|
BIOLOGICAL STUDIES
ON THE CHILEAN SEA-URCHIN LOXECHINUS ALBUS (MOLINA) (ECHINODERMATA:
ECHINOIDEA). FOOD ANALYSIS AND SEASONAL FEEDING RATE
Trabajo recibido el
28 de octubre de 1978 y aceptado para su publicación el 27 de noviembre
de 1978.
L. F. BÜCKLE
Royal Swedish Academy of Science, Kristineberg Marine Biological
Station, Fiskebäckskil, Sweden.
K. ALVEAL
Department of Marine
Biology, University of Concepcion, Chile.
E. TARIFEÑO
Institute of
Experimental and Evolutionary Biology, University of California, Los Angeles,
U.S.A.
CH. GUISADO,
L. CÓRDOBA
Department of Anatomy, Duke University Medical Center,
Durham, North Carolina, U.S.A.
C. SERRANO,
J. VALENZUELA
Se ha estudiado durante un año
el alimento en particular y la taza de alimentación del erizo comestible
Loxechinus albus que habita las costas de Chile. En el
análisis del contenido estomacal se identificaron 22 algas como
también animales pertenecientes a diferentes grupos zoológicos.
Se discute la presencia de arena en el estómago de los erizos.
Según la información obtenida no fue posible concluir si esta
especie tiene preferencia por determinado alimento. La taza de
alimentación anual fue tentativamente dividida en cuatro periodos a
saber: A = periodo de Declinación hasta Inanición de
Otoño-Invierno, B = periodo de Recuperación de
Invierno-Primavera, C = periodo de Declinación hasta Inanición de
Primavera-Verano, D = periodo de Recuperación de Verano-Otoño. La
secuencia de A + B + C + D definida como un ciclo, fue comparada con la taza de
alimentación registrada en erizos cautivos en las provincias de
Valparaíso y Chiloé que fueron alimentados con un número
determinado de especies de algas. Los ciclos de alimentación observados
en estos dos experimentos fueron similares al encontrado en una
población natural. No se encontró una relación directa
entre el desarrollo de las gonadas y la taza de alimentación durante un
ciclo anual. Probablemente los procesos son independientes y sus variaciones,
el resultado de una interacción compleja de factores fisiológicos
y ambientales.
The stomach contents of Lexechinus
albus a coastal species in the province of Valparaiso, has been
analyzed during a period of fourteen months in order to determine its
composition and quantative variation. Twentytwo species of algae were
identified together with sand and food animal origin. In search of a pattern of
seasonal variation, the feeding rate in terms of dry weight was compared with
the gonad index. No direct correlation was found. It was also compared with the
feeding rate of two previous experiments performed with sea-urchins maintained
in captivity and fed with few algae; one in the province of Valparaiso and the
other 1200 kms to the south, in the province of chiloé. On the basis of
these results the fluctuations of the feeding rate of one annual cycle have
been divided into four distinct periods apparently influenced by the effects of
a complex system of physiological and environmental factors.
The present investigation is the continuation of a series initiated in 1968 in orden to study the biology of the sea-urchin Loxechinus albus, which inhabits the coast of Chile. This species is exploited commercially by fishermen for its edible gonads. This very old fishery is important among the invertebrate fisheries of the country and the sustained overexploitation of the populations has brought about the disappearance of the commercially sized animals in several provinces, especially in the north and central part of the country. Knowing the gravity of the problem, our laboratory investigated the feeding of sea-urchins reared in captivity in the province of Valparaiso in 1970-71 and in the province of Chiloé in 1972-73. The animals were fed to satiation with a few selected species of algae. Although the mortality rate was very low, growth was slower Bückle, et al., 1977a) . Similar results have also been reported by Elbert (1967) for Strongylocentrotus purpuratus. These results have necessitated a survey of the feeding rate of a natural population in order to identify the variety and quantities of the plant material consumed in the course of one year. This information could be used to develop an adequate diet for animals maintained in captivity. These results could contribute to an improvement of sea-urchin Aquaculture techniques, and also provide some knowledge of the impact of this species on the natural plant communities. Information regarding the feeding and food preferences for this species is somewhat limited. Some observations have been made by Mortensen (1943), Alveal (1970), Dayton et al. (1973), Gutiérrez and Otsu (1975). None of them however, followed the yearly variations of the feeding rates under natural conditions suggested by Lawrence (1975). MATERIAL AND METHODSThe sub-samples analyzed in this study were taken randomly from more general samples which were utilized for other biological observations. The detailed description of the sampling method of the sea-urchins has been reported by Bückle et al. (1977c) . After opening the seaurchins, the gut contents were carefully removed for examination having been preserved in pure formalin (40%). For the analyses, the samples were spread in petri dishes under a binocular mieroscope to determine the systematic and quantative composition. The primary analysis was considered as one person's work to avoid bias, especially in the particular quantative estimation of the items, when the occurrence of each item was divided in 10%, 10-50% and 50% of the sample. In the identification of the algae, Lessonia spp. and Durvillaea sp. were not separated due to the difficulty in determining the genera correctly. The presence of food of animal origin in the stomach contents was recorded but not included in the calculations. The samples were dried at 100ºC and weighed after twenty-four hours to the nearest 0.01 g using a Sartorius balance. The feeding rate was calculated as dry weight (g) food ingested/ month/animal, regardless of the differing densities of the algal species. The animals cultivated in captivity in the province of Valparaiso (average diameter = 2.28 cm) were fed with Lessonia nigrescens, Ulva lactuca, Porphyra columbina, and Tridaea laminarioides and those in the province of Chiloé (average diameter = 6.92 cm) with Macrocystis pyrifera. The feeding rate in these experiments was calculated as wet weight (g) food ingested/month /animal. The gonad index was calculated as the ratio of the wet gonad weight to the diameter3 X 1000. RESULTSThe gut contents of Loxechinus albus have the appearence of a cluster of little balls of different size and colour, due to the predominant items forming them. In the analysis of these balls, 22 plants were identified (Table 1) together with sand and various items of animal origin but two of these species are grouped together in the analyse due to difficulties of identification. Of these plants 22.7% were Chlorophyta (5 species), 27.2% Phaeophyta (7 species) and 45.5% Rhodophyta (10 species). In addition to these algae the stomachs contained remains of hydrozoans and bryozoans in almost 80% of the samples. Molluscs like Choromytilus sp. and Perumytilus sp. and eggs of Concholepas concholepas as well as different copepods were also present. Sand covered either in a bacterial membrane or in Plocamium sp and remains of sea-urchin test and spines was also present. Even if 58% of the animals contained sand in their stomachs, it is difficult to consider this item as a basic element in their diet. There are many alternative explanations for the presence of sand, also observed in other species like Strongylocentrotus droebachiensis (Scott 1902) and Diadema antillarum (Lewis, 1964). One explanation could be the casual ingestion of the sand in places where plants are not available. Alveal et al. (1973) describes the protected and exposed front in the area of Montemar in Valparaiso, a sand layer mixed with mud, fluctuating between 10 and 16 meters' depth. The sea-urchins migrating over this zone could ingest sand on the way. Another possibility could be a relation of sand and mud needed for the digestion of the plants in the stomach, using them as a source of specific bacteria. Interstitial fauna could provide a further food source. The high percentage of Bryozoa, in the samples are explicable in terms of their use of algae, particularly Plocamium sp., as a substrate.  TABLE 1 ITEMS IDENTIFIED IN THE DIGESTIVE TRACT CONTENTS AND THE RESPECTIVE PERCENTAGE OF THTOTAL SAMPLING (N = 527 SEA URCHINS). PH = PHAEOPHYTA, RH.= RHODOPHYTA, CHL = CHLOROPHYTA The contribution of the different food types to the overall diet was assessed on the basis of the percentage of animals containing each species. The results are presented in Table 2. The subjective estimation of the observer who divided the items of the samples in three levels (10% , 10-50% and 50%) has been tested under the hypothesis that there are no significant differences among them. The frequencies observed in each level were contrasted with a variance test, and the high probability found for the hypothesis (F2;34 = 0.3998: P = 0.7073 1-P = 0.2927) suggests that the operator was impartial in the quantative criteria, and that there were no significant differences among the three subjective levels. The combination of Lessonia spp. and Durvillaea sp. appear to represent an important component of the sea-urchin's diet. Lessonia nigrescens and Durvillaea antartica occur together in the upper limit of the marine zone. These are replaced by Lessonia flavicans which is common to a depth of about thirteen meters, the exact depth depending on the degree of exposure (Alveal et al., 1973). 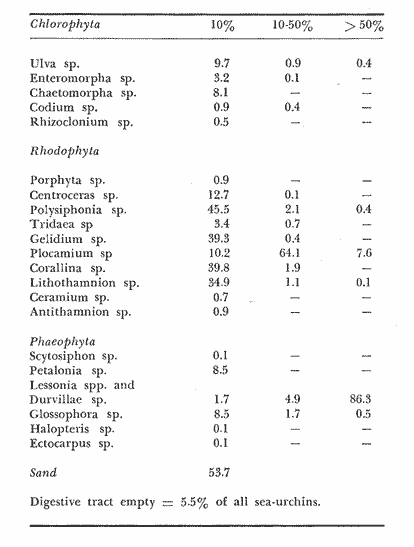 TABLE 2 PERCENTAGE OF OCURRENCE OF EACH ITEM IN THE THREE LEVELS, USED TO SEPARATE THE STOMACH CONTENTS OF THE SEA URCHINS. The dry weight of the samples of both sexes has been listed separately in Table 3 and the monthly variation of the quantities in Fig. 1. It is evident that almost in every season the feeding rate of the animals diminishes, especially so in summer time. The significant correlation (r = 0.531) for the fluctuations of the dry weight is similar for both males and females. The monthly mean percentage of the dry weight (Table 3) of the items ingested by this species is compared with the gonad index in Fig. 2. The fluctuations of the variables are uneven, which reveals that the processes are different. Only in July is the fall of the gonad index parallel. with the fall in dry weight. This does not mean that one is the consequence of the other, rather it looks more probable that the variables are influenced by reductions in temperature (Fig. 2a) . During the main breeding season, which in this species extends from late autum till spring, a marked decrease in nitrogen level occurs (especially in the females) together with the water content of the gonads, the other extract apparently replacing these compounds (Bückle et al., 1977). The energy lost during this period may be compensated for by increased feeding activity. In summer time the fonad index increases but foods intake diminishes to the lowest level found in this investigation. This may be related to an increase in water temperature at this perior (Fig. 2a) . The maximum temperature of 14.8ºC was registered in January 1969, and the lowest feeding activity, one month before. A relatively low feeding rate was observed when the gonads were fully developed. 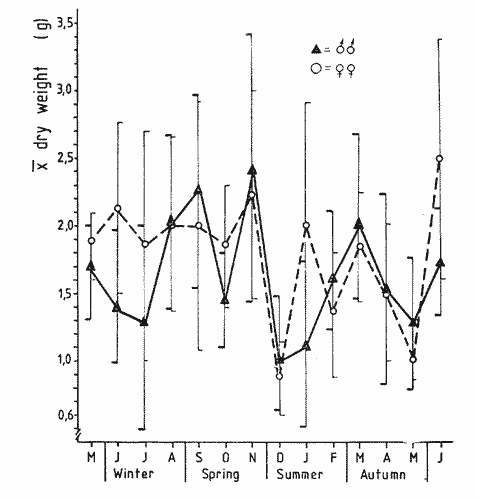 Fig. 1. Graph showing the seasonal variations in feeding rate in terms of dry weight in males (A-) and females (o --- ). The vertical lines represent the 95% It is possible that the variations of the feeding rate during one annual cycle maintain a certain pattern which fluctuates seasonally through the effect of a large spectrum of factors. The presence of animals with empty stomachs in the population specially in winter and summer time (Fig. 3d), may mark a biological stress situation (also noticed in summer time in the captive sea-urchins in Valparaiso, marked with a star in Fig. 3b) . These hypothetical stress periods were used to define major phases in the annual feeding cycle. The following feeding periods are suggested: 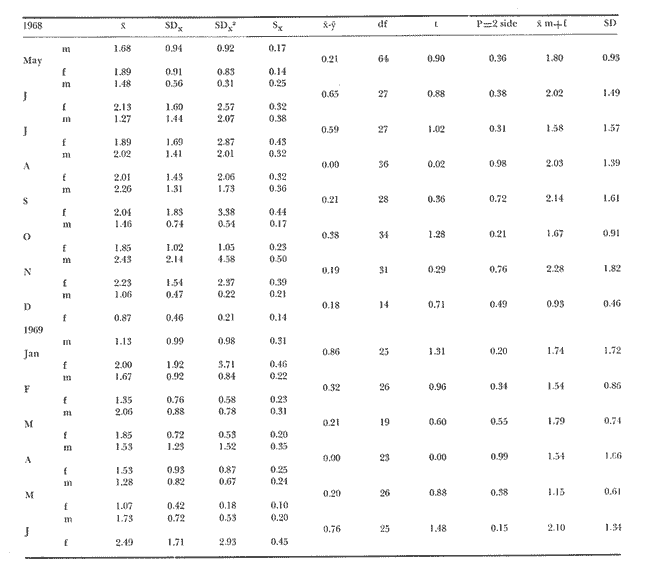 TABLE 3 DRY WEIGHT OF THE SAMPLES OF THE MALES (M) AND FEMALES (F) AND THE OOLED MEAN DRY WEIGHT IN A NATURAL POPULATION OF SEA-URCHINS IN THE PROVINCE OF VALPARAISO A: Autumn-winter decline to starvation; in Fig.3c with heavy dots. In this phase the decrease in temperature, could be one of the main factors affecting the feeding rate. The precise moment of the lowest temperature in winter varies from one year to another. Thus the decrease observed in the digestive tract contents is not necessarily an exact replica of the temperature variations. For instance in 1968 the water temperature reached 11.8ºC in August, and the lowest quantity of plants eaten one month before. Furthermore the high number (33% in July) of starved sea urchins in this period would support the hypothesis (Fig. 3d) . Empty animals were observed from May until September, but the lack of quantative information on the algal populations precludes any direct comparison with the sea-urchin starvation. Another explanation in the presence of starved animals may lie in the heavy storms typical for this season and the need for the animals to seek protection in places where foods is not easily available. The loss of gonad weight in this period can possibly be eplained by the loss of energy supporting life and this apparently caused primarily by the temperature change. 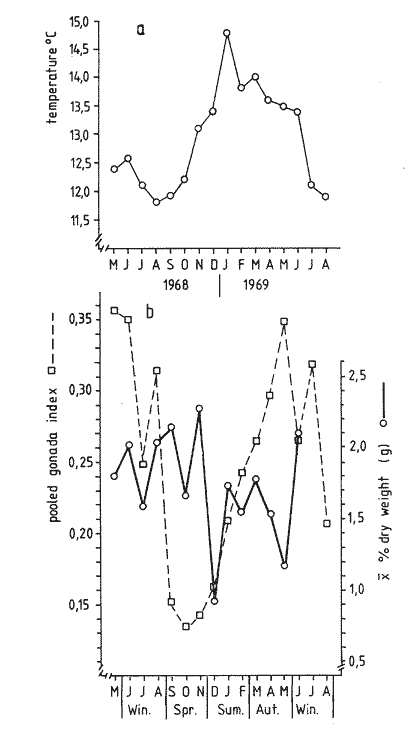 Fig. 2. a, Surface temperature of the water in the studied locality at 08.00 hours, b, Comparison between the pooled mean gonad index as a function of time and the feeding rate a natural population in the province of Valparaiso. B: Period of winter-spring recovery; in Fig. 3c with heavy lines. At this time the general tendency of the sea-urchins is to eat more in spite of the temperature of the sea water reaching the lowest value observed in 1968. This period corresponds to the main breeding season of this species and extends from late winter to October, when testes and ovaries are in full activity and the gamete extrusion is characterized by the decrease in some chemical compounds in the gonads (Bückle et al., 1977). The trend of the feeding rate is temporally interrupted just at the end of the effective spawning. Another factor which might influence the recovery of the feeding activity could be an increase in the algal population suggested by the reduction in the number of starved animals. It seems that the recovery of the feeding activity in this period can be explained more in terms of physiological necessity (or response) than on direct environmental influence. 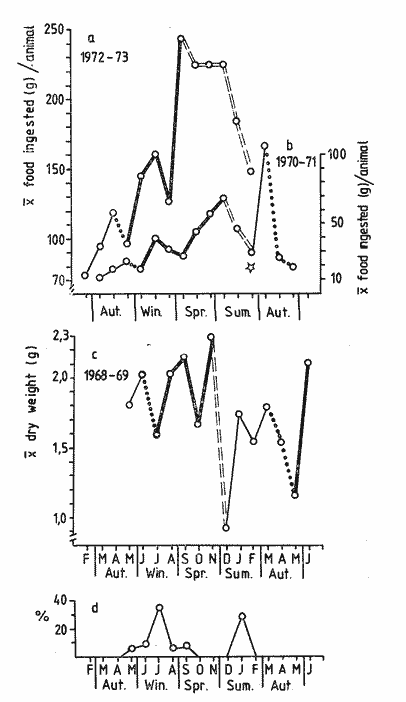 Fig. 3. Seasonal variation of the feeding rate: a, in one experiment with nimals reared in captivity in the province of Chiloe. b, with captive sea-urchinsreared in the province of Valparaiso. c, in a natural population in theprovince of Valparaiso. d, animals with empty stomachs identified in the natural population of sea-urchins. C: Spring-summer declinate to starvation. (Fig. 3c marked with double lines.) This phase is characterized by an increase in water temperature and the creatures could be affected in such a way that the consequences are very similar to the first period described. Here again around 30%, of the males have empty stomachs. No starved females were recorded though this does not preclude their occurrence. The period preceeding the highest summer temperature is marked by a reduction in the feeding rate. The gonad index instead increases steadily, possibly as a result of the energy accumulated in the previous period. This also suggests that food quality might be a critical factor in the uptake of energy for fast genadal growth. D. Summer-autumn recover period; in Fig. 3c marked with a fine line. The increase in feeding activity may be a response to the energy loss in the previous period, when the demands of reproductive development and arduous conditions where at their greatest. This period is comparable to the second hypothetical breeding period in the reproductive cycle of L. albus which is characterized by part of the population being still in spawning condition at this time. Maturation of the gonads, however, is delayed by the ensuiing winter. The increase in food intake may provide energy in this developmental phase until the animals are influenced by the food, shortages of the first period described. Thus closing an apparent natural chain of relations. The characteristics typical. of each period may be caused by the interaction of variety of factors. The periods defined above were compared with other experiments in the intention of finding a pattern in the feeding rate of this species. Dry weight variations of the samples were compared with those of seaurchins kept in a basket installed on the coast for one year. The duration of the described periods is not equal in the variables a, b, c, in Fig. 3. However, they were ordered, to contrast them with a variance test, as follows:  duration of periods in months F3;13 = 2.4061 P >= 0.1142 (1 - P = 0.8857). The low probability found for the hypothesis suggests that the periods themselves are significantly different to each other and consequently well. defined. The cycles of the same figures, defined as the sequence of the periods A + B + C + D, were contrasted with a Variance test under the hypothesis that there are no significant differences among them. The cycles were ordered as follows: a = feeding rate of the captive animals in Chiloé, b) = feeding rate of the captive animals in Valparaiso, c = feeding rate of a natural population in Valparaiso.  F2;14 = 0.6458: P = 0.5391 (1 - P = 0.4608). The result obtained suggests that there is a reasonable probability that there are variations in the feeting rate during the annual cycle, however, their cycles remain essentially similar despite the differing ecological conditions and the wide geographical separation. It also confirms that this species has a seasonal feeding pattern. These results were not tested with animals reared indoors. The results suggests that the main periods may however, be longer so that- A = autumn or autumnwinter or winter declination to starvation-winter, B = winter spring recover, C = springer-winter summer or summer declination to starvation, D = summerautumm recover. DISCUSSIONThe plant species identified in the digestive tract of Loxechinus albus do not necessarily imply that these animals have a restricted preference towards certain species though the 10 Rhodophyta species identified in the analysis(corresponding to 45.5% of all algae) suggest that they have some importance as food source. Alveal et al. (1970) found values of between 48 and 71.4% in this group in the different ecological fronts of the studied area. Furthermore this study showed that the Phaeophyta were from 5 to 40% and the Chlorophyta between 12 and 20% of the algae, which are similar to the 27.2 and 22.7% respectively found in seaurchin stomachs in this study, though too much importance should not be attached to these results. However, it is more reasonable to consider that the results are pointing to a varied feedin capacity. The animal food identified in the stomach contents, however, seems have been ingested casually and only formed a small fraction to the total intake. Previous experiments with animals reared in captivity (Bückle et al., 1977b) , doubtless in a stress situation, and fed to saturation with a only few number of plant species proved that the animals could survive well. The growth however was retarded, caused probably by an unbalanced diet. A normal diet would seem to be characterized by a wide variety of food types. Sand grains may be ingested with other food items. This large food spectrum is for aquaculture purposes an important factor which can help in the establishment of a cheap diet using waste material from food factories combined with algae. The relationship between the feeding rate and the gonad development has been studied by many authors (Lawrense 1975) in a variety of species and environmental conditions. In L. albus there is no close relationship between the variables. The results may suggest that the variables are influenced directly or indirectly by physiological and environmental factors, though further investigations are needed to clarify this complicated system. An attempt was made to divide the yearly fluctuations of the feeding rate in a natural population of sea-urchin, in four periods corresponding to a natural process of changes, strongly related to biological and environmental factors. Food availability seems not to be the only factor which determines these periods. In spite of the small number of algal species given to the captive sea-urchins reared in Valparaiso and in Chiloé, they maintained periodical fluctuations in feeding. Similar to those of the animals of the natural population. The latter had the choice of a wide selection of food species. The periods do not depend on food availability as long as it is sufficient to support basic life. Fuji (1962) describing the feeding patern of Strongylocentrotus intermedius, cultivated in aquaria, established that in the "preand incompletion-spawning period the animals fed only rarely", and afterwards they became very voracious throughout the winter until the next breeding season. It seems that this species reared in laboratory conditions have only two periods in the feeding pattern, and no starvation period. By contrast S. purpuratus appears to show a well defined starvation period (Lasker and Boolootian, 1960). The results of the present investigation suggest that these experiments with S. purpuratus were conducted under more natural conditions, and caution should be exercized when interpreting the results of feeding experiments conducted under artificial conditions. On the other hand the results of this study also show that separate natural populations of seaurchins of the same species, although they retain food in their stomachs, maintain regular annual fluctuations in their feeding. The retention of food does not alter the annual cycle of ingesting algas especially, which would constitute a natural phase of the trophic process. It is therefore necessary to revise the concept of retention of food in these animals as a basic impediment in considering wether studies undertaken on this matter are valid. Conclusiones1 . The food and the feeding rate of the edible seaurchin Loxechinus albus which inhabits the coast of Chiloé in the Province of Valparaiso has been studied. 2. In the analysis of the stomach contents 22 algae were identified together with different animal species belonging to different zoological groups. The presence of sand in the samples is discussed. 3. The results do not permit any firm conclusions concerning the food preference in this species. 4. The annual feeding cycle has been tentatively divided into four feeding periods as follows; A autumn winter declination to starvation, B winter spring recovery, C = spring summer declination to starvation, D = summer autumn recovery. 5. The sequence of A + B + C + D defined a cycle, and has been compared with the feeding rate recorded in captive sea-urchins from Valparaiso and Chiloé fed with a restricted number of algal species. The feeding cycle observed in these two experiments were essentially similar to those observed in natural populations. 6. No direct relationship has been found between gonad development and feeding rate, Probably the processes are independent and their variations the result of a complex interaction of physiological and environmental factors. AgradecimientosThe first author of this paper is deeply grateful to the Swedish International Development Authority (SIDA) whose generous award of a fellowship made it possible to continue scientific work; to The Swedish Institute and to the Royal Swedish Academy of Science for the granted support and in particular to Armin Lindquist Director of the Institue of Marine Research, to the late Bertil Swedmark and to, Jarl-Ove Strömberg Director of the Marine Biological Station of Kristineberg. LITERATURAALVEAL, K., Estudios ficoecológicos en la región costera de Valparaíso. Rev. Biol. mar. Valparaíso, 1970. 7-88. (1) 14 -------- , H. ROMO y J. VAL'NZUELA, Consideraciones ecológicas de las regiones de Valparaíso y Magallanes. Rev. Biol. mar. Valparaíso, 1973. 1-29. (1) 15 BÜCKLE, L. F., CH. GUISADO, C. SERRANO, L. CÓRDOBA y E. VÁZQUEZ, An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México. Estudio del crecimiento en cautiverio del erizo Loxechinus albus (Molina) en las costas de Valparaíso, Chiloé, Chile. 1977a. 141-152. (1): 4 An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Estudios de la alimentación en cautiverio del erizo Loxechinus albus (Molina) en las castas de Valparaíso y Chiloé, Chile. 1977b. 153-160. (1): 4 ------ , CH. GUISADO, E. TARIFEÑO, A. ZULETA, L. CÓRDOVA y C. SERRANO, Ciencias Marinas (México) Biological studies on the seaurchin Loxechinus albus (Molina) (Echinoidea: Echinodermata). IV.- Maturation cycle and gonadal biochemical changes. 1977c. (2): 4 DAYTON, P. K., R. L. ROSENTHAL y L. C. MAHAN, Kelp communities in the Chilean archipelago. R/V Hero Cruise 72-5. Antartic. Jl.U.S., 1973. 34-35. 8: EBERT, T. A., Science, Negative growth and longevity in the purple sea-urchin Strongylocentrotus purpuratus (Stimpson). 1967. 557-558. 157: FUJI, A., Studies on the biology of the sea unchin. V. Food consumption of Strongylocentrotus intermedius. Jap. J. Ecol., 1962. 181-186. (5): 12 GUTIÉRREZ, J. and I. OTSU, Periodicidad en las variaciones biométricas de Loxechinus albus Molina. Rev. Biol. mar. Valparaíso, 1975. 179-199. (2): 15 LAWRENCE, J. M., On the relationships between marin plants and sea urchins. Oceanogr. Mar. Biol. Ann. Rev., 1975. 213-286. 13: LEWIS, J. B., Feeding and digestion in the tropical sea urchin Diadema antillarum Philippi. Can. J. Zool., 1964. 549-557. 42: MORTENSEN, TH., A Monograph of the Echinoidea. III. 3. C. A. Reitzel, Copenhagen, 1943. b. 446 p. SCOTT, F. H., Food of the sea urchin Strongylocentrotus droebachiensis. Contr. Canad. Biol., 4901 1902. 49-54. 2:
|