|
ECOLOGÍA DE LOS SISTEMAS
FLUVIO-LAGUNARES ASOCIADOS A LA LAGUNA DE TÉRMINOS. EL HABITAT Y
ESTRUCTURA DE LAS COMUNIDADES DE PECES
Trabajo presentado en el VI
Congreso Nacional de Oceanografía, Ensenada B. California,
México, 10 al 13 de abril, 1978. Proyecto Especial del Programa Regional
de Desarrollo Científico y Tecnológico de la Organización
de los Estados Americanos (OEA). Recibido el 26 de abril de 1978 y aceptado
para su publicación el 18 de julio de 1978.
FELIPE AMEZCUA-LINARES
ALEJANDRO YÁÑEZ-ARANCIBIA
Universidad Nacional Autónoma de México, Centro de
Ciencias del Mar y Limnología, Laboratorio de Ictiología y
Ecología Estuarina. Contribución 122 del Centro de Ciencias del
Mar y Limnología, UNAM.
Un total de 3,838
ejemplares, de 37 colectas diurnas, comprendiendo 47 especies y 26 familias,
fueron muestreadas con una red de arrastre en los 4 sistemas fluvio-lagunares
de Laguna de Términos; Le., 1) Candelaria-Panlau, 2) Chumpan-Balchacah,
3) Palizada-del Este, 4) Pom-Atasta, entre septiembre de 1976 y octubre de
1977.
Se considera que los ríos Candelaria, Chumpan y
Palizada tienen en conjunto una descarga mínima total de 6 x 109
m3/año y esto es determinante en la ecología del área.
Transparencia entre 20 y 40%, salinidades de 2 a 26 ppm, temperaturas de 20 a
28ºC (secas) y 26 a 32ºC (lluvias), sustrato limo-arcilloso con
menos de 25% de CaC03. La vegetación predominante son bosques de
Rhizophora mangle y la macrofauna béntica es la
típica de arrecifes de Crassostrea virginica; estas
asociaciones son distintas en detalle.
En este habitat la
diversidad íctica, abundancia y productividad pesquera, fueron
estudiadas para determinar sus variaciones estacionales en las 4 zonas y en
función del tiempo. Fueron considerados seis índices. Cambios
estacionales en diversidad y abundancia relativa resultan básicamente
por el reclutamiento de juveniles terminando el verano y comienzos de
otoño (a fines del periodo de lluvias). Los resultados muestran una gran
afinidad ecológica (58 a 76%) entre las comunidades de los 4 sistemas.
Los índices de diversidad fueron bajos no como consecuencia de
áreas contaminadas sino por condiciones de variación de
salinidad, temperatura, turbidez y disponibilidad de alimento. Estos factores
limitan las especies nectónicas capaces de vivir dentro de las lagunas.
Algunas especies contribuyen con gran número de ejemplares a las
características del habitat permaneciendo con gran densidad a
través de todo el año y, aparentemente, la energía es
optimizada. Los consumidores de primer orden fueron 16 especies (34%) con una
biomasa de 2.51 g/m2/año; los consumidores de segundo orden fueron 21
(42.5%) con una biomasa de 4.95 g/m2/año; los consumidores de tercer
orden fueron 10 (23.5%) con una biomasa de 1.0 g/m2/año; con lo cual se
estima una productividad pesquera de los sistemas fluvio-lagunares de 8.5
g/m2/año.. Cuatro especies se consideran residentes permanentes del
habitat: Arius melanopus, Sphoeroides testudineus, Eucinostomus gula y
Micropogon furnieri. Además de éstas, las especies típicas
por biomasa, número y frecuencia son: Bairdiella chrysura,
Bairdiella ronchus, Chaetodipterus faber, Epinephelus guttatus, Cichlasoma
fenestralum, Dasyatis sabina, Arius felis, Anchoa mitchilli mitchilli,
Cetengraulis edentulus, Bagre marinus y Achirus lineatus.
Estos sistemas presentan baja producción pesquera aunque de
especies comerciales de alta productividad en la Bahía de Campeche. La
importancia ecológica de estas zonas es fundamental puesto que
representan áreas de crianza y/o alimentación de numerosas
poblaciones de peces. Deben tener prioridad en el manejo y protección al
utilizar la zona costera, puesto que exportan energía hacia la propia
Laguna de Términos y consecuentemente a la Bahía de
Campeche.
A total of 3,838 individuals, from 37 diurnal collection,
comprising 47 fish species and 26 families, were trawled in four fluvial-lagoon
systems, i.e., 1) Candelaria-Panlau, 2) Chumpan-Balchacah, 3) Palizada-del
Este, 4) Pom-Atasta, between September 1976 and October 1977.
It is estimated that the rivers Candelaria, Chumpan, and Palizada have
in conjunction a minimum total discharge of 6 x 109 m3/year and this factor
determines the ecology of the area. The turbidity is high with transparency
from 20 to 40%, salinities from 2 to 26 ppt, temperatures from 20 to
28ºC (dry season) and 26 to 32ºC (rainy season), the substrate is
silty-clay with less than 25% of CaCo3. The marginal vegetation is
predominantly composed of Rhizophora mangle swamps, and the
benthic macrofauna is characterised by constituent communities typical of
Crassostrea virginica reefs; these association are detailed
discused.
In these habitat the fish diversity, abundance,
and fishery productivity were investigated in order to know its seasonal
variations in the four zones and during time. Six indices were studied. A
seasonal change in relative abundance resulted primarily from the influx of
juveniles in the late summer and autum (ending rainy season). The results shows
a great ecological afinity (58 to 76%) among the communities from the four
systems. Species number and weight diversity values were low not by consequence
of contaminated arcas by the stressful conditions; variations in salinities,
temperatures, turbidity, and availability of food. This factor limit the
species of nekton able to live inside the lagoons. Some species contribute most
of number in response to the characteristics of the habitat, and remain high in
density throughout the year, and apparently, energy is optimized. The first
order consumers were 16 species (34%) with a biomass of 2.51 g/m2/year; the
second order consumers were 21 (42.5%) with a biomass of 4.95 g/m2/year, the
third orden consumers were 10 (23.4%) with a biomass of 1.0 g/m2/year. The
fishery productivity estimated was 8.5 g/m2/year. Four species are considered
permanent resident of the habitat: Arius melanopus, Sphoeroides testudineus,
Eucinostomus gula, and Micropogon furnieri. This species and the following are
typical in the ecosystem by biomass, number, and frequency: Bairdiella
chrysura, Bairdiella ronchus, Chaetodipterus faber, Epinephelus guttatus,
Cichlasoma fenestratum, Dasyatis sabina, Arius felis, Anchoa mitchilli
mitchilli, Cetengraulis edentulus, Bagre marinus, and Achirus
lineatus.
This ecological system has low fishery
productivity, but of comercial species which present high productivity in the
Campeche Bay. The ecological importance of this zones is fundamental due to
they represent nursery and/or feeding arcas for various fish populations. It
should be priority it management and protection when man use the coastal zone,
because they act to export of energy to Terminos Lagoon and consequently to the
Campeche Bay.
El estudio de las lagunas costeras de la República Mexicana, reviste en la actualidad una considerable importancia debido a que una gran parte de la producción pesquera nacional se extrae de estos medios ambientes, que por otra parte resultan más accesibles ya que no es necesario contar con equipo costoso para la extracción del recurso. Dentro de estos sistemas costeros, la Laguna de Términos, en el estado de Campeche, presenta un particular interés debido a que es una de las Lagunas costeras más extensas del país con una área aproximada de 2,500 km2, y se encuentra situada frente a la zona más productiva del Golfo de México, la Sonda de Campeche. Además, por sí misma es un sistema con enorme potencial. Con anterioridad se ha puntualizado la alta productividad de estos medios ambientes así como el hecho de que un considerable porcentaje de especies de peces costeros con interés comercial, visitan estas áreas en algún momento de su vida. El escaso aprovechamiento de estos recursos costeros, hace necesario el conocimiento integral del ecosistema con el fin de efectuar una evaluación real y concreta del potencial pesquero del sistema, tanto para especies de peces actualmente explotadas como aquellos que aún no sufren presión de pesca alguna y que representan un recurso económico potencial. De acuerdo con estos lineamientos, el Centro de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, dio inicio en septiembre de 1976, tomando como base la Estación de Investigaciones Marinas "El Carmen", situada en Ciudad del Carmen, Camp., a un proyecto de Investigaciones de carácter multidisciplinario que forma parte de un amplio programa auspiciado por la Organización de Estados Americanos (OEA) a través de su Programa Regional de Desarrollo Científico y Tecnológico denominado "Recursos Bióticos en Lagunas Costeras, Manglares y Areas Adyacentes en América Latina (Proyectos Piloto Comparativos entre el Golfo de Nicoya, Costa Rica; Bahía de Puerto Viejo, Azua, República Dominicana; y Laguna de Términos, Campeche, México"). Uno de los planteamientos iniciales del mencionado proyecto, ha sido analizar la productividad biológica de la Laguna de Términos con el propósito de establecer el significado pesquero de la propia laguna y de plantear normas de explotación racional y adecuada. Dentro de este enfoque y como parte de los actuales estudios interdisciplinarios en desarrollo, el Laboratorio de Ictiología y Ecología Estuarina del Centro de Ciencias del Mar y Limnología, ha tomado a su cargo estudios específicos como el presente trabajo sobre los sistemas fluvio-lagunares asociados a la Laguna de Términos, los cuales se han desarrollado bajo los siguientes obejtivos: 1. Compilación biogliográfica de los estudios efectuados con anterioridad en la Laguna de Términos. 2. Descripción ecológica del área. 3. Presentación de un catálogo de la fauna ictiológica de los sistemas fluvio-lagunares, incluyendo una colección de referencia. 4. Análisis de la distribución, frecuencia y abundancia (densidad y biomasa) de las especies en el área, tanto en el espacio como en el tiempo. 5. Caracterización de grupos (familias) y especies predominantes. 6. Conocer las relaciones y niveles tróficos de los peces presentes. 7. Caracterización ictioecológica de los sistemas fluvio-lagunares de acuerdo a: a) Parámetros ambientales. b) Grupos ictiofaunísticos presentes. c) Indices de diversidad. d) Indices de afinidad. e) Productividad íctica en términos de densidad y biomasa ("standing crop"). f) Tendencia alimenticia de las especies. g) Presencia de macroinvertebrados y vegetación acuática. ANTECEDENTESCon anterioridad, la Laguna de Términos, ha sido objeto de gran variedad de estudios con diversos enfoques y diferentes objetivos. Sin embargo la mayoría coincide en considerar a la laguna como una área aparentemente de un alto potencial pesquero, que presenta una gran diversidad de habitats. Actualmente la laguna tiene una importancia comercial bien establecida, y de sus aguas se extrae gran variedad de peces y mariscos dentro de los que destacan por su volumen: el ostion, el camarón y caracoles. El deficiente aprovechamiento de estos recursos conjuntado con la falta de planeación en las pesquerías hace necesario una integración de todos los conocimientos que en la actualidad se tienen del sistema con el fin de concretar una correcta evaluación a corto y largo plazo del potencial pesquero de cada una de las especies explotadas, o bien de abrir perspectivas en especies aun inexplotadas, sobre todo de peces. Entre los trabajos más relevantes se puede mencionar el estudio biológico preliminar de Zarur (1961) ; el de foraminíferos de Ayala-Castañares (1963) ; de micromoluscos de García-Cubas (1963) y de ostrácodos de Morales (1966) . Entre los estudios planctónicos están: Silva-Bárcenas (1963), Loyo-Rebolledo (1965), Suarez-Caabro y Gómez-Aguirre (1965), Gómez-Aguirre (1965a y b y 1974), Laguarda-Figueras (1957) y Marrón-Aguilar y López-Ochoterena (1969). De bentos están los trabajos de Marrón-Aguilar (1957) y Solís-Wolfovits (1973). De vegetación sumergida: Sauer (1967) y Ortega (1969). De geología y procesos geológicos existen varios trabajos como los de Thom (1969), Phleger y Ayala-Castañares. (1971), Yañez-Correa (1963 y 1968) y el reciente de Cruz-Orozco et al. (1977). De recursos naturales generales están las publicaciones de Coll de Hurtado (1975) y, S.R.H. (1976). De recursos pesqueros y productividad: Carvajal (1973 y 1975), Signoret (1974), Toral (1971), Toral y Reséndez (1974), Ruiz (1975), y Bravo-Núñez y Yáñez-Arcibia (1979). De todos los estudios mencionados sólo unos pocos tratan de la fauna ictiológica: Toral (1971), Toral y Reséndez (1974) ; y sólo uno de ellos Carvajal (1975) trata específicamente sobre aspectos biológicos de dos especies de robalos. A su vez, Bravo-Núñez y Yáñez-Arancibia (1979) discuten la ecología y estructura de las comunidades de peces en la Boca de Puerto Real, y Yáñez-Arancibia y Amezcua-Linares (1979) analizan la ecología de una raya en particular. ÁREA DE ESTUDIOLa Laguna de Términos (Fig. l), se encuentra en el sureste del Golfo de México, dentro del estado de Campeche, entre los meridianos 91º10' y 92º 00' longitud oeste y los paralelos 18º20' y 19º00' de latitud norte. Tiene una forma elipsoide en sentido E a 0, su longitud es de 70 km y su parte más ancha de 30 km. El área aproximada es de 2,500 Km2 está separada del Golfo de México por la Isla del Carmen y presenta dos bocas permanentes que la comunican con el mar, el Carmen y Puerto Real. El patrón de circulación general del Golfo influye directamente en el embalse (Fig. 2). CLIMADe acuerdo con García (1973) el clima del área de la Laguna de Términos es A m w, es decir clima cálido húmedo, con lluvias en verano. Se registra una temperatura máxima de 36ºC, y mínima de 17ºC durante los meses de invierno. Las precipitaciones pluviales alcanzan valores entre 1,200 y 2,000 mm anuales; siendo los meses de julio a noviembre el periodo de mayor precipitación y en menor grado de noviembre a enero; la precipitación más baja sucede al final del invierno y principio de la primavera. Los vientos dominantes tienen una dirección NE y SE, sin embargo durante el invierno se presentan tormentas tropicales denominadas 11 nortes" precisamente por la predominancia de los vientos del cuadrante norte. Estos "nortes" son causa principal de las lluvias esporádicas que tienen lugar de noviembre a enero. El clima es importante en la medida que determina el suelo, la vegetación y la precipitación que condiciona en última instancia el drenaje que repercute en la evolución geomorfológica de la zona. En el trabajo de Bravo-Núñez y Yáñez-Arancibia (1979) se resumen datos climáticos para el área. Se considera época de lluvias de mayo a octubre y de secas de noviembre a abril. HIDROGRAFÍAVarios ríos de importancia descargan sus aguas a la Laguna de Términos, entre ellos destacan por su aporte el río Candelaria en el extremo noroccidental y cuya cuenca se localiza principalmente en la Península de Yucatán. Se ha calculado que el aporte de este río con sus afluentes es de 21.5 M3/S S.R.H. (1976), el cual conjuntamente con el río Mamantel forman la Laguna de Panlau la que comunica a Términos por medio de la Boca de los Pargos. Este conjunto forma el Sistema Candelaria Panlau. 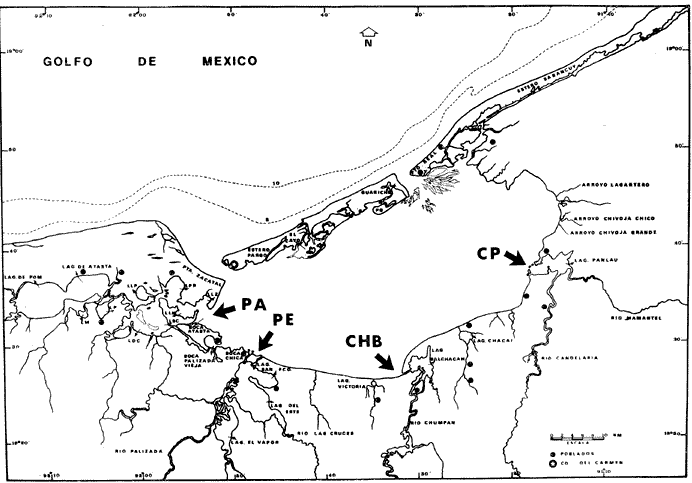 Fig. 1. Laguna de Términos, Campeche. Se señalan los principales rasgos fisiográficos y toponí micos. Las abreviaciones corresponden a: LLZ=Laguna Lodazal, LLN = Laguna de los Negros, LPR=Laguna de Puerto Rico, LLP=Laguna de los Patos, LDC=Laguna del Corte, LP=Laguna Palancares, LM=Laguna de los Muertos. Las flechas indican los cuatro sistemas fluvio-lagunares estudiados: CP=Candelaria-Panlau, CHB=Chumpan-Balchacah, PE=Palizada-del Este, PA=Pom-Atasta. 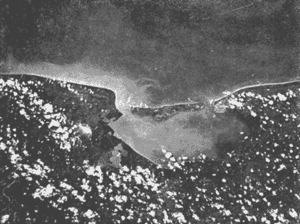 Fig.2. Vista aérea tomada por la nave espacial Geminis 5, que muestra la Laguna de Términos y áreas adyacentes. El patrón de circulación litoral determina la presencia de un delta interior en la Boca de Puerto Real y aguas claras en el sector oriental de la laguna así como también un delta externo en la Boca del Carmen y aguas turbias en el sector occidental, esto último también afectado por descargas fluviales. Fotografía, gentileza del Dr. Agustín Ayala-Castañares. Al extremo oriental de la Laguna de Términos desemboca el río Sabancuy y los arroyos Colax, Lagartero, Chivojá chico y Chivojá grande. El río Chumpan se forma en la planicie costera por los ríos Salsipuedes y San Joaquín, y desemboca finalmente a la Laguna de Balchacah. El cauce de este río tiene una área de 1,874 Km2 y un volumen de escurrimiento anual de 1,368 millones de m3. Río y laguna forman el Sistema Chumpan-Balchacah. El río Palizada forma parte de la red hidrológica de los ríos Mexcalapa, Grijalva y Usumacinta. Los ramales de este río, junto con otros ríos menores anexos dan origen a lagunas interiores; por un lado a las lagunas del Vapor, el Este y San Francisco que en su conjunto constituyen el Sistema Palizada-del Este, y por el occidente el Sistema Pom-Atasta con varias lagunas menores anexas. HIDROLOGÍALos vientos juegan un importante papel en la circulación del agua de la laguna. Los procedentes del sector norte dan origen a una circulación anticiclónica, mientras que vientos provenientes del sector sur generan una circulación ciclónica en la laguna. La corriente litoral del sistema fluye en dos sentidos principales, hacia el oriente y hacia el occidente, aunque es mucho más marcada en sentido occidental. La velocidad promedio de esta corriente es de 24.10 m/s Vargas (1978). La salinidad de la Laguna de Términos varía de acuerdo a la estación del año en relación a época de lluvias o de secas. El rango mínimo es de 12%o. Usualmente los valores más altos se registran en las inmediaciones de la Boca de Puerto Real por donde penetra agua marina del Golfo (38.2%o durante mayo, Botello 1978), decreciendo gradualmente a partir de la isla hacia tierra firme, de modo que aproximadamente al centro de la laguna existen valores intermedios (30%o, Botello 1978), encontrándose los valores más bajos en la zona afectada directamente por la descarga de los sistemas fluvio-lagunares; este fenómeno es más marcado en la porción occidental de la laguna. En general durante la época de lluvias cuando el aporte fluvial se incrementa, las salinidades disminuyen. Yáñez-Correa (1963) encontró entre los meses de marzo y abril de 1959, que existe un gradiente salino a partir de la Boca de Puerto Real donde se localizan los valores más altos, 39%o hacia el interior de la laguna decreciendo rápidamente hacia la porción continental donde se encuentra el área más afectada por el aporte fluvial; en estas zonas la salinidad de fondo cambia mucho menos intensamente que la de superficie. La salinidad decrece hacia la parte occidental de la isla hasta 29%o en la porción más ancha; a partir de este sitio aumenta rápidamente hacia el Canal del Carmen donde existe una marcada estratificación. Botello (1978), menciona que existe muy poca o ninguna estratificación vertical en la laguna. En este estudio, las salinidades en el Sistema Candelaria-Panlau fluctuaron durante mayo entre 5%o, en las zonas más altas, a 25%o enfrente de la Boca de Pargos dentro de la Laguna de Términos; durante agosto se registraron valores ligeramente más altos entre 8%o y 26%o en las localidades mencionadas. En el sistema Chumpan-Balchacah el rango salino fue de 26 a 28%o en mayo y de 10%o a 22%o en octubre debido, en este último caso, a la influencia de las lluvias. En Palizada-del Este se observaron valores un poco más bajos que en los sistemas anteriores ya que aquí la mínima fue de 2%o en enero y la máxima 25%o en marzo. En el Sistema Pom-Atasta los valores oscilaron entre 2%o y 19%o. En los cuatro sistemas fluvio-lagunares se observa por lo general rangos que van desde salobres (25%o) a casi dulceacuícolas (2%o) que varían de acuerdo a la temporada de precipitación pluvial y al sitio específico del mismo sistema, ya que se puede reconocer un gradiente desde las porciones más altas hasta la desembocadura del sistema con la Laguna de Términos. La temperatura sufre variaciones estacionales dentro de un rango de 15.7ºC. Las más bajas se registran durante el invierno (23.4ºC) en el área de Palizada Vieja y la más alta en abril en el área de Ensenada. De acuerdo con Toral (1971) el valor promedio de las temperaturas en la laguna es de 29.9ºC; Yáñez-Correa (1963) localiza la isoterma superficial de los 30ºC frente a Palizada Vieja; mientras que en Boca de Atasta localiza la isoterma superficial de los 28ºC. En general la temperatura del agua se mantiene uniforme en el cuerpo principal de la laguna. En este estudio, para el sistema Candelaria-Panlau se obtuvieron valores desde 28 a 30ºC en mayo y 29 a 31ºC en agosto. En Chumpan-Balchacah y Palizada-del Este los rangos son similares 28 a 31ºC, aunque en este último la temperatura es menor durante marzo, 24ºC. En Pom-Atasta se obtuvo la variación máxima con un rango de 20ºC en enero a 32ºC en agosto. Los valores de oxígeno disuelto por lo regular son cercanos o mayores a la saturación debido a la alta oxigenación causada por la intensa acción de los vientos. Botello (1978) reporta durante el mes de mayo rangos de saturación entre 86 y 150% con valor promedio de 99.6% de saturación. En general el oxígeno disuelto tiene una variación entre los 3.6 m/l en la zona del Cayo (abril 1970) y los 8.1 m/l en Ensenada (julio 1970) Toral (1971). Los productores primarios también influyen en la buena oxigenación. En el presente trabajo se registraron valores de 02 desde 2.2 ml/l en el Sistema Chumpan-Balchacah durante mayo, a 7.2 ml/l, en Candelaria-Panlau en agosto. En general el oxígeno disuelto siempre se encuentra muy por arriba de los rangos mínimos tolerables de los peces en sistemas estuarinos. La transparencia de la laguna varía considerablemente de acuerdo a la localidad específica de que se trate, así mientras que para la Boca de Puerto Real se registran valores altos en septiembre, para los sistemas fluvio-lagunares la transparencia es mucho menor debido al abundante aporte sedimentario de los ríos que enturbian grandemente estos sistemas. BATIMETRÍA Y SEDIMENTOSEl perfil batimétrico de la laguna es bastante homogéneo y en general tiene poca profundidad, ya que se encuentra saturada por la gran cantidad de sedimentos aportados por los ríos que en ella confluyen, los que representan un volumen aproximado de 19.24 x 10.9 m3 (Vargas, 1978) . Las zonas más profundas son precisamente las bocas donde existen profundos canales; así entre la Punta Xicalango y la Isla del Carmen se registran profundidades hasta de 12 y 15 m, siendo por esto la parte más profunda de la laguna, ya que en la Boca de Puerto Real, entre la Isla del Carmen e Isla Aguada, la profundidad máxima está entre 8 y 10 m. En promedio, la profundidad es entre 2 y 3 m. Al centro de la laguna hay 4 m de profundidad, y disminuye paulatinamente hacia la periferia donde existen fondos muy someros de unos 0.3 m. La mayor parte de la laguna está ocupada por sedimentos de limo y arcilla. Yáñez-Correa (1963) indica que toda la porción norte que se encuentra frente a la isla del Carmen presenta fondos de arena con concha de grano fino y grueso, mientras que en la porción suroriental existen sedimentos limo-arcillosos y en la suroccidental es fangosa. Los sedimentos de la laguna muestran un alto contenido de carbonato de calcio, localizándose los valores más altos en las inmediaciones de la Boca de Puerto Real, donde Yáñez-Correa (1963) reporta entre un 50 y 70%. La alta concentración de carbonatos que se disuelven en el agua dan lugar a un pH alcalino; Botello (1978) reporta para el mes de mayo valores de 8.1 a 8.2 en la parte central y occidental de la laguna, mientras que en la porción oriental el pH aumenta de 8.2 a 8.38. Para otros meses el mismo autor, coincide con Yáñez-correa (1963) en estimar la presencia de mayor cantidad de carbonatos en la porción oriental de la laguna. En las zonas ocupadas por bancos de Thalassia testudinum el contenido de materia orgánica en los sedimentos es muy alto. En los sistemas fluvio-lagunares el sedimento es limo-arcilloso con menos de 25% de carbonato de calcio. Lo más notable de estos sistemas es la presencia de arrecifes ostrícolas establecidos sobre todo en Candelaria-Panlau, Chumpan-Balchacah y Pom-Atasta. En este último sistema existe una abundante explotación del molusco ya que de aquí se extrae casi la totalidad de la producción. Los ostiones le dan a los sistemas características particulares en el habitat. Además da origen a zonas muy someras, siendo las zonas más profundas las desembocaduras de los ríos y canales donde existe una profundidad promedio de 2 m. VEGETACIÓNEl rasgo notable es que la vegetación costera emergente en los márgenes de la laguna, están casi por completo cubiertos por manglares y los sistemas fluvio- lagunares, esteros, canales y pequeñas lagunetas asociadas no son las excepción (Fig. 3). La importancia del mangle dentro de la laguna ha sido reconocida desde los estudios de Zarur (1961) quien reconoce a estas asociaciones como fijadoras del suelo que permiten una posterior implantación de flora terrestre. Además se ha visto que en otros lugares de características similares es muy significativo que el aporte de materia orgánica al sistema, proviene de los manglares; Heald (1969 y 1970), Heald et al. (1974), Odum y Heald (1972 y 1975) Odum et al. (1973), Snedaker y Lugo (1973).  Fig. 3. Vegetación costera emergente predominantemente en los sistemas estudiados. Pueden encontrarse Rhizophora mangle, Avicenia germinans y Laguncularia racemosa La especie más abundante en los manglares es Rizophora mangle; sin embargo se encuentra asociada con Avicenia germinans y Laguncularia racemosa. Sobre la Isla del Carmen existen conjuntamente con manglares, grandes cocotales, Junto con matorrales de Acacia sp. Randia acubate y Coccoloba uvifera.  Fig. 4. Parches de Thalassia testudinum con algunas macroalgas Phaeophycaea y Rhodophycaea "sueltas" que pueden encontrarse en las bocas de algunos sistemas. Esto es más característico en el sistema Candelaria-Panlau donde además se presenta Ruppia maritima en el interior. 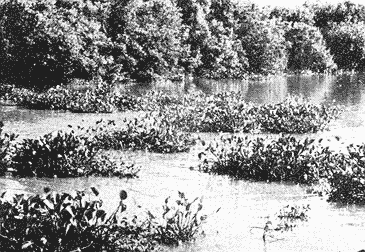 Fig.5. Lirio acuatico, Eichhornia chassipes es abundante, particucularmente en el interior del Sistema Palizada-del Este. Las aguas son turbias. La vegetación sumergida está formada principalmente por grandes praderas de Thalassia testudinum tanto en la porción norte de la laguna como frente a las bocas de algunos de los sistemas fluvio-lagunares (Fig. 4). También se ha encontrado Ruppia maritima en el interior de Candelaria-Panlau. Así mismo existen grandes extensiones cubiertas de macroalgas del tipo Phaeophyceae como Gracilaria sp y otras en la comunicación de estos sistemas con la Laguna de Términos. El lirio acuático Eichhornia crassipes se encuentra cubriendo grandes extensiones en la porción alta del Sistema Palizada-del Este (Fig. 5). Aquí mismo existen abundantes pastos de pantanos del tipo "Juncos" (Typha sp.) y "carrizales", lo mismo sucede en el resto de los sistemas pero en menor proporción (Fig. 6).  Fig.6. Los pastos de pantanos, tipo "juncos" (Typhasp) y "carrizales"completan el cuadro de productos primarios en el lirio y el manglar MATERIAL Y MÉTODOSLas capturas de peces fueron efectuadas con una red de arrastre camaronera de 10 m de largo, 9 m de amplitud con 5 m de abertura de trabajo, equipada con tablas de 0.8 x 0.5 m y luz de maya de 3/4", a bordo de una lancha de fibra de vidrio con 7 m de eslora y motor fuera de borda de 40 H.P. Se efectuaron colectas diurnas en los sistemas fluvio-lagunares durante un ciclo anual repartidos como sigue: la. colecta del 20 al 24 de septiembre de 1976 2a. colecta del 18 al 27 de octubre de 1976 3a. colecta del 18 al 22 de enero de 1977 4a. colecta del 2 al 11 de marzo de 1977 5a. colecta del 2 al 6 de mayo de 1977 6a. colecta del 16 al 20 de agosto de 1977 7a. colecta del 24 al 26 de octubre de 1977 En todos los casos las capturas no sobrepasaron los 4 m de profundidad y fueron efectuadas a diversos niveles, desde las bocas de los sistemas fluvio-lagunares con la Laguna de Términos, hasta la parte más interna donde arriban los ríos (Figs. 7 a 10). En cada estación se determinó el área muestreada con el fin de cuantificar densidad y biomasa de la fauna ictiológica. Se hizo un total de 17 estaciones repartidas de la siguiente manera: Sistema Candelaria-Panlau, 4 estaciones; Sistema Chumpan-Balchacah, 3 estaciones, y Sistema Palizada-del Este y Pom-Atasta con 5 estaciones cada una. Tales estaciones se denominaron con las iniciales de cada sistema, así las estaciones en Candelaria-Panlau se denominaron CP; las de Chumpan-Balchacah CHB, las de Palizada-del Este PE, y las Pom-Atasta PA. Con cada captura se tomaron los siguientes parámetros ambientales: temperatura, salinidad y oxígeno de superficie y fondo, transparencia, profundidad, tipo de fondo y mareas. La temperatura fue medida por medio de un termómetro de cubeta marca Kahlsico con precisión de 0.1ºC. La salinidad se midió con un refractómetro American Optical de lectura directa y con temperatura compensada. El oxígeno disuelto se cuantificó por medio del método químico de Winkler y mediante un oxímetro, marca Kahlsico. La transparencia se hizo por medio de un disco de Secchi. Adicionalmente, se efectuaron observaciones de la vegetación sumergida, macrofauna bentónica y datos climáticos. Los individuos capturados, fueron preparados con una incisión ventral y fijados en una solución de formalina al 10% neutralizada con borato de sodio. Todo el material debidamente etiquetado y envasado fue trasladado al laboratorio para su análisis. MATERIAL DE ESTUDIODe un total de 37 colectas en las 17 estaciones, se obtuvieron 3,838 ejemplares repartidos de la siguiente manera: Sistema Candelaria-Panlau 2,003 ejemp. Sistema Chumpan-Balchacah 582 ejemp. Sistema Palizada-del Este 402 ejemp. Sistema Pom-Atasta 851 ejemp. ACTIVIDADES DE LABORATORIOEn el laboratorio se utilizaron diversos instrumentos para el estudio, dentro de los principales están: microscopios estereoscopios marca SWIFT con diversos aumentos, balanza granataria marca OHAUS con capacidad de 2,610 g y lectura mínima de 0.1 g, báscula marca "JACOBS" con capacidad de 10 kg y división mínima de 25 g, y un ictiómetro convencional de 50 cm. Los peces, flora y fauna acompañante fueron separados, lavados, reetiquetados y reenvasados en frascos de vidrio, usando alcohol metílico al 70% como preservador, para su estudio. La identificación taxonómica se hizo por medio de la literatura básica, fundamentalmente con los trabajos clásicos de Jordan y Everman (1896-1900) y Meek y Hildebrand (1923, 1925, 1928), así como los de Hildebrand (1943), Castro (1965) y Reséndez (1970 y 1973). Para las mediciones se utilizó la longitud total. 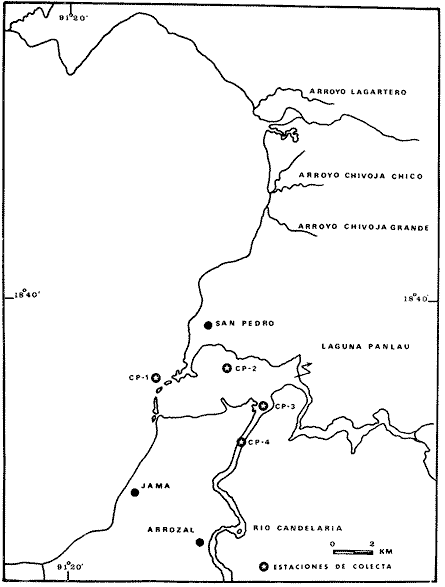 Fig. 7. Sistema fluvio-lagunar Candelaria-Panlau en el sureste de la Laguna deTérminos. Cuatro estaciones de colecta: CP-1 en la boca del sistema sobre bancos de Crassostrea virginica, Thalassia testudinum y macroalgas; CP-2 y CP-3 en el interior del sistema sobre bancos de Ruppia maritima; CP-4 en ambiente fluvial con gran cantidad de detritus. Orillas de Rhizophora mangle. 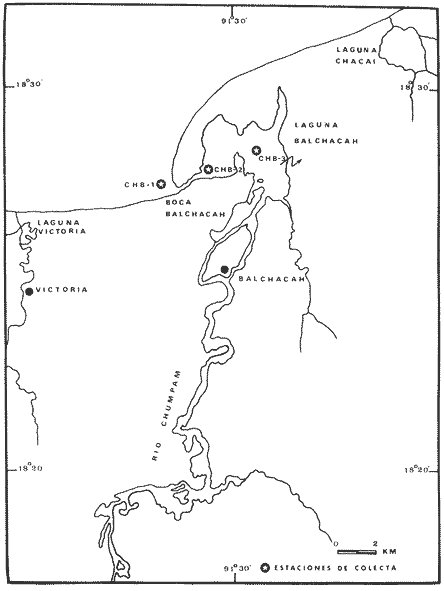 Fig. 8. Sistema fluvio-lagunar Chumpan-Balchacah en el sur de la Laguna de Términos. Tres estaciones de colecta: CHB-1 en la boca del sistema; CHB-2 en el canal de entrada sobre bancos de Crassostrea virginica; CHB-3 en el interior del sistema en ambiente fluvial con gran cantidad de detritus. Orillas con Rhizophora mangle. 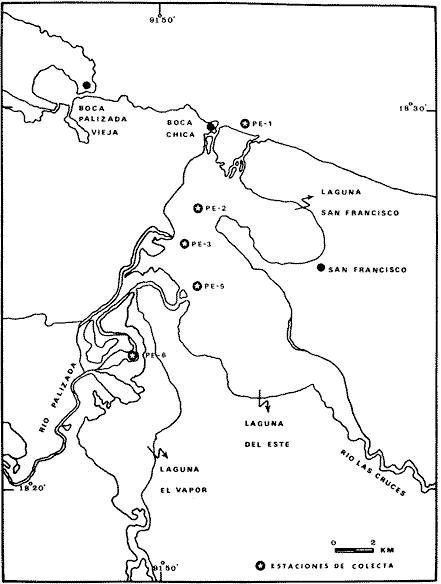 Fig. 9. Sistema fluvio-lagunar Palizada-del Este en el sur de la Laguna deTérminos. Cinco estaciones de colecta: PE-1 en la entrada del sistema; PE-2 y PE-3 en el interior frente a bosques de Rhizophora mangle; PE-5 y PE-6 sobre el delta del Río Palizada con vegetación de lirio acuático (Eichhornia crassipes) y pastos de pantanos y carrizales. 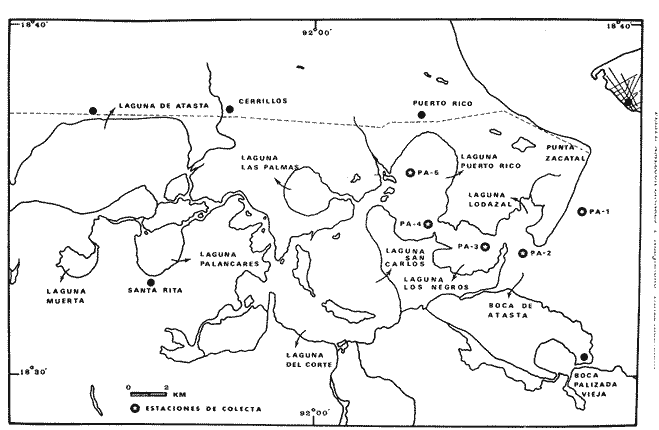 Fig. 10. Sistema fluvio-lagunar Pom-Atasta al oeste de la Laguna de Términos. Cinco estaciones de colecta. PA-1 y PA-2 en la boca del sistema frente a bosques de Rhizophora mangle; PA-3, PA-4 y PA-5 en el interior del sistema con la misma vegetación y gran cantidad de detritus. Bancos de Crassostrea virginica se encuentra en las estaciones PA-2 y PA-3. La transparencia del agua fue estimada porcentualmente relacionándola con la profundidad de acuerdo a la siguiente fórmula: 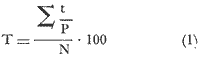 dondeT = porcentaje anual de la transparencia; t = transparencia en un muestreo; P = profundidad, en el muestreo correspondiente y N = número total de eventost/P . Las comunidades de peces están integradas por diversos componentes, éstos fueron determinados de acuerdo a su origen y frecuencia de aparición en los muestreos de los sistemas fluvio-lagunares. Así, las especies con porcentajes defrecuencia cercana al 100% serían las residentes permanentes, mientras que especies con bajo porcentaje corresponden a especies visitantes ocasionales. Tales valores se obtienen de acuerdo a la relación: 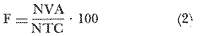 donde F = porcentaje de frecuencia; NVA = número de veces de aparición de una especie en una estación dada y NTC = número total de colectas en la estación correspondiente. La diversidad se consideró como el número de especies para cada colecta y su relación con el número total de individuos de todas las especies de la colecta. Se consideró uníndice de diversidad para cada colecta de acuerdo a la expresión matemática propuesta por Shannon y Weaver (1963): 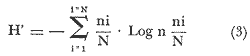 dondeni = número de individuos de una especie en una colecta, y N = número total de individuos de todas las especies en la colecta. Aún cuando esta función considera diversidad y abundancia, es recomendable utilizar también otros índices que tratan estos aspectos por separado. Así, para el cálculo de la riqueza de especies como componente de la diversidad, se usó la siguiente expresión que ha sido utilizada por Margalef (1969) y otros autores en áreas de lagunas costeras. D = (S - 1) /log n (4) donde S es el número de especies y N el número de individuos. El índice de equitatividad propuesto por Pielou (1966), obtiene información que permite discutir la abundancia relativa de las especies; éste se expresa como sigue: J' = H'/ H max = H'/ log nS (5) donde log de S es el valor máximo de H'. Entonces H' = H' max. cuando las especies son igualmente abundantes. Del mismo modo, se consideró un índice de biomasa que se obtiene utilizando la fórmula de Shannon y Weaver (1963), en la que se reemplaza el número de individuos de una estación en una colecta, por gramos de las mismas, y el número total de individuos de todas las especies de la colecta por los gramos totales de la misma. Esta modificación ha sido inicialmente considerada por Wilhm (1968). La función matemática queda entonces: 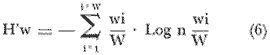 donde wi son los gramos de una especie en una colecta yW los gramos totales de todas las especies de la colecta. Todos los cálculos de diversidad se efectuaron sobre la base de logaritmos naturales. Ahora de todo lo anterior, la abundancia de la ictiofauna fue determinada como densidad y biomasa, esto es indiv. m-2 y g m -2. Se estableció un índice deafinidad sobre la base deespecies y poblaciones comunes de peces. Esto se obtiene comparando todas las áreas entre sí, en este caso los 4 sistemas fluvio-lagunares en estudio, y se objetiva graficando en un diagrama de afinidad. La expresión matemática para estos cálculos es: 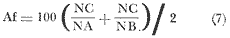 donde Af = al índice porcentual de afinidad,Na = número de especies del áreaA, NB = número de especies del área B y NC = número de especies comunes entre el área A y B. La categorización ictiotrófica fue hecha en base a las categorías propuestas para peces de ambientes lagunares-estuarinos por Yáñez-Arancibia (1977 y 1978). Tales categorías son las siguientes: 1. Consumidores Primarios: en esta categoría se incluyen peces planctófagos (fito y zoo) , detritívoros y omnívoros que consumen detritus, vegetales y fauna de tamaño pequeño. 2. Consumidores Secundarios: aquí se incluyen peces predominantemente carnívoros, aún cuando pueden incluir en su dicta algunos vegetales y detritus, pero con poco significado cuantitativo. 3. Consumidores de Tercer Orden: incluye peces exclusivamente carnívoros, donde los vegetales y el detritus son alimentos accidentales. RESULTADOS
COMPOSICIÓN DE LA FAUNA ICTIOLÓGICADel análisis de 3,838 ejemplares obtenidos en 37 capturas repartidas en 17 estaciones, fueron identificadas 47 especies agrupadas en 40 géneros y 26 familias. El ordenamiento sistemático fue hecho de acuerdo al criterio de Greenwood et al. (1966 y 1967) para categorías supragenéricas, mientras que para géneros y especies se adoptó principalmente el criterio de Jordan y Evermann (1896-1900), Meek y Hildebrand (1923-1928) y Cervigón (1966 a y b, 1975). LISTA TAXONÓMICA DE LAS ESPECIES
Clase Chondrichthyes
Orden Batoidea
Suborden Myliobatoidea
I Familia Dasyatidae
Género 1. Dasyatis Rafinesque, 1810
1) Dasyatis sabina (Le Sueur, 1824)
Clase Osteichthyes
Divición I Taeniopaedia
Superorden Clupeomorpha
Orden Clupeiformes
Suborden Clupeoidei
II Familia Clupeidae
Género 2. Sardinella Cuvier y Valenciennes, 18472) Sardinella Macrophthalmus (Ranzani, 1842) 3) Sardinella humeralis (Cuvier y Valenciennes, 1847) Género 3. Opisthonema Gill, 18614) Opisthonema oglinum Le Sueur, 1782 III Familia Engraulidae
Género 4. Anchoa Jordan Evermann, 19275) Anchoa Mitchilli mitchilli (Cuvier y Valenciennes 1848) 6) Anchoa hepsetus hepsetus (Linnaeus, 1785) Género 5. CetengraulisGünther, 18687) Cetenngraulis edentulus (Cuvier, 1829 División III Euteleosyei
Superorden Protacanthopterygii
Orden Salmoniformes
Suborden Myctophoidei
IV Familia Synidontidae
Género 6. Sinodus Scopeli, 17778) Synodus foetens (Linnaeus, 1766 Superorden Ostariophysi
Orden siluriformes
V Familia ictaluridae
Género 7. Ictalurus Rafinesque, 18209) Ictalarus meridionalis (Günther, 1860 VI Familia Ariidae
Género 8. Bagre Oken, 181710) Bagre Marinus (Mitchill, 1915) Género 9. Arius Cuvier y Valenciannes, 193711) Arius felis (Linnaeus, 1766) 12) Arius melanopus Günther, 1864. Superorden Paracanthopterygii
Orden Batrachoidiformes
Familia Batrachoidiformes
Género 10. Opsanus Rafinesque, 181813) Optanus beta (Goode y Bean, 1937) Género 11. Nautopaedium Jordan, 191914) Nautopaedium Prosissimum (Cuvier y Valenciennes, 1937 Superorden Atherinomorpha
Orden Atheriniformes
Suborden Exocoetoidei
VIII Familia Exocoetidae incluye Hemirhamphidae
Género 12. Hyporhamphus Gill, 186015) Hyporhamphus unifasciatus (Ranzani, 1842) Superorden Acanthopterygii
Orden gasterosteiformes
Suborden Syngnathoidei
IX Familia Syngnathidae
Género 13. Syngnathus Linnaeus, 185816) Syngnathus mackayi (Swani y Meek, 1884) Orden Perciformes
Suborden Percoidei
X Familia Centropomidae
Género 14. Centropomus Lacépède, 180217) Centropomus undecimalis (Bloch, 1729) XI Familia Serranidae
Género 15. Epinephelus Bloch, 179318) Epinephelus guttatus (Linnaeus, 1758) XII Familia Carangidae
Género 16. Caranx Lacépède, 180219) Caranx hippos (Linneus, 1766) Género 17. Chloroscombrus Girard, 195920) Chloroscombrus Chrysurus (Linnaeus, Género 18. Selene Lacépède, 180321) Selene vomer (Linnaeus, 1758) Género 19. Olipoplites Gill, 186322) Oligoplites saurus (Bloch y Schneider, 1801) XIII Familia Lutjanidae
Género 20. Lutjanus Bloch, 197023) Lutjanus griseus (Linnaeus, 1758) XIV Familia Gerridae
Género 21. Eucinostomus Baird y Girard, 185424) Eucinostomus gula (Cuvier y Valenciennes, 1830) 25) Eucinostomus argenteus Baird y Girard, 1854 Género 22. Eugerres Jordan y everman, 192726) Eugerres plumieri (Cuvier y Valenciennes, 1830) Género 23. Diapterus ranzani, 184027) Diapterus rhombeus (Cuvier y Valenciennes, 1830) XV Familia Sparidae
Género 24. Archosargus Gill, 186528) Archosargus prabatocephalus (Walbaum, 1792) XVI Familia Sciaenidae
Género 25. Micropogon Cuvier y Valenciennes, 183029) Micopogon furnieri (desmarest, 1802) Género 26. Bairdiella Gill, 186130 Bairdiella chrysura (Lacépède, 1802) 31) Bairdiella ronchus (Cuvier y Valenciennes, 1830) Género 27. Cynoscion Gill, 185432) Cynoscion nebulosus (Cuvier y Valenciennes, 1830) 33) Cynoscion nothus (Holbrook, 1850) XVII Familia Ephippidae
Género 28. Chaetodipterus lacépède, 180334) Chaetodipterus faber (Braussonet, 1872) XVIII Familia Cichlidae
Género 29. Cichlidae35) Cichlasoma urophthalmus (Günther, 1862) 36) Cichlasoma fenestratum (Günther, 1860) Suborden Polynemoidei
XIX Familia Polynemoide
Género 30. Polynemus Linnaeus, 175837) polynemus octonemus Giard, 1858 Suborden Gobioidei
XX Familia Gobiidae
Género 31. Gobionellus Girard, 185838) Gobionellus oceanicus (Pallas, 1770) Género 32. Gobiosoma Girard, 185839) Gabiosoma bosci (Lacépède, 1758) Suborden scombroidei
XXI Familia Trichiuridae
Género 33. Trichiurus Linnaeus, 175840) Trichiurus lepturus Linnaeus, 1758 XXII Familia Scombridae
Género 34. Scomberomorus Lacépède, 180241) Scomberomorus maculatus (Mitchill, 1815) Orden Pleuronectiformes
Suborden Pleuronectoidei
XXIII Familia Bothidae
Género 35. Cytharichthys Bleeker, 186242. Cytharichthys spilopterus Günther, 1862 Género 36. Etropus Jordan y Gilbert, 188143) Etropus crossotus Jordan y Gilbert, 1881 Suboreden Soleoidei
XXIV Familia Soleidei
Género 37. Achirus Lacépède, 180344) anchirus lineatus. (Linnaeus, 1725) Género 38. Trinectes Rafinesque, 183145) Trinectes maculatus (Bloch y Schneider, 1801) XXV Familia Cynoglossidae
Género 39. Symphurus rafinesque, 181046) Symphurus plagiusa Linnaeus, 1766 Orden Tetraodontiformes
Suborden Tetraodontoidei
XXVI Familia tetraodontisae
Género 40. Shoeroides Dumeril, 180647) Shoeroides testudineus linnaeus, 1743 ESTRUCTURA DE LAS COMUNIDADES
COMPONENTES COMUNITARIOSLa dinámica de los sistemas fluvio-lagunares de la Laguna de Términos determinan que las poblaciones de peces que allí se encuentran pueden ser agrupadas en tres componentes comunitarios: 1) Especies visitantes ocasionales,tanto de origen marino como dulceacuícola. A este grupo corresponde una frecuencia entre 1 y 30%, con un total de 25 especies (55.3%). 2) Especies que utilizan los sistemas fluvio-lagunares como áreas naturales de crianza y/o para alimentarse. Corresponde una frecuencia de 31 a 70% con un total de 17 especies (36.2%). 3) Especies residentes permanentes, con frecuencia en las capturas de 71 a 100% y con sólo 4 especies (8.5%). En el Sistema Candelaria-Panlau, se presentan 15 especies (32%) que son visitantes ocasionales; 17 especies (36%) que utilizan el habitat como áreas de crianza y/o para alimentarse; y 4 especies (8.5%) que son residentes permanentes. En el Sistema Chumpan-Balchacah, 6 especies (13%) son visitantes ocasionales; 11 especies (23%) utilizan el habitat como áreas de crianza y/o para alimentarse; y 4 especies (8.5%) son residentes permanentes. En Palizada-del Este se encontraron 9 especies (19%) visitantes ocasionales; 15 especies (32%) que utilizan el habitat como áreas de crianza y/o para alimentarse; y 4 especies (8.5%) son residentes permanentes. Por último en el Sistema Pom-Atasta aparecieron 14 especies (30%) como visitantes ocasíonales; 15 especies (32%) que utilizan el habitat como áreas de crianza y/o para alimentarse; y 4 especies (8.5%) son residentes permanentes. La cuantificación de las tablas 1 a 4 se basa en el registro total anual. Por lo tanto el panorama refleja valores máximos. Para cada uno de los sistemas, los componentes comunitarios varían en el tiempo, de acuerdo a los patrones ecológicos en juego. Este cambio sucesional comunitario, puede apreciarse en la tabla 9 y en la figura 11.  TABLA 1 COMPONENTES COMUNITARIOS DEL SISTEMA FLUVIO-LAGUNAR CANDELARIA-PANLAU  TABLA 2 COMPONENTES COMUNITARIOS DEL SISTEMA FLUVIO-LAGUNAR CHUMPAN-BALCHACAH  TABLA 3 COMPONENTES COMUNITARIOS DEL SISTEMA FLUVIO-LAGUNAR PALIZADA-DEL ESTE  TABLA 4 COMPONENTES COMUNITARIOS DEL SISTEMA FLUVIO-LAGUNAR. POM-ATASTA 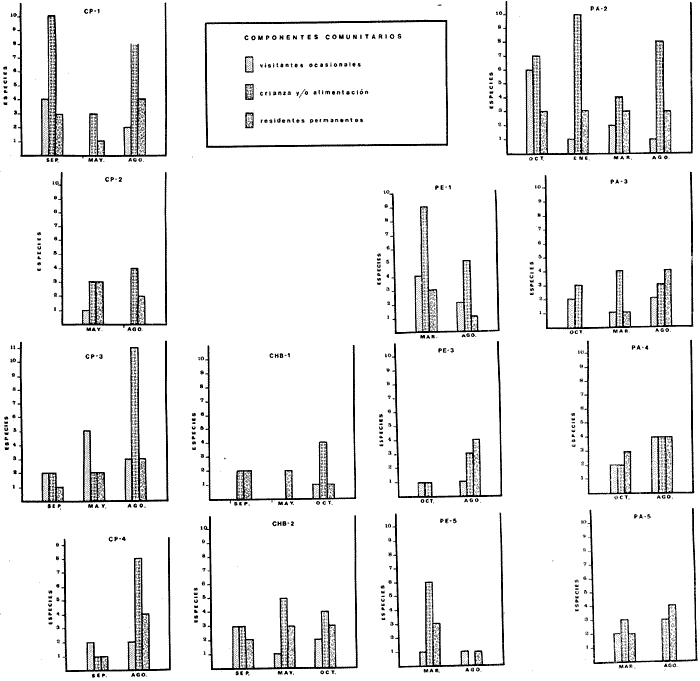 Fig. 11. Variacion temporal y proporciones entre los 3 tipos de componentes comunitarios en la estructura de las ictiocomunidades de los 4 sistemas fluvio- lagunares estudidos .CP=Candelaria-Panlau, CHB=Chumpan-Balchacah, PE=Palizada -del Este y PA= Pom-Atasta.Los números asociados corresponden a estaciones de colecta. 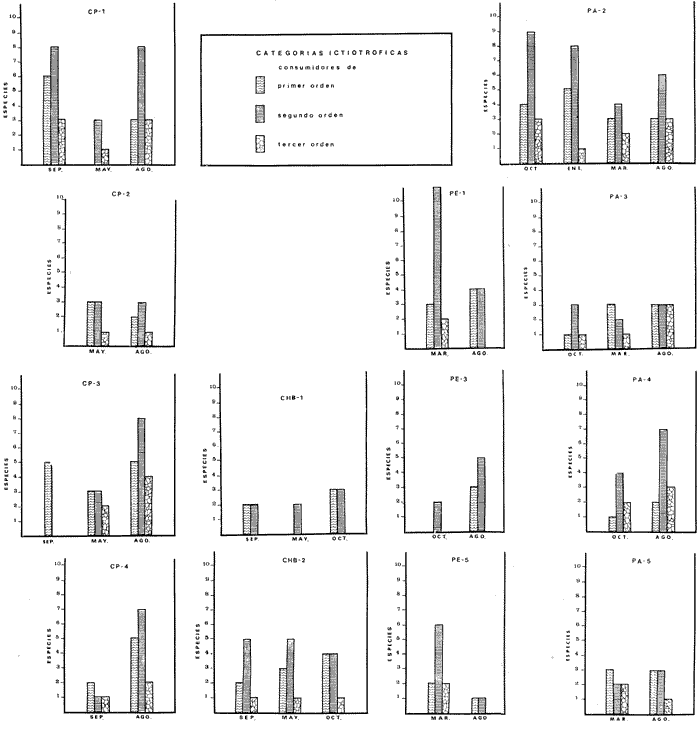 Fig.12. Variación temporal y proporciones entre las 3 categorias ictiograficos en las comunidades de peces de los 4 sistemas fluvios-lagunanes estudiados CP =Candelaria-Panlau,CHB= chumpan-Balchacah, PE=Palizada del Este y PA= Pom-Atasta .Los números asociados corresponden aestaciones de colecta. CATEGORÍAS ICTIOTRÓFICASDe acuerdo con Yáñez-Arancibia (1978 a, b y c), la ictiofauna puede quedar agrupada en 3 categorías ictiotróficas: consumidores primarios, secundarios y terciarios. De la totalidad de las especies encontradas, el 34% son consumidores primarios, el 42.5% corresponden a consumidores de segundo orden, y sólo el 23.4% son consumidores de tercer orden. Estos porcentajes sin embargo, pueden variar en un momento dado, ya que algunas especies suelen cambiar de nivel trófico de acuerdo a la disponibilidad del alimento, estación del año, edad del pez y localidad específica. Como ejemplo de esto, se puede citar al género Arius cuyas especies en ciertas épocas del año se convierten en consumidores de tercer orden, es decir se vuelven exclusivamente carnívoros. La cuantificación de las tablas 5 a 8 se basa en el registro total de especies del área correspondiente. Por lo tanto el panorama refleja valores máximos para cada área. Sin embargo los componentes comunitarios ictiotróficos varían en el tiempo de acuerdo a los patrones ecológicos en juego. Este cambio puede apreciarse en la figura 12 donde se muestran las variaciones de cada estación por área.  TABLA 5 CATEGORÍAS ICTIOTRÓFICAS DEL SISTEMA FLUVIO-LAGUNAR CANDELARIA-PANLAU  TABLA 6 CATEGORÍAS ICTIOTRÓFICAS DEL SISTEMA FLUVIO-LAGUNAR CHUMPAN-BALCHACAH  TABLA 7 CATEGORÍAS ICTIOTRÓFICAS DEL SISTEMA FLUVIO-LAGUNAR PALIZADA-DEL ESTE  TABLA 8 CATEGORIAS ICTIOTRÓFICAS DEL SISTEMA FLUVIO-LAGUNAR POM-ATASTA FRECUENCIAEl análisis de la tabla 9 ha permitido establecer los rangos de frecuencia para determinar las especies que son visitantes ocasionales, las que utilizan a los sistemas fluvio-lagunares como áreas naturales de crianza y/o para alimentarse y las especies residentes permanentes (Tablas 1 a 4). Las especies Arius melanopus, Eucinostomus gula, Micropogon furnieri y Sphoeroides testudíneus, son consideradas por frecuencia residentes permanentes de las comunidades. De las especies que utilizan estos sistemas como áreas naturales de crianza y/o para alimentarse, las que presentan una mayor frecuencia han sido: Arius felis, Bairdiella chrysura, Anchoa mitchilli mitchilli, Cetengraulis edentulus, Bagre marinus, Chaetodipterus faber y Achirus lineatus. La alta frecuencia de las especies Arius melanopus y principalmente Sphoeroides testudineus, se debe a que ambas especies se encuentran asociadas a bancos de ostión, los que constituyen el principal alimento de esta última especie quien los consume en estado adulto. Arius melanopus ataca juveniles del bivalvo, el cual se encuentra abundantemente en los sistemas fluvio-lagunares estudiados. En la tabla 9 se muestra la frecuencia de cada una de las especies por estación y época del año; en la columna final se considera el porcentaje total. DIVERSIDAD, ESPECIES DOMINANTES Y DISTRIBUCIÓNEl uso de índices para el estudio de la diversidad en comunidades nectónicas de sistemas lagunares costeros ha sido propuesto por varios autores como Harima y Mundy (1970), Copeland y Birkhead (1972) ; Hester y Copeland (1975) y Subrahmanyam y Drake (1975), entre otros. Sin embargo los resultados son difíciles de manejar e interpretar por cuanto se trata con comunidades de desplazamiento activo. Así mismo, el uso aislado de uno o más índices realmente no tiene significado, pero no es así cuando se considera información adicional y se integran con parámetros ambientales en un amplio contexto ecológico. En las figuras 13 a 16 se establece la relación de diferentes índices mismos que luego serán discutidos. En general, los índices de diversidad difícilmente rebasaron el orden de 2 unidades. De acuerdo con Shannon y Weaver (1963), esto indicaría un probable grado de contaminación. Sin embargo, estudios recientes han verificado la inexistencia de contaminación alguna en la Laguna de Términos, Botello (1978), por lo que los bajos índices de diversidad encontrados en la mayoría de las estaciones, deben obedecer a la dinámica ambiental del habitat. El índice de diversidad total anual varía en cada uno de los sistemas, así en Candelaria-Panlau corresponde 2.24; en Chumpan-Balchacah 1.28, en Palizada-del Este 1.91 y en Pom-Atasta 2.52. La diversidad calculada como índice numérico H'n, D y J' muestran la variación temporal. Estos índices permiten resumir información concerniente a la estructura numérica de las poblaciones. Su valor empírico aislado es vago y debe considerarse como información ecológica adicional. Del total de 47 especies consideradas en la tabla 10, en el Sistema Candelaria-Panlau se encuentran 33 especies (70%), en Chumpan-Balchacah 21 especies (45%), Palizada-del Este 28 especies (60%) y en el Sistema Pom-Atasta se encuentran 33 especies (70%) al igual que en Candelaria-Panlau. El incremento en el porcentaje de diversidad del Sistema Palizada del Este en relación a Chumpan-Balchacah, se puede explicar por la presencia de algunas especies de origen dulceacuícola como Ictalurus meridionalis y Cichlasoma fenestratum (Tablas 10 y 11) . En las mismas tablas 10 y 11, se pueden observar las especies dominantes, en relación al número de individuos y biomasa, así para el primer Sistema, Candelaria-Panlau se observa de la relación porcentual que Bairdiella chrysura (20%),Eucinostomus gula (19.6%), Diapterus rhombeus (17%), Arius melanopus (12.8%) y Micropogon furnieri (10.7%) constituyen las especies que apuntan con el 79.4% del total anual de componentes ícticos comunitarios. Asimismo, la dominancia en biomasa, obtenida en la tabla 11, está representada por Arius melanopus (23%), Chaetodipterus faber (17.6%), Micropogon furnieri (9.5%) y Bairdiella chrysura (8.1%) apuntando todas con el 58.2% del total anual. El Sistema Chumpan-Balchacah está dominado numéricamente por una especie, Arius melanopus ya que representa el 71.8% del total anual, y únicamente es seguida por Bairdiella ronchus con sólo el 6%, ambas especies constituyeron conjuntamente el 77.8% del total anual. La dominancia en biomasa (Tabla 11) también está ampliamente aportada por Arius melanopus con 69.7% del total, sólo seguido de Epinephelus guttatus con 8.3%, ambos hacen un total de 78% del total anual. En Palizada-del Este, la comunidad está dominada numéricamente por la misma especie que en el sistema anterior, Arius melanopus con 56%, y sólo le siguen con 8.9% Micropogon furnieri, tales especies hacen un total de 64.9%. En biomasa las especies dominantes están representadas por Arius melanopus con 52.3% y en menor grado por Cichlasoma fenestratum con 13.2% ambas hacen un total anual de 65.5 %. Por último, en el Sistema Pom-Atasta, las especies dominantes numéricamente fueron: Eucinostomus gula (31 %), Sphoeroides testudineus y Micropogon furnieri (10% cada una) ; las tres hacen un total anual de 51 %. Las especies dominantes por su biomasa en este mismo sistema fueron Chaetodipterus faber (42%) Sphoeroides testudineus (10%), Dasyatis sabina (8.7%), Arius melanopus (8.2%) y Eucinostomus gula (7%) que hacen un total de 76%. ABUNDANCIALa abundancia puede ser evaluada de tres puntos de vista: a. Densidad (ejem /m2) b. Abundancia numérica relativa (Tabla 10) c. Biomasa (g/m2) (Tabla 11). La densidad y la biomasa se analizan para cada una de las estaciones y como se puede observar en las figuras 13 a 16 no necesariamente existe una relación directa entre ambos parámetros, aunque, por lo general, valores altos de densidad pueden coincidir con los de biomasa. En general la densidad se mantiene por debajo de la unidad, mientras que la biomasa es generalmente mayor llegando hasta 14 g/m2 en la estación CHB-2 durante septiembre de 1976. La densidad máxima es de 1.27 ejem/m2. Este hecho pudiera ser explicado por la presencia de pocos ejemplares de considerable tamaño. En el Sistema Candelaria-Panlau durante el mes de septiembre de 1976, el promedio de densidad fue de 0.055 ejem /m2, mientras que para mayo de 1977 fue de 0.06, registrándose el valor más alto en agosto del mismo año con 0. 16 ejem /m2. Los valores de biomasa en las mismas fechas fueron: 4.96 g/m2, 0.87 g/m2 y 3.15 g/m2 respectivamente.Para el Sistema Chumpan-Balchacah, los promedios de densidad son: en septiembre de 1976 0.13 ejem/m2, en mayo de 1977 0.006 ejem /m2 y para octubre del mismo año 0.03 ejem /m2, mientras que la biomasa tuvo promedio para los mismos meses de 7.04 g/m2 (el valor más alto obtenido de todos los muestreos), 0.21 g/n12 y 4.418 g/n12 respectivamente. Se observa que mientras los valores de densidad son muy bajos, los de biomasa, por lo contrario, son altos. Esto se interpreta por la presencia de pocos ejemplares pero de gran tamaño, lo que indicaría que las especies de origen marino, que son la mayoría en estos sistemas, se encuentran en pleno crecimiento y no abandonan el habitat hasta avanzado estado de desarrollo o bien alcanzando su madurez gonádica. En Palizada-del Este la densidad tuvo valores promedio de 0.29 ejem /m2 para octubre de 1976, 0.034 ejem /m2 en marzo de 1977 y el menor, 0.013 ejem /m2 en agosto del mismo año. La biomasa fue del orden de 1.52 g/m2 en octubre de 1976, 0.464 en marzo de 1977 y 0.79 g/m2 en agosto del mismo año. Por último, en el Sistema Pom-Atasta, la densidad promedio fue de 0.023 ejem/ m2 en octubre de 1976, 0.025 ejem /m2 en enero de 1977, 0.006 en marzo y 0.056 ejem /m2 en agosto del mismo año. La biomasa también aquí tuvo valores más altos que la densidad; así en octubre de 1975 fue de 0.297 g/m2, en los siguientes meses de enero, marzo y agosto de 1977 fue de 0.253 g/m2, 1.92 g/m2 y 0.83 g/m2 respectivamente.
Tabla 9 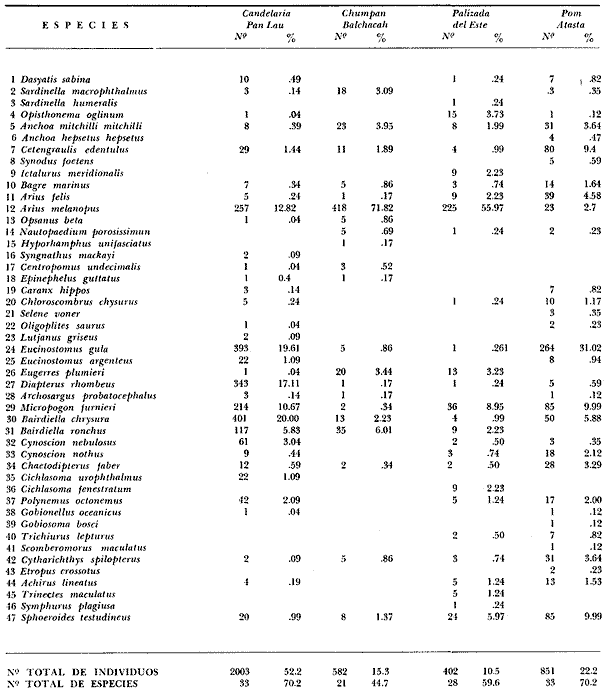 TABLA 10 DIVERSIDAD Y ABUNDANCIA NUMÉRICA RELATIVA EN LOS SISTEMAS FLUVIO-LAGUNARES ESTUDIADOS 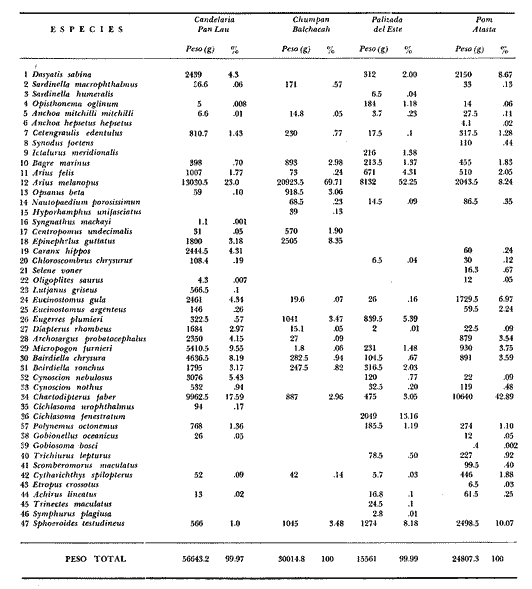 TABLA 11 DIVERSIDAD Y ABUNDANCIA DE BIOMASA EN LOS SISTEMAS FLUVIO-LAGUNARES ESTUDIADOS 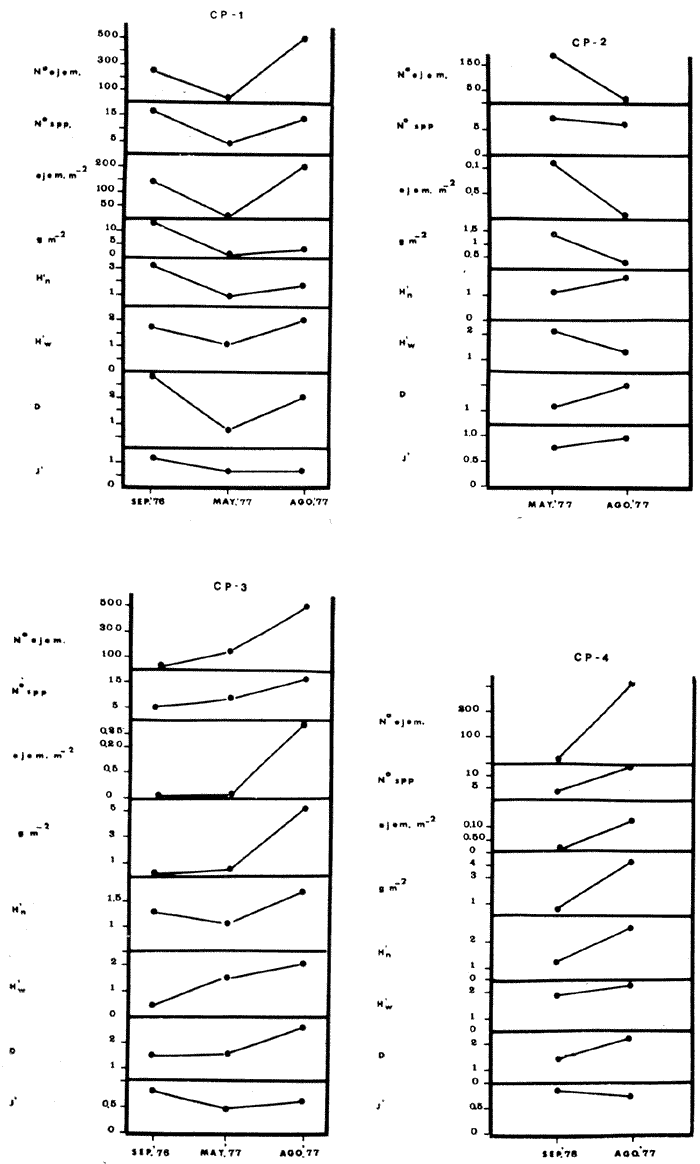 Fig. 13. Variación temporal y comparación entre la diversidad, densidad, biomasay los índices respectivos en el sistema fluvio-lagunar Candelaria-Panlau.Los números asociados corresponden a estaciones de colecta. 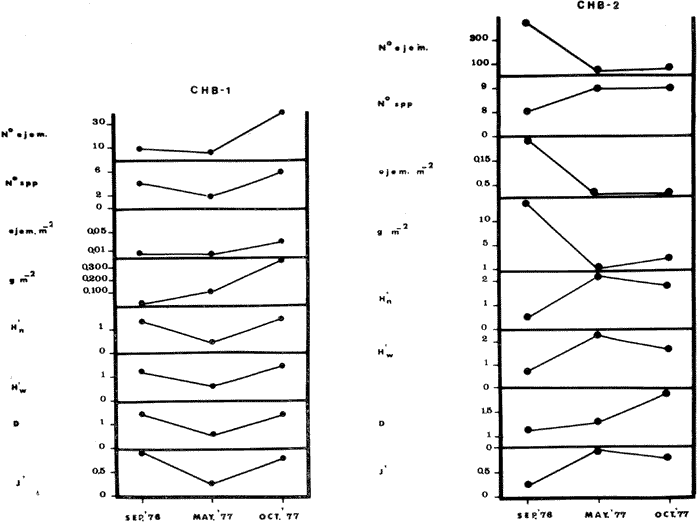 Fig. 14. Variación temporal y comparación entre la diversidad, densidad, biomasa y los índices respectivos en el sistema fluvio-lagunar Chumpan-Balchacah.Los números asociados corresponden a estaciones de colecta. 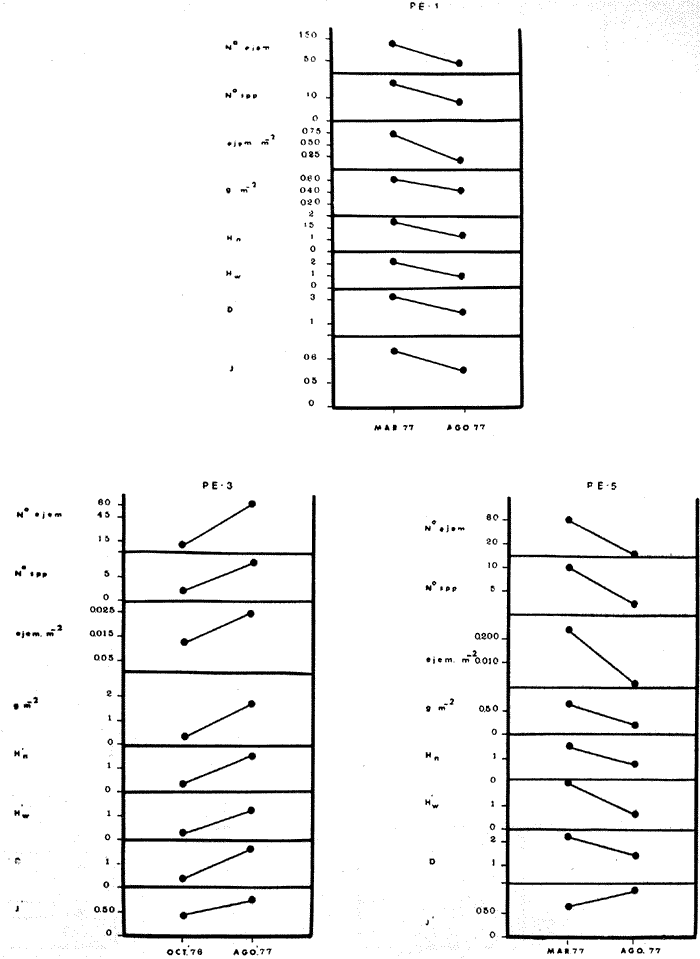 Fig. 15. Variación temporal y comparación entre la diversidad, densidad, biomasa y los índices respectivos en el sistema fluvio-lagunar Palizada-del Este.Los números asociados corresponden a estaciones de colecta. 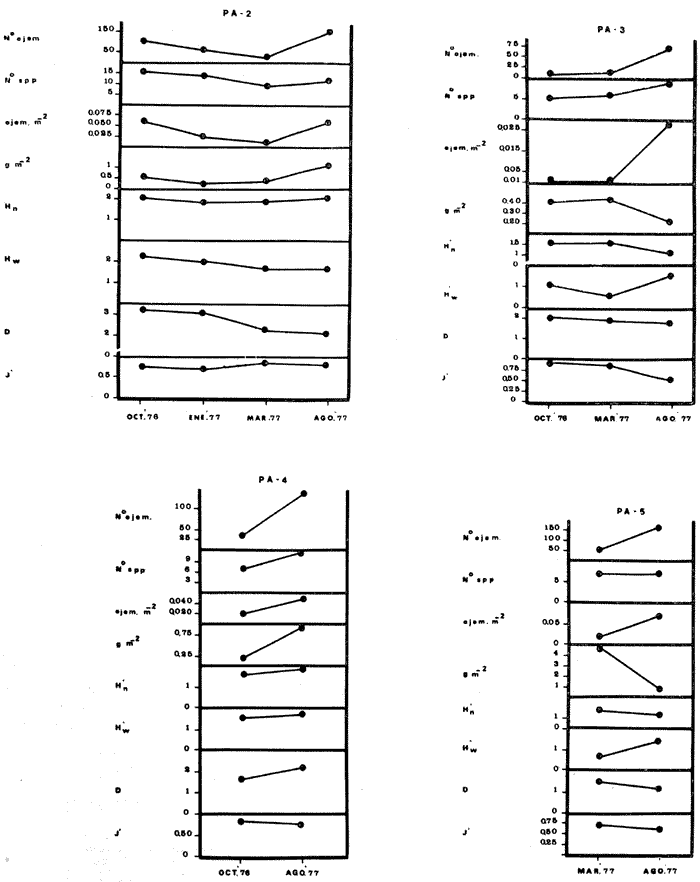 Fig. 16. Variación temporal y comparación entre la diversidad, densidad, biomasa y los índices respectivos en el sistema fluvio-lagunar Pom-Atasta. Los números asociados corresponden a estaciones de colecta. AFINIDADSe puede observar que entre los 4 sistemas fluvio-lagunares estudiados existe un alto grado de afinidad en relación a las especies comunes. Esta afinidad se manifiesta principalmente entre el Sistema Candelaria-Panlau y Pom-Atasta, los cuales tienen entre sí 25 especies comunes de un total de 33, lo que corresponde a una afinidad de 76%. Este alto porcentaje de afinidad puede deberse a ciertas características ecológicas comunes entre ambos sistemas, como son la presencia de considerables bancos de ostión y la relativa cercanía de cada una de las bocas con que cuenta Laguna de Términos, lo que proporciona notable influencia marina que se manifiesta en una mayor diversidad íctica. Esta observación refuerza la similitud ecológica de ambos sistemas. Por otra parte, los Sistemas Chumpan-Balchacah y Palizada-del Este también presentan condiciones similares entre sí, ya que la afinidad entre ambos es de 62% con 15 especies comunes de un total de 28. La afinidad entre los Sistemas Candelaria-Panlau con Chumpan-Balchacah y Palizada-del Este, varía de 75 a 70% respectivamente; este mismo porcentaje de afinidad (70%) se registra entre Palizada-del Este y Pom-Atasta, ambos con 25 especies en común. Los sistemas menos afines entre sí fueron Chumpan-Balchacah y Pom-Atasta, cuya afinidad fue de 58% considerando 15 especies en común de un total de 33. Sin embargo, este valor es considerable a pesar de ser el menor registrado, lo que indica que entre los 4 sistemas fluvio-lagunares existe una marcada afinidad ecológica. El mismo porcentaje 25.5% (12 especies) se presentó para especies existentes en 3 de los sistemas estudiados, estas son: Anchoa mitchilli mitchilli Cetengraulis edentulus Bagre marinus Arius felis Arius melanopus Eucinostomus gula Diapterus rhombeus Micropogon furnieri Bairdiella chrysura Chaetodipterus faber Cytharichthys spilopterus Sphoeroides testudineus Analizando las especies presentes, se pudo establecer que el 25.5% (12 especies) estuvieron presentes en los 4 sistemas; estas especies son: Dasyatis sabina Sardinella macrophthalmus Opisthonema oglinum Nautopaedium porosissimum Chloroscombrus chrysurus Eugerres plumieri Archosargus probatocephalus Bairdiella ronchus Cynoscion nebulosus Cynoscion nothus Polynemus octonemus Achirus lineatus Esto determina que 24 especies (51%) están variadamente distribuidas en los sistemas fluvio-lagunares estudiados, siendo esto otro aspecto importante que refuerza la similitud entre los 4 sistemas. DISCUSIÓN
EL HABITATLas características ambientales predominantes, analizadas en la descripción de las áreas, determinan dos hechos significativos: abundantes bosques de mangle rojo, Rhizophora mangle y, grandes e importantes arrecifes de ostión, Crassostrea virginica. Bosques de manglarLos bosques de manglares son ciertamente los productores primarios dominantes en los pantanos asociados, aún cuando existen algas filamentosas, pastos de pantano, lirio acuático y otros. No obstante la producción de materia orgánica proveniente del mangle, es considerablemente mayor y aparentemente la más significativa en las asociaciones ecológicas lagunares y estuarinas tropicales y subtropicales donde se presentan (ver: Heald, 1970; Odum, 1971; Odum y Heald, 1972; Odum et al., 1973; Heald et al., 1974; Odum y Heald, 1975; Yáñez-Arancibia, 1978 a, b y c; Edwards, 1973 a y b, 1978). La biota asociada a Rhizophora mangle puede ser muy variada, desde numerosas epífitas como algas unicelulares, algas filamentosas verdes y verdeazules y macroalgas, que pueden distribuirse de acuerdo a sus características ecológicas, desde raíces aéreas hasta el interior de los sedimentos (Kuenzler, 1974). Numerosos tipos de animales se encuentran en estos pantanos de manglar, siendo los más importantes organismos bentónicos, probablemente los crustáceos y moluscos donde la mayoría pueden clasificarse como comedores de fondo o filtradores. Especies de las familias Ocypodidae, Grapsidae, Gecarcinidae, Pinnotheridae, Penaeidae, Portunidae, Palemonidae y Balanidae son comunes entre los crustáceos del habitat. Moluscos gastrópodos y bivalvos de los géneros Cerithium, Littorina, Brachydontes, Crassostrea y otros son comunes en el área. La mayoría de estos animales no obtienen su alimento directamente del manglar, sino que filtran partículas del agua, fragmentos y sedimentos para obtener la materia orgánica. Sin embargo, los árboles de manglar en los pantanos sirven a las poblaciones animales más como un lugar protegido donde vivir que como fuente de alimento. Kuenzler (1974), ha discutido que la distribución de los animales en los bosques de manglares está controlada por: 1) nivel del agua, 2) resistencia a la pérdida de agua, 3) protección de insolación, 4) grado de consolidación del substrato y, 5) la disponibilidad en las capas superiores de un substrato de microflora y microfauna y detritus orgánico disponibles como alimento. En Laguna de Términos podría agregarse a esto la fuerza del flujo de la corriente y la turbidez del agua como factores probablemente importantes que determinan la mayor abundancia de filtradores o de comedores de fondo. Arrecifes de ostiónLos arrecifes de ostión, Crassostrea virginica, resultan de la colonización de ostiones juveniles sobre cualquier substrato adecuado. Este arrecife va creciendo subsecuentemente conforme nuevos ostiones colonizan el substrato viejo de ostiones muertos (Chestnut, 1974). En la boca de los 4 sistemas fluvio-lagunares estudiados, es característico la comunidad béntica de arrecifes de ostión. Un examen de los arrecifes de ostión en diferentes latitudes, muestran una gran abundancia de formas animales asociados con las ostras; desde micro, medio y macrofauna de vida libre, como también los mismos grupos sésiles utilizando las conchas como substrato sólido. Estos últimos incluyen numerosas formas de algas, hidrozoos, platelmintos, nemertinos, briozoos, cirripedios, moluscos, poliquetos tubícolas y de vida libre, y tunicados, todos estos como parte de la epifauna. Por otra parte pueden encontrarse formas que horadan las conchas, como esponjas, moluscos, algas y poliquetos. Diversos organismos buscan su alimento en los intersticios del arrecife, pero otros predan sobre ellos como algunos caracoles, cangrejos y peces. Importantes grupos faunísticos circundan el arrecife, entre estos destacan los peces (discutidos más adelante) y diferentes tipos de decápodos de los géneros Penaeus, Callinectes, Palemonetes, Rhitropanopeus y representantes de otras familias como Pinnotheridae, Porcellanidae y Majidae. El tipo y número de especies en la comunidad del arrecife de ostión, debe depender básicamente del medio ambiente, salinidad, temperatura, corrientes, turbidez y otros factores limitantes de la distribución y el crecimiento. Puesto que Crassostrea virginica es una especie sedentaria, no es capaz de escapar de sus predadores ni de cambios drásticos en condiciones naturales. Sin embargo, los cambios o variaciones temporales pueden ser tolerados por mecanismos de de defensa: físicos (cierre de la concha) y fisiológicos (osmorregulación). En Laguna de Términos excesiva precipitación y aumento del flujo pluvial pueden provocar la total mortalidad de algunos arrecifes. Con subsecuente incremento en salinidad, las larvas pueden nuevamente colonizar y formar una nueva comunidad. Parámetros ambientalesLa turbulencia causada principalmente por las corrientes de los ríos de los sistemas fluvio-lagunares estudiados, además de la acción de los vientos, determina que las aguas presenten valores de oxígeno disuelto muy por encima de los valores letales para peces en sistemas de aguas salobres (i.e., 1.8 ml O2 / l; Muus, 1967), no obstante la considerable cantidad de materia orgánica aportada a los sistemas por los productores primarios especialmente por hojas de Rhizophora mangle. En el sistema Candelaria-Panlau (Tabla 12, Fig. 17), el más bajo valor de O2 se registra a fines del periodo de secas y comienzos del periodo de lluvias, y el máximo valor de oxígeno ocurre precisamente en pleno periodo de lluvias. La temperatura muestra una gran estabilidad anual con un rango de 28 a 30ºC a fines del periodo de secas y ligeramente más alta en la época de lluvias, con rango de 29 a 31ºC Sin embargo, la salinidad varía considerablemente desde la zona de la boca (25 a 26%) hacia el interior fluvial (5 a 8%). En el sistema Chumpan-Balchacah (Tabla 13, Fig. 18) , el oxígeno fue superior a los valores letales para peces, aún cuando sólo hay registro para la época de secas donde el consumo debe ser aparentemente mayor. La temperatura muestra, al igual que en el sistema anterior, una estabilidad variando entre 28 a 31ºC, pero con un registro extremo de 35ºC en el mes de septiembre de 1976. La salinidad se mantiene alta durante la época de secas con un rango de 26 a 28%o y varía significativamente durante la época de lluvias con un rango de 10 a 22%o. En Palizada-del Este (Tabla 14, Fig. 19), el oxígeno se mantiene alto durante todo el año, con un amplio rango de 2.8 a 6.8 ml/l en la época de secas. La temperatura muestra alguna variación anual con un mínimo de 24ºC durante la época de secas y un máximo de 31ºC en la época de lluvias. La salinidad es baja y se ha registrado un máximo de 14%o en la zona de la boca. Este sistema es predominantemente dulceacuícola. En el Sistema Pom-Atasta (Tabla 15, Fig. 20), el oxígeno se mantiene alto todo el año. La temperatura muestra los valores más bajos en invierno durante la época de secas, variando entre 20 a 26ºC, y los más altos en la época de lluvias de 28 a 32ºC. La salinidad se comporta muy similar al Sistema Candelaria-Panlau con mínimas de 2%o hacia el interior y 20%o en la zona de la boca; el rango es amplio. En términos generales, los cuatro sistemas estudiados son muy similares, con salinidad en un gradiente desde 2 a 26%o; relativa estabilidad térmica; buena oxigenación de las aguas y alta turbidez como se aprecia en la tabla 16, donde se observa una mayor similitud nuevamente entre los Sistemas Candelaria-Panlau y Pom-Atasta, así como también entre Chumpan-Balchacah y Palizada-del Este. Esto se refuerza al analizar las tablas 12 a 15. El aporte de agua dulce a través de estos sistemas hacia la Laguna de Términos es elevado y Phleger y Ayala-Castañares (1971), estiman un volumen de aproximadamente 6 x 109 m3/año. 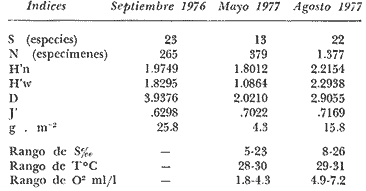 TABLA 12 ANÁLISIS DE LA VARIACIÓN DE LOS FACTORES AMBIENTALES Y SUS EFECTOS SOBRE LA COMPOSICIÓN, DIVERSIDAD Y ABUNDANCIA DE LAS COMUNIDADES DE PECES, EN COLECTAS DE "TRAWL" EN EL SISTEMA FLUVIO-LAGUNAR CANDELARIA-PANLAU 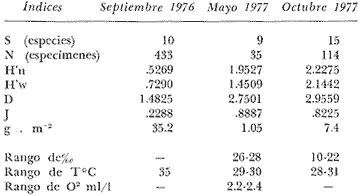 TABLA 13 ANÁLISIS DE LA VARIACIÓN DE LOS FACTORES AMBIENTALES Y SUS EFECTOS SOBRE LA COMPOSICIÓN, DIVERSIDAD Y ABUNDANCIA DE LAS COMUNIDADES DE PECES, EN COLECTAS DE "TRAWL" EN EL SISTEMA FLUVIO-LAGUNAR CHUMPAN-BALCHACAH 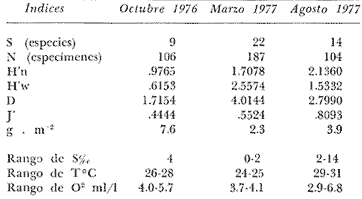 TABLA 14 ANÁLISIS DE LA VARIACIÓN DE LOS FACTORES AMBIENTALES Y SUS EFECTOS SOBRE LA COMPOSICIÓN, DIVERSIDAD Y ABUNDANCIA DE LAS COMUNIDADES DE PECES, EN COLECTAS DE "TRAWL" EN EL SISTEMA FLUVIO-LAGUNAR PALIZADA-DEL ESTE 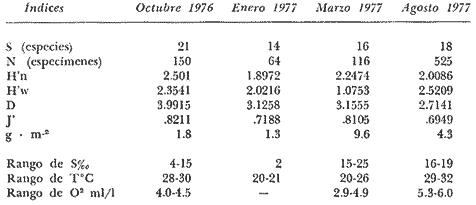 TABLA 15 ANÁLISIS DE LA VARIACIÓN DE LOS FACTORES AMBIENTALES Y SUS EFECTOS SOBRE LA COMPOSICIÓN, DIVERSIDAD Y ABUNDANCIA DE LAS COMUNIDADES DE PECES, EN COLECTAS DE "TRAWL" EN EL SISTEMA FLUVIO-LAGUNAR POM-ATASTA  TABLA 16 PORCENTAJE DE TRANSPARENCIA ANUAL EN LOS SISTEMAS FLUVIO-LAGUNARES EL MUESTREOLas poblaciones nectónicas de activo desplazamiento no son difíciles de colectar, pero sí puede ser problemática la interpretación de la colecta. Se considera que sólo desde el año de 1972 se han venido desarrollando sistemáticamente métodos cuantitativos para evaluar poblaciones de necton en lagunas costeras y estuarios. Dependiendo del área de estudio, de la localidad, de la dinámica del ambiente, de la profundidad de la turbidez y de la topografía del fondo entre los factores más importantes, es que se han utilizado: 1) atarraya, 2) chinchorro, 3) agallera, 4) red de arrastre ("trawl") y 5) red estacionaria ("drop net") (Bechtel y Copeland, 1970; Loesh et al., 1972; Kushland, 1974; Hester y Copeland, 1975; Kjelson et al., 1975; Kjelson, 1977; Kjelson y Colby, 1977; Kjelson y Johnson, 1974 y 1978; Yáñez-Arancibia, 1978 a, b y c; Bravo Núñez y Yáñez-Arancibia., 1979), además de otros importantes trabajos considerados en las tablas 17 y 18. El análisis de esta literatura, permite establecer que en la actualidad son 2 los sistemas de colecta preferidos por los autores: red de arrastre y red estacionaria. En Laguna de Términos ambos métodos parecen ser los más adecuados. Se considera que la red estacionaria (drop-net) entrega mayor información, pero su desarrollo es muy reciente y aún está poco difundida entre los investigadores de poblaciones nectónicas de sistemas estuarinos. No obstante, la tendencia es a utilizarse como una red standard en el futuro. A su vez, la red de arrastre de prueba camaronera se encuentra ampliamente utilizada en la literatura. Al respecto, existen numerosas pruebas de eficiencia de captura, considerándose que un arrastre a la velocidad entre 2 y 3 nudos durante 10 a 30 minutos de tiempo promedio, la eficiencia de captura promedio para las poblaciones de peces en general es de 20%. Este último criterio se ha adoptado en estos estudios de Laguna de Términos. ESTRUCTURA DE LAS COMUNIDADESFueron encontradas las 3 categorías ictiotróficas que se han propuesto para lagunas costeras; esto es: consumidores de primer orden, consumidores de segundo orden y consumidores de tercer orden. De éstos, las especies más abundantes numéricamente correspondieron a consumidores de segundo orden (21 especies, 44.7% con una biomasa total anual de las 4 localidades de 4.95 g/m2/año) . A su vez los consumidores de primer orden representaron 16 especies, 34.0%, con una biomasa total anual de las 4 localidades de 2.51 g/m2/año. Los consumidores de tercer orden representaron 10 especies, 21.0%, con una biomasa total anual de las 4 localidades de 1.00 g/m2/año. Los consumidores de segundo orden se identifican en gran medida con los componentes comunitarios que utilizan los sistemas fluvio-lagunares como áreas naturales de crianza y/o para alimentarse (más del 50%) por lo cual es razonable suponer que estos componentes tanto estructural como funcionalmente son los que en forma más significativa regularían y condicionarían en tiempo y espacio a las comunidades en cuestión. En términos ecológicos Yáñez-Arancibia y Nungent (1977) pueden establecer que los sistemas fluvio-lagunares estudiados, por la dinámica de sus comunidades ícticas, son exportadores de energía (productividad pesquera) hacia la propia Laguna de Términos y subsecuentemente a la plataforma continental adyacente. Los índices de diversidad, tanto en número de especies (H'n) como de biomasa (H'w) en general no son altos y concuerdan con valores de áreas de características similares donde predominan pastos marinos, pastos de pantanos y manglares en centro y norte América (Tablas 17 y 18) . Sin embargo, se registraron algunos valores superiores a las 2 unidades tanto en diversidad numérica como en biomasa, sobre todo en los sistemas Candelaria-Panlau y Pom-Atasta donde alcanzaron valores para el índice de diversidad H'n ligeramente superiores a 3 unidades en algunas estaciones. El hecho de que por lo general se registran índices bajos en la mayoría de las estaciones dentro de los sistemas fluvio-lagunares, no reflejan necesariamente que se trate de áreas contaminadas como ha sido propuesto, ya que estudios recientes del área por Botello (1978) y Botello y Mandelli (1978) han demostrado la ausencia de niveles drásticos de agentes contaminantes. Por lo tanto, es razonable suponer que las presiones sobre la comunidad son naturales por la dinámica de un ambiente en modificación constante en su hidrología, sedimentación, disponibilidad de alimento, etcétera, a través de un ciclo anual. Esto representa una fuerte presión sobre las poblaciones de las especies que se encuentran colonizando estos habitats. 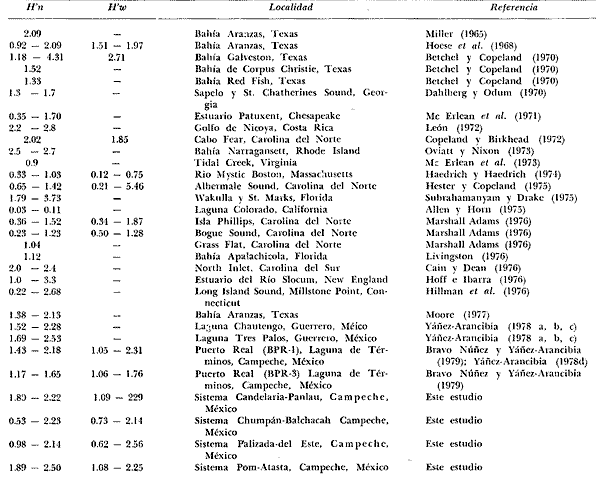 TABLA 17 VALORES DE DIVERSIDAD (H'n - Número; H'w = Peso) DE COMUNIDADES DE PECES EN SISTEMAS ESTUARINOS DEL CENTRO Y NORTE AMÉRICA Del análisis de la tabla 18, donde se compara la biomasa de comunidades de peces de diferentes sistemas ecológicos estuarinos con pastos y manglares, se deduce que, aparentemente, estos habitats son sistemas ecológicos eficientes capaces de convertir la energía radiante en materia orgánica suficiente para sustentar importantes poblaciones de peces. Sin embargo, ¿es realmente alta la biomasa de los peces? En realidad diferentes estudios como los de la tabla 18 indican que ésta es considerablemente menor que el potencial de productividad primaria de las lagunas. No obstante, algunas excepciones a esto. se presentan en algunas lagunas costeras tropicales con bocas efímeras donde la productividad pesquera puede ser muy alta en un periodo breve de tiempo (Yáñez-Arancibia, 1978 c; tabla 18). Además existen otros valores altos de biomasa en peces, en lagunas tropicales, que pueden complementar a la tabla 18 y ejemplificar lo recién arriba discutido: Lagler et al. (1971) quienes encuentran entre 33.7 y 59.2 g/m2 en la laguna Kafue Flats, Africa. Mago (1970) encuentra 100 g/m2 en una laguna de Venezuela. Bonetto et al.(1970) quienes encuentran entre 55 y 128.7 g/m2 en cuatro lagunas permanentes del Paraná. Hilcík (1970) encuentra 32.1 g/m2 en la Laguna Sabanilla y 32.5 en la Laguna Luisa, ambas en Cuba. Detallada discusión de esto y otros aspectos de la comunidad de peces tropicales hace Lowe-Mc Connell (1975) con énfasis en aguas dulces. 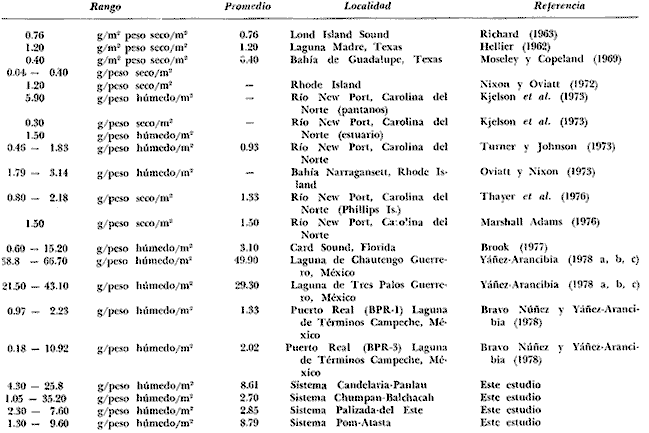 TABLA 18 VALORES DE BIOMASA DE COMUNIDADES DE PECES EN SISTEMAS ESTUARINOS DE NORTE AMÉRICA Por los resultados de este estudio, se puede agregar la importancia de la comunidad de arrecife de ostión, donde Dame (1975) ha discutido el significado del flujo energético en comunidades de estos arrecifes y la trascendencia de ello hacia los macroconsumidores. La biomasa de los peces aquí encontrados depende directamente de las características ecológicas del habitat. Resulta relevante el hecho de que Arius melanopus se encuentre presente en cifras significativas en los 4 sistemas fluvio-lagunares. Esto determina que esta especie sea la más común y conspicua dentro de los residentes permanentes con una frecuencia de 85%. Con cifras menos significativas en abundancia, pero con un porcentaje de frecuencia de 92% en las colectas, la especie Sphoeroides testudineus es otro residente permanente que en general se encuentra asociado a bancos de ostión. Esta especie preda sobre Crassostrea virginica. Las otras dos especies residentes permanentes son: Eucinostomus gula y Micropogon furnieri, presentaron 77% de frecuencia anual. La categorización aquí propuesta, esto es, residentes permanentes, visitantes ocasionales y los que utilizan los sistemas como áreas naturales de crianza o para alimentarse, es muy similar a la sugerida por Subrahmanyan y Drake (1975) en la cual reconocen cuatro categorías para comunidades de peces en ecosistemas parecidos: especies residentes permanentes, especies que utilizan los pantanos salobres como áreas naturales de crianza, especies que utilizan los pantanos salobres para alimentarse y especies esporádicas (visitantes ocasionales) , que presentan movimientos erráticos. Con la diferencia de que en este estudio, se fusionan en una sola categoría las especies que utilizan las áreas para crianza y las que la utilizan para alimentarse, ya que algunas de estas especies presentaron ambas características y tales funciones corresponden a un comportamiento ecológico similar. Yáñez-Arancibia (1977 y 1978 a, b y c) propone seis componentes comunitarios, cuatro de ellos coinciden con Subrahmanyan y Drake (1975), y el resto son los denominados dulceacuícolas y en tránsito. En este estudio, dentro de los visitantes ocasionales se incluyen tanto dulceacuícolas como de origen marino; y especies en tránsito (anádromos y catádromos) no fueron encontrados. A su vez Bravo-Núñez y Yáñez-Arancibia (1979), reconocieron para la Boca de Puerto Real (un subsistema inestable y transicional de la Laguna de Términos) especies ocasionales, especies en tránsito, y residentes permanentes. Esa categorización guarda cierta similitud con la de Subrahmanyan y Drake (1975), pero son esencialmente diferentes. La categorización de Puerto Real parece más afín con la propuesta por Kikuchi (1966) en la cual reconoce para comunidades de pastos marinos: residentes permanentes, residentes estacionales, visitantes temporales que se alimentan en una área más amplia que la solamente ocupada por pastos marinos y especies migratorias ocasionales. Es probable que todas estas categorizaciones de componentes comunitarios tengan similitudes y diferencias, básicamente por ser ecosistemas de aguas salobres pero de diferentes latitudes. Todas tienen una validez local; sin embargo, la amplitud de la efectuada por Yáñez-Arancibia (varios trabajos) permite englobar a las otras. Para ubicar a las diferentes especies en cualquiera de las categorías de componentes comunitarios aquí propuestas se considera, además del análisis de frecuencia ya mencionado, los patrones de migración, hábitos alimenticios, tallas y peso de los ejemplares y el estado de sus gónadas. Sobre esto último es fundamental destacar que 19 especies se encuentran en la Boca de Puerto Real y en los sistemas fluvio-lagunares, pero las tallas de los ejemplares de estas poblaciones son considerablemente mayores en estos últimos habitats, corroborándose la apreciación de Bravo-Núñez y Yáñez-Arancibia (1978) de que las poblaciones de peces utilizan la Boca de Puerto Real como una de las entradas del sistema ecológico del interior de Laguna de Términos. Además, los resultados de este estudio confirman que los sistemas fluvio-lagunares son áreas de crianza y fuente alimenticia para numerosas especies, algunas de las cuales penetraron a través de la Boca de Puerto Real. Las 19 especies comunes para la Boca de Puerto Real (50 especies en total) y para los sistemas fluvio-lagunares estudiados (47 especies en total) determina que exista una afinidad ecológica de 16% entre ambos subsistemas. Esta afinidad es muy baja como reflejo de la dinámica ambiental que representan, observándose que dentro del complejo ecológico de Laguna de Términos existen diferentes comunidades reflejo de los diferentes habitat existentes. Las poblaciones de especies comunes tienen una gran significancia en la transferencia de flujo energético entre diferentes habitat. El planteamiento del comportamiento similar de los 4 sistemas fluvio-lagunares estudiados se fundamenta en el análisis de afinidad de sus poblaciones de peces, mostrando éstas una notoria similitud. En las figuras 17 a 20 se relacionan individualmente para cada localidad los parámetros ambientales con algunos datos de la estructura numérica de las poblaciones donde se aprecia que el incremento de la diversidad y de la abundancia relativa se encuentra en relación directa con la finalización del periodo de lluvias, esto a fines de verano y comienzos de otoño con un considerable reclutamiento de especies juveniles al interior de los sistemas. 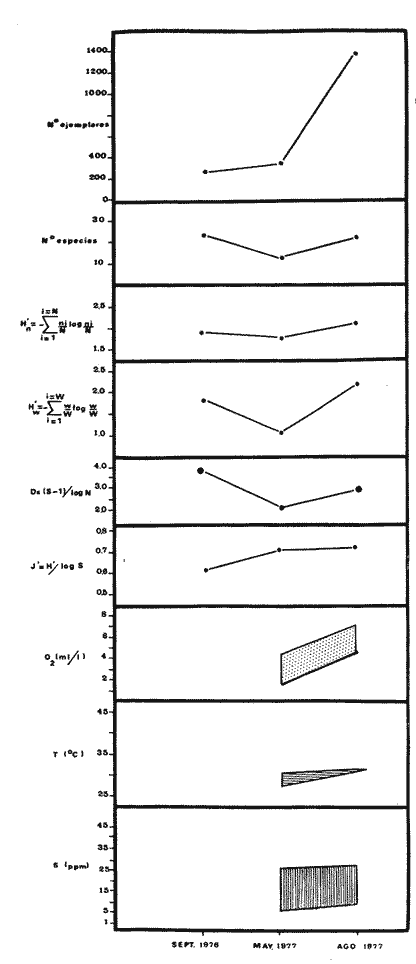 Fig. 17. Variación temporal y comparación entre la diversidad, abundancia relativa, sus respectivos índices y su posible relación con los parámetros ambientales; en el sistema fluvio-lagunar Candelaria- Panlau. 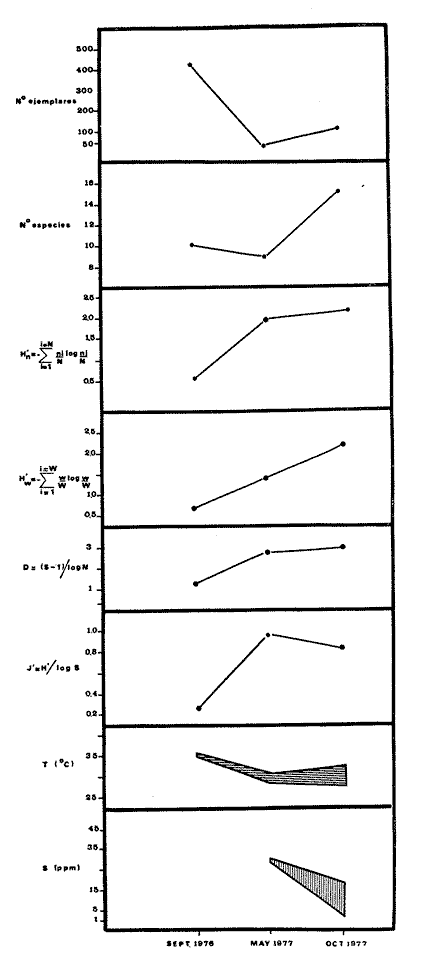 Fig. 18. Variación temporal y comparación entre la diversidad, abundancia relativa, sus respectivos índices y su posible relación con los parámetros ambientales; en el sistema fluvio-lagunar Chumpan-Balchacah. 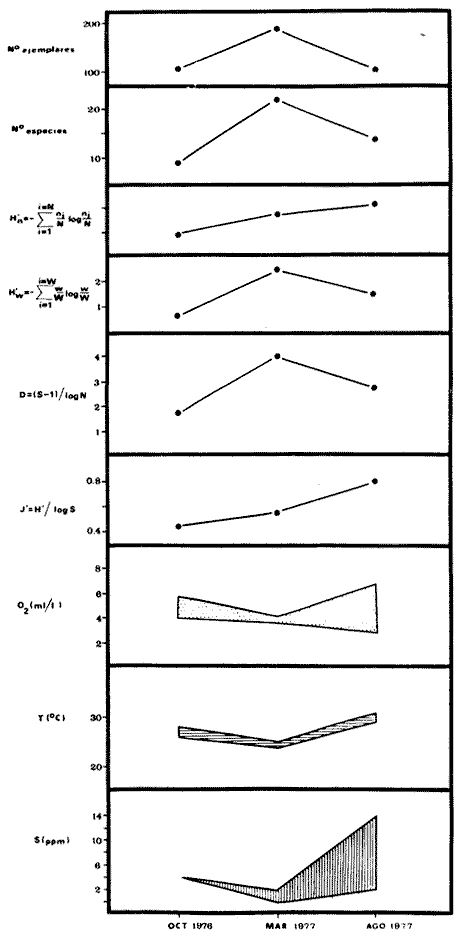 Fig. 19. Variación temporal y comparación entre la diversidad, abundancia relativa, sus respectivos índices y su posible relación con los parámetros ambientales; en el sistema fluvio-lagunar Palizada-del Este. 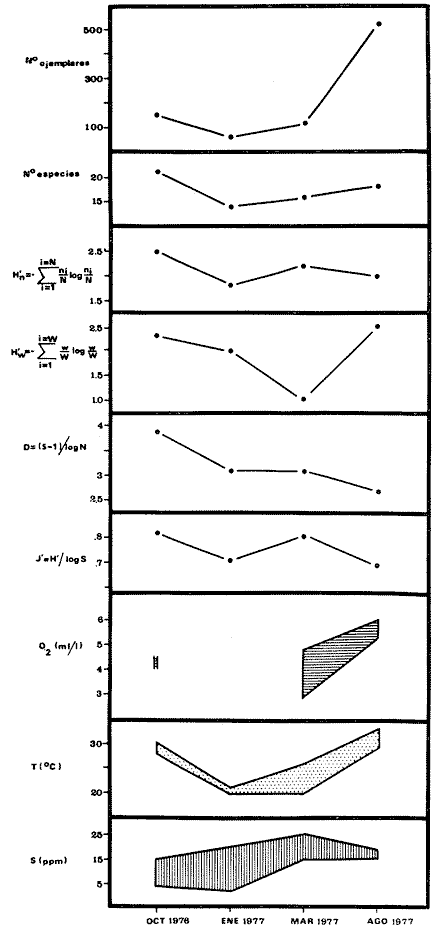 Fig. 20. Variación temporal y comparación entre la diversidad, abundancia relativa, sus respectivos índices, y su posible relación con los parámetros ambientales; en el sistema fluvio-lagunar Pom-Atasta. 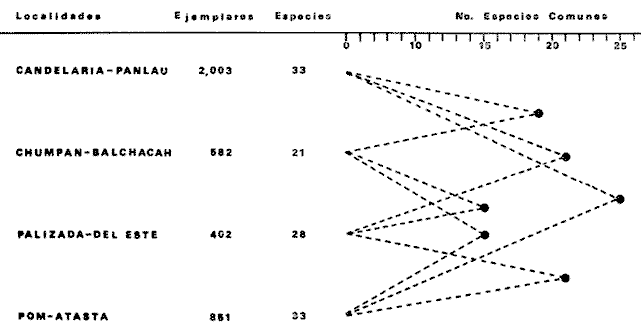 Fig. 21. Relación de la diversidad, abundancia numérica relativa y número de especies comunes entre los 4 sistemas fluvio-lagunares estudiados. 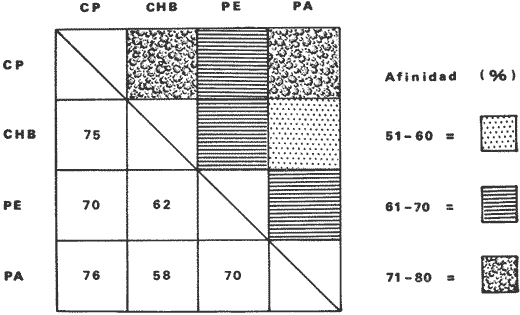 Fig. 22. Diagrama de enrejado de la relación porcentual de afinidad ecológica, entre los 4 sistemas fluvio-lagunares estudiados. PC = Candelaria-Panlau, CHB = Chumpán-Balchacah, PE = Palizada-del Este y PA = Pom-Atasta. 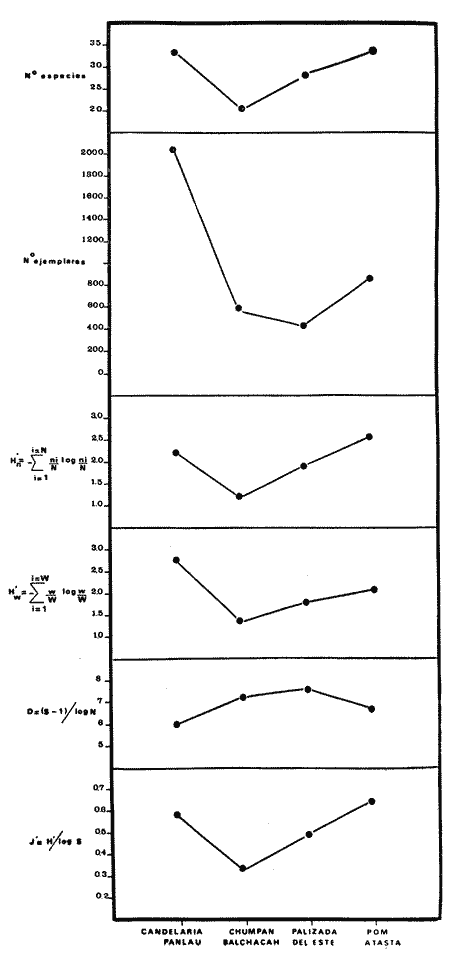 Fig. 23. Comparación de la diversidad, abundancia relativa y sus respectivos índices entre los 4 sistemas fluvio-lagunares estudiados. La mayor afinidad se presenta entre Candelaria- Panlau y Pom-Atasta, así como también entre Chumpán-Balchacah y Palizada-del Este. Ver figura 21 y 22 y discusión en el texto. Los valores de H'n son bajos y reflejan una distribución de pocas especies contribuyendo con numerosos organismos a las características estuarinas del ambiente. Rápidas fluctuaciones de la salinidad, disponibilidad de alimento y turbidez limitan a las especies de peces capaces de vivir dentro de los sistemas fluvio-lagunares. La temperatura puede ser también un factor importante. El análisis de las figuras 17 a 20 sugiere que poblaciones de numerosos individuos colonizan estos habitats a fines del periodo de lluvias, misma época en que se produce un reemplazo de poblaciones casi adultas como lo sugieren los valores de la época de secas. Puesto que existen dos épocas climáticas marcadas, es explicable que poblaciones de diferentes especies utilizan estos estuarios durante diferente época del año y por diferentes razones. Del total de 47 especies encontradas, la dispersión es variada pero similar; esto se observa en la figura 21 donde se analizan el número de especies comunes entre las diferentes localidades. El análisis de la afinidad muestra un alto grado de significancia estadística (Fig. 22) entre los 4 sistemas fluvio-lagunares donde se aprecia una mayor similitud entre las áreas más distantes; así como también entre las dos áreas intermedias. La concentración de información de la estructura numérica de las poblaciones (Fig. 23) resalta esta observación, donde Candelaria-Panlau y Pom-Atasta muestran gran similitud ecológica e igual cosa sucede entre Chumpan-Balchacah y Palizada-del Este. La mayor diversidad se presenta en Candelaria-Panlau y Pom-Atasta donde también son varias las especies dominantes. Parece razonable una relación directa de estas situaciones entre ambos sistemas por el hecho de que ambos sistemas fluvio-lagunares se encuentran próximos a las bocas de Laguna de Términos y presentan además los más importantes arrecifes de ostiones, particularmente en Pom-Atasta. El índice de diversidad total anual (H'n) indica 2.24 para Candelaria-Panlau, 1.29 para Chumpan-Balchacah, 1.91 en Palizada-del Este y 2.52 para Pom-Atasta, tales valores varían dentro de los rangos indicados en la tabla 17. Esta misma tabla muestra los rangos de variación del índice de biomasa (H'w) y la tabla 18 señala el rango y promedio de biomasa (g/peso húmedo /m2) para las localidades presentándose 8.61 en Candelaria-Panlau, 2.70 en Chumpan-Balchacah, en Palizada-del Este 2.85 y 8.79 en Pom-Atasta. Cuatro especies (8.5%) son residentes permanentes en los sistemas estudiados: Arius melanopus, Eucinostomus gula, Micropogon furnieri y Sphoeroides testudineus. Quince especies (32%) son típicas de estos sistemas ecológicos estudiados, por biomasa, número y frecuencia en los muestreos: Bairdiella chrysura Eucinostomus gula Bairdiella ronchus Arius melanopus Micropogon furnieri Sphoeroides testudineus Chaetodipterus faber Epinephelus guttatus Cichlasoma fenestratum Dasyatis sabina Arius felis Anchoa mitchilli mitchilli Cetengraulis edentulus Bagre marinus Achirus lineatus Además de estas 15 especies que caracterizan a las comunidades de peces estudiados, existen 24 especies (51%) variadamente distribuidas en todos los sistemas fluvio-lagunares estudiados (ver resultados de afinidad) , siendo esto otro aspecto importante que refuerza la similitud ecológica entre los cuatro sistemas ecológicos estudiados. ConclusionesLas comunidades de peces de los 4 sistemas fluvio-lagunares son reflejo del habitat y los patrones ecológicos característicos; esto es: a) gradiente de salinidad, b) temperatura c) sustrato d) transparencia, e) vegetación, f) macrofauna béntica y g) disponibilidad de alimento. 1. El habitat se caracteriza por presentar un gradiente salino de 2 a 26%o, temperaturas de 20 a 28°C (excepcionalmente hasta 30°C) durante la época de secas y de 26 a 32°C (excepcionalmente hasta 35ºC) durante la época de lluvias, substrato de fango limo-arcilloso con menos de 25% de CaCO3, transparencia baja entre 20 y 40%, vegetación predominante de Rhizophora mangle, macrofauna béntica típica de arrecifes de Crassostrea virginica. En conjunto, los sistemas fluviales aportan una descarga mínima anual dentro de la Laguna de Términos de 6 x 109 m3 /año. 2. Fueron encontradas 47 especies de peces repartidas entre 26 familias y 40 géneros. Las familias mejor representadas en diversidad fueron: Sciaenidae (5 especies) , Carangidae y Gerridae (4), Clupeidae, Engraulidae y Ariidae (3). 3. Estas 47 especies se componen, desde el punto de vista de su permanencia en la comunidad en: especies residentes permanentes 4 (8.5%), especies que utilizan los sistemas como áreas naturales de crianza y/o para alimentarse 17 (36.2%) y visitantes ocasionales 25 (55.3%). 4. La diversidad ictiofaunística (H'n) total anual fue de 2.24 en Candelaria-Panlau, 1.29 en Chumpan-Balchacah, 1.91 en Palizada-del Este y 2.52 en Pom-Atasta. 5. El promedio de la biomasa total anual (g/peso húmedo /m2) fue de 8.61 en Candelaria-Panlau, 2.70 en Chumpan-Balchacah 2.85 en Palizada-del Este y 8.79 en Pom-Atasta. 6. Las categorías ictiotróficas indicaron 16 especies (34%) consumidoras de primer orden, aportando una biomasa de 2.51 g/m2/ año, 21 especies (42.5%) consumidores de segundo orden con una biomasa de 4.95 g/m2/año y consumidores de tercer orden 10 especies (23.4%) con una biomasa de 1.00 g/m2/año. 7. Esto determina que los sistemas fluvio-lagunares tengan una productividad pesquera de 8.5 g/m2/año. (Nota: g/m2 x 10 da Kg/Ha.) 8. La afinidad ecológica en los 4 sistemas fluvio-lagunares muestra un alto grado de significado estadístico (58 a 76%) básicamente por la distribución y presencia de las mismas especies residentes permanentes, además de compartir muchas de las especies que dominan en biomasa, número y frecuencia. 9. Las especies Arius melanopus, Eucinostomus gula, Micropogon furnieri y y Sphoeroides testudineusson especies tipo y residentes permanentes en estas comunidades. Además de éstas, las especies a continuación son características de estos sistemas por biomasa, número y frecuencia: Bairdiella, chrysura, Bairdiella ronchus, Chaetodipterus faber, Epinephelus guttatus, Cichlasoma fenestratum., Dasyatis sabina, Arius felis, Anchoa mitchilli mitchilli, Cetengraulis edentulus, Bagre marinus yAchirus lineatus 10. Los 4 sistemas fluvio-lagunares representan áreas naturales de crianza y/o alimentación para un gran número de especies de importancia comercial en la Bahía de Campeche y que se reclutan en estos estuarios en grandes cardúmenes de juveniles a fines del verano y comienzo del otoño. Estos sistemas tienen productividad pesquera baja, aunque de especies comerciales que tienen una alta productividad pesquera en la Bahía de Campeche. Por lo tanto, la importancia ecológica de los sistemas fluvio-lagunares asociados a la Laguna de Términos es fundamental y deben tener prioridad en el manejo y protección al utilizar la zona costera. Estos sistemas son exportadores de energía hacia la propia Laguna de Términos y, consecuentemente, a la Bahía de Campeche. AgradecimientosLos autores agradecen a Alfredo Laguarda-Figueras, José L. Rojas Galavíz y Rodolfo Cruz-Orozco, el apoyo y facilidades otorgadas para la realización de este estudio. A Antonio García-Cubas, Jorge Carranza Fraser, Luis R. Segura y Alfredo Laguarda-Figueras la revisión y comentarios del manuscrito, a Violeta Leyton por su ayuda en algunas colectas y confección de mapas y gráficas. A Zoila Castillo por su ayuda en la elaboración de tablas. Auspiciaron este estudio el Centro de Ciencias del Mar y Limnología de la UNAM y la Organización de los Estados Americanos (OEA) a través del Programa Regional de Desarrollo Científico y Tecnológico, Oficina de Proyectos Especiales. Programa de estudios denominados: "Recursos Bióticos en Lagunas Costeras, Manglares y Areas Adyacentes en América Latina (Proyectos Pilotos Comparativos en el Golfo de Nicoya, Costa Rica, Bahía de Puerto Viejo, República Dominicana, Laguna de Términos, Campeche, México)." LITERATURAALLEN, L. G. M. H. HORN, Abundance, diversity and seasonality of fishes in Colorado Lagoon, Alamitos Bay, California. Estuar. Coast. mar. Sci. 1975. 371-380. 3 AYALA-CASTAÑARES, A., Sistemática y distribución de los foraminíferos recientes de la Laguna de Términos, Campeche, México. Bol. Inst. Geol. Univ. Nal. Autón. México, 1963. 1-130. (3): 67 BETCHEL, T. J. B. J. COPELAND Fish species diversity indices as indicators of pollution in Galveston Bay, Texas. Publs. Inst. Mar Sci. Univ. Texas, 1970 10-132 15 BONETTO, A. W. DIONI C. PIGNALBERI, Physis, Nuevos datos sobre las poblaciones de peces en ambientes lénticos permanentes del Paraná Medio. B. Aires 1970 54-141. 80 30 BOTELLO, A. V., An. Centro de Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Variación de los parámetros hidrológicos en las épocas de sequía y lluvias (mayo y noviembre de 1974) en la Laguna de Términos, Campeche, México. 1978. 159-178. (1): 5 ---, E. F. MANDELLI . Bull. Environ. Contam. Toxicol. Distribution of N-Paraffisn in seagrasses, benthic algae, oysters and recent sediments from Térninos Lagoon, Campeche, México. 1978 162-170. (2): 19 BRAVO-NÚÑEZ, E. A. YÁÑEZ-ARANCIBIA An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México Ecología de la Boca de Puerto Real, Laguna de Términos, Sur del Golfo de México. 1. Descripción del área y análisis estructural de las comunidades de peces. 1979. 125-182. (1): 6 CAIN, R. L. J. M. DEAN Annual occurrences, abundance and diversity of fish in a South Carolina Intertidal creek. Mar. Biol., 1976. 369-379. 36: CARVAJAL, R. J. Lagena, Condiciones ambientales y productividad de la Laguna de Términos, Campeche, México. 1973. 35-58. 31: Contribución al conocimiento de los robalos Centropomus undecimalis y C. poeyi en la Laguna de Términos, Campeche, México. Bol. Inst. Oceanogr. Univ. Oriente, 1975. 51-70. (1): 14 CASTRO, J. L., An Inst. Nal. Inv. Biol.-pesq Peces sierra, rayas, mantas y especies afines de México. 1965. 170-256. 1: CERVIGON, M. F., Los Peces Marinos de Venezuela. Estación de Investigaciones Marinas de Margarita, Fundación La Salle de Ciencias Naturales, Caracas, 1966. Monogr. 11 y 12 1-951 (2 vols.). Los Peces Marinos de Venezuela. Complemento IV. Contribución Científica 5, Universidad de Oriente, Nueva Esparta, 1975. 45 p. CHESTNUT, A. F., Oyster Reefs. Coastal Ecological System in the United States. , In: Odum, H. T., B. J. Copeland y E. A. Mc Mahan (Eds.) The Conservation Foundation Wash. D. C. 1974. 171-203. 2: 1974, COLL DE HURTADO A., El sureste de Campeche y sus recursos naturales. Inst. Geogr. Univ. Nal. Autón. México. Serie Cuadernos, 1975. 85 p. COPELAND, P. J. W. S. BIRKHEAD First Annual Report to Carolina Power and hight Co., Raleigh, N. C., Some ecological studies of the lower Cape Fear River estuary, ocean out fall and Dutchman Greek, 1971. 1972. 1-105. 71: CRUZ-OROZCO, R. F. LEY LOU R. LECUANDA F. ROSALES-LÓPEZ Programa de Estudios: Recursos Bióticos en Lagunas Costeras, Manglares y Areas Adyacentes de América Latina. Informe preliminar sobre los estudios de geología, Laguna de Términos, Campeche, México. Centro Cienc. del Mar y Limnol., UNAM-OEA. Mimeografeado 1977. 21 p. DAME, R. F. Estuar. Energy flow in an Intertidal oyster Population. Coast. mar. Sci. 1975. 243-253. 4: DAHLBERG, D. M. E. P. ODUM Annual cycles of species ocurrence, abundance, and diversity in Georgia Estuarine fish populations. Amer. Midl. Natur., 1970. 382-392. (2): 83 DAY, J. W. W. G. SMITH P. R. WAGNER W. C. STONE Community structure and carbon budget of a salt marsh and shallow bay estuarine system in Louisiana. Publ. No. LSU-56-7204 Center of Wetland Resources Louisiana, State University, Baton Rouge, La. 1973b. 79 p. EDWARDS, R. R. C., Estuar. Production ecology of two caribbean marine ecosystems. I. Phisical environment and fauna. Coast. mar Sci. 1973a. 303-318. 1: Estuar. Production ecology of two caribbean marine ecosystems. 11. Metabolism and energy flow. Coast. mar Sci.., 1973b. 319-333. 1: Estuar. Ecology of a coastal lagoon complex in México. Coast. mar Sci. 1978. 75-92. 6: GARCÍA, E., Modificaciones al Sistema de Clasificación Climática de Koppen (para adaptarlo a las condiciones de la República Mexicana). Instituto de Geografía Univ. Nal. Autón. México, 1973. 246 p. GARCÍA-CUBAS, A. J., Sistemática y ditribución de los micromoluscos de la Laguna de Términos, Campeche, México Bol. Inst. Geol. Univ. Nal. Autón. México, 1963. 1-55. 67: GÓMEZ-AGUIRRE, S., Tesis Profesional Comportamiento estacional del plancton de la Laguna de Términos, Campeche, México. Fac. Ciencias Univ. Nal. Autón. México. 1965a. 106 p. An. Inst. Biol. Univ. Nal. Autón. México, Algunas consideraciones acerca del fi toplancton primaveral en la Boca de Paso Real, Campeche. 1965b. 65-69. (1-2): 36 GÓMEZ-AGUIRRE, S. An. Centro Cíenc. del Mar y Limnol. Univ. Nal. Autón. México, Reconocimientos estacionales de hidrología y plancton en la Laguna de Términos, Campeche, México. 1974. 61-82. (1): 1 GREENWOOD, P. H. D. E. ROSEN S. H. WEITZMANN G. S. MYERS Bull. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Amer. Mus. Nat. Hist., 1966. 339-456. (4): 131 GREENWOOD, P. H. G. S. MYERS D. E. ROSEN S. E. WEITZMANN Named main divisions of teleostean fishes. Proc. Biol. Soc. Wash. 1967. 227-228. 80: HAEDRICH, L. R. S.O. HAEDRICH Estuar. A seasonal survey of the fishes in the Mystic River, a polluted estuary in downtown Boston, Massachusetts. Coast. mar Sci. 1974. 59-73. 2: HEALD, E. J. W. E. ODUM The contribution of mangrove swamps to Florida fisheries. Proc. Gulf Carib. Fishes. Inst. 1970. 130-135. 22: ------ D. C. TABB Environment of South Florida: Present and Past. Mangroves in the estuaine food. chain. In: Gleason, J. (Ed.) Miami Geological Society Memoir 1974. 182-189. 2: HELLIER, T. R., Fish production and biomass studies in relation to photosynthesis in the Laguna Madre of Texas. Publs. Inst. Mar Sci. Univ. Texas, 1962. 212-215. 8: HESTER, J. W. B. J. COPELAND Sea Grant Program Publ. UNC-SG75-02, Nekton population dynamics in the Albermal Sound and Neuse River Estuaries. University of North Carolina. 1975. 129 p. HILDEBRAND, S. F., Bull. A review of the American anchovies (Family Engraulidae), Bingham Oceanogr. Coll., 1943. 1-165. (2) 8 HILLMAN, R. E. N. W. DAVIS J. WENNEMER Estuar. Abundance, diversity and stability in shore zone fish communities in an arca of Long Island Sound affected by the thermal dicharge of Nuclear Power Station. Coast. mar. Sci., 1977. 355-381. 5: HOESE, H. D. B. J. COPELAND F. N. MOSELEY E. D. LANE Fauna of the Aransas Pass Inlet, Texas. III. Diel and seasonal variations in trawlable organisms of the adjacent arca. Tex. J. Sci., 1968. 33-60. 20: HOFF, G. J RICHARD M. IBARRA Estuar. Factors affecting the seaonal abundance, composition and diversity of fishes in a Southeastern New England Estuary. Coast. mar Sci., 1977. 665-678. 5: HOLCIK, J., Vestnik. Cs. spol. Zool. (Acta soc. zool. Bohemoslov.), Standing crop, abundance, production, and some ecologial aspect of fish populations in some inland waters of Cuba. 1970. 184-201. 34: HORN, M. H. L. G. ALLEN Bull. Numbers of species and faunal resemblance of marine fishes in California bays and estuaries. South Cal. Acad. Sci., 1976. 159-170. (2): 75 JORDAN, D. S. B. W. EVERMAN Bull. The fishes of North and Middle America. U.S. Nat. Mus., 1896-1900. 1-3313. (47): 1-4 KIKUCHI, T., An ecological study on animal communities of the Zostera marina belt in Tornioka Bay, Amakusa, Kyushu. Publ. Amakusa Mar. Biol. Lab., 1966. 1-106. 1: KJELSON, M. A., Proceedings of the Conference on Assessing the Effects of Power-Plant Induced Mortality on Fish Populations, Estimating ahe size of juvenile fish populations in Southeastern coastal-plain estuarries. In: Webster Van Winkle (Ed.) Pergamon Press, New York. Gatlinburg, Tenn., 1977. 1977. 71-90. Catch efficiencies of a 6.1 meter otter trawl for estuarine fish populations. Trans. Am. Fish. Soc., 1978. 246-254. (2): 107 ----- D. R. COLBY Estuarine Processes, The evaluation and use of gear efficiencies in the estimation of estuarine fish abundance. In: Wiley M. L. (Ed.). Academic Press, Inc., New York, 1977. 416-424. 2: G. N. JONSHON R. L. WATSON Annual Report to the Atomic Energy Comission. Biomass and densities of the fish and macrovertebrates of the New Port Estuary, North Carolina In: Rice, T. R. (Ed.) U.S. Dept. Commerce, National Marine Fisheries Service, Beaufort, N.C., 1973. :169-182. ---, W. R. TURNER G. N. JONSHON Description of a stationary drop-net for estimating nekton abundance in shallow waters. Trans. Am. Fish. Sci. 1975. 46-49. 1 104 KUENZLER, E. J., Coastal Ecological System of the United States. Mangrove swarnp systems. In: Odum, H. T., B. J. Copeland y E. A. Mc Mahan (Eds.) The Conservation Foundation. Wash. D.C. 1974, 1974. 346-371. 1: KUSHLAN, J. A., Quantitative sampling of fish population in shallow water environments. Trans. Am. Fish. Soc. 1974. 348-352 (27): 103 LAGLER, K. F., J. M. KAPETSKY, D. J. STEWART, The fisheries of the Kafue flats, Zambia, in relation to the Kafue George dam. Univ. Michigan Tech. Rept., FAO, Rome, N9 FI:FS/ZAM II Tech. Rept., v: 1971. 1-161. LAGUARDA-FIGUERAS, A., Estudio sistemático y distribución de Sagitta friederici en la Laguna de Términos, Campeche, México. An. Inst. Biol. Univ. Nal. Autón. México. Ser. Cienc. del Mar y Limnol., 1967. 47-57. 1 38 LEÓN, E. P., Ecología de la ictiofauna del Golfo de Nicoya, Costa Rica, un estuario tropical. Rev. Biol. Trop., 1973. 5-30. (1) 21 LIVIGSTON, J. R., Estuar. Diurnal and seasonal fluctuations of organisms in a North Florida estuary Coast. mar. Sci., 1976. 373-400. 4: LOESCH, H., J. BISHOP, A. CROWE, R. KUCKYR, P. WAGNER, Technique for estimating trawl efficiency in catching brown shrimp (Penaeus aztecus), Atlantic croaker (Micropogon undulatus) and spot (Leitostomus xanthurus). 5(2):29-33. Gulf Res. Rep., 1972. LOWE-MC CONNELL, R. H., Fish Communities in Tropical Freshwaters. Longman Inc. New York, 1975. 337 p. LOYO-REBOLLEDO, M. E., Notas acerca de la flora de diatomeas de Laguna de Términos, Campeche, México. An. Inst. Bíol. Univ. Nal. Autón. México, 1965. 61-64. (1-2) 36 MAGO, F. M. L., Acta Venez., Estudios preliminares sobre la ecología de los peces de los llanos de Venezuela. 1970 71-102 7 MARRÓN-AGUILAR, M. A., Tesis doctoral Estudio cuantitativo y sistemático de los poliquetos (Annelida, Polychaeta) bentónicos de la Laguna de Términos, Campeche, México. Fac. Ciencias Univ. Autón. México, 1975. 143 p. ---, E. LÓPEZ-OCHOTERENA, Protozoarios ciliados de México. XVI. Sistemática de algunas especies del orden Tintinnida, Kofoid y Campbell, de la Laguna de Términos, Campeche, México. Revta. Soc. mex. Hist. Nat., 1969. 43-64. 30: MARSHALL ADAMS, S., The ecology of eelgrass, Zostera marina (L.) fish communities. I. Structural analysis. J. exp. mar. Biol. Ecol., 1976. 269-291. 22: Mc ERLEAN, A. J., S. G. O´CONNOR, J. A. MIHUSKY, CH. I. GIBSON, Estuar. Abundance, diversity, and seasonal pattens of estuarine fish populations. Coast, mar. Sci., 1973. 19-36. 1 MÉEK, E. S., S. F. HILDERBAND, The marine fishes of Panama. , Field Mus. Nat. Hit. Publ., Zool. Ser. 1923-1928. 1-1045. (215, 226 y 249) 15 MILLER, J. M., A trawl survey of the shallow gulf fishes near Port Aransas, Texas. , Publ. Ins. Mar. Sci. Univ. Texas 1965. 80-107. 10: MOORE, H. R., Estuar. Variation in the diversity of summer estuarine fish populations in Aransas Bay, Texas, 1966-1973. Coast. mar. Sci., 1978. 495-501. 6: MORALES, G. A., Ecology, distribution and taxonomy of recent Ostracoda of the Laguna de Términos, Campeche, México. Bol. Inst. Geol. Univ. Nal. Autón. México, 1966. 1-103. 81: MOSELEY, F. N., B. J. COPELAND, A portable dropnet for representative sampling of nekton., Publ. Inst. Mar. Sci. Univ. Texas 1969. 37-45. 14: MUUS, B. J., The fauna. of Danish estuaries and lagoons. Meddelelson foa Danmarks Fiskeri-og Havundessolgelsen, Ny Serie 1967. 1-316. (1) 5 NIXON, S. W., C. A. OVIATT, Preliminary measurement of midsummer metabolism in beds of eelgrass, Zostera marina. Ecology, 1972. 150-153. 53: ODUM, W. E., Bull. Pathways of energy floy in a South Florida estuary. Sea Grant Program Tech.Univ. Miami, 1971 1-162 7 ODUM, W. E., E. J. HEALD, Bull. Trophic analysis of an estuarine mangrove community. Mar. Sci., 1972. 671-738. (3) 22 The detritus-based food web of an estuarine mangrove community. Chemistry, Bialogy and the Estuarine System. Academic Press Inc. Nueva York, 1975. 265-286. 1: ---, J. C. ZIEMAN, E. J. HEALD, Proceeding of the Coastal Marsh and Estuary Management Symposium. The importance og vascular plant detritus to estuaries In: Chabreck, R. A. (Ed.) Louisiana State University, Baton Rouge, La., 1973. 91-114. July 1972, ORTEGA, M. M. Tesis doctoral Contribution á l'étude de la vegetation de la Laguna de Téminos (Campeche, Mexique). Fac. Ciencias Univ. Paris, Francia, 1969. 60 p. OVIATT, A. C., S. W. NIXON Estuar. The demersal fish of Narraganett Bay: An analysis of communíty structure, distribution and abundance. Coast. mar. Sci., 1973. 361-378. 1: PHLEGER, F. B., A. AYALA-CASTAÑARES, Bull. Processes and history of Términos Lagoon, México. Am. Ass. Petrol. Geol., 1971. 2130-2140. (2) 55 RESÉNDEZ, M. A. An. Inst. Biol. Univ. Nal. Autón. México, Ser. Cienc. del Mar y Limnol., Estudio de los peces de la Laguna de Tamiahua, Veracruz, México 1970. 79-146 (1) 41 Revta. Soc. mex. Hist. Nat., Estudio de los peces de la Laguna de Alvarado, Veracruz, México. 1973. 183-281. 34: RICHARD, S. W., Bull. The demersal fish populations of Long Island Sound. Bingham Oceanogr. Coll., 1963. 1-101. 18: RUIZ, M . E., Tesis profesional, Estudio ecológico preliminar de las almejas comerciales del sistema lagunar de Términos, Campeche, Rangia cuneata (Gray, 1831). Fac. Ciencias Univ. Nal. Autón. México, 1975. 80 p. SAUER, J., Geographic reconnaissance of seashore vegetation along the Mexican Gulf Coast. Univ. Press. Coastal Studies Series, Louisiana State, 1967. 1-59. 21 SHANNON, C. E., WEAVER The Mathematical Theory of Communication. University of Illinois Press. Ubana. 1963. 119 p. SIGNORET, M., An. Inst. Biol. Univ. Nal. Autón. México. Ser. Zoología, Abundancia, tamaño y distribución de camarones (Crustacea Peneidae) de- la Laguna de Términos, Campeche, México y su relación con algunos factores hidrológicos. 1974. 119-140. (1): 45 SILVA-BÁRCENAS, A., Sistemática y distribución de los géneros de diatomeas de la Laguna de Términos, Campeche, México. Bol. Inst. Geol. Univ. Nal. Autón. México, 1963. 1-131. (2) 67 SNEDAKER, S. C., A. E. LUGO, Rep. Bur. Sports Fisheries and Willife Contract. No.14-16-008-606 The role of mangrove ecosystems in the maintenance of environmental quality and a high productivity of desirable fishes. 1973. 381 p. SOLÍS-WOLFOWITZ, V., Contribución al conocimiento de las ascidias de la Laguna de Términos. Problema de Investigación. (mimeogr.). Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, 1973. 62 p. S. R. H., Estudio de la calidad del agua en la Laguna de Términos, Campeche, Secretaría de Recursos Hidráulicos, Subsecretaría de Planeación, Dirección de Control de la Contaminación del Agua. Contrato ATECMAR, S. A., Núm. SP75-C-I. 1976. SUÁREZ-CAABRO, J. A., S. GÓMEZ-AGUIRRE, Bull. Observaciones sobre el plancton de la Laguna de Términos, Campeche, México Mar. Sci., 1965. 1072-1120. (4) 15 SUBRAHMANYAM, C. B., S. H. DRAKE, Bull. Studies on the animal communities in two north Florida salt marshes. Part 1: Fish communities. Mar. Sci., 1975. 445-465. (4): 25 THAYER, G. W., S. MARSHALL ADAMS M. W. LACROIX, Estuarine Research. Chemistry, Biology and the Estuarine System. Structural and functional aspects of a recently stablished Zostera marina communities. In: Cronin, L. E. (Ed.). Academic Press, Inc. New York, 1975. 1: THOM, B. G., Problems of the development of Isla del Carmen, Campeche, México. Zutschr, Geomorphologie, N. F. Bd., 1969. 406-413. (4) 13 TORAL, S., Tesis profesional, Estudios de los Cichlidae (Pisces, Perciformes) de la Laguna de Términos y sus afluentes. Fac. Ciencias Univ. Nal. Autón. México, 1971. 32 p. ----, A. RESENDEZ, Los cíclidos (Pisces, Perciformes) de la Laguna de Términos y sus afluentes. Rev. Biol. Trop., 1974. 254-274. (2) 21 TURNER, W. R. G. N. JONSHON, Distribution and relative abundane of fishes in Newport River, North Carolina. NOAA Tech. Rep. NMES SSRF-666 1973. 1-23. VARGAS, F. M., Tesis profesional, Las corrientes y el transporte neto de agua en la Laguna de Términos, Camp. Fac. Ciencias, Univ. Nal. Autón. México. 1978. 94 p. WILHM, J. L., Ecology, Use of biomass units in Shanno's formula. 1968. 153-156. (1) 19 YÁÑEZ-ARANCIBIA, A., Symposium on Progress in Marine Research in the Caribbean and Adjacent-Regions, Caracas Venezuela, 12-16 July 1976. Paper on Fisheries, Aquaculture and Marine Biology. Piscicultura en lagunas costeras: Perspectivas en México. In: Stewart, H. B. (Ed.) FAO Fish. Rep., 1977. 529-547. (200): An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Patrones ecológicos y variación cíclica de la estructura trófica de las comunidades nectónicas en lagunas costeras del Pacífico de México. 1978a. 285-306. (1) 5 Seminar on Present and Future Research in Coastal Lagoons. Fishes ocurrence, diversity, and abundance in two tropical coastal lagoon with ephemeral inlets on the Pacific Coast of México. Duke University Marine Laboratory, IABO and UNESCO, Beaufort N.C. 1978b. Aug. 28 - Sep. 2, 1978 (in press). Taxonomía, ecología y estructura de las comunidades de peces en nueve Lagunas Costeras del Pacífico de México. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Publ. Espec., 1978c. 1-306. 2: Seminar on Present and Future Research in Coastal Lagoon. Ecology in the entrance of Puerto Real, Términos Lagoon, South Gulf of México 11. Discusión on trophic structure of fish communities in banks of Thalassia testudinum. Duke University Marine Laboratory, IABO, and UNESCO, Beaufort NC. 1978d. Aug. 28-Sep. 2, 1978. (In press.) ------, R. S. NUGENT An. Cento Cienc. del Mar y Limnol. Univ. Nal. Autón. México El papel ecológico de los peces en estuarios y lagunas costeras. 1977. 107-114. (1) 4 --- , F. AMEZCUA-LINARES, An Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México, Ecología de la raya Urolophus jamaicensis (Cuvier) en Laguna de Términos, un sistema estuarino del Sur del Golfo de México. (Pisces: Urolophidae.) 1979. (2): 6 YÁÑEZ-CORREA, A., Batimetría, salinidad, temperatura y distribución de los sedimentos recientes en la Laguna de Términos, Campeche, México. Bol. Inst. Geol. Univ. Nal. Autón. México, 1963. 1-47. (1) 67 Littoral processes and sediments of the Inner continental Shelf of the Southern Bay of Campeche, M. S. Thesis, Texas A and M Univesity, 1968. 80. p. ZARUR-MENEZ, A., Tesis profesional, Estudio biológico preliminar de la Laguna de Términos, Campeche, México. Fac. Cienc. Univ. Nal. Autón. México, 1961. 64 p.
|