|
ANÁLISIS COMPARATIVO DE LOS PARÁMETROS DE CRECIMIENTO DE
TRES ESPECIES DE BIVALVOS DEL GÉNERO MACOMA EN EL PACÍFICO
NORORIENTAL
Trabajo presentado en el VI Congreso Nacional de
Oceanografía, Ensenada B. California, México, 10 al 13 de abril,
1978. Recibido el 22 de diciembre de 1978 y aceptado para su publicación
el 21 de febrero de 1980.
ARTURO MUHLIA-MELO
Comisión Interamericana del Atún
Tropical, c/o Scripps Institution of Oceanography, La Jolla, California 92037,
USA.
ROBERT OLSON
San Diego State University, 5300 Campanile Dr., San
Diego, California 92182, USA.
WILLIAM HAZEN
Universidad Autónoma de Baja California, Escuela de
Ciencias Marinas. Apartado Postal 453, Ensenada, Baja California,
México.
El presente estudio está basado
en el análisis de las muestras obtenidas en sustratos arenosos de dos
localidades: Estero de Punta Banda, B.C., México y Estero de Tijuana,
California, EUA.
Las lecturas de edad se obtuvieron mediante
la medición de los anillos anuales de crecimiento en las conchas. La
talla máxima promedio encontrada fue superior enMacoma
secta, intermedia enM. nasuta e inferior
enM. indentata.
Los parámetros de
crecimiento se calcularon mediante el uso de las ecuaciones de Bertalanffy y
Richards respectivamente. Los resultados obtenidos mediante el ajuste de estos
dos métodos para los parámetros de crecimiento Lα (talla
máxima a la asintota), b (constante de escala, Lα-L0/Lα), K
(tasa de crecimiento) y n (parámetro de la forma de la curva) se
discuten comparativamente mediante un análisis estadístico
detallado y se concluye que el crecimiento en las 3 especies es
cracterísticamente diferente.
This study is based on the analysis of
samples taken from sandy substrates of two locals: Punta Banda Estuary, B. C.
Mexico and Tijuana Estuary, California USA.
Age was
determined by means of annual rings on empty shells. The length of shells at
each rings was measured. The average maximum size found was greatest
inMacoma secta,intermediate inM.
nasuta,and lowest inM. indentata.Growth
parameters, Lα (asymptotic size), b (scaling constant,
Lα-L0/Lα), K (growth rate constant), and n (curve-shape parameter)
calculated by both functions are presented and compared statistically. It is
concluded that growth of the three species in characteristically
different.
Desde el trabajo inicial realizado por Petersen, los miembros del géneroMacomahan venido siendo utilizados como especies indicadoras de las comunidades de sedimentos marinos no consolidados. Estos trabajos fueron revisados por Thorson (1957), la distribución de los miembros de este género es bastante amplia en el hemisferio norte lo que ha contribuido a la realización de algunos estudios. Se han descrito comunidades características de Macoma nasutayMacoma secta en las costas del Pacífico de Norteamérica, no obstante se han descuidado los detalles de la ecología, aunque Addicott (1952) trabajando en Elkhorn Slough mostró las diferencias en los hábitos de M. secta y M. nasutalos cuales están probablemente relacionados con el tamaño de las partículas del substrato. M. nasuta se encuentra en substratos que van desde arena hasta arcilla y M. secta en cambio se encuentra en substratos arenosos de tamaño medio hasta arenas muy finas. Por otro lado Macoma indentata es menos común y ha sido poco estudiada. La relación taxonómica del género fue recopilada en el trabajo de Coan (1971) en el que muestra información sobre los rangos de las tallas, los substratos y las medidas del tamaño de las conchas, información que fue tomada de ejemplares de museo. M. nasuta está distribuida desde Kodiak Island en Alaska hasta Cabo San Lucas, B.C., viviendo comúnmente en áreas de mareas en las bahías y en aguas profundas sobre la costa, su tamaño máximo hasta de 110 mm. M. secta o comúnmente conocida como la almeja de la arena se distribuye desde la Isla de Vancouver, Canadá hasta la Bahía Magdalena en B.C. Sur (México), encontrándose principalmente en substratos arenosos de las zonas de mareas en las bahías y a corta distancia de la costa en zonas semiprotegidas. Su tamaño es hasta de 120 mm. M. indentata se encuentra distribuida desde Trinidad (California) hasta la Isla Cerralvo, B.C. Sur y Guaymas, Sonora (México) encontrándose en sedimentos de arcilla y arena en la zona de mareas de las bahías y más común en áreas poco profundas, su tamaño es hasta 63 mm. A pesar de la abundancia e importancia de los miembros de este género, muy poco se ha hecho en cuanto a estudios de crecimiento. Entre los estudios que más destacan aparece el de M. balthica realizado por Gilbert (1973) y un estudio de laboratorios realizado sobre M. nasuta por Hylleberg y Gallucia (1975). Desafortunadamente no es posible comparar los resultados de este último con los obtenidos en el presente trabajo debido a que fue solamente sobre pocos meses y con esa información no se puede predecir una tasa anual de crecimiento que combina un período de crecimiento rápido con uno de crecimiento lento. El investigador en general se encuentra con 2 tipos de problemas en relación a la teoría de crecimiento: (1) qué teoría de crecimiento escoger de acuerdo a los antecedentes con los que se cuenta y (2) saber si los datos se ajustan adecuadamente a dicha teoría. La curva de crecimiento de von Bertalanffy que es una función del tipo de disminución exponencial, se ha utilizado muy frecuentemente para describir el crecimiento animal, Bertalanffy (1960) presenta su modelo fisiológico y una revisión de la literatura sobre el crecimiento, en el que muestra que en muchos casos los datos se ajustan bien a su formulación. Sin embargo, la ecuación ha venido siendo criticada desde diferentes puntos de vista; entre ellos, por ejemplo, el que la curva no presenta un punto de inflexión que se ha encontrado en las curvas de crecimiento de muchos animales en los que el ciclo de vida se conoce detalladamente. El objeto de este trabajo es analizar comparativamente el modelo de von Bertalanffy (1948) y la función de Richards (1959) para obtener el mejor ajuste de los parámetros de crecimiento de 3 especies del géneroMacoma. ÁREAS DE ESTUDIOUna de las zonas de estudio fue el estuario de Tijuana localizado en Imperial Beach (California) al norte de la frontera de México y Estados Unidos en la latitud 32º 33' Norte. Los sitios son áreas arenosas de canales de bajas mareas, los rangos de mareas y la velocidad de las corrientes tienen un máximo de 1.4 m y 9.27 km/h, respectivamente (Peterson, 1975). El agua dulce que fluye hacia el estero es prácticamente insignificante conservándose la salinidad igual a la del área de la costa circundante (a unos 34%), con excepción de la época de las lluvias de invierno. La actividad de los pescadores de conchas y cavadores en busca de carnada es frecuente en este estuario; sin embargo, Peterson (1976) concluye que su influencia en la abundancia de invertebrados es poca. Macoma nasuta ocurre en los substratos arenosos y lodosos de este estuario, pero solamente se utilizaron en este estudio aquellas conchas encontradas en áreas arenosas. Otra zona de estudio fue en el Estero de Punta Banda localizado al extremo sur de la Bahía de Todos Santos a 15 Kms al sur de Ensenada (México), ubicado en la latitud 31º45' Norte. El estero se extiende al sur aproximadamente unos 10 Kms, es un marisma continuo hasta su extremo oriental. El rango del tamaño de los elementos que forman el fondo va desde arenas gruesas hasta arcillas finas, aunque la colección de las conchas para este estudio se obtuvo en el lecho del estuario de fondo grueso arenoso. MATERIAL Y MÉTODOSLa colección de conchas deMacoma nasutafue obtenida mediante excavaciones y colecta de la superficie del fondo en las áreas arenosas de los canales del estuario de Tijuana, en cambio, las deMacoma sectayMacoma indentata fueron solamente colectadas de la superficie. Las conchas fueron separadas del fondo con un cernidor de una malla de 5 mm de luz. Por lo consiguiente, las conchas de menor tamaño pasaron a través de la malla y no estuvieron representadas en la muestra. Se separaron y enumeraron las valvas izquierdas y derechas. utilizando para la determinación del crecimiento las valvas derechas debido a que ese lado es más convexo y facilita la lectura de los anillos. Las medidas se hicieron a la máxima longitud antero-posterior de cada anillo anual de crecimiento con un vernier o pie de rey. También se utilizó un microscopio de disección para poder distinguir entre los anillos anuales y aquellos causados por disturbio. Para las estimaciones de las tasas de crecimiento se usó la información de las medidas de longitud y la edad. MODELO DE BERTALANFFYEl modelo de von Bertalanffy es un modelo bien conocido en el medio científico y cuenta con una serie de versiones para su ajuste. En este caso se utilizó el programa elaborado por Tomlinson (1971), versión BGC4, para muestras de marca y recaptura. El ajuste lo hace mediante el método de mínimos cuadrados requiriendo los valores de longitud o talla inicial, longitud o talla final y tiempo, el proceso se hace ajustando la siguiente ecuación: Lt+Δt = LtRΔt + A (1-RΔt); 0 < R < 1 (1) donde Lt = longitud inicial; Lt+Δt = longitud final y Δt = período de tiempo; Lα= A y K = -logeR. Dando un cierto número de tercias de datos (Lt,Lt+Δt, Δt) y la ecuación (1), el programa minimiza la función Q= N/Σ [Lt+Δt - LtRΔt - A (1-RΔt)]² FUNCIÓN DE RICHARDSEntre los trabajos más recientes referentes a las técnicas de ajuste y la aplicación de la función de Richards destacan: Nelder (1961), Causton (1967), Fletcher (1975) y Ebert (en manuscrito). En la mayoría de los procedimientos propuestos es necesario conocer el tamaño y la edad de los organismos, datos que en general son obtenidos de programas de marcado y recaptura. Sin embargo es posible manejar la técnica para muestras cuya información contiene las medidas de longitud a diferentes períodos de tiempo, como es el caso de las medidas de los anillos de crecimiento en las conchas. La función sugerida por Richards (1959) ecuación (2) es un modelo flexible o modelo generalizado de la ecuación de von Bertalanffy; Lt -1/n =Lα-1/n- (1-be-Kt) (2) en la cual uno de los casos particulares es la curva de Bertalanffy cuando n = -1. De esta función se pueden derivar otros modelos en los que n toma otros valores como lo son la curva logística (n=1) y toda la familia de curvas que abarca el modelo. Una dificultad mayor consiste en dar un sentido biológico a la n en cuanto ya se ha decidido utilizar un parámetro extra (n) que podría ajustar mejor los datos pero que a su vez ese mejor ajuste puede compensarse por la mayor dificultad de interpretar al curva biológicamente. Sabemos que un parámetro adicional puede generar un mejor ajuste pero no está bien claro que el uso del parámetro (n) se justifique a menos que los datos nos den una buena razón para creer, por ejemplo, que existe un punto de inflexión. Actualmente la manera más práctica de utilizar este método es mediante el uso de un programa de computadora creado para este objeto, este solo requiere los datos de longitud o talla y el proceso encuentra un valor mínimo para n asociado con la suma de los cuadrados de los errores de la regresión de la gráfica de Walford (Walford, 1946) en la que los valores para el eje están dados por Lt+T-1/n contra los valores del eje X dados por Lt-1/n el programa finalmente obtiene los valores asociados para n, K, LΑ y b (Lα-L0/-Lα, donde L0es la longitud al tiempo cero) con intervalo de confianza de 95%. La ecuación 1 en forma lineal se puede escribir como sigue: Lt+T-1/n= Lα-1/ n(1-e-KT) +Lt-1/ne-KTen donde la pendiente (m) = e-KT y el intercepto (c) = Lα-1/n(l-eKT) por consiguiente K = -n(m)/T y Lα = (c/(l-m))-n y T el intervalo de tiempo. Así, la suma del cuadrado de los errores a la recta está representado por: SRC=n/Σ/i=1 [Yi- (mXi+ c)-n]² (2) en donde Y =Lt+T y X=Lt-1/n. De tal modo que el programa hace la búsqueda del valor de n por medio del sistema prueba y error, hasta que encuentra un valor menor a un límite propuesto. RESULTADOS Y DISCUSIÓNEl número de las muestras deMacoma nasutay M. Sectay M. indentata fue de 169, 20 y 17 conchas. respectivamente. En las tablas 1, 2 y 3 se muestra el rango, la media y la desviación normal de las tres especies deMacoma que formaron este estudio. Para poder distinguir cuál de las dos funciones (Bertalanffy y Richards) antes mencionadas es la más apropiado para representar el crecimiento en poblaciones cuyo ciclo de vida es poco conocido, es necesario hacer un análisis de la forma lineal de los datos en la gráfica de Walford, que es un método utilizado para la estimaciones de los parámetros de crecimiento del modelo de Bertalanffy. En el que Lt, la longitud al tiempo t, se gráfica en contra de la longitud a un intervalo de tiempo más tarde Lt+T, en la figura 1 se muestran las gráficas de Walford de las tres especies de Macoma. Si en realidad. en las curvas de crecimiento de estas almejas existiera un punto de inflexión este haría que en la gráfica de Walford los puntos no cayeran a lo largo de la línea recta y tomaran en cambio una forma más curvilínea. Esto es difícil de apreciar visualmente debido a la variabilidad de los puntos alrededor de la línea, no pudiendo fácilmente ser tomada una decisión respecto a su linearidad. 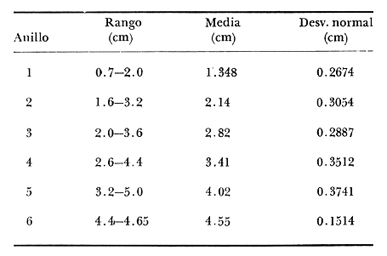 TABLA 1 DATOS SOBRE EL RANGO, LA MEDIA Y LA DESVIACIÓN NORMAL DE LOS ANILLOS ANUALES DE CRECIMIENTO DE MACOMA NASUTA 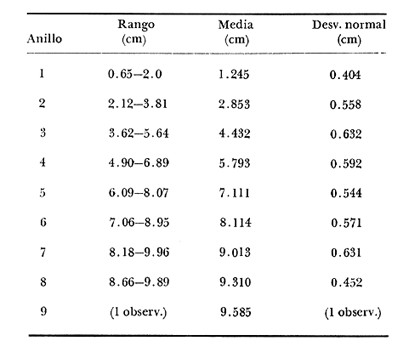 TABLA 2 DATOS SOBRE EL RANGO, LA MEDIA Y LA DESVIACIÓN NORMAL DE LOS ANILLOS ANUALES DE CRECIMIENTO DE MACOMA SECTA 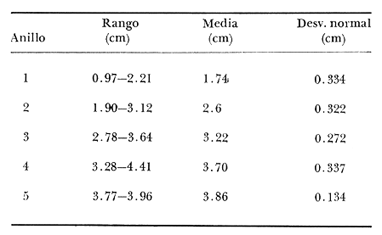 TABLA 3 DATOS SOBRE EL RANGO, LA MEDIA Y LA DESVIACIÓN NORMAL DE LOS ANILLOS ANUALES DE CRECIMIENTO DE MACOMA INDENTATA Las curvas de crecimiento de otros bivalvos parecen tener claramente un punto de inflexión (Hallam, 1967), de aquí que se haya considerado la posibilidad de que la función de Richards pudiese ajustar la forma de crecimiento de estas especies ya que tiene una parámetro extra que la permite mayor flexibilidad. Debido a las consideraciones anteriores, se hicieron las pruebas estadísticas de linearidad y homogeneidad de la varianza de los puntos alrededor de la línea (Dixon y Massey, 1957), partiendo de la premisa de que entre más lineal sea la forma, mejor será el ajuste con el modelo de Bertalanffy y por el contrario entre más curvilínea sea la forma, mejor será el ajuste hacia la función de Richards. Los resultados obtenidos de estas pruebas estadísticas se muestran en la tabla 4. Como se podrá notar para las tres especies, los datos caen dentro de los límites de linearidad y respecto a la homogeneidad de la varianza puede decirse que los datos muestran ser homogéneos alrededor de la línea con excepción deMacoma nasuta que tiene un valor ligeramente superior al límite. Sin embargo, las diferencias encontradas en cuanto a la precisión de los métodos parecen no ser significativas ya que los valores de la suma residual de cuadrados de los errores es muy pequeña (ver Tabla 5). Es posible argumentar que en general, lo más apropiado para el ajuste es el modelo más simple que describa los datos, de aquí que dados los resultados de linearidad y homogeneidad de la varianza el modelo de Bertalanffy es el modelo que describe en forma más apropiada el crecimiento de las tres especies deMacoma. 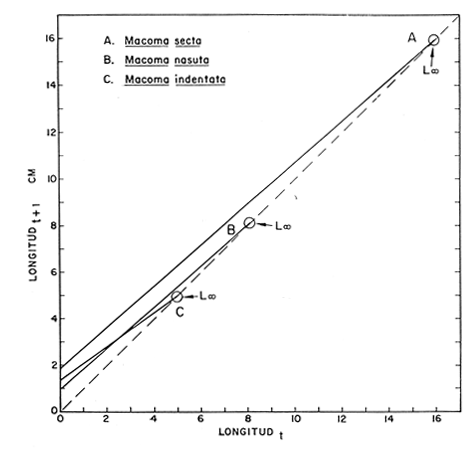 Fig. 1. Gráfica de Ford-Walford de la estimación de los parámetros de crecimiento de las 3 especies de Macoma en donde (Y) es la longitud a la edad Lt+1 y (X) es la longitud a una unidad de tiempo anterior, Lt. En la tabla 5 se presenta una lista de los valores calculados de los parámetros de crecimiento (Lα, K, b y n) para la comparación de los dos modelos, así como también los valores de la suma residual de cuadrados de los errores (SRC) con relación a la recta en la que los Modelos hacen su mejor ajuste. En la figura 2, se presentan las curvas de crecimiento de Macoma secta, generadas por los dos métodos como un ejemplo que nos muestra objetivamente los ajustes. Nótese que los dos métodos tienen un ajuste idéntico en la primera fase de la curva hasta donde hay datos, de allí en adelante siguen una trayectoria diferente. Esta gran diferencia en la parte superior de las curvas puede explicarse como consecuencia de la falta de datos en esa región de la gráfica, además de que el parámetro LΑ es la talla máxima teórica de un organismo, que en realidad no tiene mucho significado biológico.
* La significancia de los valores
obtenidos F, ys1, y2 se puede
obtener mediante el uso de las tablas dela distribución de
Fisher.
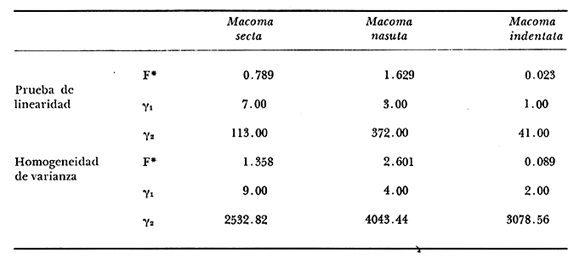 TABLA 4 PRUEBAS ESTADÍSTICAS DE LINEARIDAD Y HOMOGENEIDAD DE LA VARIANZA PARA LAS TRES ESPECIES DE MACOMA 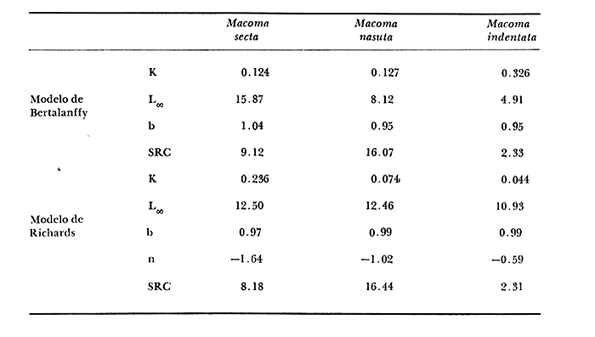 TABLA 5 PARÁMETROS DE CRECIMIENTO CALCULADOS POR LOS DOS -MÉTODOS Y PARA LAS TRES ESPECIES DE MACOMA. TAMBIÉN SE INCLUYE LA SUMA RESIDUAL DE CUADRADOS DE LOS ERRORES A LA RECTA (SRC) 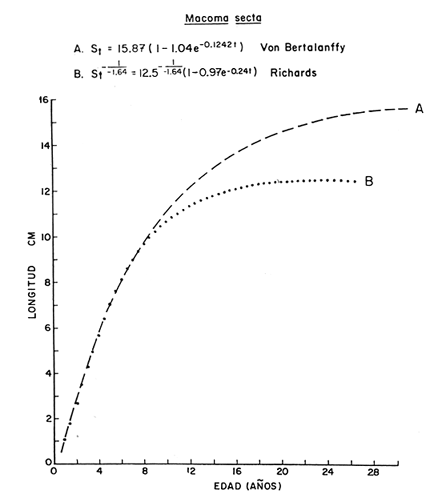 Fig. 2. Gráfica en la que se muestra comparativamente. A la curva del modelo de von Bertalanffy y B la curva de la función de Richards, ambas para Mocoma secta. También es propio considerar que nuestra información tiene escasez de datos de los individuos pequeños menores de un año de edad. Smith (1974) menciona que es difícil identificar el primer anillo de crecimiento en las conchas, resultando una probabilidad de error en la parte baja de la curva. De aquí que exista una pequeña posibilidad de que haya un punto de inflexión en esa parte baja de la curva. Los puntos de la talla media de cada anillo anual están sobrepuestos en las Curvas de crecimiento de Bertalanffy en las tres especies deMacoma (Fig. 3), de Modo que en esa figura se ilustra como se ajustan las curvas a los datos en los Tres casos. Parece ser que las curvas describen bastante bien a los datos, pero debe notarse que la talla máxima no está bien determinada debido a que los datos solo abarcan la zona baja de la curva, después de lo cual la curva solo sigue la proyección del ajuste. La longitud máxima teórica calculada para Macoma indentata es la más confiable de las tres especies ya que está más cerca de la talla máxima encontrada. Así que Macoma indentata parece ser característicamente distinta a las otras dos especies por su ciclo de vida más corto, una tasa de crecimiento relativamente alta y un valor asintótico bajo. En el caso de Macoma secta la función de Richards parece hacer un mejor ajuste de acuerdo al argumento de que la longitud máxima teórica LΑestá más cercana a la máxima longitud encontrada en los datos (ver Fig. 3). Se hizo finalmente, para las tres especies, un análisis de covarianza con los datos en su forma lineal, con el objeto de determinar si son estadísticamente distintos en términos de su pendiente, media y media lineal, y si las líneas pueden ser descritas por una regresión. Los resultados de este análisis (Tabla 6), muestran que las líneas son diferentes entre sí, bajo el criterio de la prueba. 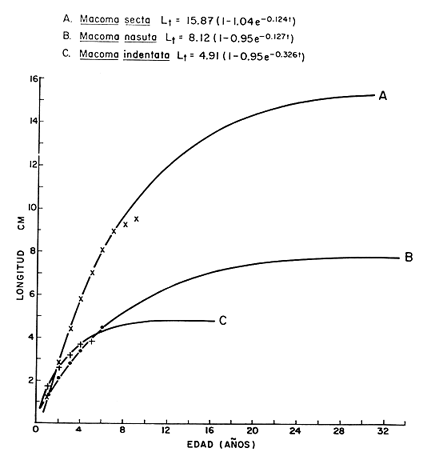 Fig. 3. Curvas de crecimiento de tres especies del género Macoma; M. secta y M. indentata del estero de Punta Banda, México y M. nasuta del estero de Tijuana, California, EUA. Los símbolos (X, +, .) indican la talla media correspondiente a las medidas de los anillos de crecimiento en las conchas. 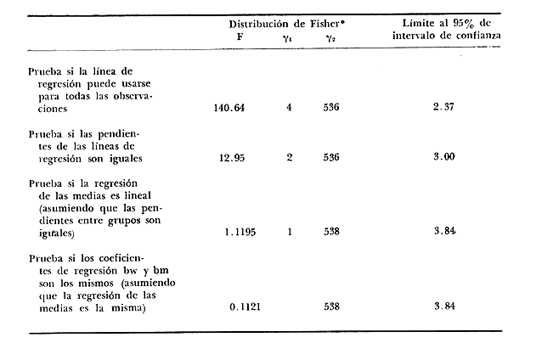 TABLA 6 ANÁLISIS DE COVARIANZA COMPARANDO LAS LÍNEAS DE REGRESIÓN DE LAS TRES ESPECIES DE MACOMA
*El
resultado de la prueba puede obtenerse localizando los datos en la tabla de
Fisher.
Por lo tanto con estos resultados se puede decir que el crecimiento en estas 3 especies es característicamente diferente en términos de su pendiente media lineal y líneas de regresión. Finalmente, en consecuencia de los resultados del análisis estadístico es posible suponer que las características ecológicas del medio en que se desarrollan también son diferentes. AgradecimientosSe agradece a Thomas Ebert por su ayuda a entender los problemas de crecimiento en poblaciones naturales, y a Patrick Tomlinson por su ayuda en el análisis de los datos. LITERATURAADDICOTT, W. O.,Ecological and natural history studies of the pelecypod genus Macoma in Elghorn Slough, California. M. A. thesis, Dept. Bio. Sci. Stanford University,1952.89 pp. BERTALANFFY, L. VON, A quantitative theory of organic growth. Hum. Biol.,1948.181-213.10(2): ------------- Fundamental Aspects of Normal and Malignant Growth. Principles and theory of growth, Wiktor W. Nowinski (Ed.), Amsterdam: Elsevier.1960.p: 137-259. CAUSTON, D. R., Some mathematical properties of growth curves and applications to plant growth analysis. Ph. D. Dissertation University of Wales.1967. COAN, E. V., The northwest America Tellinidae.Veliger 14 (supplement):1971.1-63. DIXON, W. J. T. J. MASSEY, Introduction to Statistical Analysis.McGraw-Hill Co.New York,1957.488 p. EBERT, T. A. (manuscrito). Estimating growth parameters in Richards function from mark-recapture data. FLETCHER, R. T., A general solution for the complete Richards function.Math. Biosci.1975.349-360.27: GILBERT, M. A., Growth rate, longevity, and maximum. size of Macoma balthica (L.) Biol. Bull.,1973.119-126.145: HALLAM, A., Paleontology., The interpretation of size-frequency distributions in mollusc death assemblages.1967.25-42.10: HYLLBERG, J. V. F. GALLUCIA, Selectivity in feeding by deposit-feeding bivalve, Macoma nasuta. Mar. Biol.,1975.167-178.32: NELDER, J. A., The fitting of a generalization of the logistic curve.Biometrics.,1961.89-110.17: PETERSON, C. H., Stability of species and communitv for the benthos of two lagoons.Ecology,1975.598-965.56: -------------Lethaia, Relative abundance of living and dead lolluscs in two Californian lagoons.1976.137-148.9: RICHARDS, F. J., A flexible growth functíon for empirical use.J. Exp. Botany,1959.290-300.10: SMITH, S. H., The growth and mortality of the little neck clam. Protothaca staminea in Tijuana Slough. M.Sci. thesis, San Diego St. Univ.1974.116 p. THORSON, G., Bottom communities. In: J. W. Hegpeth (ed.). Geol. Soc. Amer. Mem.1957.67. TOMLINSON, P. K., Abramson, N. J. Computer programs for fish assessment.BGC4-von Bertalanffy growth curve fitting.FAO Fish. Tech. Paper.1971.4 p. WALFORD, L. A., A new graphic method of describing the growth of animals.Biol. Bull.,1946.141-147.90:
|