|
MODERN OSTRACOD SPECIES
DIVERSITY, DOMINANCE, AND BIOFACIES PATTERNS VERACRUZ-ANTON LIZARDO REEFS,
MEXICO
Trabajo recibido el 25 de julio de 1979 y aceptado para su
publicación el 21 de febrero de 1980.
PAUL R. KRUTAK
Department of Geology, University
of Nebraska, Lincon, Nebraska 68508. Part of the field expenses for this study
were defrayed by a Senior Summer Research Fellowship (1972) awarded by the
Research Council of the University of Nebraska, USA.
SUE E. RICKLES
Amoco Production Co. Denver, Colorado 80206, USA.,
RAÚL GÍO-ARGÁEZ
Universidad Nacional
Autónoma de México, Instituto de Geología, México
20, D. F.
Los corales
escleractineos que se desarrollan enfrente de la Ciudad de Veracruz y la Punta
Antón Lizardo son bastantes extensos. Están separados en dos
ambientes deposicionales notablemente diferentes, distantes entre si por
escasos 20 km. Esta separación deposicional está dada por la
desembocadura del Río Jamapa. El análisis de las especies
dominantes de 33 muestras colectadas en estos arrecifes, indican la presencia
de dos diferentes biofacies. Loxocorniculum
tricornatumdomina el área de Veracruz mientras
queL. cf.L. postdorsoalata caracteriza
las estaciones de Antón Lizardo. Dos especies de
ostrácodosL. tricornatumyMorkhovenia
insconspicua,son cosmopolitas y se presentan en las 33 estaciones.
Las especies poco comunes que se encuentran en los dos complejos arracifales
están generalmente restringidas a los arrecifes de Veracruz. El
análisis de diversidad simple (número de especie/estación)
demuestra que el área de Veracruz es más compleja que la de
Antón Lizardo, sin embargo, al utilizar el método de
Shannon-Weiner (Valores en función de equivalencia) indican que las
diferencias de diversidad son mínimas y no reflejan las
características ambientales observadas. De este modo, los datos de
distribución más simples reflejan mejor el contraste observado en
el campo, principalmente por aguas turbias cargadas de sedimento en el complejo
Antón Lizardo y claras en el complejo de Veracruz.
Scleractinian patch reefs living off Veracruz and Anton Lizardo are
extensive. They are separated into two strikingly different depositional
environments by the Rio Jamapa, yet lie only 12 miles apart. Analysis of the
dominant ostracod species in 33 samples collected on the reefs indicates two
distinct biofacies exist.Loxocorniculum tricornatumdominates
the Veracruz group; whereas,L. cf.L.
postodorsoalatacharacterizes the Anton Lizardo stations. Two ostracod
species, L. tricornatumandMorkhovenia
inconspicua, are cosmopolitan, and occur at all 33 sampling stations.
Rare species living on the two reef complexes are generally restricted to the
Veracruz complex. Simple diversity analysis (# of species per station)
demonstrates that the Veracruz reefs are more diverse than those of Anton
Lizardo. However, Shannon-Weiner information function and equitability values
indicate that diversity differences are minimal, and do not reflect the
observed environmental differences. Thus the more simple distributional data
better reflects the constrasts in depositional environments observed in the
field (primarily more turbid, sediment -laden waters on the Anton Lizardo
complex- vice-versa on the Veracruz
complex).
Only three modern scleractinian reefs exist in the western. Gulf of Mexico: (1) Flower Garden Banks, (2) the Isla de Lobos group, and (3) the Veracruz-Anton Lizardo complex. Study of these living reefs, and others like them, has been the key to the recognition of similar structures in the fossil record. Fossil reefs have enormous hydrocarbon potential (Pugh 1950, Friedman 1969). Therefore many modern and ancient reefs have been studied (Leblanc and Breeding 1957, Frostet al.1977). Benthic ostracods are part of the microbiota of such reefs and make up a sizable fraction of the skeletal components of the sand-sized fraction. The habitat of these microscopic crustaceans makes them extremely valuable as facies indicators and as potential indices of depositional environments. Furthermore, since they are microscopic, they can be easily recovered from well cuttings and cores. Nevertheless, few publications exist on modern reefal ostracods. Patch reefs growing in the Gulf of Mexico offshore from the port of Veracruz and the village of Anton Lizardo, Mexico, occur in a unique depositional environment. Here corals not only live, but flourish in some locations despite their proximity to terrigenous sediments. This paper will outline the modern ostracod species diversity, dominance, and biofacies patterns of these reefs. The authors feel that shelf margin reefs, like these, may serve as models for predictable trends and facies patterns in fossil limestone oil and gas reservoirs, and hope that this paper will provide a baseline for future studies of their ostracod faunas. PREVIOUS WORKBold (1977, Fig. 2, p. 177) outlined the distributíon of ostracod biogeographic provinces in Recent sediments of the Gulf of Mexico and the Caribbean, and indicated a "transition zone" between the Gulf and Caribbean. This zone consists of the shallow carbonate platforms between Florida, the Bahamas, Cuba, and Yucatan. Bold (1977, Fig. 1, p. 176) pointed ou that only one Recent sediment sample (Miskito, Keys off Nicaragua) has been examined from the area between the Goajira Peninsula and Panama. Teeter (1975) described in detail the modern ostracods from the barrier reef off Belice. These two widely spaced localities represent the only two published works known to the writers in the northwestern Caribbean Basin, although Palacios-Fest (1975) summarized the distribution of bairdiacean ostracods in his professional thesis at Universidad Nacional Autonoma de Mexico. It is clear that the western Caribbean ostracod fauna is very poorly understood. Maddocks (1974) presented a comprehensive bibliography of ostracod studies made in the Gulf of Mexico. She also described, in reconnaissance, the ostracod fauna of the Flower Garden Banks. The West and East Flower Garden Banks represent the northern limit of flourishing coral-reef growth in the Gulf of Mexico (Rezak 1977, p. 27). There is only one other living scleractinian reef complex between the Flower Garden Banks (lat. 27º52.6'N.) and the Veracruz-Anton Lizardo complex (lat. 19º10'N.). The Isla de Lobos grup, offshore from Cabo Rojo (lat. 21º30'N.), was studied by Rigby and Mcintire (1966), but nothing is known of its ostracod fauna. Two major complexes of coral reefs (total of 23 emergent coral reefs and islands-Edwards 1969, p. 1) occur in the Gulf of Mexico at latitude 19º10'N. and longitude 96º10'W. (Fig. l). The northernmost group, designated here as the Veracruz complex, lies off the coast from the port of Veracruz, Mexico. A second group, the Anton Lizardo complex, occurs 12 miles (7 nautical miles) to the southeast. Together these reefs form the largest scleractinian development in the western Gulf of Mexico (Edwards 1969, p. 1). The two complexes lie northwest and southeast, respectively, of the mouth of one of two major rivers influencing sediment deposition in the area, the Rio Jamapa. Rio Jamapa descends through the foothills of the Sierra Madre Oriental approximately 50 miles west of Veracruz, and carries andesitic sediments derived from Mount Orizaba, an extinct composite volcanic cone (Krutak 1974, p. 11; Morelock and Koenig 1967, p. 1001). The terrigenous volcaniclastics are deposited in the Gulf of Mexico near Boca del Rio, a small village situated about halfway between Veracruz, and Anton Lizardo. Rio Papaloapan also affects sedimentation in the reef complex area. It descends through Precambrian and Paleozoic igneous, sedimentary, and metamorphic terranes from 200 km. to the west, in the Sierra de Juarez (Freeland 1971, pp. 33, 37; Edwards 1969, p. 8) . The river enters Alvarado Lagoon near the town of the same name about 24 miles southeast of the headland of Anton Lizardo. SEDIMENTATION PATTERNS - HYPOTHESES AND ASSUMPTIONSVery little is known about bottom currents in the area. Rio jamapa sediments may be carried southeast by strong currents generated during prevailing winter winds ("Los Nortes" from a north-northeasterly direction) , and by spring and summer trade winds blowing, from the northeast (Edward 1969, p. 6). Such directional transport seems likely, as no delta exists at Boca del Rio. If this is in fact true, these sediments would be deposited between, and on the flanks of the Anton Lizardo Reefs. Edwards (1969, p. 40) hypothesized that sediments deposited in the Gulf from Rio Papaloapan are transported in northerly direction by prevailing longshore drifts, and deposited around the Anton Lizardo headland. This is partially borne out by Holocene beach ridges seen on reflected infra-red satellite photographs. We assume, therefore, that sediments from the Papaloapan source terrance are also deposited on and around the Anton Lizardo reef complex. Assuming both cases above are true, then terrigenous sedimentation rates on the Anton Lizardo reefs are significantly faster than such rates on the Veracruz reefs, which are relatively uncontaminated by terrigenous debris. It follows that the two reefs, complexes exist, in strikingly different depositional enviroments, although they lie only 12 miles apart. Field observations bear this out. The Anton Lizardo complex, with its relatively more turbid, sediment-laden waters, has a poorly developed scleractinian fauna. Underwater observations on these reefs indicate they are being choked by large amounts of river-borne sediments. This is further substantiated by Morelock and Koenig (1967, p. 1003) and Emery (1963, p. 13). They recorded the percentages of calcium carbonate present in sediments between the reefs and on the reefs slopes and beaches of Anton Lizardo and Veracruz respectively. Percentages are significantly lower in the Anton Lizardo group than in comparable areas of the Veracruz group. The lower percentages in sediments adjacent to the reefs buildups at Anton Lizardo indicate that terrigenous deposition in taking place more rapidly there than at Veracruz. Krutak (1977) noted that trubidity increased significantly in the Anton Lizardo area during summer river floods. Conditions here appear to be marginal for coral growth and survival. On the other hand, inter-reef sediments of the Veracruz complex contain higher calcium carbonate percentages, and underwater observaations by Krutak in 1972 indicate turbidity is significantly lower. Typical West Indian corals (Acropora palmata, A. cervicornis, Porites sp., Diploria strigosa, Montastrea annularis)flourish here, and several lagoonal, reefs, and surrounding deeps-water habitats and communities may be differentiated. Flourishing growth of scleractinian framebuilders and the associatedThalassia-Halimeda community is characteristic. Conditions certainly appear better for coral growth and survival than at Anton Lizardo. These inferred differences in sedimentation and ecology between Anton Lizardo and Veracruz reef complexes suggest that differences should exist in the sediment microbiota. Lidz and Lidz (1966), using foraminifera, established biofacies on the Veracruz reefs, but did not note the differences in sedimentation regimes. To date, no studies have made use of the common ostracods living on these two reef complexes. The purpose of this paper is to determine whether or not the ostracod populations actually do, in any way, reflect the physical differences observed in the field. STUDY AREA AND COLLECTING PROCEDURESReefs at Veracruz-Anton Lizardo occur in two distinct groups (Fig. 1). The northernmost group lies off the port of Veracruz and consists of seven major reefs: Gallega, Blanquilla, Hornos, Pajaros, Isla Sacrificios, Isla Verde, and Anegada de la Adentro. Southeast, offshore from Anton Lizardo, are Giote, Chopas, Salmedina, Rizo, Enmedio, Cabeza, Anegadillo, Santiaguilla, and Anegado de Afuera. The reefs rise to the surface from a very gently sloping shallow shelf less than 25 fathoms deep, and are oriented northwest-southeast parallel to the shoreline (Morelock and Koenig 1967, P. 1101). During July and August 1972, Krutak collected sediment samples from 33 stations on reefs of both complexes (Fig. 2). Twenty five of these samples were from reefs of the Veracruz group and eight were taken on three of the nine reefs composing the Anton Lizardo group. A 3.5 cm. inside diameter Phleger core tube 0.30 m. long was used for sampling. Taking actual vertical cores on the reefs was impossible as sediment cover over the hard coral frame-work rarely exceeded 15 cm. Samples were surface scrapes. Loose sedimentary material coverina the reefs was collected by hand into the core tube, or the tube itself was scraped along the reef surface until at least half of it was filled with sediment. At each sample site, the following data were recorded: date and hour of sampling, depth, salinity, pH, temperature., Eh, and dissolved oxygen (Table l). Sample sites were plotted by resection from known shore points on the Punta del Morro to Puerto de Alvarado Mexican East Coast Map of 1955. 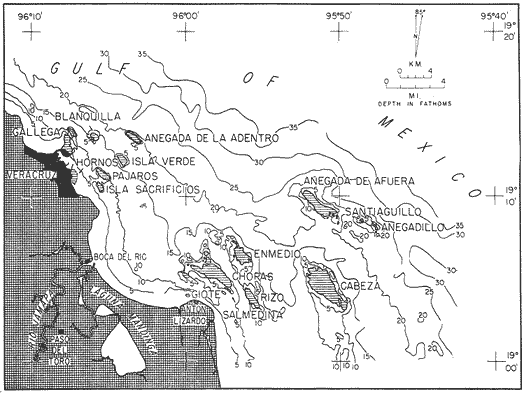 Fig. 1. Location of study area. 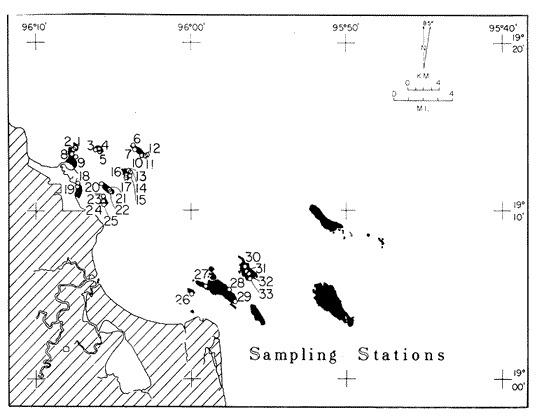 Fig. 2. Location of sampling stations on reefs of the Veracruz and Anton Lizardo complexes. Eh and pH were measured in the field with a Sargeant-WeIch model PBL pH meter. Dissolved oxygen content and water temperature were taken with a Yellow-Springs Instrument Corporation Model 51A Oxygen Meter. Salinity values were obtained with a Martek TDC (temperature, depth, conductivity) probe and depth measurements were taken using a calibrated sounding line and checked against the TDC meter. On recovery aboard ship, pH and Eh measurenment were made directly in the Phleger tube. Immediately thereafter, each core tube and Rose Bengal stain to mark living ostracods, and make possible the later distinction between living and dead specimens. The sediments were retained and transported in the Phleger core tubes until such time as laboratory processing began. 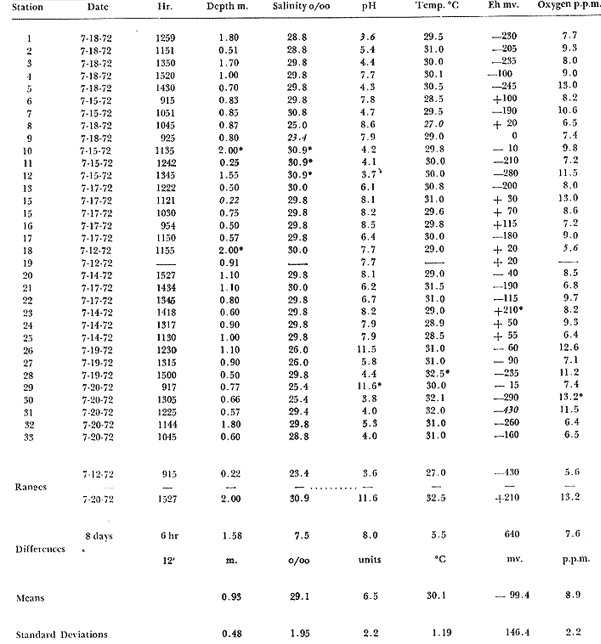 TABLE 1 SUMMARY OF ECOLOGIC DATA COLLECTED AT SAMPLING LOCALITIES ON THE VERACRUZ AND ANTON LIZARDO REEFS COMPLEXES MINIMA UNDERLINED, MAXIMA ASTERISKED. LABORATORY METHODSBecause of the sampling technique it was impossible to collect in situequal volume samples. However, in the laboratory, 100 cc. wet volume aliquots were taken from each Phleger tube. These 100 cc. sediment samples were washed over a 200 mesh screen (openings 74-88 microns) and oven-dried. Each of the 33 dry samples was then picked. Following Dryden (1931) and Kafescioglu (1975), Rick1es picked a minimum of 300 specimens from each sample (if indeed, 300 specimens could be found). Dryden and Kafescioglu demonstrated that beyond a count of 300 individuals, the number of additional species or mineral varieties found did not increase significantly. Therefore, 300 specimens are an accurate representation of relative species abundance at the sampling locality. Live-dead counts were not made, although it would be possible to do so at later date. Counts in this paper represent total populations (live + dead) at each sampling site. These are thanatocoenoses in the strictest sense, and represent what paleontologists recover from ancient sedimentary rocks. Fig. 3 is a map showing the number of ostracod individuals picked from each sample. Only 4 stations out of the 33 produced less than the statistically significant count of 300 specimens (12, 14, 20, and 27). 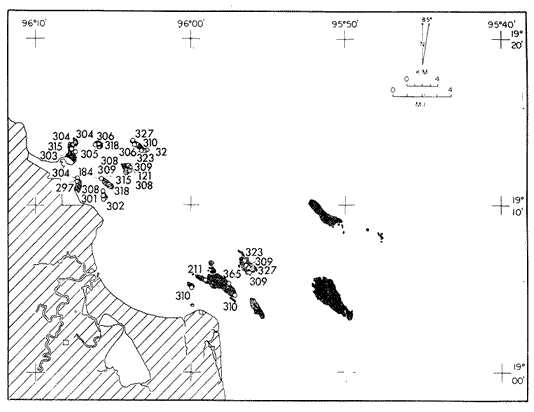 Fig. 3. Number of ostracod individuals picked from each sampling locality. OSTRACOD POPULATION STRUCTUREEighty five species of ostracods distributed among 9601 individuals occur in the 33 samples collected on the reefs of Veracruz and Anton Lizardo (see Rickles 1977 Appendix II). A complete faunal list including obbreviated diagnoses and distributions of illustrated species appears in Rickles 1977 (Appendix III). TOTAL SPECIES DOMINANCEIn the total assemblage of 85 species only 17 attain a total relative abundance higher than 1% (Table 2). We emphasize the wordtotal here. For example,Loxocorniculum tricornalum comprises 24.8% or 2384 out of the 9601 individuals counted from all 33 stations on the reefs. All 17 of the dominant species are common to both the Veracruz and Anton Lizardo complexes. Two of these 17 species, Loxocorniculum tricornatum andMorkhovenia inconspicua, are completely cosmopolitan in that they are present at all 33 sampling stations.Loxocorniculumcf. L. Postdorsoalata in almost ubiquitous, occurring at 31 of the 33 stations. The distribution of the dominant species (numerically most abundant) at each sample station is shown on Fig. 4. 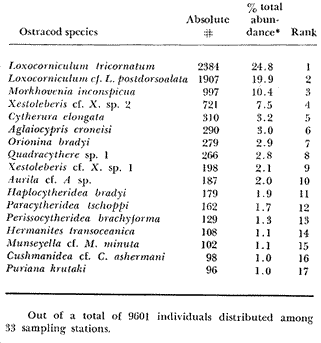 TABLE 2 RANK ORDER CHART. INCLUDES ONLY SPECIES WITH TOTAL RELATIVE ABUNDANCES GREATER THAN OR EQUAL TO 1% Loxocorniculumcf.L. posdorsoalataand Morkhovenia inconspicua are dominant species at stations on both reef complexes.Loxocorniculum tricornatum andHaplocytheridea bradyi are dominant only at stations of the Veracruz group. The dominance of H. bradyiin two stations of the Veracruz complex is puzzling in that it constitutes only 1.9% of the total population, while L. tricornatum, Loxocorniculum cf.L. posdorsoalataandMorkhovenia inconspicua each represent greater than 10% of the total population. This dominance ofH. bradyiis, in addition, curiously restricted to the nearshore reef of Hornos. Stations dominated byLoxocorniculum tricornatumall occur in the Veracruz reef group, even though this species occurs at 100% of the sampling stations on both reef complexes.L. tricornatum is dominant in 40% of the sampling localities at Veracruz. Loxocorniculumcf. L. posdorsoalata, the next species most nearly ubiquitous, was found at 92% of the Veracruz sampling stations and at all of the stations at Anton Lizardo. However, this species is dominant at 75% of the sample localities in the Anton Lizardo complex, while it is dominant in only 24% of the stations at Veracruz. RELATIVE ABUNDANCE DETERMINATIONSRelative abundances were calculated for each species occurring at the 33 sampling localities (Rickles 1977, Appendix II). Stations 12, 14, 20, and 27 were not included in these calculations as the number of specimens picked from the quantitative 100 cc. aliquot did not reach the statistica lly significant count of 300 individuals. 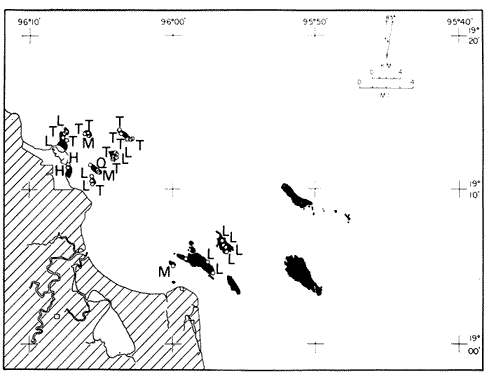 Fig. 4. Species dominance map. H = Haplocytheridea bradyi, L = Loxocorniculum cf. L . postdorsoalata, M = Morkhovenia in conspicua, Q = Quadracythere sp. 1, T = Loxocorniculum tricornatum. Stations 12, 14, 20, and 27 not plotted because of insufficient number of specimens. Since 85 species occur on the Veracruz-Anton Lizardo reefs, it is not possible here to make detailed comparisons of their relative abundances. Therefore, the three dominant and most nearly ubiquitous species have been chosen for consideration at this point. Loxocorniculum tricornatumis numerically the most abundánt species in the study area. Relative abundances of this species at individual sample sites are shown in Fig. 5. The relative abundance ofL. tricornatum at each station rarely drops below 10%. Five stations show less than 10 %, wtih 2 of those at 9.2 % and 9.7% and one at an anomalous low of 0.32%. Sixteen of the 29 stations included in this analysis show relative abundances for this species greater than 20%. The highest percentage of L. tricornatumrecorded was at Station 3, with 66.3%. Stations 18 and 19 are two of the sample sites with less than 10% relative abundances ofLoxocorniculum tricornatum (9.2% and. 5.7% respectively) . It is interesting to note that these two stations are those which showHaplocytheridea bradyi as the dominant species (Fig. 4). We examined the ecologic parameters recorded at the time of sample collection for a possible explanation of this phenomenon (Table l). Eh and pH readings for the two stations are exactly the same. Comparisons of salinity, temperature and dissolved oxygen content were not possible as no readings were recorded for Station 19. However, dissolved oxygen at Station 18 was the lowest measured at any of the sample sites. Further investigation is obviously necessary to determine what other factors may be contributing to this distribution phenomenon. 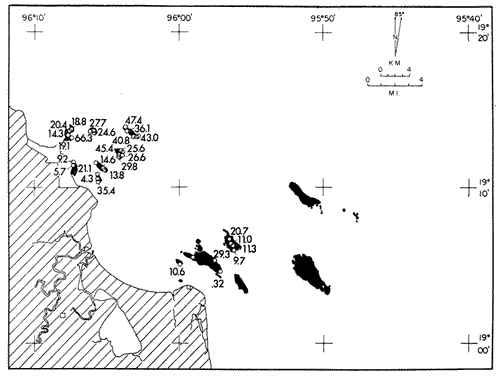 Fig. 5. Distribution of Loxocorniculum tricornatum showing, relative abundance percentages. Data from Rickles 1977, Appendix II. The very low relative abundance value of 0.32% for L. tricornatumwas recorded at Station 29, on Chopas of the Anton Lizardo complex. This surprisingly low value occurs at the station showing the highest pH reading (11.6) of any sample site. Correspondingly, Station 3, with the highest relative abundance of L. tricornatum., shows a relatively higher salinity reading and significantly lower pH and Eh values. Distributional control of this species would appear to be related to pH and Eh. conditions. Fig. 6 shows the relative abundances of Loxocorniculum of L. postdorsoalata,which ranks second in total abundance to L. tricornatum.Eight stations show less than 10% values, and 12 out of the 27 stations included in the analysis (L. cf. L. postdorsoalata was not present at 2 stations) show relative abundances greater than 20%. The highest value occurred at Station 24, where L. cf. L. postdorsoalata composed 73.1% of the population. Environmentally, Station 24 has a relatively high salinity reading, low temperature and median pH and Eh values. Station 9, however, shows the lowest relative abundance value for this species, 0.66%. Station 9 correspondingly records the lowest salinity value of any sampling locality and direaly opposes the high salinity value for Station 24. All other parameters compare satisfactority. This example indicates the striking control salinity has over faunal distribution in a localized area. 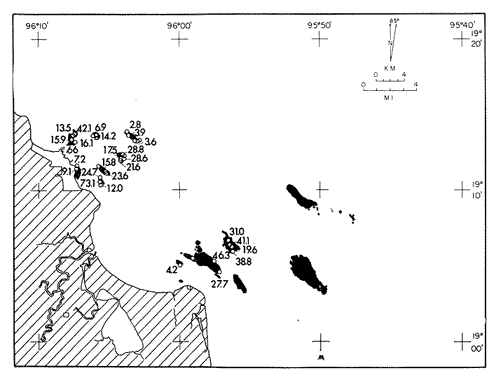 Fig. 6. Distribution of Loxocorniculum cf. L. postdorsoalata showing relative abundance percentages. Data from Rickles 1977. Appendix II. Morkhovenia inconspicua is third most abundant in the study arca and the only other totally cosmopolitan species. This species exhibits significantly lower relative abundances (Fig. 7) than those previously mentioned. Only 4 stations show values greater than 20%. The lowest value, 0.32% occurs at Station 8, and the highest. 61.3% at Station 26. A review of ecologic conditions at Station 26 reveals very high pH, dissolved oxygen content and temperature readings. Station 8, on the other hand, shows comparatively low pH and dissolved oxygen values. The temperature at Station 8 was the lowest recorded at any sampling station, and probably partially controls the distribtuion of this species. SPECIES DIVERSITYIn order to test the validity of the hypotheses and assumptions outlined under Sedimentation Patterns, we employed simple species diversity (S), equitability (E), and the Shanon-Weiner information function (H(S)). Details and methods of calculation are outlined in Gibson and Buzas (1973). See Rickles 1977, Appendix II for ostracod distribution data and absolute and relative abundance percentages of the species at each station. Table III is a summary of the results from each sampling station, and Figs. 7, 8, 9, and 10 are plots of S, E, and H(S) respectively. Table III also shows the maximum H(S) possible for each sample station. Mean S (number of species) values for Veracruz and Anton Lizardo differ (Table 3 and Fig. 7). Twenty six species occur on the average on the Veracruz group; whereas, only 22 occur at Anton Lizardo. The relatively higher species diversity at Veracruz may indicate greater numbers of ecological niches. Whether this is linked directly to water clarity is debateable, but it does demonstrate a diversity difference. This conclusion is supported more dramatically by the range of S in the two groups. The Anton Lizardo reef group shows a low of 11 species at Station 26 to a high of 33 at Station 31. Sampling stations at Veracruz range from 8 species (Station 12) to 51 species (Station 19), almost twice the range of Anton Lizardo. Equitability (E = eH(S)/S) is number of equilly distributed species necessary to obtain a particular H (S) value. E = 1.0 when all species are equally distributed (have the same relative abundance). Low E values indicate significant deviation from a sample of equally distributed species. Equitability values and their ranges for sampling stations of both complexes appear in Table 3 and Fig. 7. The range of E values at Veracruz is higher than that of Anton Lizardo (0.4352 at Veracruz versus 0.3036 at Anton Lizardo) ; however, the mean E of Veracruz (0.3818) sampling sites is only sligthly higher than the mean E at Anton Lizardo (0.3795). This slight difference may indicate a less equitable, possibly declining environmental situation at Anton Lizardo. Slightly more dominance exists in the Anton Lizardo group, and this may indicate less niche partitioning. Both inean equitability values are quite low, indicating significant deviation from equally distributed species for both complexes. However, the numerical differences are small, and the results inconclusive. 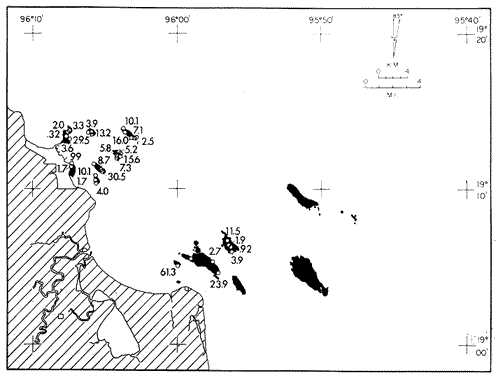 Fig. 7. Distribution of Morkhovenia inconspicua showing relative abundance percentages. Data from Rickles 1977, Appendix II. 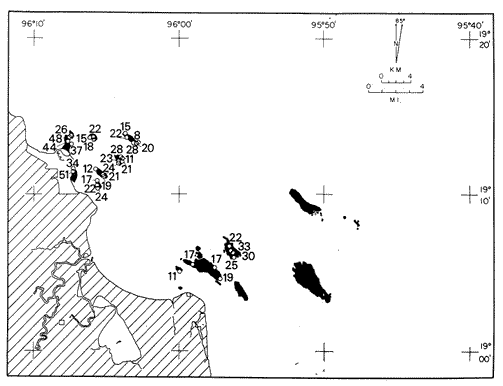 Fig, 8. Number of species, S, occurring at each sample site. Data from Rickles 1977, Appendix II. Appendix II.  Fórmula the number of species in a sample, pi is the relative abundance of each species at a particular location, and in is the natural log of that proportion) for stations of both complexes are shown in Fig 9 and Table 3. The range of H(S) values is high at Veracruz, with 1.17 at Station 3 to 3.37 at Station 19. The spread is somewhat less in the Anton Lizardo area, ranging from 1.39 at Station 26 to 2.87 at Station 32. The smaller spread between H(S) values at Anton Lizardo may indicate a more uniform environment there characterized by less diversity, reflecting the dead and dying coral growthand subsequent decline in the number of available niches. Means H(S) for Veracruz (2.1949) and the means H(S) for Anton Lizardo (2.0596) are only slightly different. In each case, the mean amount of information conveyed is high. The mathematical difference between mean H(S) and mean maximum expected H(S) indicates the degree to which the respective complexes as a whole approach equal distribution of all species. Comparisons of this difference show the Veracruz complex with a mean H(S) only slightly closer to its mean maximum expected H(S) than the difference at Anton Lizardo. In summary, H(S) values indicate differences are minimal between the two groups. 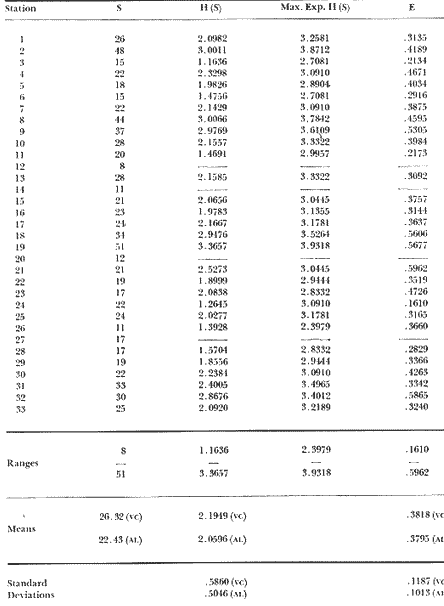 TABLE 3 SUMMARY OF SHANNON-WEINER INFORMATION FUNCTION (H(S)), EQUITABILITY (E), AND SIMPLE SPECIES DIVERSITY (S) AT SAMPLE SITES OF THE VERACRUZ (VC AND ANTON LIZARDO (AL) REEFS COMPLEXES. STATION 12,14, 20, AND 27 NOT PLOTTED BECAUSE OF INSUFFICIENT NUMBER OF SPECIMENS. BASIC DATA FROM RICKLES 1977, APPENDIX II. 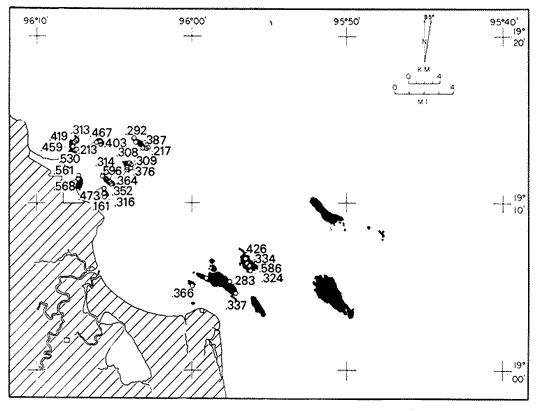 Fig. 9. Plots of equitability, E, values at each sample site. Basic: data for calculation from Rickles 1977, Appendix II. ConclusionesAnalysis of dominant ostracod species among the reefs indicates that distinct biofacies exist.Loxocorniculum tricornatumKrutak 1971 dominates the majority of the Veracruz group.Loxocorniculumcf.L. postdorsoalatagroup (Teeter) 1975 characterizes the Anton Lizardo stations. Most of the other 85 identified species overlap between the reef complexes. Two ostracod species are completely cosmopolitan, and occur at all 33 sample stations. In most cases they comprise significant numerical abundances. These species are L. tricornatumandMorkhovenia inconspicua (Brady) 1880. L. cf. L. postdorsoalatais almost ubiquitous and also quite abundant. It occurs at 31 of the 33 sampling localities. Rare species found on the, two roof complexes, are confined to the Veracruz group. 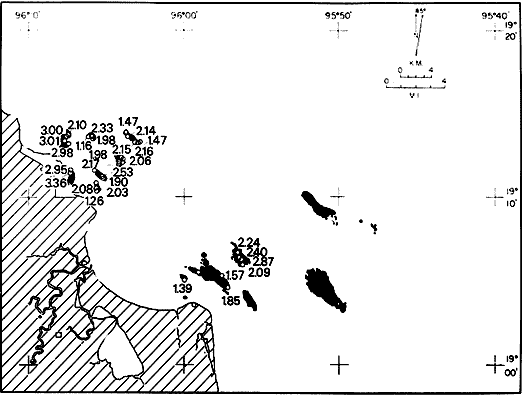 Fig. 10. Plots of Shannon-Weiner information function at each sample site. Basic data for calculation from Rickles 1977, Diversity analysis comparing only the number of ostracod species (S) indicates that the Veracruz reefs are more diverse than those of Anton Lizardo. Shannon-Weiner information function analysis indicates that diversity differences between the two groups are minimal. Comparisons of mean equitability values also reveal very little difference between the ostracod populations of the two reef groups. Terrigenous sedimentation is gradually overwhelming the reefs at Anton Lizardo. As a result, the number of available niches for ostracod speciation has been reduced. The Veracruz reefs are relatively uncontaminated by terrigenous debris. Turbidity is lower, water transparensy is higher. Zoozanthellae symbionts are able to thrive, and roof growth is relatively more rapid and healthy at Veracruz. Thus far more microniches are available for ostracod speciation. LITERATURABOLD, W. A. VAN DEN, 6th International Ostracod Symposium, Saalfelden:Distribution of Marine Podocopid Ostracoda in the Gulf. of Mexico and. the Caribbean:1977,175-186. BRADY, G. S., Report of the Ostracoda dredged by the H.M.S. Challenger during the years 1873-1876: Report on the Scientific Results of the Voyage of H.M.S. Challenger: Zoology:1880,1-184. DRYDEN, A. L., Accuracy in percentage representation of heavy mineral frequencies:National Academy Science, Proceedings,1931,223-238.17: EDWARDS, G. S., Distribution of Shelf Sediments, Offshore from Anton Lizardo and the port of Veracruz, Veracruz, Mexico: M.S. Thesis: Texas A & M University,1969,p. 174. EMERY, K. O., Coral Reefs of Veracruz, Mexico.Geofísica International,1963,11-17.3: FREELAND, G. L.,Carbonate Sediments in a Terrigenous Province: The Reefs of Veracruz, Mexico: Ph. D. Dissertation, Rice University:1971,1-268. Depositional Environments in Carbonate Rocks: FRIEDMAN, G. M. (ed.),Society Economic Paleontologists & Mineralogists, Spec. Publ.1969,1-421.14: Reefs and Related Carbonates - Ecology and Sedimentology: EROST, S. H. et al. (ed.),American Association Petroleum Geologists, Studies in Geology,1977,1-421.4: GIBSON, T. G. M. A. BUZAS, Species Diversity: Patterns in Modern and Miocene Foraminifera of the Eastern Margin of North America:G. S. A. Bull.,1973,217-238.84: KAFESCIOGLU, I. A., Quantitative Distribution of Foraminifera on the Continental Shelf and Uppermost Slope of Massachussets:Micropaleontology,1975,261-305.21:(3): KRUTAK, P. R., The Recent Ostracoda of Laguna Mandinga, Veracruz, Mexico:Micropaleontology1971,1-30.17(1): --------- Standing Crops of Modern Ostracods in Lagoonal and Reefal Environments, Veracruz, Mexico: West Indies Laboratory Fairleigh Dickinson University, St. Croix, U.S. Virgin Islands,Spec. Publ. 6,1974,11-14. ---------- Feefs and Related Carbonates - Ecology and Sedimentology: Change in Ostracod Standing Crop, Laguna Mandinga, Veracruz, Mexico: in Frost, S. H. et al. (ed.), Am. Assoc. Petrol. Geol. Stud. Geol.1977,209-218.4: Regional Aspects of Carbonate Deposition: LEBLANC, R. J. and J. C. BREEDING, (ed.), Society Economy Paleontologist & Mineralogists, Epec. Publ.1957,1-178.5: LIDZ, L. B. LIDZ, Foraminiferal Biofacies of the Veracruz Reefs:Bull. American Association Petroleum Geologists,1966.1514-1517.50(7): MADDOCKS, R. F., Biota of the West Flower Garden Bank, Flower Garden Ocean Research Center: :Biota of the West Flower Garden Bank: Ostracodes Gulf Publishing Co.,Houston, Texas.1974, MORELOCK, J. K. J. KOENIG, Terrigenous Sed¡mentation in a Shallow Water Coral Reef Environment:Jour. Sedimentary Petrology1967,1001-1005.37(4): PALACIOS-FEST, M. R., Contribución al Conocimiento de la Superfamilia Bairdiacea (Ostracoda, Latreille) en el Mar Caribe de MéxicoTesis profesional. Univ. Nal. Autón. México,1975,73 p. Bibliography of Organic Reefs, Bioherms and Biostromes: PUGH, W. E. (ed.), Seismograph Service Corporation, Tulsa, Oklahoma,1950,1-139. RICKLESS, S. E. Ecology, Taxonomy and Distribution of Holocene Reefal Ostracods, Veracruz, Mexico: M. S. Thesis, University of Nebraska,1977,1-131. REZAK, R., Reefs and Related Carbonates Ecology and Sedimentology: West Flower Garden Bank, Gulf of Mexico: in Frost, S. H. et al. (ed.), American Association Petroleum Geologists, Studies in Geology,1977,27-35.4: RIGBY, J. K. W. G. MCINTIRE, The Isla de Lobos and Associated Reefs, Veracruz, Mexico: Brigham Young Research Studies, Geology,1966,3-46.13: TEETER, J. W., Belize Chelf - Corbonate Sediments, Clastic Sediments, and Ecology: Distribution of Holocene Marine Ostracoda from Belize: in Wantland, K. F. and Pusey, W. C. III (ed.), American Association Petroleum Geologists, Studies in Geology,1975,400-499.2:
|