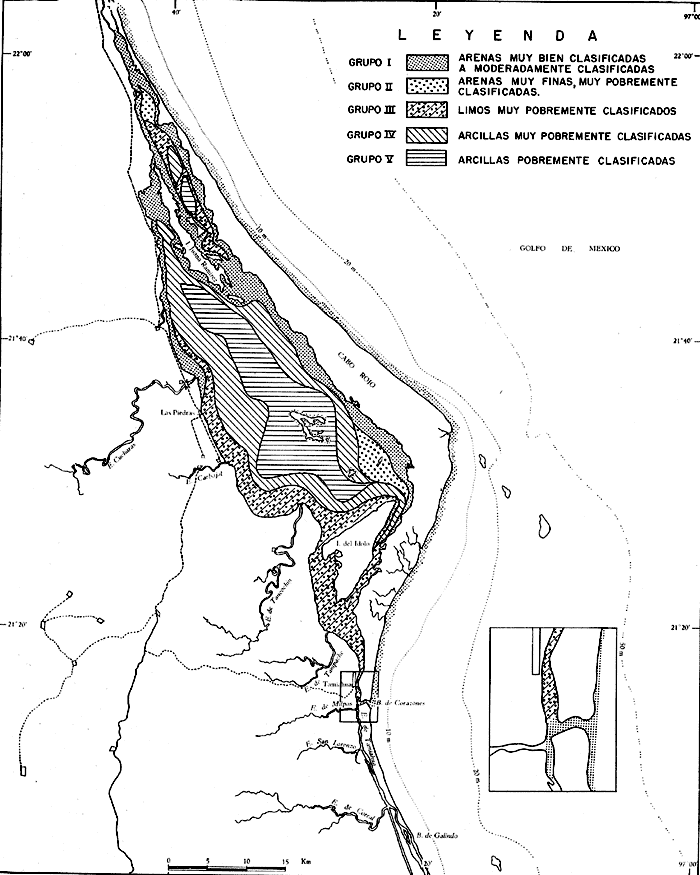 Fig. 6. Distribución de los grupos texturales establecidos (según Cruz, 1968). 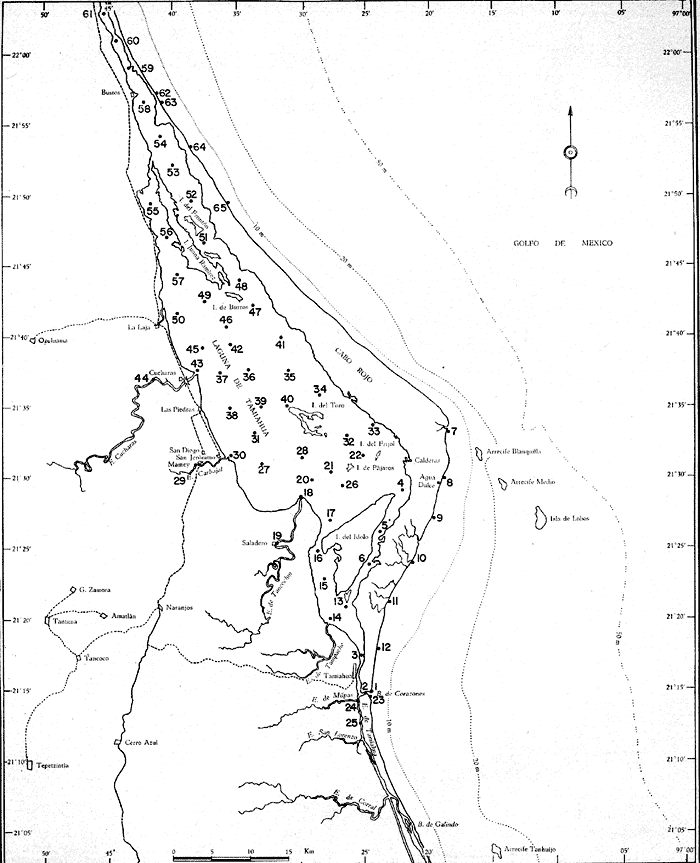 Fig. 7. Localización de las estaciones de observación y muestreo.
Tabla 1 Tabla 2 Tabla 3 Tabla 4 Tabla 5 Tabla 6 Tabla 7 Tabla 8 Tabla 9 Tabla 10 POBLACIONES TOTALES DE FORAMINÍFEROSLas poblaciones totales oscilaron entre 14 y 17,600 ejemplares por muestra (Fig. 8). Hacia la parte norte de la laguna las poblaciones tienden a ser altas, quizás debido a la influencia de las aguas del río Pánuco, que está conectado hacia esta región por un estrecho canal. POBLACIONES MAYORES Y MENORES DE 0.149 mmLos porcentajes obtenidos entre las poblaciones mayores y menores de 0.149 mm son más o menos equivalentes en la mayor parte de las muestras, sin embargo, porcentajes mayores al 90% se encuentran en las estaciones ubicadas a lo largo de la barrera de Cabo Rojo, frente al litoral (Fig. 9). NÚMERO DE GÉNEROS DE FORAMINÍFEROSLa distribución de los géneros de foraminíferos por muestra en la Laguna de Tamiahua osciló entre 2 y 31 géneros (Fig. 10), encontrándose un promedio general de 4 géneros. En áreas próximas a la desembocadura de los esteros se encontró un hecho muy significativo que tiene estrecha relación con ella y aquellas bajo la influencia del agua marina que penetra por la Boca de Corazones, en ellas el número de géneros es mayor, variando entre 11 y 31, asimismo a lo largo del litoral de la barrera arenosa de Cabo Rojo, se encuentra el mayor número de géneros por muestra, entre 3 y 22. NÚMERO DE ESPECIES DE FORAMINÍFEROSFueron identificadas 70 especies de foraminíferos. Sin embargo, se encontró un amplio rango de variación de especies por muestra, entre 2 y 54 (Fig. 11). En los esteros se encontraron entre 2 y 3 especies, en tanto que la laguna propiamente dicha, el número varía entre 5 y 8 especies; esa cifra se elevó hacia el S y en las cercanías de la Boca de Corazones se encuentra el número de especies más alto, entre 23 y 54. Las estaciones de las playas presentan un número de especies que fluctúa entre 5 y 37. Lo anterior pone en evidencia que en la Laguna de Tamiahua, como en otras muchas lagunas, el número de especies aumenta desde las áreas menos marinas, bajo el efecto de los ríos, hasta las más marinas, provenientes del Golfo de México. PORCENTAJES TOTALES DE FORAMINÍFEROS AGLUTINADOSSu distribución en toda la laguna es amplia, pero existe una tendencia de mayores porcentajes desde la porción central, hacia el norte. En general, los porcentajes oscilan entre 0.1 y 1.68% como máximo, siendo el promedio de 2.0%, (Fig. 12). Las condiciones ecológicas que predominan en la desembocadura de esteros y que posiblemente influyen, son principalmente la poca profundidad, la baja salinidad, la presencia de arcillas en los sedimentos, así como la existencia, cerca de estas áreas, de grandes bancos de ostiones y también la turbidez que va de 0.51 a 0.99 m, haciendo de esta área una porción típicamente lagunar. La influencia de los esteros se deja sentir, con una tendencia a mayores porcentajes de formas aglutinadas. PORCENTAJES TOTALES DE MILIÓLIDOS EN LAS POBLACIONESLos miliólidos en la Laguna de Tamiahua, se distribuyen principalmente a lo largo de la barrera arenosa frente al litoral, con porcentajes que van de 1.2 a 43% (Fig. 13), y también en la Boca de Corazones donde la influencia marina es mayor, observándose en dichas áreas, así como en las porciones cercanas a la misma, un aumento en el porcentaje que va de 1.7 hasta 33%; tanto en la porción central de la laguna, como muy hacia el N, se advierten porcentajes muy bajos con franca tendencia a desaparecer. 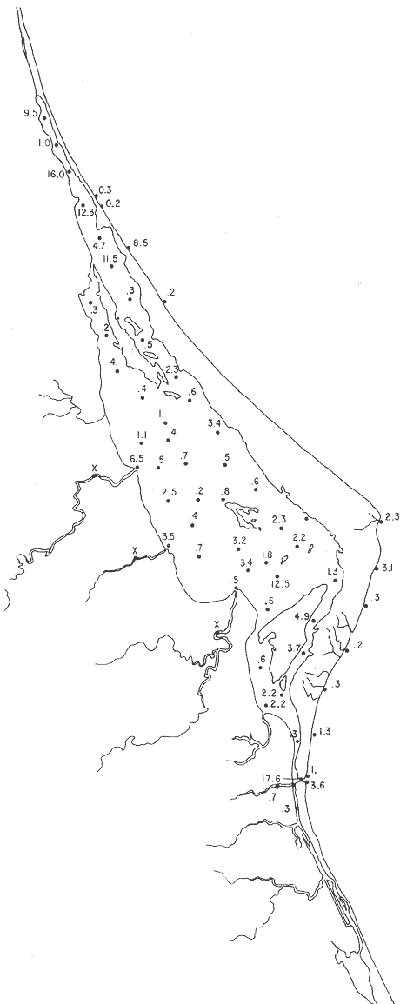 Fig. 8. Poblaciones totales de foraminíferos en miles de ejemplares. 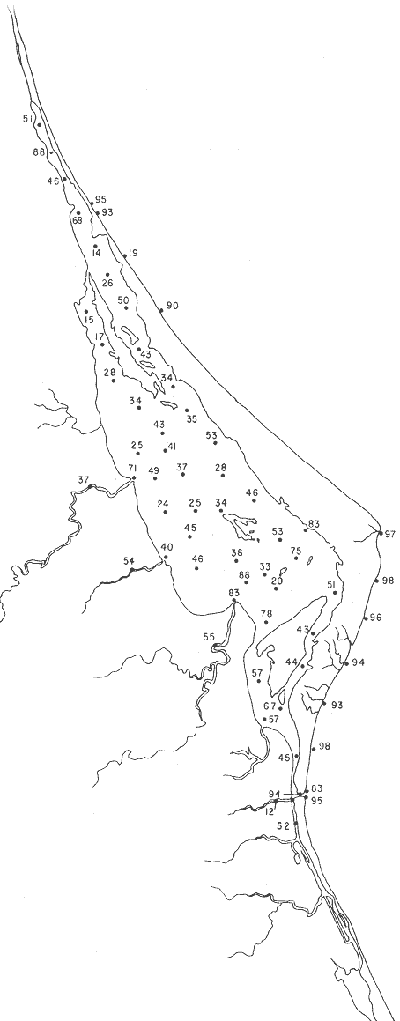 Fig. 9. Población mayor de 0.149 mm. 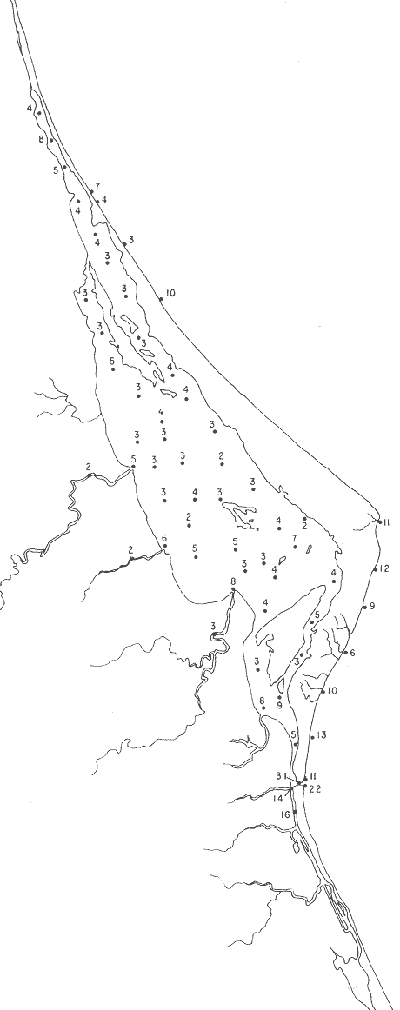 Fig. 10. Número de géneros de foraminíferos por muestra. 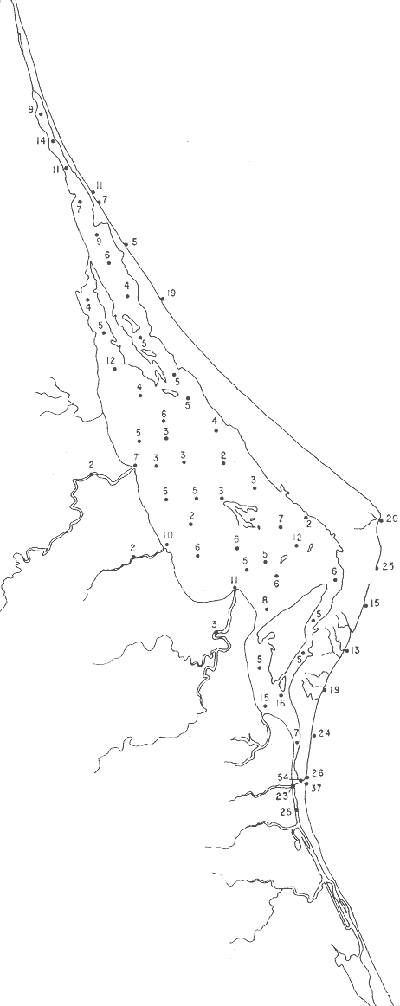 Fig. 11. Número de especies de foraminíferos por muestra. 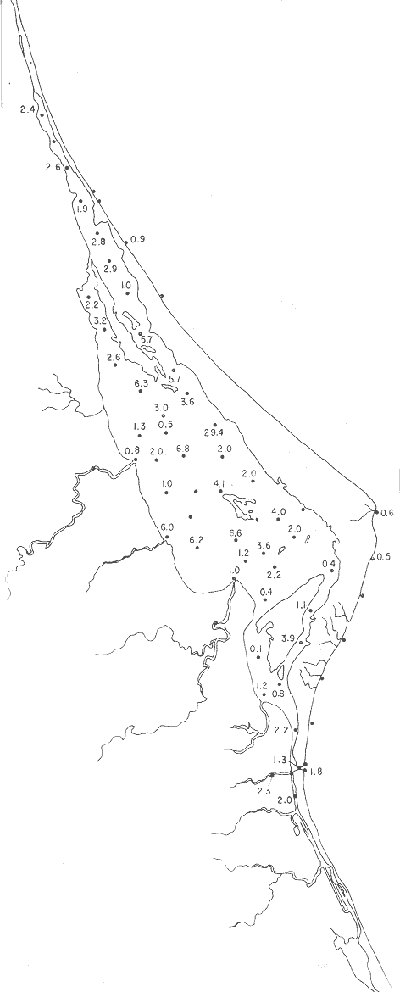 Fig. 12. Porcentajes totales de foraminíferos aglutinados. 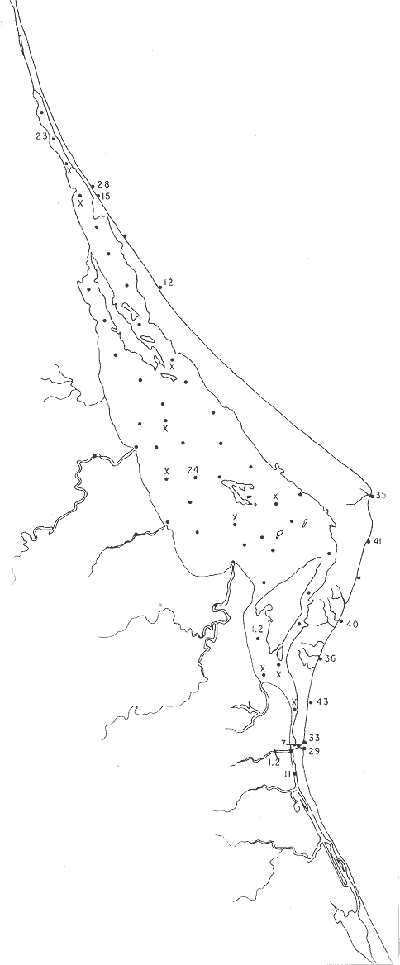 Fig. 13. Porcentajes totales de miliólidos en las poblaciones. Algo similar observó Ayala-Castañares (1963) en la Laguna de Términos, Campeche, en donde los miliólidos existen con mayores porcentajes en las porciones influenciadas por el mar. POBLACIONES VIVIENTES DE FORAMINÍFEROS EN NÚMERO DE EJEMPLARESLas poblaciones vivientes por unidad de muestra se han considerado (de acuerdo con las ideas de F. B Phleger y otros investigadores) como indicadores relativos de la productividad orgánica. Esto ha podido ser comprobado por otros métodos en diversas regiones. En el área de esta investigación las poblaciones vivientes son pequeñas, variando entre 0 y 113 ejemplares por muestra (Fig. 14). Las poblaciones mayores se encuentran en los esteros y áreas afectadas por ellos, indicando la mayor productividad orgánica, posiblemente como un efecto de los nutrientes aportados por esas corrientes. Otra área que presenta poblaciones más altas corresponde a los canales hacia el N de la laguna y se cree que sea debido a la influencia del río Pánuco. Las zonas de fondo arenoso, en particular las playas, área de la boca y sotavento de la barrera arenosa, no presentan ejemplares vivos o éstos son muy escasos, lo cual es evidencia de productividad orgánica baja. Si se comparan las poblaciones vivientes de foraminíferos de la Laguna de Tamiahua, con las de las otras áreas conocidas del Golfo de México, especialmente las lagunas Madre y Términos, así como Bahías de Texas, Sonda del Mississippi; se observa que la productividad orgánica en la Laguna de Tamiahua parece ser más baja que en las otras regiones y ello pudiera ser una evidencia de que, por alguna causa, se hubieran alterado las condiciones ecológicas de la misma, durante la época de la colecta de las muestras. PORCENTAJE DE EJEMPLARES VIVOS DE FORAMINÍFEROS EN LAS POBLACIONES TOTALESEsos porcentajes de foraminíferos vivos han sido considerados como un indicador relativo de la velocidad de depósito, de modo que a mayor porcentaje, la velocidad de sedimentación sería más alta. Dichos porcentajes fueron en general bajos, variando desde 1 a 22%, aunque se observó que en la parte media de la laguna y en las desembocaduras de los esteros tienden a ser mayores (Fig. 15). Las mayores abundancias relativas se encuentran en las áreas situadas en la porción central y hacia el N. ESPECIES DE FORAMINIFEROSLas estaciones subrayadas en los mapas indican poblaciones vivientes.
Género Ammobaculites Cushman, 1910Escaso, en la parte sur de la laguna, en canales y áreas cercanas a los esteros y sin ejemplares vivos; en límos pobremente clasificados, a profundidades de 1 a 2 m, con salinidad entre 20 y 30% Ammobaculites dilatatus Cushman y Brönnimann(Lám. 1, figs. la-c) Ammobaculites dilatatus Cushman y Brönnimann, 1948b. Contr. Cushman. Lab. foramin. Res.,24(2): 39, lám. 7, figs. 10, 11. Ammobaculites dilatatus Cushman y Brönnimann. Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol. Bol., 67 (3): 49, lám. 1, fig. 2. Escasa, sin ejemplares vivos, confinada principalmente a las desembocaduras de los esteros, en limos pobremente clasificados y a profundidades no mayores de 1m; salinidad entre 21 y 36%, (Fig. 16). Hipotipos: CCML-7-Mi, CCML-8-Mi. Ammobaculites sppSe encontraron ejemplares escasos muertos, no identificados específicamente, en la desembocadura de los esteros y canales de la parte meridional de la laguna. 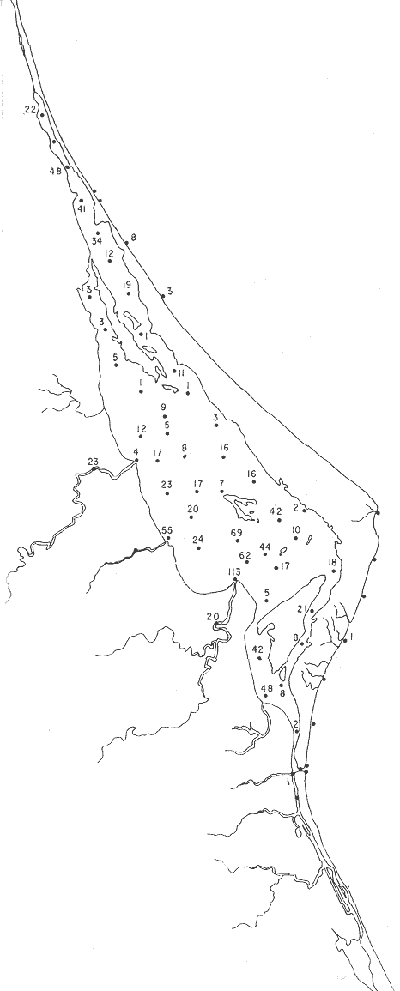 Fig. 14. Poblaciones vivientes de foraminíferos en número de ejemplares. 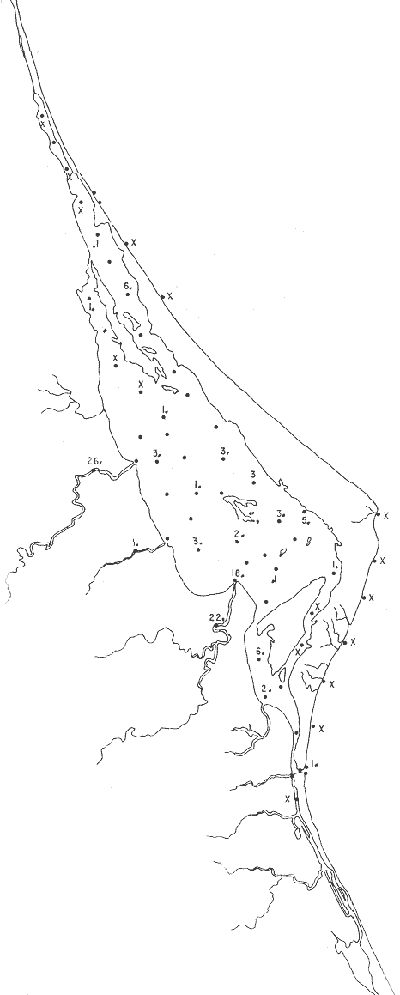 Fig. 15. Porcentajes de ejemplares vivos de foraminíferos en las poblaciones totales. Género Ammonia Brünnich, 1772Está distribuido ampliamente; es común en casi todas las estaciones, con ejemplares vivos en la mayoría de ellas, representando los porcentajes más altos dentro de las poblaciones totales. Ammonia beccarii (Linnaeus) variantes A y B(Lám. 1, figs. 2a-c, 3a-c). Nautilus becarii Linnaeus, 1758, Syst. Nat., 10 Ed. Stockholm: 710. Ammonia beccarii(Linnaeus). Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol. Bol., 67(3): 87, lám. 7, figs. 4a-c, 5a-c. Es la especie más abundante en la laguna, con porcentajes que van de 15.3 a 97.3%, en todos los grupos texturales y temperaturas. La mayor parte de las veces, se hallaron ejemplares vivos (Fig. 17). Hipotipos: CCML-25-Mi, CCML-26-Mi, CCML-27Mi, CCML-28-Mi, CCML-29-Mi, CC ML-30-Mi, CCML-32-Mi, CCM1-33-Mi, CC ML-34-Mi. Género Ammotium Loeblich y Tappan, 1953Individuos muertos de este género, en porcentajes bajos, se encontraron en casi todas las estaciones de la parte central y norte de la laguna, a profundidades entre 1 y 3 m, en todos los tipos de fondo. Ammotium salsum (Cushman y Brönnimann)(Lám. 1, figs. 4a-c) Ammobaculites salsum Cushman y Brönnimann, 1948a. Contr. Cushman Lab. foramin. Res., 24 (1):16, lám. 3, figs- 7-9. Ammotium salsum(Cushman y Brönnimann). Parker y Athearn, 1959, J. Paleont.,33(2):340, lám. 50, figs. 6, 13. Especie representada en muchas estaciones, muerta, con porcentajes en general bajos dentro de la población total, del orden de 0.3 a 29.4% como máximo (Fig. 18). Hipotipos: CCML-55-Mi, CCML-56-Mi. Ammotium sppSe encontraron ejemplares muertos no identificados específicamente, con porcentajes bajos dentro de la población total, entre 0.7 y 2.9%, en la parte norte de la laguna. Género Archaias De Montfort, 1804Se encontró solamente en unas cuantas estaciones de las playas de la barrera arenosa, con porcentajes muy bajos. Archaias angulatus (Fitchel y Moll)Nautilus angulatus Fichtel y Moll., 1978, Test. Micr.: 113, lám. 22, figs. a-e. Archaias angulatus (Fichtel y Moll). Cushman, 1930, Bull. U.S. natn. Mus.,104 (7): 46, lám. 16, figs. 1-3, lám. 17, figs. 3-5. Esta especie se encontró solamente muerta en una estación, en la parte meridional de la laguna; su porcentaje no es significativo dentro de la población total. Hipotipo: CCML-77-Mi. Archaias sppSólo se hallaron ejemplares muertos, no identificados específicamente, en las estaciones de la parte externa de la barrera arenosa, con porcentajes bajos dentro de las poblaciones totales. Hipotipos: CCML-78- Mi, CCML-79-Mi. Género Articulina d'Orbigny, 1826
Articulina sppSe encuentran escasos ejemplares muertos, no identificados específicamente, en dos estaciones de la parte meridional de la laguna, cercanas a la Boca de Corazones. Hipotipos: CCML-70-Mi, CCML-71-Mi, CC ML-72Mi, CCML-73-Mi, CCML-74-Mi. 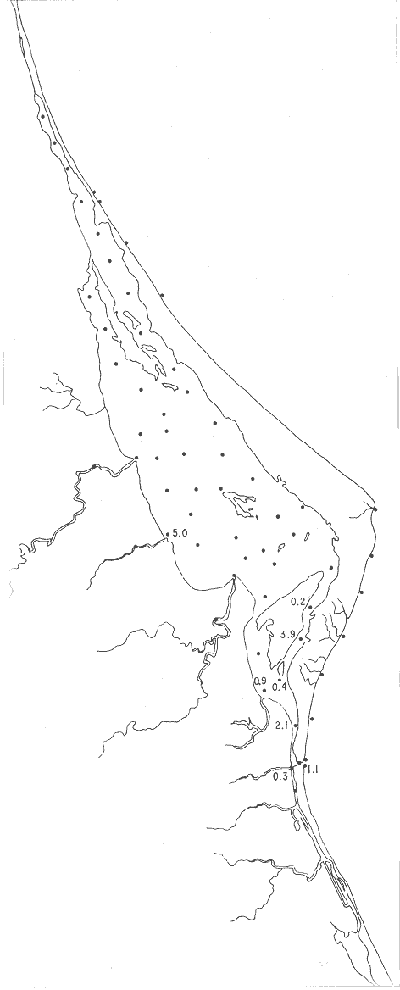 Fig. 16. Porcentajes totales de Ammobaculites dilatatus. 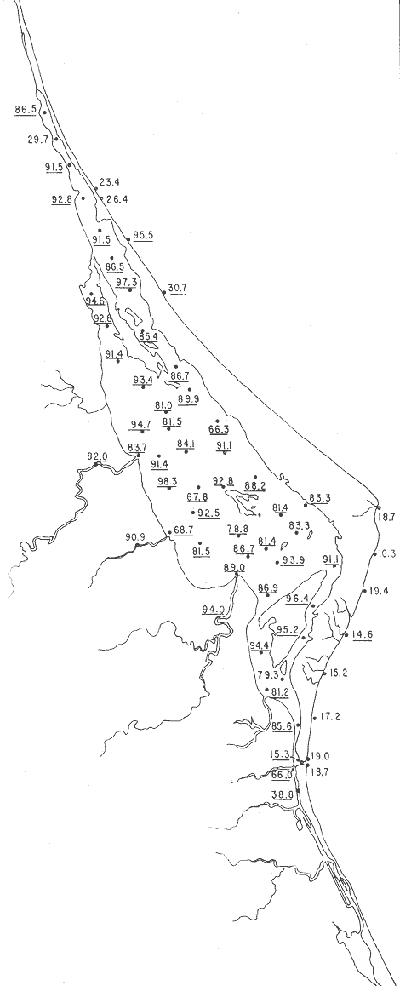 Fig. 17. Porcentajes totales de Ammonia beccarii y vars. Género Bigenerina d'Orbigny, 1826
Bigenerina sp(Lám. 1, figs. 5a-c) Es muy escasa, cerca de la boca, sin ejemplares vivos. Hipotipo: CCML-924-Mi. Género Bolivina d'Orbigny, 1839
Bolivina sppFue encontrado principalmente en las desembocaduras de los esteros y canales de las estaciones de la parte sur de la laguna, con porcentajes muy bajos dentro de la población total. Se observaron ejemplares vivientes en las estaciones YZ-14-64 e W-18-64. Hipotipos: CCML-120-Mi, CCML-121-Mi. Género Brizalina Costa, 1856
Brizalina striatula (Cushman)(Lám. 1, figs. 6a-d) Bolivina striatula Cushman, 1922, Carnegie Inst. Wash. Publ., 17 (311):27 y 28, lám. 3, fig. 10. Brizalina siriatula (Cushman). Wantland, 1967. Ph. D. Dissert. Rice Univ.: 196, lám. 11, figs. 6-7. Se encontró escasa en 4 estaciones de la parte meridional de la laguna, en limos pobremente clasificados, salinidades de 26 a 36% y profundidades de 1 m. Ejemplares vivos se encontraron en - la estación YZ-13-64. Hipotipo: CCML-928-Mi. Género Buccella Andersen, 1952
Buccella hannai (Phleger y Parker)(Lám. 1, figs. 7a-b) Eponides hannaiPhleger y Parker, 1951, Mem. geol. Soc. Am.,46(2): 21, lám. lo, figs. lla,b, l2a,b, 13a,b, 14a,b. Buccella hannai (Phleger y Parker). Andersen, 1961, Bull. geol. Surv. La., 35 (2):106, lám. 22, figs. 4a-c. En la estación YZ-2-64, cercana a la Boca de Corazones, se encontraron dos ejemplares muertos de la especie. Hipotipo: CCML-927-Mi.
Género Buliminella Cushman, 1927
Buliminella elegantissima (d)Orbigny)(Lám. 1, fig. 8a) Bulimina elegantíssima d'Orbigny, 1839a, Voy.. Ani. Mérid. Paris Strasb., 5 (5):51 y 52, lám. 7, figs. 13, 14. Buliminella elegantissima(d'Orbigny). Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol. Bol,. 67 (3):71, lám. 5, figs. 2a,b, 3a,b. Ejemplares muertos de esta especie, fueron encontrados en dos estaciones de la parte meridional de la laguna. Hipotipo: CCML-926-Mi. Género Discorbis Lamarck, 1804
Discorbis bulbosa Parker(Lám. 1, figs. 9a-b) Discorbis bulbosa Parker, 1954b, Bull. Mus. comp. Zool. Harvard, 111 (10):523, lám. 8, figs. 10-12. Discorbis bulbosa Parker. Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol. Bol., 67 (3):78, lám. 5, figs. l3a-c. Se encontró un solo ejemplar muerto, en la estación YZ-264, cercana a la Boca de Corazones. Hipotipo: CCML-930-Mi. Discorbis sppEjemplares muertos del género, no identificados específicamente, se encontraron en la estación YZ-7-64, de la barrera arenosa. Hipotipo: CCML-218-Mi. Género Elphidium Montfort, 1808Junto con el Ammonia, constituye uno de los géneros dominantes de la laguna, común en casi todas las estaciones y en todos los tipos de fondo, en profundidades de 1 a 3 m y salinidades de 20 a 36%, sus porcentajes van de 0.8 a 58.9%, dentro de la población total, se encontró viviente en dos estaciones (Fig. 19). 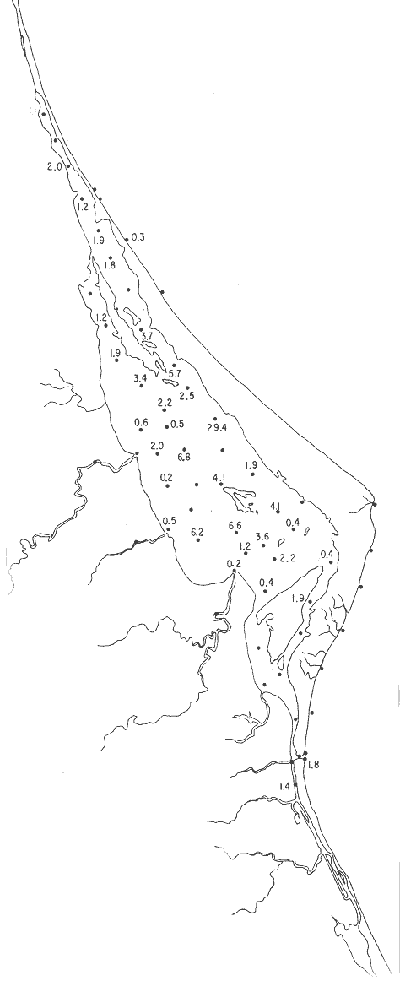 Fig. 18. Porcentajes totales de Ammotium salsum. 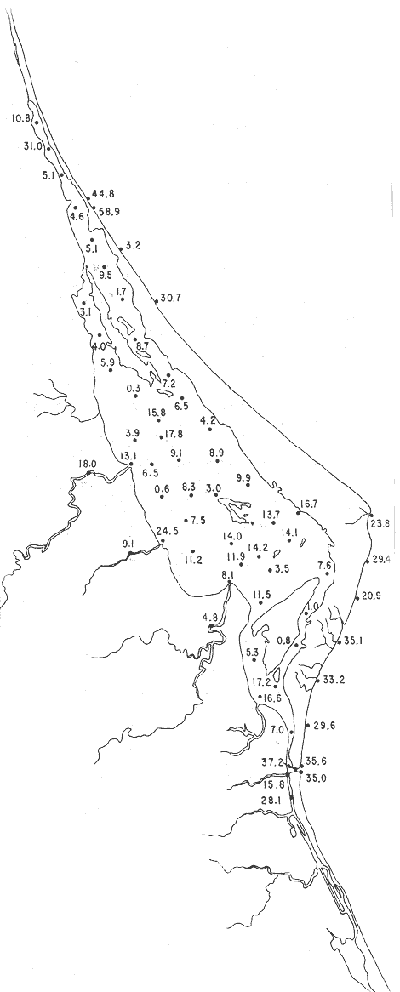 Fig. 19. Porcentajes totales del género Elphidium. Elphidium advenum (Cushman)(Lám. 2, figs. 1 a-e) Polystomella advena Cushman, 1922, Carnegie Inst. Wash. Publ., 17 (311):56 y 57, lám. 9, figs. 11, 12. Elphidium advenum (Cushman). Cushman, 1939, Prof. Pap. U.S. geol. Surv., (191):60, lám. 16, figs. 31-35. Está esporádicamente distribuida en el interior de la laguna, principalmente en los canales de la región septentrional, así como en algunas estaciones cercanas a la Boca de Corazones. Es frecuente en las estaciones del litoral, junto a la barrera a renosa (Fig. 20). Hipotipo: CCML-931-Mi. Elphidium delicatulum Bermúdez(Lám. 2, figs. 2a-e) Elphidium delicatulumBermúdez, 1949, Spec. Publs Cushman Lab. foramin. Res., (25):168, lám. 11, figs. 22-23. Elphidium delicatulum Bermúdez. Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol., Bol.67(3):92, lám. 8, figs. la-b. Escasos ejemplares muertos, fueron encontrados en tres estaciones de la parte meridional de la laguna, en un canal cercano a la Boca de Corazones. Hipotipo: CCML-258-Mi. Elphidium discoidale (d'Orbigny)(Lám. 2, figs. 3a-d) Polystomella discoidalisd'Orbigny, 1839b, In: De la Sagra Hist. Phys. Pol. Île de Cuba "Foraminifères", 8: 56 y 57, lám. 6, figs. 23, 24. Elphidium discoidale (d'Orbigny). Cushman, 1930, Bull. U.S. natn. Mus.,104 (7): 22, lám. 8, figs. 8-9. Esta especie se encontró en las muestras del litoral frente a la barrera arenosa, así como canales de la parte meridional de la misma. Sin ejemplares vivos (Fig. 21). Hipotipo: CCML-259-Mi. Elphidium galvestonense Kornfeld(Lám. 2, figs. 4a-d) Elphidium gunteri Cole var.galvestonensis Kornfeld, 1931, Contr. Dep. Geol. Stanford Univ., 1 (3): 87-88, lám. 15, figs. 1-3. Elphidium galvestonense Kornfeld. Parker, Phleger y Peirson, 1953. Spec. Publs Cushman Fdn. foramin. Res., (2): 7, lám. 3, figs. 15-16. Común en las estaciones del litoral frente a la barrera arenosa, no se encontraron ejemplares vivientes (Fig. 22). Hipotipo: CCML-279-Mi, CCML-281-Mi, CCML-282-Mi, CCML-283-Mi, CCML-284-Mi. Elphidium gunteri Cole(Lám. 2, figs. 5a-e) Elphidium gunteri Cole, 1931, Bull. geol. Surv. Fla. St., (6): 34, lám. 4, figs. 9-10. Elphidium gunteri Cole. Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol. Bol.,67 (3): 93, lám. 8, figs. 5a-b. Esta especie se encuentra frecuentemente en todas las estaciones de la laguna, en todo tipo de fondos, profundidades y salinidades; se encontraron ejemplares vivos en las estaciones YZ-2-64 e YZ-53-64 (Fig. 23). Hipotipo: CCML-932-Mi. Elphidium incertum mexicanum Kornfeld(Lám. 3, figs. la-e) Elphidium incertum (Williamson) var. mexicanum Kornfeld, 1931, Contr. Dep. Geol. Stanford Univ., 1 (3): 89, lám. 16, figs. 1-2. Elphidium incertum (Williamson) var.mexicanum Kornfeld. Cushman, 1939, Prof. Pap. U.S. geol. Surv., (191):57, lám. 16, figs. 3. Ejemplares muertos de esta especie, se encontraron en la parte externa de la laguna, así como en las desembocaduras de los esteros, en los canales y de manera notoria en las estaciones cercanas a la Boca de Corazones, en profundidades de 1 a 3 m (Fig. 24). Hipotipo: CCML-934-Mi. Elphidium cf. E. koeboeense Le Roy(Lám. 3, figs. 2a-c) Elphidium koeboeense Le Roy, 1939, Natuurk. Tjdschr. Ned. Indie,99: 240, lám. 9, figs. 6, 7. Elphidium cf. E. koeboeense Le Roy. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2):8, lám. 3, figs. 22, 23. 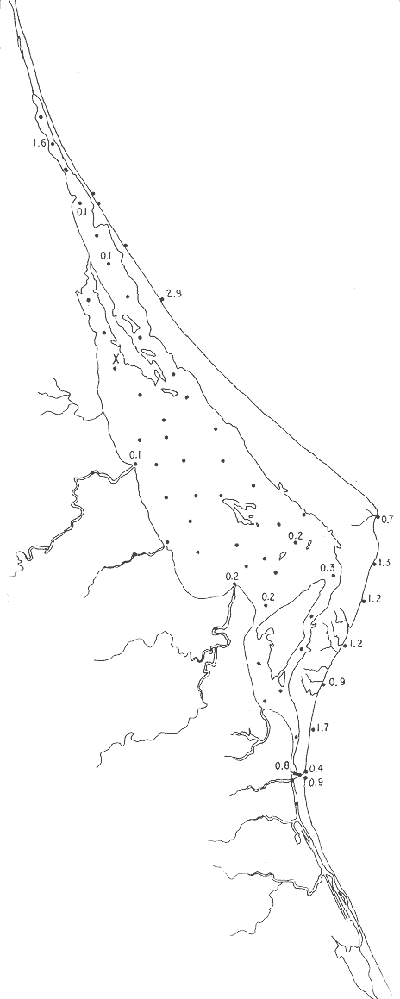 Fig. 20. Porcentajes totales de Elphidium advenum. 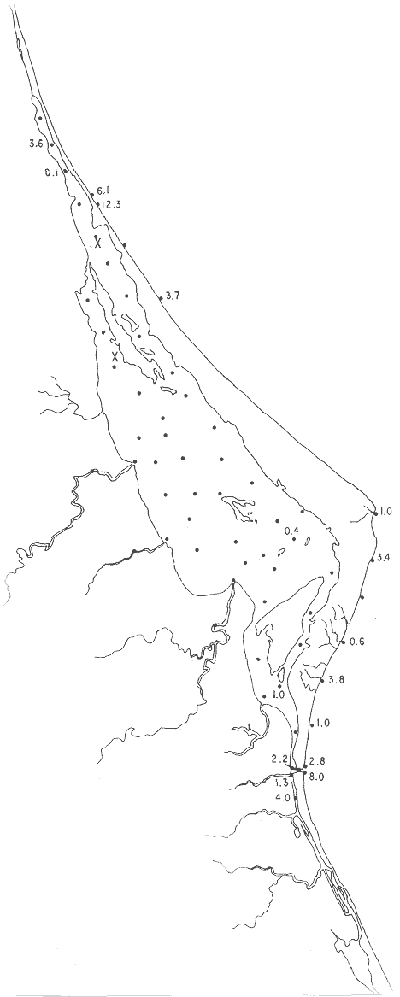 Fig. 21. Porcentajes totales de Elphidium discoidale. 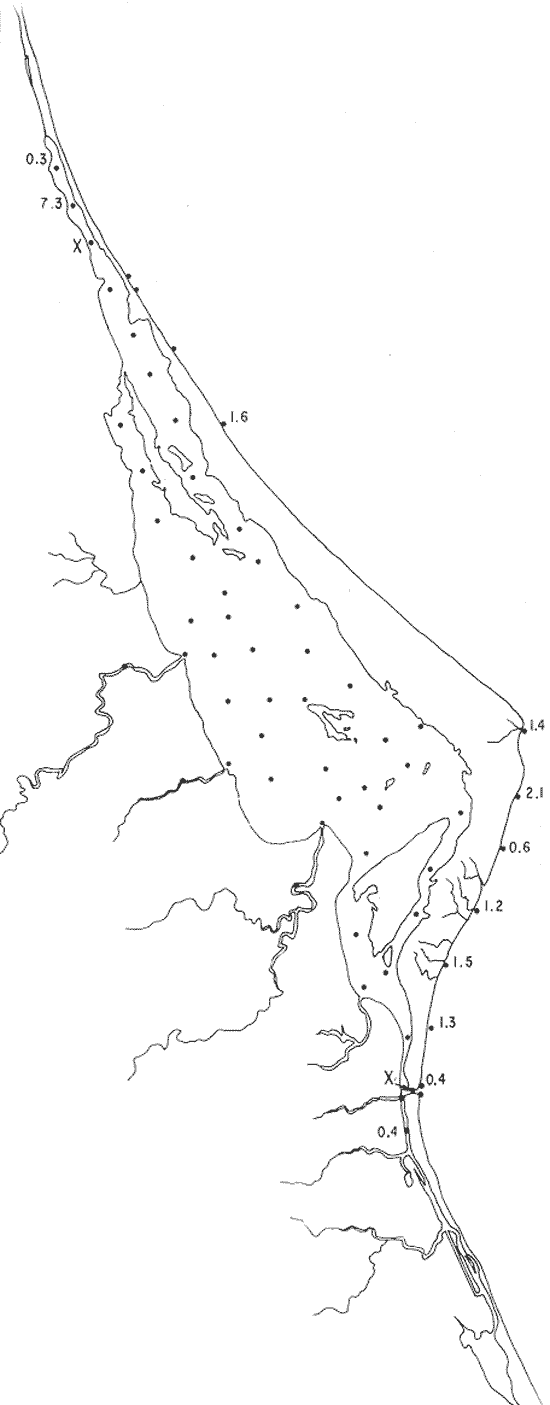 Fig. 22. Porcentajes totales de Elphidium galvestonense. 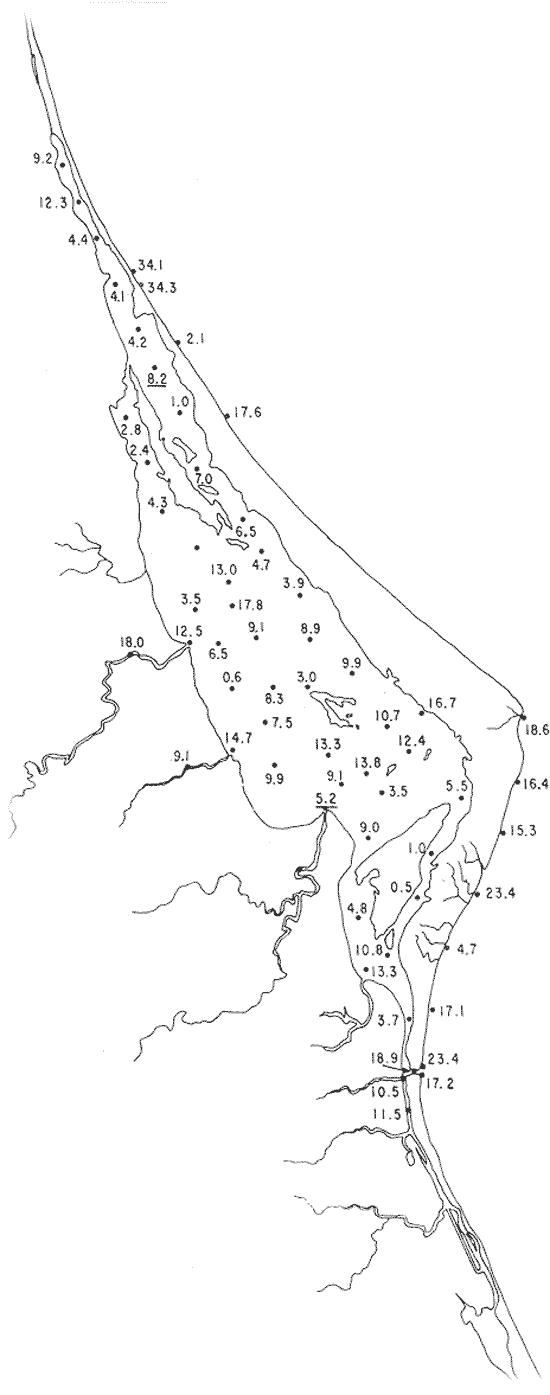 Fig. 23. Porcentajes totales de Elphidium gunteri. En las estaciones en la parte meridional de la laguna y en las cercanias a la Boca de Corazones, se encontraron escasos ejemplares muertos. Hipotipo: CCML-933-Mi. Elphidium matagordanum (Kornfeld)Nonion depressula (Walker y Jacob) var.matagordana Kornfeld, 1931, Contr. Dep. Geol. Stanford Univ.1 (3): 87, lám. 13, figs. 2a,b. Elphidium matagordanum (Kornfeld). Parker, Phleger y Peirson, 1953, Spec. Pbls Cushman Fdn. foramin. Res., (2):8, 9, lám. 3, figs. 24-25. Presente sólo en la estación YZ-20-64, con ejemplares muertos. Hipotipos: CCML-331-Mi, CCML-332-Mi. Elphidium poeyanum (d'Orbigny)(Lám. 3, figs. 3a-d) Polystomella poeyana d'Orbigny, 1839b,In: De la Sagra. Hist. Phys. Pol. Nat. Île de Cuba, "Foraminiféres", 8: 55-56, lám. 6, figs. 25, 26. Elphidium poeyanum (d'Orbigny). Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol., Bol. 67 (3): 65, lám. 9, figs. 1a, b. Especie escasa, muerta, observada en las partes septentrional y meridional de la laguna, tanto en las desembocaduras de los esteros como en los canales y estaciones cercanas a la Boca de Corazones; también se halló en las muestras del litoral junto a la barrera arenosa (Fig. 25). Hipotipos: CCML-340-Mi, CCML-341-Mi, CCML-342-Mi, CCML-343-Mi, CCML-344-Mi, CCML-345-Mi, CCML-346-Mi. Elphidium sppEjemplares muertos no identificados específicamente, fueron encontrados en casi todas las estaciones de la laguna, en general con porcentajes bajos, aunque algunas muestras de la parte meridional, contienen porcentajes más altos (Fig. 26). Hipotipo: CCM-347-Mi. Género Eoguttulina Cushman y Ozawa, 1930
Eoguttulina spEn la estación YZ-25-64, se encontraron ejemplares muertos de este género. Hipotipo: CCML-354-Mi. Género Eponidella Cushman y Hedberg, 1935
Eponidella gardenislandensis Akers(Lám 3, figs. 4a-c) Eponidella gardenislan densis Akers, 1952, J. Paleont., 26 (4): 648, figs. 2a-c. Eponidella gardenislandensisAkers. Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol., Bol. 67 (3):81, lám. 6, figs. 5a-c, 6a-c. Individuos muertos, escasos, se encontraron en dos estaciones cercanas a la Boca de Corazones. Hipotipos: CCML-363-Mi, CCML-364-Mi. Eponidella sppEn la estación YZ-24-64, cercana a la Boca de Corazones, se hallaron pocos ejemplares muertos. Género Eponides De Montfort, 1808
Eponides antillarum (d'Orbigny)(Lám. 3, figs. 5a-d) Rotalina antillarum d'Orbigny, 1839b, In: De la Sagra Hist. Phys. Pol. Nat. Île de Cuba, "Foraminiféres", 8: 75, lám. 5, figs. 4-6. Eponides antillarum(d'Orbigny). Phleger y Parker, 1951, Mem. geol. Soc. Am.,46(2): 20, lám. 10, figs. 9a,b, 10a,b. Un ejemplar muerto se identificó en la estación YZ-2-64 (Fig. 27). Hipotipo: CCML-935-Mi. Eponides repandus (Fichtel y Moll)(Lám. 3, figs. 6a-e) Nautilus repandus Fichtel y Moll, 1798, Test. Micr.: 35, lám. 3, figs. a-d. 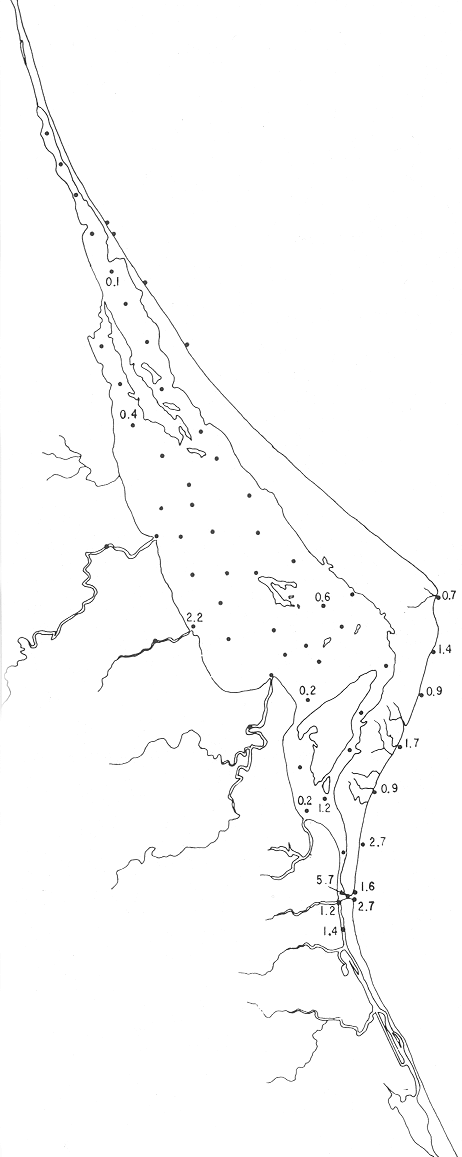 Fig. 24. Porcentajes totales de Elphidium incertum mexicanum. 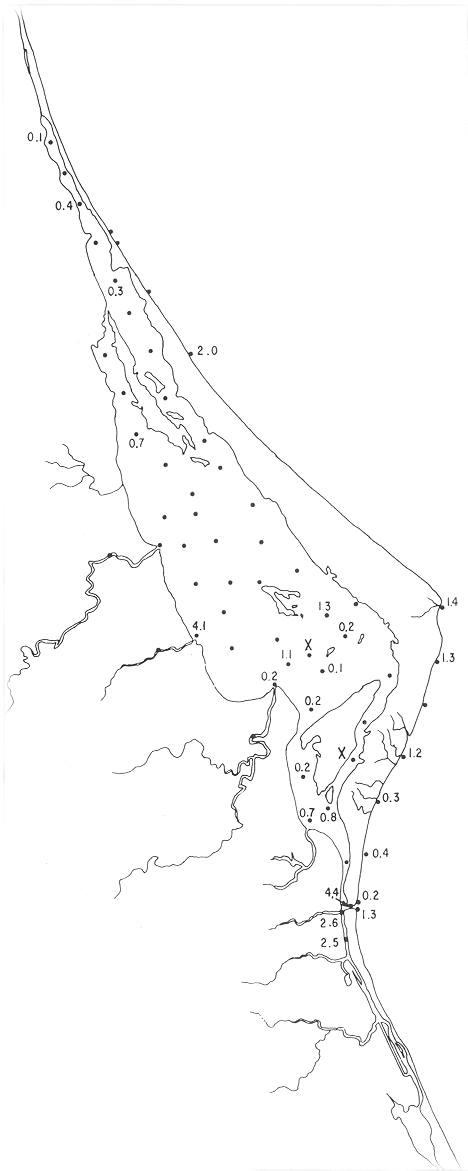 Fig. 25. Porcentajes totales de Elphidium poeyanum. 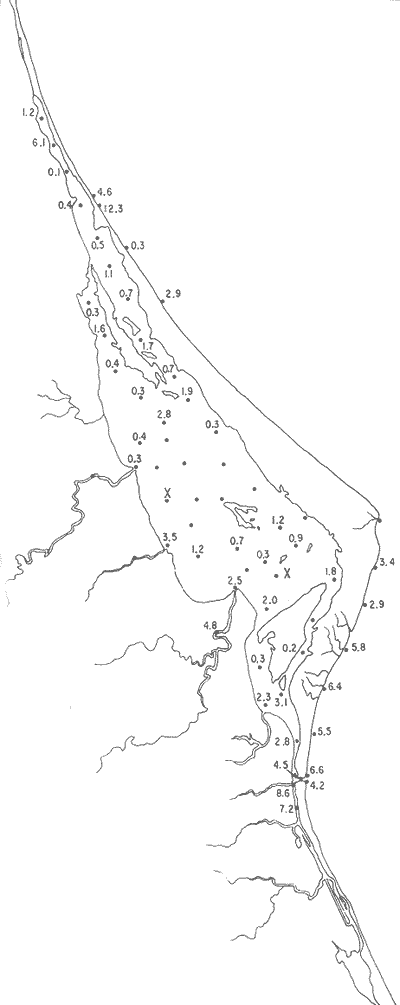 Fig. 26. Porcentajes totales de Elphidium spp. 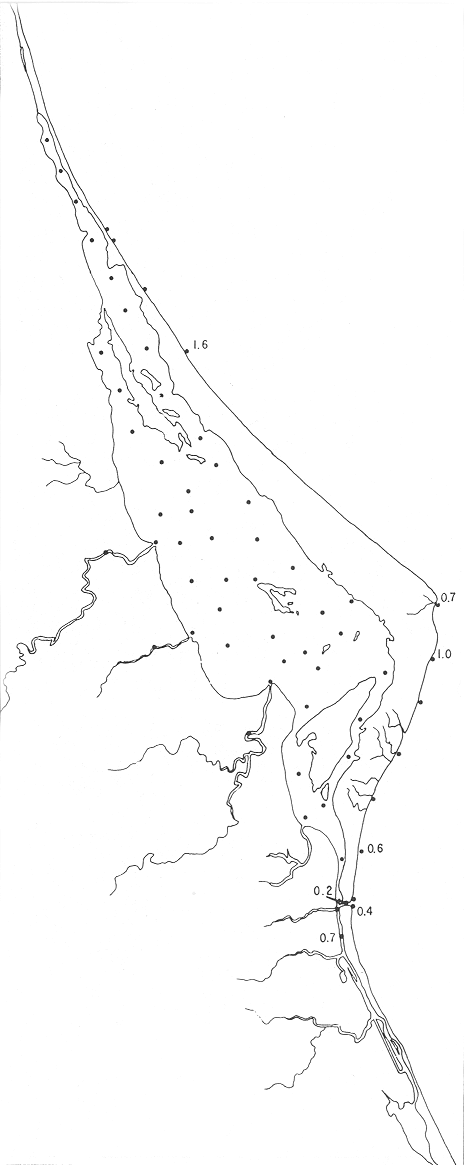 Fig. 27. Porcentajes totales de Eponides antillarum. Eponides repandus (Fichtel y Moll). Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 21, lám. 11, figs. 5a,b, 6a,b. Esta especie se encontró principalmente en las estaciones del litoral frente a la barrera arenosa, con excepción de 3 estaciones situadas cerca de la Boca de Corazones. Hipotipos: CCML-374-Mi, CCML-375-Mi, CCML376-Mi, CCML-377-Mi. Género Fissurina Reuss, 1850
Fissurina sppSe encontraron dos ejemplares muertos en la estación YZ-2-64, en - la parte meridional de la laguna. Hipotipo: CCML-382-Mi. Género Globocassidulina Voloshinova, 1960
Globocassidulina spEjemplares muertos de este género, se localizaron en la estación YZ-24-64, situada en la parte meridional de la laguna, cercana a la Boca de Corazones. Hipotipo: CCML-413-Mi. Género Globulina d'Orbigny, 1839
Globulina spSolamente se encontraron 8 ejemplares muertos del género, en la estación YZ-2-64. Hipotipo: CCML-429-Mi. Género Glomospira Rzehak, 1885
Glomospira sp(Lám. 4, figs. 1a-d) Se encontró en la estación YZ-25-64, entre los esteros de Milpas y San Lorenzo, con ejemplares muertos. Hipotipo: CCML-430-Mi. Género Guttulina d'Orbigny, 1839
Guttulina australis (d'Orbigny)(Lám. 4, figs. 2a-d) Globulina australis d'Orbigny, 1839a, Voy. Am. Mérid. Paris Strasb.,5 (5): 60, lám. 1, figs. 1-4. Guttulina australis (d'Orbigny). Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2):9, lám. 3, figs. 9-10. Especie distribuida principalmente en la porción sur de la laguna y en una estación del litoral frente a la barrera arenosa; sin ejemplares vivos (Fig. 28). Hipotipo: CCML-431-Mi. Guttulina sppEjemplares muertos fueron encontrados en 4 estaciones. Género Hanzawaia Asano, 1944
Hanzawaia concentrica (Cushman)(Lám. 4, figs. 3a-d) Truncatulina concentrica Cushman, 1918b, Bull. U.S. geol. Surv., (676):64, lám. 21, fig. 3. Hanzawaia concentrica(Cushman). Ayala-Castañares, 1963. Univ. Nal. Autón. México. Inst. Geol., Bol. 67 (3):107, lám. 11, figs. 6a-c. Esta especie se encontró principalmente en estaciones del litoral frente a la barrera arenosa (Fig. 29). Hipotipos: CCML-453-Mi, CCM-454-Mi, CCML455-Mi, CCML-456-Mi, CCML-457-Mi. Hanzawaia sppEscasos ejemplares muertos fueron encontrados en 14 estaciones tanto en la entrada de la laguna, como en muestras del litoral frente a la barrera arenosa representando un porcentaje muy bajo dentro de la población total. 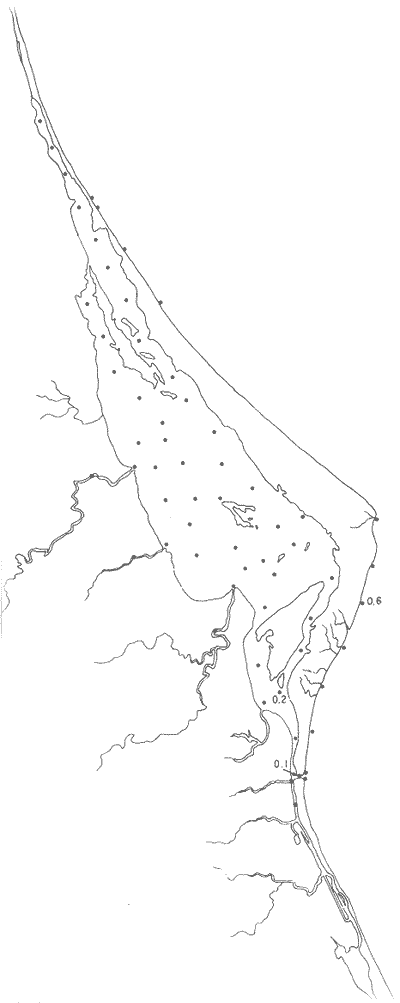 Fig. 28. Porcentajes totales de Guttulina australis. 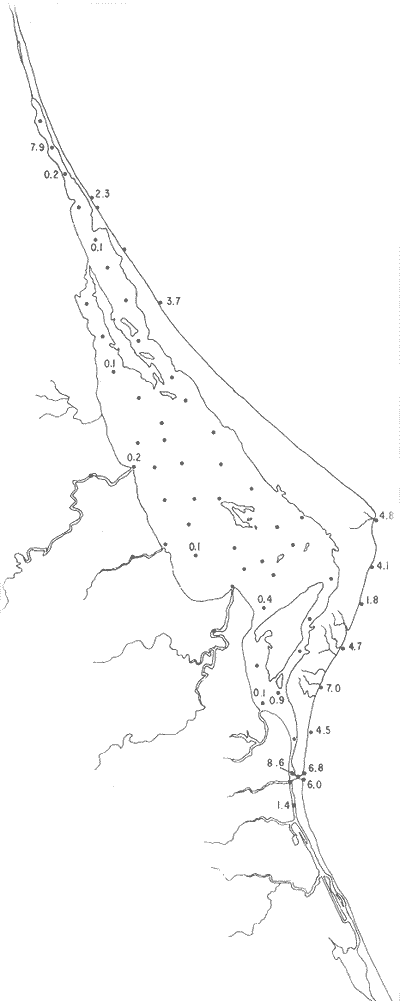 Fig. 29. Porcentajes totales de Hanzawaia concentrica. Género Heterolepa Franzanau, 1884
Heterolepa sppEjemplares no identificados específicamente fueron encontrados muertos en 3 estaciones de la parte sur de la laguna. Género Massilina Schlumberger, 1893
Massilina peruviana (d'Orbigny)(Lám. 4, figs. 4a-c) Quinqueloculina peruviana d'Orbigny, 1839a, Voy. Am. Mérid. Paris, Strasb., 5 (5):73, lám. 4, figs. 1-3. Massilina peruviana(d'Orbigny). Ayala-Castañares y Segura, 1968. Univ. Nal. Autón. México. Inst. Geol. Bol., (87):42-44, lám. 2, figs. 4a-c. Esta especie se encontró muerta en sólo dos estaciones de la porción sur de la laguna, cercanas a la Boca de Corazones. Hipotipo: CCML-942-Mi. Massilina protea Parker(Lám 4, figs. 5a-e) Massilina Protea Parker, 1953. In: Parker, Phleger y Peirson. Spec. Publs Cushman Fdn. foramin. Res., (2):10, lám. 2, figs. 1-4, fig. texto 2. Massilina proteaParker. Wantland, 1967. Ph. D. Dissert. Rice Univ.: 158, lám. 6, figs. 6a, b, c. Especie escasa, con 8 ejemplares muertos en la estación YZ-8-64. Hipotipo: CCML-943-Mi. Massilina sppEjemplares muertos no identificados específicamente, se encontraron en 3 estaciones, una la YZ-60-64 en los canales de la porción norte y 2 estaciones del litoral frente a la barrera arenosa. Los porcentajes son bajos en relación a la población total. Género Miliammina Heron-Allen y Earland, 1930
Miliammina fusca (H. B. Brady)(Lám. 5, figs. la-d) Quinqueloculina fusca H. B. Brady, 1870, Ann. Mag. nat. Hist., 6 (34): 286-288, lám. 11, figs. 2a-c, 3a-b. Mialimmina fusca (H. B. Brady). Parker, 1952, Bull. mus. comp. Zool. Harvard,106 (9): 404-405, lám. 3, figs. 15, 16. Un total de 36 ejemplares muertos, se encontraron en las estaciones del litoral frente a la barrera arenosa y en los canales norte y sur de la laguna, además del área cercana a la Boca de Corazones. Esta especie, coincide en su distribución ecológica con lo citado por Ayala Castañares y Segura (1968). Hipotipos: CCML-518-Mi, CCML-519-Mi, CCML-944-Mi. Género Miliolinella Wiesner, 1931
Miliolinella circularis (Bornemann)(Lám. 5, figs. 2a-d) Triloculina circularis Bornemann, 1855, Z. dt. geol. Ges.,7:349, lám. 19, fig. 4. Miliolinella circularis (Bornemann).In: Parker, 1958, Rep. Swed. deep Sea Exped.,8 (4): 255, lám. 1, figs. 16, 17. Presente en las estaciones del litoral frente a la barrera arenosa, con porcentajes bajos dentro de la población total; no se encontraron ejemplares vivientes (Fig. 30). Hipotipo: CCML-945-Mi. Género Nonionella Cushman, 1926
Nonionella atlantica Cushman(Lám. 5, figs. 3a-f) Ninionella atlantica Cushman, 1947, Contr. Cushman Lab. foramin. Res.,23 (4): 90, 91, lám. 20, figs. 4, 5. Nonionella atlantica Cushman. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2):11, lám. 3, figs. 30. Ejemplares muertos de esta especie se encontraron distribuidos esporádicamente en toda la laguna, principalmente en las estaciones cercanas a la Boca de Corazones (Fig. 31). Hipotipo: CCML-542-Mi, CCML-543-Mi, CCML-544-Mi, CCML-545-Mi. 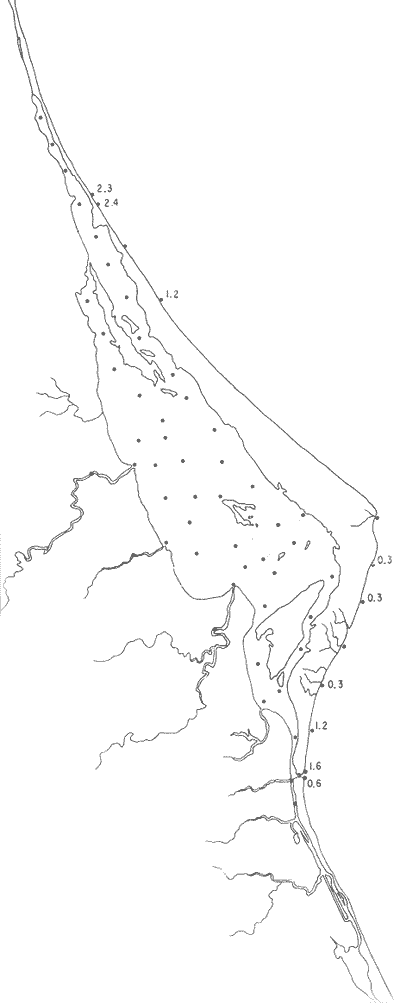 Fig. 30. Porcentajes totales de Miliolinella circularis. 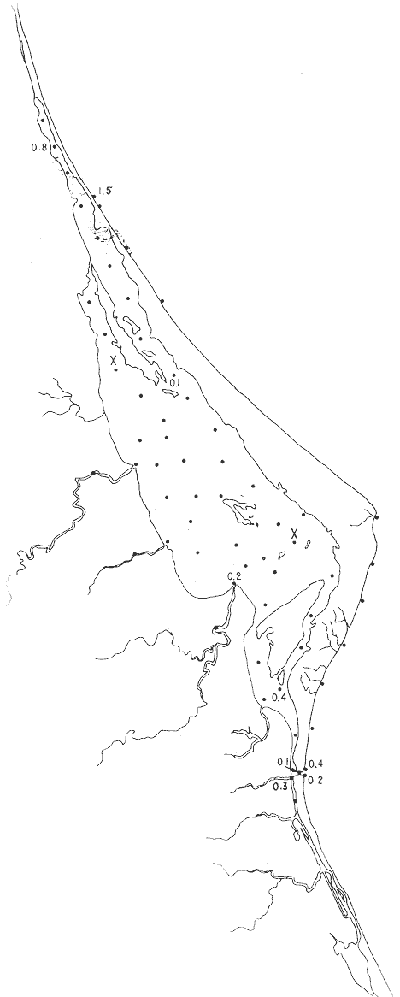 Fig. 31. Porcentajes totales de Nonionella atlantica. Nonionella sppCon ejemplares muertos en 2 estaciones de la porción sur de la laguna. Hipotipo: CCML-550-Mi. Género Palmerinella Bermúdez, 1934
Palmerinella palmerae Bermúdez(Lám. 5, figs. 4a-e) Palmerinella palmerae Bermúdez, 1934, Mems. Soc. cub. Hist. Nat.,8 (2): 84, figs. texto 1-3. Palmerinella palmerae Bermúdez. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn foramin. Res., (2):11, lám. 4, figs. 42, 44. Es una especie escasa en el centro de la laguna, común en los esteros y canales, en particular hacia la porción sur de la misma, en la cual se encuentra en casi todas las estaciones, aunque en general con porcentajes bajos dentro de la población total. Sin ejemplares vivos (Fig. 32). Hipotipos: CCML-622-Mi, CCML-623-Mi, CCML-624-Mi, CCML-625-Mi, CCML-626-Mi, CCML-627-Mi. Palmerinella sppSe encontró con 8 ejemplares muertos en la estación YZ-264. Género Peneroplis De Montfort, 1808
Peneroplis sppIndividuos muertos y con porcentajes muy bajos dentro de la población total, se encontraron en 5 estaciones del litoral frente a la barrera arenosa y 2 cercanas a la Boca de Corazones. Hipotipos: CCML-637-Mi, CCML-638-Mi. Género Planorbulina d'Orbigny, 1826
Planorbulina mediterranensis d'Orbigny(Lám. 5, figs. 5a-c) Planorbulina mediterranensis d'orbigny, 1826, Annls. sci. nat.,7 (2): 280, lám. 14, figs. 4-6. Planorbulina mediterranensis d'Orbigny. Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 33, lám. 19, figs. 5a, b. En la estación YZ-2-64 se identificó un solo ejemplar muerto de esta especie. Hipotipo: CCML-954-Mi. Género Planulina d'Orbigny, 1826
Planulina sppPresente solamente en algunas estaciones de la barrera arenosa externa, en su porción sur y otras Cercanas a la Boca de Corazones; porcentajes bajos y sin ejemplares vivos. Hipotipos: CCML-571-Mi, CCML-572-Mi, CCML-573-Mi, CCML-574-Mi. Género Poroeponides Cushman, 1944
Poroeponides cf. P. lateralis (Terquem)(Lám. 6, figs. la-b) Rosalina lateralis Terquem, 1878, Mem. Soc. geol. Fr., ser. 3,1: 25, lám. 2, figs. lla-c. Poroeponides cf. P. lateralis(Terquem). Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 23, lám. 12, figs. 5a, b. Especie representada por un solo ejemplar muerto, encontrado en la estación YZ-2-64. Género Pseudomassilina Lacroix, 1838
Pseudomassilina sppEjemplares muertos, fueron encontrados en las estaciones del litoral frente a la barrera arenosa (Fig. 33). Hipotipos: CCML-586-Mi, CCML-587-Mi, CCML-558-Mi, CCML-589-Mi, CCML-590-Mi. 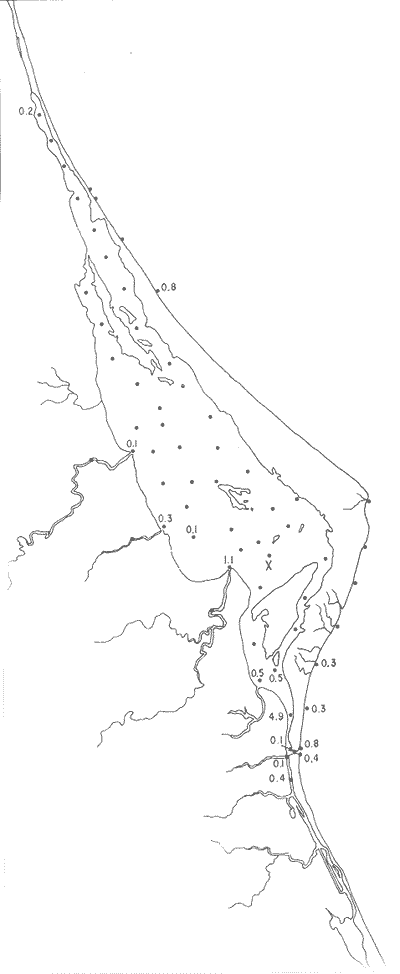 Fig. 32. Porcentajes totales de Palmerinella palmerae. 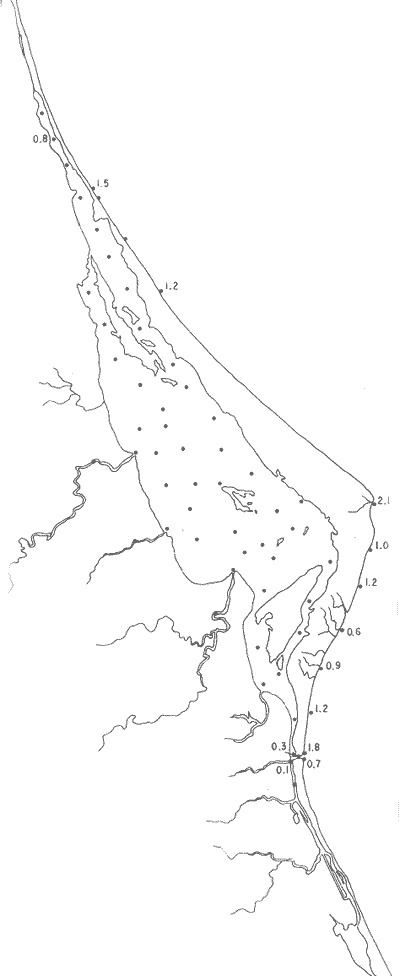 Fig. 33. Porcentajes totales de Pseudomassilina spp. Género Pyrgo Defrance, 1824
Pyrgo cf. P. elongata (d'Orbigny)(Lám. 6, figs. 2a-d) Biloculina elongatad'Orbigny, 1826, Annls. Sci. nat., 7:298, n. 4. Pyrgo elongata (d'Orbigny).Cushman, 1929, Bull. U.S. natn. Mus., 104 (6): 70, lám. 19, figs. 2-3. Escasos individuos muertos de esta especie, se encontraron en una sola estación cercana a la Boca. Hipotipo: CCML-949-Mi. P yrgo cf. P . nasutus(Lám. 6, figs. 3a-e) Pyrgo nasutus Cushman, 1935. Smithson, misc. Collns., 91 (2l): 7, lám. 3, figs. 1-4. Pyrgo nasutus Cushman. Wantland, 1967, Ph. D. Dissert. Rice Univ.: 161, lám. 7, figs- 1a-b. Especie muy escasa, con ejemplares muertos y solamente presentes en la estación YZ-2-64. Hipotipo: CCML-950-Mi. Pyrgo subsphaerica (d'Orbigny)(Lám. 6, figs. 4a-d) Biloculina subsphaerica d'Orbigny, 1839h. In: De la Sagra Hist. Phys. Pol. Nat. Île de Cuba "Foraminiféres",8:162, 163, lám. 8, figs. 25-27. Pyrgo subsphaerica (d'Orbigny). Cushman, 1929, Bull. U.S. natn. Mus., 104 (6): 68, lám. 18, figs. 1-2. Un solo ejemplar muerto de esta especie se halló en la estación YZ-2-64. Hipotipo: CCML-952-Mi. Pyrgo sppSolamente se encontró un ejemplar, no identificado específicamente, en la estación YZ-2-64. Género Quinqueloculina d'Orbigny, 1826Está distribuido principalmente en las estaciones localizadas en la parte externa de la barrera arenosa y algunas de la parte media y sur de la laguna; en general los porcentajes mayores se presentan en las estaciones del litoral frente a la barrera y en aquellas cercanas a la Boca de Corazones. Quinqueloculina compta Cushman(Lám. 6, figs. 5a-d) Quinqueloculina compataCushman, 1947, Contr. Cushman Lab. foramin. Res., 23 (4): 87, 88, lám. 19, fig. 2. Quinqueloculina compta Cushman. Phleger y Parker, 1951, Mem. geol. Soc. Am.,46 (2): 7, lám. 3, figs. 16a, b, 17a, b. Especie representada por escasos ejemplares muertos, en estaciones del litoral frente a la barrera arenosa y algunas cercanas a la boca, con porcentajes bajos dentro de la población total (Fig. 34). Hipotipo: CCML-653-Mi, CCML-654-Mi, CCML-952-Mi. Quinqueloculina lamarckiana d'Orbigny(Lám. 7, figs. 1a-e) Quinqueloculina lamarckiana d'Orbigny, 1839.In: De la Sagra, Hist. Phys. Pol. nat. Île de Cuba "Foraminiféres", 8:189, lám. 11, figs. 14, 15. Quinqueloculina lamarckianad'Orbigny. Plileger y Parker, 1951, Mem. geol. Soc. Am.,46(2): 7, lám. 4, figs. 1,ab. Escasos ejemplares muertos de esta especie, se hallaron en 4 estaciones de la porción sur de la laguna, cerca de la Boca de Corazones. Hipotipo: CCML-959-Mi. Quinquleoculina poeyana d'Orbigny(Lám. 7, figs. 2a-e) Quinqueloculína Poeyana d'Orbigny, 1839b.In: De la Sagra Hist. Phys. Pol. nat. Île de Cuba "Foraminiréres", 8: 191, lám. 11, figs. 25-27. Quinqueloculina poeyana d'Orbigny. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn foramin. Res., (2):12, lám. 2, figs. 13, 14. 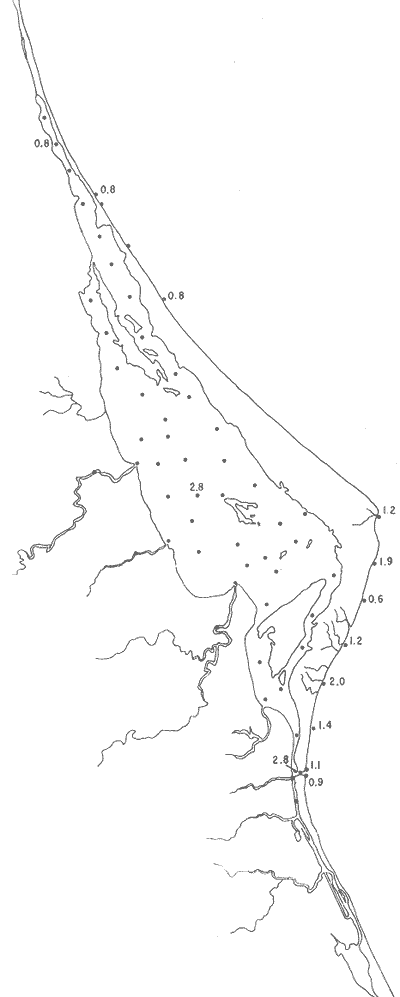 Fig. 34. Porcentajes totales de Quinqueloculina compta. 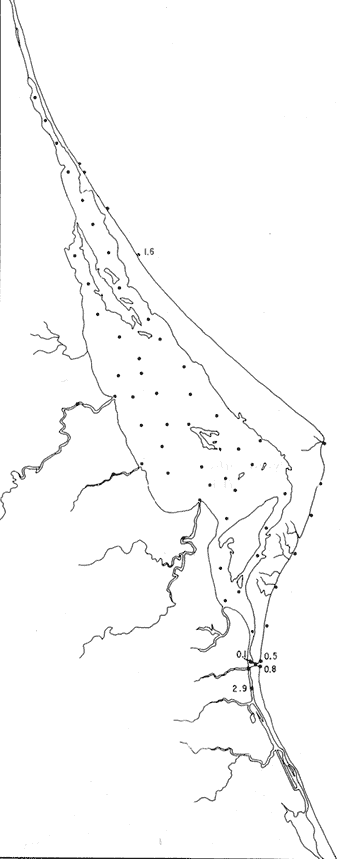 Fig. 35. Porcentajes totales de Quinqueloculina poeyana. Especie restringida casi a las estaciones cercanas a la Boca de Corazones; sin ejemplare vivos (Fig. 35). Hipotipo: CCML-958-Mi. Quinqueloculina seminulum (Linnaeus)(Lám. 7, figs. 3a-c) Serpula seminulum Linnaeus, 1758, Syst. Nat., 10 Ed Stockholm: 786. Quinqueloculina seminulum(Linnaeus). Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. fora min. Res., (2): 12, lám. 2, figs. 18, 19. Abundantes ejemplares muertos de esta especie, se encontraron principalmente en las estaciones del litoral frente a la barrera arenosa y áreas cercanas a la Boca de Corazones (Fig. 36). Hipotipo: CCML-961-Mi. Quinqueloculina sppEscasos ejemplares, todos muertos, con porcentajes bajos dentro. de la población total, fueron encontrados en forma esporádica en el interior de la laguna y algunas estaciones del litoral frente a la barrera arenosa particularmente en el área cercana a la boca. Quinqueloculina tenagos Parker(Lám. 7, figs. 4a-d) Quinqueloculina rhodiensisParker, 1953.In: Parker, Phleger y Peirson, Spec. Publs Cushman Fdn. foramin. Res., (2):12, lám. 2, figs. 15-17. Quinqueloculina tenagos Parker. Parker, 1962, Contr. Cushman Fdn. foramin. Res., 13 (3): 110. Presente en la estación YZ-2-64, con 56 ejemplares todos muertos, no siendo significativo el porcentaje de esta especie dentro de la población total. Hipotipo: CCML-962-Mi. Quinqueloculina vulgaris d'Orbigny(Lám. 7, figs. 5a-d) Quinqueloculina vulgaris d'Orbigny, 1826, Annls. Sci. nat., 7: 302, n. 33. Quinqueloculina vulgaris d'Orbigny. Cushman, 1944, Spec. Publs Cushman Lab. foramin. Res., (12):15, lám. 2, fig. 22. Especie representada en casi todas las estaciones localizadas en el litoral frente a la barrera arenosa, sin ejemplares vivientes (Fig. 37). Hipotipos: CCML-728-Mi, CCML-729-Mi, CCML730-Mi, CCML-731-Mi. Quinqueloculina wiesneri Parr(Lám. 7, figs. 6a-d) Quinquelocuilna anguina (Terquem) var.wiesneri Parr, 1950, B.A.N.Z. Antarctic Res. Exped. 1929-1931, Rep. Ser. B., 5 (6): 290, lám. 6, fig 10. Quinqueloculina wiesneri Parr. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2):13, lám. 2, figs. 20-22. Ejemplares muertos de esta especie fueron ,encontrados particularmente en algunas estaciones del litoral frente a la barrera arenosa y áreas cercanas a la Boca de Corazones (Fig. 38). Hipotipo: CCML-963-Mi. Género Reophax Montfort, 1808
Reophax nana RhumblerReophax nana Rhumbler, 1913, Ergebn. PlanktonExped. Humboldt., 3 (2): 471, lám. 8, figs. 6-12. Reophax nana Rhumbler. Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2): 13, lám. 1, fig. 11. Se encontraron tres ejemplares muertos de esta especie, en la estación YZ-2-64. Hipotipo: CCML-966-Mi. Reussella atlantica Cushman(Lám. 8, figs. la-b) Reussella spinulosa (Reuss) var.atlantica Cushman, 1947, Contr. Cushman Lab. foramin. Res.,23 (4): 91, lám. 20, figs. 6-7. Reussella atlantica Cushman. Ayala-Castañares, 1963. Univ. Nal. Autón. México. Inst. Geol., Bol. 67 (3): 77, lám. 5, figs. 11a, b. 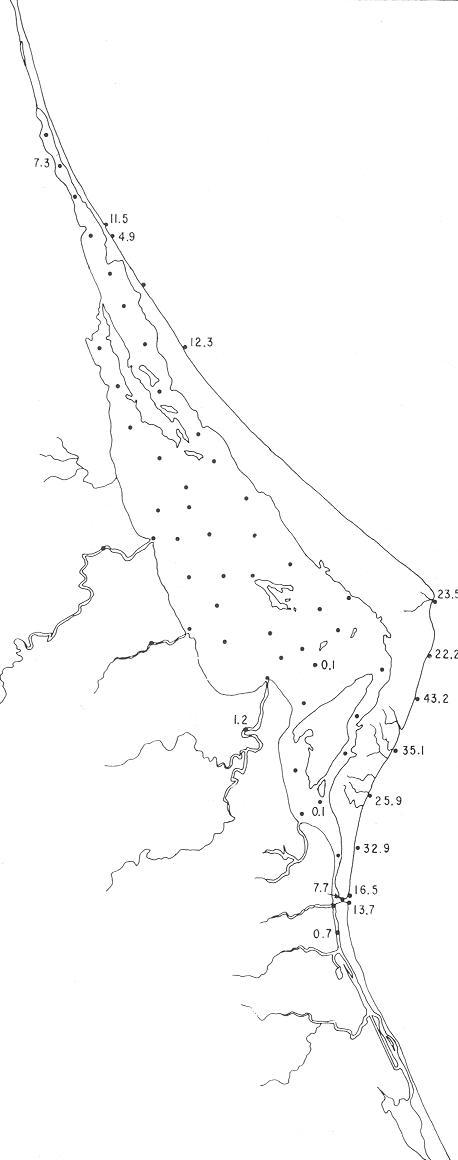 Fig. 36. Porcentajes totales de Quinqueloculina seminulum. 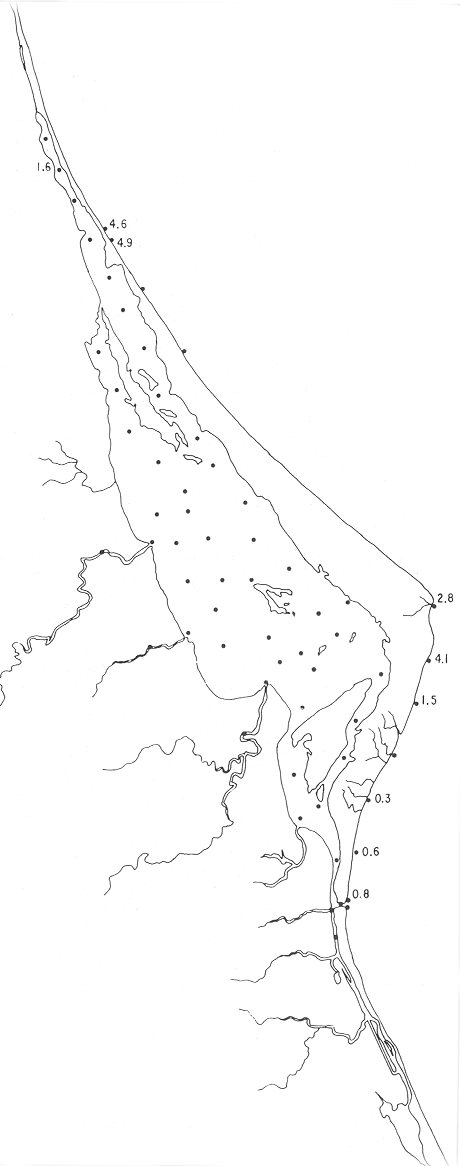 Fig. 37. Porcentajes totales de Quinqueloculina vulgaris. Especie representada por ejemplares muertos, en la estación YZ-2-64. Hipotipo: CCML-967-Mi Género Rosalina d'Orbigny, 1826
Rosalina cf. R. concinna (H. B. Brady)(Lám 8, figs 2a-b) Discorbina concinna H. B. Brady, 1884, Rep. Sci. Results Voy. H. M. S. Challenger Zool., 9:646, lám. 20, figs. 7-8. Rosalina cf. R. concinna (H. B. Brady). Ayala-Castañares, 1963, Univ. Nal. Autón. México. Inst. Geol., Bol. 67 (3): 79, Lám 6, figs. 1a-c. Especie muy escasa, con ejemplares muertos en la estación YZ-23-64, cercana a la Boca de Corazones (Fig. 39). Hipotipo: CCML-965-Mi. Rosalina floridana (Cushman)(Lám. 8, figs. 3a-b) Discorbis floridana Cushman, 1922, Carnegie Inst. Wasli. Publ., 17 (311): 39, lám, 5, figs. 11, 12. Rosalina floridana(Cushman). Ayala-Castañares y Segura, 1968, Univ. Nal. Autón. México. Inst. Geol., Bol. (87): 51, lám. 4, figs. 2a-c. Especie presente en tres estaciones del área cercana a la Boca de Corazones. YZ-2-64, YZ-23-64, YZ-25-64 y con ejemplares muertos. Hipotipo: CCML-964-Mi. Rosalina sppSe encontraron ejemplares muertos, en tres estaciones de la porción sur de la laguna, cercanas a la Boca de Corazones. Hipotipos: CCML-779-Mi, CCML-780-Mi. Género Rotalia Lamarck, 1804
Rotalia translucens Phleger y Parker(Lám 8, figs. 4a-b) Rotalia translucens Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 24, lám. 12, figs. 11a, b, l2a, b. Rotalia translucens Phleger y Parker. Parker, 1954b, Bull. Mus. comp. Zool. Harvard, 111 (10): 531, lám. 10, figs. 3, 7. Un ejemplar muerto de esta especie, se encontró en la estación YZ-2-64. Hipotipo: CCML-968-Mi. Género Siphonina Reuss, 1850
Siphonina pulchra Cushman(Lám. 8, figs. 5a-b) Siphonina pulchra Cushman, 1919, Carnegie Inst. Wash. Publ., (291): 42, lám. 14, figs. 7a-c. Siphonina pulchra Cushman. Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 24, lám. 12, figs. l5a, b. Especie representada en la estación YZ-8-64, todos muertos. Hipotipo: CCML-969-Mi. Siphonina sppEscasos ejemplares muertos, fueron encontrados en la estación YZ-2-64. Género Spiroloculina d'Orbigny, 1826
Spiroloculina sppEjemplares muertos, se encontraron en dos estaciones, YZ-2-64 e YZ-12-64. Hipotipos: CCML-822-Mi, CCML-823-Mi. Género Textularia Defrance, 1824
Textularia mayori Cushman(Lám. 8, figs. 6a-c) Textularia mayori Cushman, 1922, Carnegie Inst. Wash. Publ., 17 (311): 23, lám. 2, fig. 3. Textularia mayori Cushman. Phleger y Parker, 1951, Mem. geol. Soc. Am., 46 (2): 5, lám. 2, figs. 1-5. Esta especie se identificó muerta, escasa en dos estaciones cercanas a la Boca de Corazones (Fig. 40). Hipotipo: CCML-845-Mi. Textularia sppEjemplares muertos del género, fueron encontrados principalmente en tres estaciones del litoral frente a la barrera arenosa y en la estación YZ-2-64, de la Boca de Corazones. Hipotipos: CCML-851-Mi, CCML-852-Mi, CCML853-Mi. 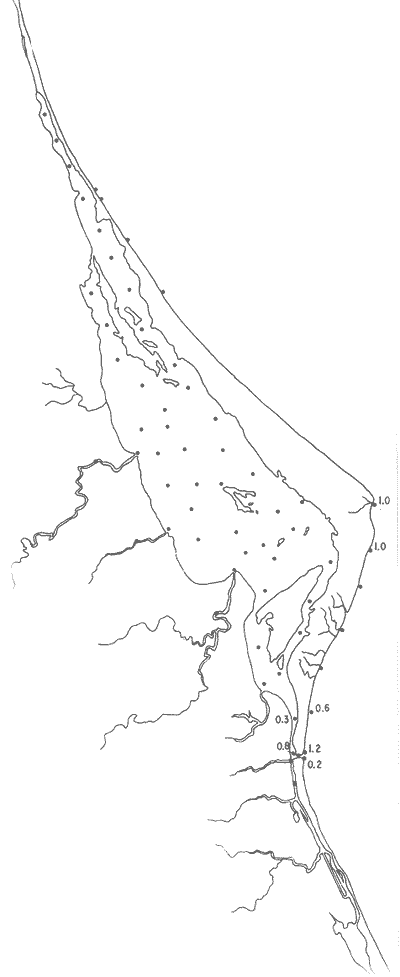 Fig. 38. Porcentajes totales de Quinqueloculina wiesneri. 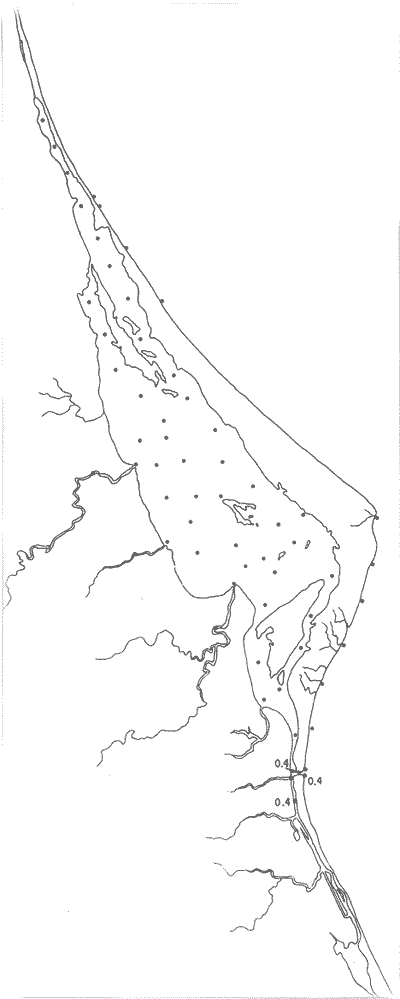 Fig. 39. Porcentajes totales del género Rosalina. 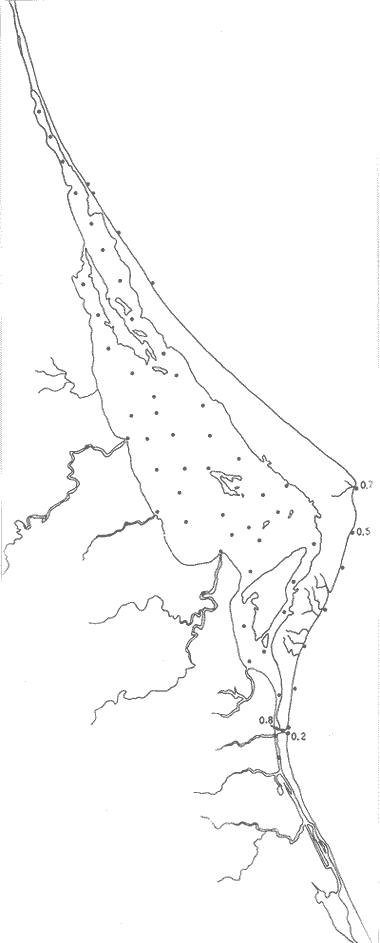 Fig. 40. Porcentajes totales del género Textularia. 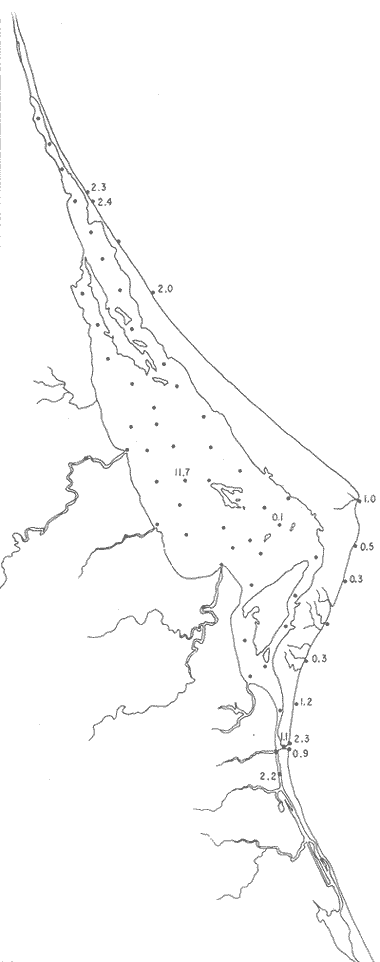 Fig. 41. Porcentajes totales del género Triloculina. Género Triloculina d'Orbigny, 1826
Triloculina oblonga (Montagu)(Lám. 8, figs. 7a-c) Vermiculum oblongus Montagu, 1803, Test. Brit.: 522,lám. 14, f ig. 9. Triloculina oblonga (Montagu). Cushman, 1929, Bull. U.S. natn. Mus., 104 (6): 57, lám. 13, figs. 4-5. Especie muy escasa, presente solamente en la estación YZ-8-64, sin ejemplares vivos (Fig. 41). Hipotipo: CCML-869-Mi. Triloculina sppFue identificada en el interior de la laguna, principalmente en las estaciones cercanas a la Boca de Corazones y en dos estaciones del litoral frente a la barrera arenosa, sin población viviente. Hipotipo: CCML-876-Mi. Triloculina trigonula (Lamarck)(Lám. 8, figs. 8a-d) Miliolites trigonula Lamarck, 1804, Annls. Mus., 3: 351, lám. 47. Triloculina trigonula(Lamarck). Cushman, 1929. Bull. U.S. natn. Mus.,104 (6): 56, lám. 12, figs. 10, 11, lám. 13, figs. 1-2. Escasa, restringida a las estaciones YZ-2-64 e YZ-2364, todas cercanas a la Boca de Corazones, sin ejemplares vivos. Hipotipo: CCML-970-Mi. Género Trochammina Parker y Jones, 1859
Trochammina inflata (Montagu)(Lám. 8, figs. 9a-b) Nautilus inflatus Montagu, 1808, Test. Brit.: 81, lám. 18, fig. 3. Trochammina inflata (Montagu). Parker, Phleger y Peirson, 1953, Spec. Publs Cushman Fdn. foramin. Res., (2): 15, lám. 3, figs. 5, 6. Muy escasa, presente solamente en la estación YZ-5 64, sin ejemplares vivos. Hipotipo: CCML-971-Mi. DISCUSIÓNDel análisis de la distribución de los foraminíferos en la Laguna de Tamiahua, Veracruz, se encontró lo siguiente: 1. La fauna de foraminíferos identificada es característica de lagunas litorales de salinidad media. 2. Se identificaron 42 géneros y 70 especies. 3. Se pudieron establecer 3 biofacies: Biofacies de esteros y ríos, Biofacies lagunar y Biofacies de Golfo Abierto, que reflejan las características hidrográficas y ecológicas del área (Fig. 42). BIOFACIES DE ESTEROS Y RÍOSComprende las zonas influenciadas por las desembocaduras de los esteros de la Laja, Cucharas, Carbajal y Tancochin. La salinidad osciló entre 18.7 y 25.0%, por lo que se le puede considerar como una zona polihalina, corresponde a la zona de menor transparencia del agua, debido al aporte de terrígenos de los esteros y está afectada por la influencia del agua dulce de los mismos. Por su contenido en foraminíferos se le puede definir de la siguiente manera: El número de ejemplares por muestra dentro de las poblaciones totales fluctuó entre 11 y 6,500 observándose que en términos generales las poblaciones son bajas, ya que solamente en las estaciones 30 y 43 se encontraron más de 11000 ejemplares. El número de géneros por muestra fue reducido, entre 2 y 8; las estaciones de foraminíferos oscilaron entre 2 y 11, los foraminíferos aglutinados, estuvieron representados principalmente por los géneros Ammobaculites y Ammoflum, presentando sus mayores porcentajes en algunas de las desembocaduras de los esteros; no se encontraron miliólidos en esta biofacies. 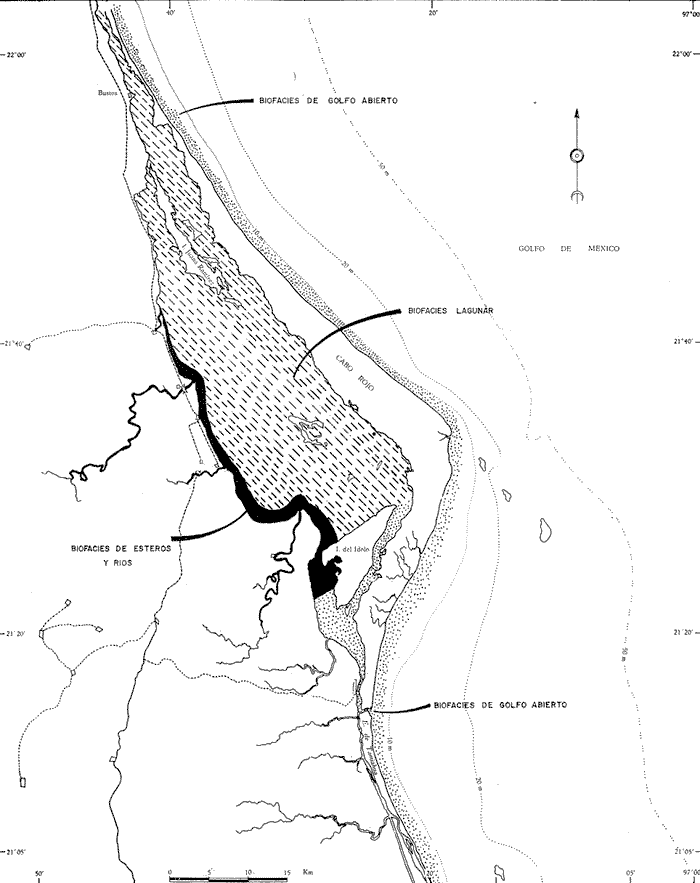 Fig. 42. Biofacies de foraminíferos. El número de ejemplares vivientes dentro de la población, fluctuó entre 4 y 113, por lo que la productividad orgánica relativa en esta biofacies, puede considerarse baja. Sin embargo algunas estaciones como la 18, 19 y 44, presentaron mayor número de ejemplares, lo que sería indicativo de una productividad orgánica relativa mayor en esas áreas. Algo semejante, observó Ayala-Castañares (1963), en la Laguna de Términos, donde el número de ejemplares vivientes tiende a ser mayor en las desembocaduras de los esteros y ríos. Los porcentajes de foraminíferos vivientes dentro de la población total fueron en general bajos, lo que indica una velocidad relativamente lenta de sedimentación. Sólo las estaciones 18 y 44 presentaron porcentajes mayores, lo que sería indicativo de un mayor depósito de sedimentos. Las especies vivientes encontradas en esta biofacies estuvieron representadas principalmente por:Ammonia beccarii y Elphidium gunteri; otras especies menos frecuentes pero no encontradas vivas fueron: Ammobaculites dilatatus, Ammotium salsum, Elphidium advenum, E. incertum mexicanum y E. poeyanum, la mayor parte de estos organismos fueron observados en la Biofacies de Golfo Abierto y se supone que invaden esta biofacies como resultado de la dinámica propia de la laguna. BIOFACIES LAGUNARSe extiende en la mayor parte de la laguna, estrechándose hacia el N y estando delimitada al W por las biofacies de esteros y ríos, y al E por la sub-biofacies lagunar de sotavento, al S colinda con la Isla del Idolo y al N, llega hasta la desembocadura del río Pánuco, por el Canal del Chijol. La salinidad determinada osciló entre 23.30 y 27.35%, por lo que se le considera polihalina. Está afectada en gran parte por las porciones arenosas del sotavento de la gran barrera de Cabo Rojo. Por su contenido en foraminíferos se le puede definir de la siguiente forma: El número total de foraminíferos encontrado en las muestras osciló entre 100 y 12,500 ejemplares; el número de géneros entre 2 y 8 y el número de especies entre 2 y 12. Los foraminíferos aglutinados estuvieron representados principalmente por AmmobaculitesyAmmotium, estos foraminíferos se ha observado que caracterizan lagunas de baja salinidad, los miliólidos casi estuvieron ausentes y sus porcentajes en la mayor parte de las estaciones de esta biofacies no excedieron al 1%, algo semejante observó Ayala-Castañares (1963), en la Laguna de Términos, Campeche. Las poblaciones encontradas de foraminíferos vivientes oscilaron entre 1 y 69 ejemplares por muestra, lo que indicaría para la misma, una productividad orgánica relativa baja, en el tiempo en que se hizo el muestreo. Los porcentajes de foraminíferos vivos dentro de la población total fluctuaron entre 0.1 y 6.4 % de lo que se dedujo, una velocidad de sedimentación lenta para esta biofacies. Las especies más comunes identificadas en esta biofacies, en su mayor porcentaje estuvieron representadas por: Ammonia beccarii y Elphidium gunteri, en menor proporción Elphidium discoidale, Palmerinella palmerae, etc. BIOFACIES DE GOLFO ABIERTOCorresponde a aquellas porciones de la laguna que están influenciadas por las aguas del Golfo de México y son principalmente las playas de Cabo Rojo, la Boca de Corazones y la porción sur de la laguna. La salinidad de esta biofacies es típicamente ultrahalina, mayor de 30%. Por su contenido en foraminíferos se define de la siguiente forma: El número de ejemplares por muestra dentro de las poblaciones totales fluctuo entre 300 y 17,000, predominando poblaciones mayores de 1,000. El número de géneros osciló entre 3 y 31, el número de especies entre 5 y 54. El porcentaje de foraminíferos aglutinados entre 0.3 y 3.9%. Los porcentajes de miliólidos son los más altos fluctuando entre 11 y 41 %, observándose que los porcentajes mayores se encuentran en la Boca de Corazones y en las muestras colectadas sobre el litoral de la barrera arenosa de Cabo Rojo; entre las especies identificadas son de mencionarse: Quin taqueloculina compta, Q. poeyana, Q. seminulum y Q. wiesneri. Las poblaciones vivientes en número de ejemplares por muestra encontrados en esta biofacies oscilaron entre 2 y 48, observándose que en general son bajas, de lo que se puede deducir, que la productividad orgánica relativa es baja para la misma. Se observó también que los porcentajes de ejemplares vivos no son significativos, pues fluctuaron entre 0.1 y 0.5, lo que sería una evidencia de depósito relativamente lento de sedimentos para esta región. Junto con las especies ya mencionadas de miliólidos, otros foraminíferos que caracterizan a esta biofacies, pero en menor proporción fueron: Ammonia beccarii, E. gunteri, E. spp., Hanwazaia concentrica, etcétera. Ecológicamente las aguas de la Laguna de Tamiahua parecen corresponder en muchos aspectos a los de la porción W en la Laguna de Términos en Campeche, la productividad orgánica relativa parece ser la más baja de las diversas áreas estudiadas, incluso aún la de la Laguna Madre, esto parece indicar que en la fecha de 1964 en que se hizo el trabajo de campo, la ecología de la región hubiera sufrido un desequilibrio cuyas causas se desconocen. AgradecimientosLos autores agradecen la ayuda de las siguientes personas e instituciones: G. P. Salas (ex-director del Instituto de Geología, UNAM); F. B Phleger (Institución Scripps de Oceanografía, Universidad de California, San Diego); A. Yáñez y R. Cruz (Instituto de Geología, UNAM); A. Madrid, S. Cavazos y R. Ávila (Gerencia de Exploración de Petróleos Mexicanos); y a la National Science Foundation de los Estados Unidos de América (con cuya aportación NSF-3420 se llevó a cabo parte de la investigación). LITERATURAAKERS, W. H., General ecology of the Foraminiferal genus Eponidella with description of a Recent Species. J. Paleont.,1952.645-649.26 (4): ÁLVAREZ DEL VILLAR, J., EstudioPeces Mexicanos (Claves). Sría. Ind. y Comercio, Inst. Nal. Inv. Biol. Pesq., ser. Invest. Pesq.,1970. 166 p. (1): ANDERSEN, H. V., Bull. geol. Surv. La., Foraminifera of the mudlumps, Lower Mississippi River Delta. 1961. 1-208.(35): AYALA-CASTAÑARES, A., Sistemática y distribución de los Foraminíferos Recientes de la Laguna de Términos, Campeche, México. Univ. Nal. Autón. México. Inst. Geol. Bol., 1963. 1-130 p.67 (3): AYALA-CASTAÑARES, A. R. CRUZ A. GARCÍA-CUBAS, Jr L. R. SEGURA Lagunas Costeras. Un Simposio. Mem. Simp. Intern. Lagunas Costeras. Síntesis de los conocimientos sobre lo Geología Marina de la Laguna de Tamiahua, Veracruz, México. In: Ayala-Castañares, A. y F. B Phleger (Eds.) UNAM-UNESCO,1969.39-48. nov. 28-30, 1967: AYALA-CASTAÑARES, A. F. B PHLEGER Bull. Am. Ass. Petrol. Geol.,Process and History of Terminos Lagoon, Mexico.1971. 2130-2140.55 (12): AYALA-CASTAÑARES, A. L. R. SEGURAEcología y distribución de los Foraminíferos Recientes de la Laguna Madre, Tamaulipas, México, Univ. Nal. Autón. México. Inst. Geol. Bol. 1968.1-89. (87): BENAVIDES G., L., Symposium sobre yacimientos de Petróleo y Gas. XX Congr. Intern. Notas sobre la Geología Petrolera de México. In: Guzmán, E. J. (Ed.). 1956.351-562. (3): BERMUDEZ, P. J., Un género y especie nueva de Foraminíferos vivientes de Cuba. Mems. Soc. cub.1934. _____, Spec.Publs Cushman Lab. foramin. Res., Tertiary Smaller Foraminifera of the Dominican Republic. 1949. 1-322. (25): BORNEMANN, J. G., Die mikroskopische Fauna des Septarienthones von Hermsdorf bei Berlin. Z. dt. geol. Ges., 1855. 307-371.7: BRAVO, H. L. S. SALAZAR A. VÁZQUEZ-BOTELLO E. F. MANDELLI Bull. Environ. Contam. Toxicol., Polyaromatic Hydrocarbons in Oysters from Coastal Lagoons along the Eastern Coast of the Gulf of Mexico, Mexico. 1978. 171-176.19 (2): BRADY, H. B., Ann. Mag. nat. Hist. ser, The Ostracoda and Foraminifera of Tidal Rivers. In: Brady G. S. y D. Robertson (Eds.). 1870.273-3096 (34): BRADY, H. B., Report on the Foraminifera dredg ed by H. M. S. during the years 1873-1876. Rep. Sci. Results Voy. H. M. S. CHALLENGER Zool.,1884.1-814 9: COLE, W. S., Bull. Fla St. geol. Surv. The Pliocene and Pleistocene Foraminifera of Florida. 1931. 1-79. (6): CRUZ, R., Tesis Prof. Esc.Sup. Ing. y Arq. Inst. Pol. Nal.,Sedimentologia de la Laguna de Tamihual Veracruz, México. 1966. 100 p. _____, Geología Marina de la Laguna de Tamiahua, Veracruz, México.Univ. Nal. Autón. México. Inst. Geol. Bol. 1968.1-47. 88: CUSHMAN, J. A., Bull. U.S. geol. Surv. Some Pliocene and Miocene Foraminifera of the Coastal Plain of the United States. 1918a5-38. (676): _____, Bull. U.S. geol. Surv. Some Miocene Foraminifera of the Coastal Plain of the United States.1918b.39-100. (676): _____, Fossil Foraminifera from the West Indies. Carnegie Inst. Wash. Publ. 1919.21-71. (291): _____, Shallow-water Foraminifera of the Tortugas Region. Carnegie Inst. Wash. Publ., 1922.1-85.17 (311): _____, Bul. U.S. natn. Mus. The Foraminifera of the Atlantic Ocean. Part 6, Miliolidae, Ophthalmidiidae and Fischerinidae. 1929.129 p. (104): _____, Bull. U.S. natn. Mus. The Foraminifera of the Atlantic Ocean. Part. 7, Nonionidae, Camerinidae, Peneroplidae and Alveolinellidae, United States. 1930. 79 p. (104): _____, Fourteen New Species of Foraminifera. Smithson. misc. Collns., 1935.1-9.91 (21): _____, A monograph of the Foraminiferal Family Nonionidae. Prof. Pap. U.S. geol. Surv. 1939.1-100. (191): _____, Spec. Publs Cushman Lab. foramin. Res.Foraminifera from the shallow water of the New England Coast. 1944.1-37. (12): _____, New Species and varieties of Foraminifera from off the Southeastein Coast of the United States.Contr. Cushman Lab. foramin. Res., 1947. 86-92.23 (4): CUSHMAN, J. k. P. BRÕNNIMANN Some new genera and Species of Foraminifera from Brackish water of Trinidad. Contr. Cushman Lab. foramin. Res., 1948a.15-21.24 (1): _____, Additional new Species of Arenaceous Foraminifera from shallow waters of Trinidad. Contr. Cushman Lab. foramin. Res., 1948b.37-42.24 (2): DE BUEN, F., Crisis Ostrícolas en México y su recuperación. Sría. Marina, Dir. Gral. Pesca eInd. Conexas: 1957.1-46. FICHTEL, L. von S. P. C. von MOLL Testacea microscopica, aliaque minuta exegeneribus Argonauta et Nautilus, ad naturam delincata et descripta 4to. Wien: 123. 1798. GARCÍA CUBAS, A., Diccionario Geográfico, Histórico y Biográfico de los Estados Unidos Americanos. Of. Tipograf. Sría. Fomento, México, 1891. 1-567. 5: GARCÍA-CUBAS, A. Jr., Ecología y distribución de los micromoluscos recientes de la Laguna de Tamiahua, Veracruz, México. Univ. Nal. Autón. México. Inst. Geol. Bol. 1969.1-80. 91: GARCÍA, E., Inst. Biol. An., Los climas del Estado de Veracruz. Univ. Nal. Autón. México. 1970.3-42.41 (1): GONZÁLEZ-MEDRANO, F., Tesis Prof. Fac. Cienc., Univ. Nal. Autón. México, La Vegetación del Nordeste de Tamaulipas. 1966. 57 p. HUMM, H. J. H. H. HILDEBRAND Marine Algae from the Gulf Coast of Texas and Mexico. Publ. Inst. Mar. sci. 1962. 227-268.8 (1): KORNFELD, M. N., Recent littoral Foraminifera form Texas and Lousiana. Contr. Dep. Geol. Stanford Univ., (3):1931.77-101.1 LAMARCK, J. B. P. A. M., Annls Mus., Suite des memoires sur les fossiles des environs de Paris.1804. 179-180, 237-245, 349-357. 5: LE ROY, L. W., Some small Foraminifera, Ostracoda and Otholiths from the Neogene (Miocene) for the Rokan Tapanoeli Arca, Central Sumatra. Natuurk. Tijdschr. Ned., Indie, 1939. 215-296.99: LINNAEUS, S., Systema Naturae per regna tria naturae. 10 Ed. Stockholm, 1758.824 p. LOEBLICH, A. R. Jr. H. TAPPAN Studies of Arctic Foraminifera. Smithson. misc. Collns., 1953. 11-50.121 (7): McINTIRE, W. C. Ho Lagunas Costeras, Un Simposio. Mem. Simp. Intern. Lagunas Costeras. Development of Barrier Island Lagoons Western Gulf of Mexico. In: Ayala-Castañares, A. y F. B. Phleger (Eds.). UNAM-UNESCO 1969. 49-62. nov. 28-30, 1967: MONTAGU, G., Testacea Britannica, of natural history of British shells marine, and a freshwater, including the most minute, printed by J. S. Hollis, England, 1803. 606 p _____, Testacea Britannica Suppl., 1808.81 p. MUÑOZ-LUMBIER, M., Algunos datos sobre las islas mexicanas para contribuir al estudio de sus recursos naturales. Univ. Nal. Autón. México. Inst. Geol. Bol., 1919.7:54. MUIR, J. M., Geology of the Tampico Region, Mexico. Am. Ass. Petrol. Geol. Tulsa., Oklahoma,1936. 280 p. NÚÑEZ-ORTEGA, A., Boln. Soc. Geogr. Estadist. Repúb. mex., Un viaje a la Laguna de Tamiahua. 2a. Época, 1869. 734-737.1: ORBIGNY, A. d', Annls. Sci. nat., Tableau Méthodique de la Classe des Céphalopodes. 1826.245-314. 7: _____, Voyage dans l'Am. Mérid. Paris, Strasb., 1839a.1-86.5 (5): _____, In: De la Sagra Hist. Phys. Pol. Nat. Île de Cuba ''Foraminíferos''. A. Bertrand Ed. Paris, 1839b.1-224. 8: PARKER, F. L., Bull. Mus. comp. Zool. Harvard., Foraminifera Species of Porstmouth, New Hamphshire.1952.391423.106 (9): _____, Gulf of Mexico, its origin waters, and marien life. Fishery Bull. Fish. Wildlife Serv. Gulf of Mexico Foraminifera: In: Galtsoff, P. S. (Ed.) 1954a. 235-241. (89): _____, Bull. Mus. comp. Zool. Harvard,Distribution of the Foraminifera in the North eastern Gulf of Mexico. 1954b.453-588.111 (10): _____, Eastern Mediterranean Foraminifera. Rep. Swed. deep Sea Exped., 1958.255.8 (4): _____, Quinqueloculina tenagos, new name for Quinqueloculina rhodiensis Parker, preoccupied. Contr. Cushman Fdn foramin. Res., 1962.110.13 (3): PARKER, F. L. W. D. ATHEARN Ecology of Marsh Foraminifera in Poponesset Bay Massachusetts. J. Paleont., 1959.333-343.33 (2): PARKER, F. L F. B PHLEGER J. F. PEIRSONSpec. Publs Cushman Fdn foramin Res. Ecology of Foraminifera from San Antonio Bay and environs, Southwest Texas. 1953. 1-75. (2): PARR, W. J., Foraminifera B.A.N.Z., Antarctic Res. Exped. 1929-1931, Rep. ser. B., 1950.232-292.5 (6): PHLEGER, F. B AYALA-CASTAÑARES Lagunas Costeras, Un Simposio. Mem. Simp. Intern. Lagunas Costeras. Marine Geology of Topolobampo Lagoons, Sinaloa, Mexico. In: AyalaCastañares, A. y F. B. Phleger (Eds.). UNAM-UNESCO, 1969. 39-48.nov. 28-30, 1967: PHLEGER, F. B F. L. PARKER Ecology of Foraminifera Noth west Gul of Mexico, Part. II, Foraminifera Species. Mem. geol. Soc. Am., 1951. 1-64.16 (2): POGGIE, J. jr., Coastal Pioneer Plant sand Habitat in the Tampico Region, Mexico. Tech. Rep. cstl Stud. Inst. La St. Univ., Contr., 1962.1-60.62 (1): RAMÍREZ-HERNÁNDEZ, E., An. Inst. Nal. Invest. Biol. Pesq., Estudios preliminares sobre los peces Marinos de México.1965.257-292. 7: RESENDEZ M., A., An. Inst. Biol., Estudio de los peces de la Laguna de Tamiahua, Veracruz, México. Univ. Nal. Autón. México. ser. Cienc. del Mar y Limnol.,1970.79-146. 41 (1): ROSEN, D. E., Bull. Fla St. Mus., Middle-American Poecillid fislies of the genus Xiphophorus. 1960. 1-242.5 (4): RHUMBLER, L., Die Foraminiferes (Thalamophoren) der Plankton Exped., etc., Pt. I Systematik. Ergebn. PlanktonExped. Humboldt., 1911. 1-33 y 333-476. 3 y 3 (1): y (2): SEGURA, L. R., Sistemática y distribución de los foraminíferos litorales de la "Playa Washington", al sureste de Matamoros, Tamaulipas, México. Univ. Nal. Autón. México. Inst. Geol. Bol.1963. 1-92. 68: SEVILLA, M. L. E. C. MONDRAGÓN An. Inst. Nal. Invest. Biol. Pesq. Desarrollo gonádico de Crassostrea virginica Gmelin, en la Laguna de Tamiahua. 1965. 53-69. (1): TAMAYO, J. L., Geografía General de México.Inst. Mexicano Invest. Econ., 1962. vols. 1-5. TERQUEM, O., Les Foraminiferes et les Entomostraces Ostracodes du Pliocene Superior de I'lle de Rhodes. Mem. Soc. geol. Fr. ser., 1878.1-133.3 (1): VILLALOBOS, F., A. et al, Informe Final de las investigaciones realizadas en la Laguna de Tamiahua. Univ. Nal. Autón. México.Inst. Biol. (Informe inédito). 1968. WANTLAND, K. F., Recent Benthonic Foraminifera of the British Honduras She1f. Ph. D. Dissert.Rice Univ., Houston, Tex. 1967. 300 p. LÁMINAS
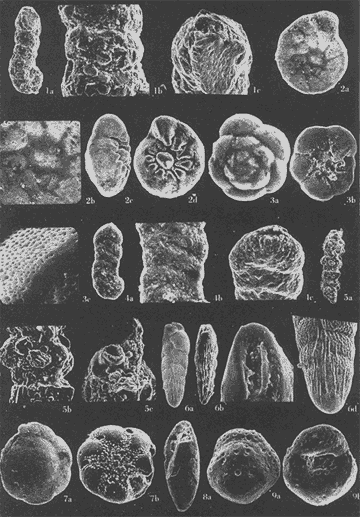 Lám. 1. Figs. 1a, b, c. Ammobaculites dilatatus Cushman y Brönnimann. Muestra YZ-23-64. Hipotipo CCML-7-Mi. Aprox. X 60. Figs. 2a, b, c, d. Ammonia beccarii (Linnaeus) variante A. Muestra YZ-2-64 Hipotipo CCML-27-Mi. Aprox. x 30. Figs. 3a, b, c. Ammonia beccarii (Linnaeus) variante B. Muestra YZ-24-64. Hipotipo CCML-34-Mi. Aprox. x 28. Figs. 4a, b, c. Ammotulm salsum (Cushman y Brönnimann). Muestra YZ-27-64. Hipotipo CCML-55-Mi. Aprox. x 45. Figs. 5a, b, c. Bigenerina sp. Muestra YZ-2-64. Hipotipo CCML-924-Mi. Aprox. x 119 Ammotuint salsum (Cushman y Brönnimann). Muestra YZ-27-64.64. Hipotipo CCML-928-Mi. Aprox. x 54. Figs. 7a, b. Buccella hannai (Phleger y Parker). Muestra YZ-2-64, Hipotipo CCML 927-Mi. Aprox. x 32. Figs. 8a. Buliminella. elegantissima (d'Orbigny). Muestra YZ-24-64. Hipotipo CCML-926-Mi. Aprox. x 25. Figs. 9a, b .Discorbis bulbosa Parker. Muestra YZ-2-64. Hipotipo CCML-930-Mi. Aprox. x 28. 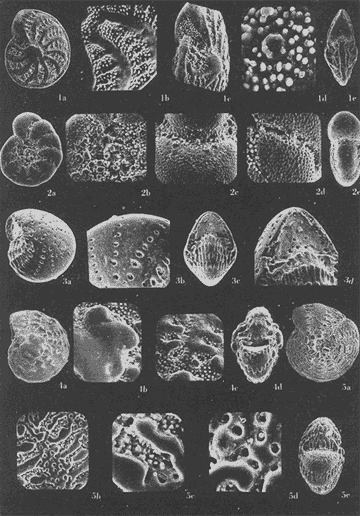 Lám. 2. Figs. 1a,b,c,d,e. Elphidium advenum (Cushman). Muestra YZ-23-64. Hipotipo CCML-931-Mi. Aprox. x 25. Figs. 2a,b,c,d,e. Elphidium delicatulum Bermúdez. Muestra YZ-3-64. Hipotipo CCML-258-Mi. Aprox. x 42. Figs. 3a,b,c,d. Elphidium discoidale (d'Orbigny). Muestra YZ-2-64. Hipotipo CCML-259-NE. Aprox. x 67. Figs. 4a,b,c,d. Elphidium galvestonense Kornfeld. Muestra W-2-64. Hipotipo CCML-279-Mi. Aprox. x 34. Figs. 5a,b,c,d. Eponides antillarum (d'Orbigny). Y71-2-64, Hipotipo CCML-932-Mi, Aprox. x 56. 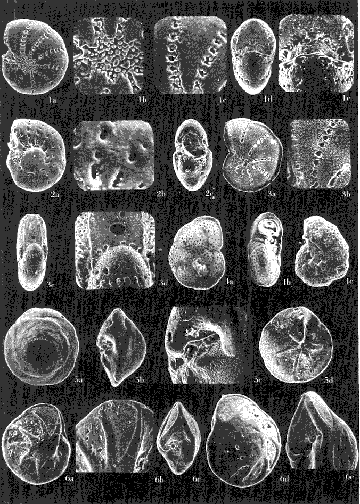 Lám. 3. Figs. 1a,b,c,d,e Elphidium incertum mexicanum Kornfeld. Muestra YZ-2-64. Hipotipo CCML-934-Mi. Aprox. x 54. Figs. 2a,b,c. Elphidium cf. E. koeboeense Le Roy. Muestra YZ-2-64. Hipotipo CCML-933-Mi. Aprox. x 52. Figs. 3a,b,c,d. Elphidium poeyanum (d'Orbigny). Muestra YZ-24-64. Hipotipo CCML-344-Mi. Aprox. x 22. Figs. 4a,b,c. Eponidella gardenislandensis Akers. Muestra YZ-23-64. Hipotipo CCML-363-Mi. Aprox. x 34 Figs. 5a,b,c,d. Eponides antillarum (d'Orbigny). Muestra YZ-2-64. Hipotipo CCML-935-Mi. Aprox. x 103. Figs. 6a,b,c,d,e. Eponides repandus (Fitchell y Moll). Muestra YZ-2-64. Hipotipo CCLM-374-Mi Aprox. x 55 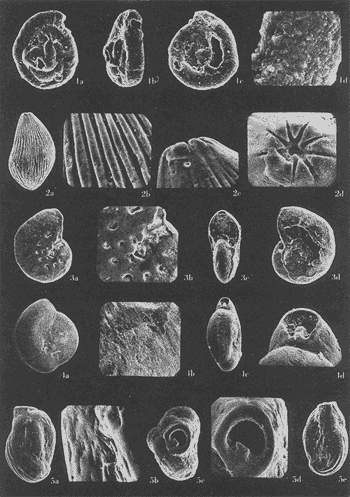 Lám. 4. Figs. 1a, b, c, d. Glomospira sp. Muestra YZ-25-64. Hipotipo CCML-430-Mi. Aprox. x 31. Figs. 2a,b,c,d. Guttulina australis (d'Orbinyg). Muestra YZ-23-64. Hipotipo CCML-431-Mi. Aprox. x 67. Figs. 3a, b, c, d. Hanzawaia concéntrica (Cushman). Muestra YZ-2-64. Hipotipo CCML-457-Mi. Aprox. x. 79. Figs. 4a, b, c, d. Massilina peruviana (d'Oibigny). Muestra YZ 23-64. Hipotipo CCML-942-Mi. Aprox. x 67. Figs., 5a, b, c, d, e. Massilina protea Parker. Muestra YZ-8-64. Hipotipo CCML 943-Mi. Aprox. x 71. 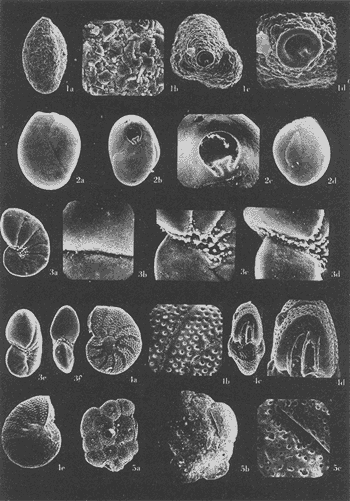 Lám. 5. Figs. 1a, b, c, d. Miliammina fusca (H. B. Brady). Muestra YZ-23-64. Hipotipo CCML-944-Mi. Aprox. x 52. Figs. 2a, b, c, d. Miliolinella circularis (Bornemann. Muestra YZ-12-64. Hipotipo CCML-945-Mi. Aprox. x 67. Figs. 3a, b, c, d, e, f. Nonionella atlantica Cushman. Muestra YZ-23-64. Hipotipo CCML-544-Mi. Aprox. x 42. Figs. 4a,b,c,d,e. Palmerinella palmerae Bermúdez. Muestra YZ-12-64. Hipotipo CCML-625-Mi. Aprox. x 56. Figs. 5a, b, c. Planorbulina mediterranensis d'Orbigny. Muestra YZ-2-64. Hipotipo CCML-954-Mi. Aprox. x 82. 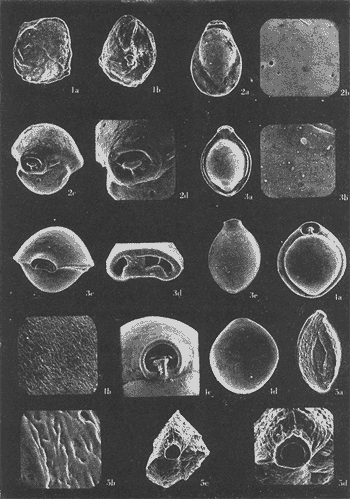 Lám. 6. Figs. la, b. Poroeponides cf.P. lateralis (Terquem). Muestra YZ-2-64. Hipotipo CCML-1331-Mi. Aprox. x 47. Figs. 2a,b,c,d. Pyrgo cf. P. elongata (d'Orbigny). Muestra YZ-2-64. Hipotipo CCML-949-Mi. Aprox. x 47. Figs. 3a,b,c,d,e. Pyrgo cf. P. nasutus Cushman. Muestra YZ-2-64. Hipotipo CCML-951-Mi. Aprox. x 447. Figs. 4a,b,c,d. Pyrgo subsphaerica d'Orbigny. Muestra YZ-2-64. Hipotipo CCML-952-Mi. Aprox. x 32. Figs. 5a,b,c,d. Quinqueloculina compta Cushman. Muestra YZ-2-64. Hipotipo CCML-957 Mi. Aprox, x 73. 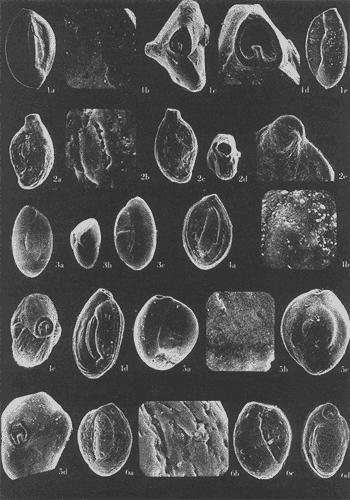 Lám. 7. Figs. 1a, b, c, d, e. Quinqueloculina lamarckiana d'Orbigny. Muestra YZ-23-64. Hipotipo CCML-959-Mi. Aprox. x 47. Figs. 2a, b, c, d, e. Quinqueloculina poeyana d'Orbigny. Muestra YZ-23-64. Hipotipo CCML-958-Mi. Aprox. x 39. Figs. 3a, b, c. Quinqueloculina seminulum (Linnaeus). Muestra YZ-23-64. Hipotipo CCML-961-Mi. Aprox. x 102. Figs. 4a, b, c, d. Quinque loculina tenagos Parker. Muestra YZ-2-64. Hipotipo CCML-962-Mi. Aprox. x 54. Figs. 5a, b, c, d. Quinqueloculina vulgaris d'Orbigny. Muestra YZ-1-64. Hipotipo CCML-728-Mi. Aprox. x 53. Figs. 6a, b, c, d. Quinqueloculina wiesneri Parr. Muestra YZ-23-64. Hipotipo CCML-963-Mi. Aprox, x 39. 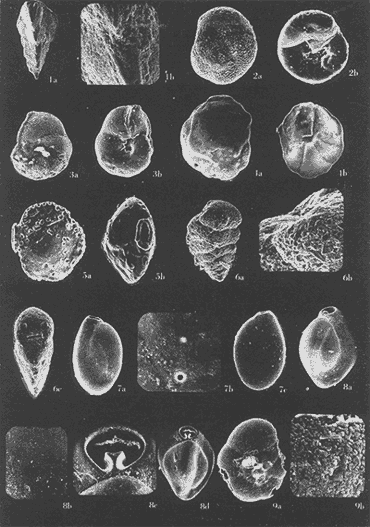 Lám. 8. Figs. 1a, b. Reussella atlantica Cushman. Muestra YZ-2-64. Hipotipo CCML-967-Mi. Aprox. x 55. Figs. 2a, b. Rosalina cf. R. concinna (H. B. Brady). Muestra YZ-23-64. Hipotipo CCML-965-Mi. Aprox. x 59. Figs. 3a, b. Rosalina floridana (Cushman). Muestra YZ23-64. Hipotipo CCML-964-Mi. Aprox. x 41. Figs. 4a, b. Rotalia translucens Phleger y Parker. Muestra YZ-2-64. Hipotipo CCML-968Mi. Aprox. x 53. Figs. 5a, b. Siphonina pulchra Cushman. Muestra YZ-2-64. Hípotipo CCML-969-Mi. Aprox. x 88. Figs. 6a, b, c. Textularia mayori Cushman. Muestra YZ-23-64. Hipotipo CCML-845-Mi. Aprox. x 90. Figs. 7a, b, c. Triloculina oblonga d'Orbigny. Muestra YZ-8-64. Hipotipo CCML-869-Mi. Aprox. x 59. Figs. 8a, b, c, d. Triloculina trigonula (Lamarck). Muestra YZ-2-64. Hipotipo CCML-970-Mi. Aprox. x 52. Figs. 1a, b. Trochammina inflata (Montagu). Muestra YZ-3-64. Hipotipo CCML-971-Mi. Aprox. 51.
|