|
ON
THECAL MORPHOLOGY OF DINOFLAGELLATES WITH SPECIAL EMPHASIS ON CIRCULAR AND
SULCAL PLATES
Trabajo recibido el 5 de octubre de 1978 y aceptado para su
publicación el 6 de noviembre de 1978.
ENRIQUE BALECH
Museo Argentino de Ciencias
Naturales "B. Ravidavia". Av. A. Gallardo 470. 1405 Buenos Aires,
Argentina.
El autor subraya la importancia
taxonómica y filogenética de los caracteres tubulares de los
dinoflagelados tecados, sobre todo de los correspondientes al
cingulum y al sulcus que son las partes
más dinámicas y, al mismo tiempo, las más estables.
Según su experiencia ambas regiones tienen variaciones menores en cada
género, con la excepción, para el cingulum,
del género Pyrophacus.
Los
Prorocentroides tienen dos valvas; las dos placas que la forman aprisionan, en
el extremo de emergencia flagelar, un conjunto de hasta ocho placas
diminutas.
Los Dinophysoidea, también bivalvares,
tienen regiones o series tabulares bien definidas y revelan, a pesar de la muy
alta diversidad morfológica, quizás extrema entre los tecados, un
plan tabular único que sólo experimenta, de un género a
otro, variaciones menores. Esta unidad tabular quedó demostrada en sus
últimas investigaciones. El autor expone las características
tabulares principales de los distintos géneros.
Los
Peridinoidea, en cambio, no son bivalvares y no tienen tampoco un plan tabular
único para el orden, pues difiere bastante aún entre
géneros próximos. Llama la atención, sin embargo, sobre el
hecho de que a veces la mayor disparidad se debe a distintos criterios
utilizados para definir las series tabulares (tal el caso de algunas de las
diferencias entre Goniodoma y Gonyaulax).
Por lo tanto propone un criterio único y sostiene que sería
más correcto llamar "perisulcales" a las placas que
hoy denominamos "antapicales".
Observa
también que el recubrimiento de las placas sigue un orden definido y,
por último, expresa su desacuerdo sobre el término
"poros de tricocistos" porque los así llamados
parecen cumplir otras funciones.
L'auteur soutent la grande importance
taxinomique et phylogenétique de la tabulation des dinoflagellés,
surtout de celles du cingulum et du
sulcus qui sont les parties plus dynamiques de la cellule
et, en méme temps, les plus stables: elles ne changent pas
fondamentelement dans chaque genre, avec l'exception, à l'égard
du cingulum, de Pyrophacus.
Les Prorocentroidet ont une structure à deux valves; les deux
plaques que forment les valves emprisonnent à l'extrême où
émergent les flagelles un centain nombre de plaques tràs petites
(jusqu'à 8).
Les Dinoprysoidea, bivalves aussi, ont
des régions tabulaires bien définies; chacune d'elles
étant formés, en principe, par 4 plaques. Malgré les
grandes differeces morphologiques qu'on voit dans cet ordre tous
répondent à un plan tabulaire unique, comme a été
constaté par nos derniàres études. On donne un
détail des principaux traits tabulaires des diffrents genres.
Les Peridinioidea ne scnt pas bivalves et n'ont par un plan tabulaire
unique puisque la tabulation varie beaucoup, même parmi des genres
très voisins morphologiquement. Quand même quelques
différences ne sont pas réelles et elles sont dues aux differents
critériums emplyés pour la détermination des séores
(telles, par exemple, quelques differences entre Gonyaulax
et Goniodoma). Pour liminers ces différences
artificielles l'auteur propose un critérium unique; il pense que les
plaques génaralement appellées "antapicales"
seraient mieux. dénommées
"perisulcales".
L'auteur observe que les
recouvremnets des plaques semolent suivre un ordre fixe et, finalement, il fait
connaître son desaccord avec le nom. "pores des trichocystes" car ces
pores semblent bien repondre à d'autres fonctions que le rejet de ces
organites.
In April 1978, a Penrose Conference was held in Colorado Springs, USA. It was intented to examine some of the most important problems concerning the mophhology and taxonomy of the extant and fossil dinoflagellates, and to review our knowledge on these points. The present paper is an invited presentation to that conference. Its aims were to clarity some of the points concerning thecal morphology of extant dinoflagellates, to stress the importance of the cingular and sulcal structure and to propose some modifications of the usual platenomenclature. RESULTS AND DISCUSSIONThe author has been dissecting thousands of thecae, during some 35 years, what gives him some experience in the anatomy and taxonomy of the thecate members of this group, which may be of some interest to the specialists of extint dinoflagellates, and perhaps also to the micropaleontologists. Through all these years he became convinced that in order to do good work on geographic distribution, ecology and biology we have to do good taxonomy, and to do so it is necessary, for the majority of the species, to dissect them and to. take into account a number of characters (even sculpture has some taxonomic value). Nevertheles some of the characters are more important than others. The tabular are of utmost importance but the tabulation of every regions has not the same weight. Undoubtedly the epitheca is the one which shows more variation in the number and relations of the plates: the number varies in some species, especially when they live in unusual ecological conditions. That is frequently seen in cultures where there are quite a number of abnormal individuals, and they survive only because the natural mechanisms of the elimination of the least apt does not function. The tabulation of the hypotheca is always much more conservative and that can also be said of the cingular and sulcal series. It is interesting to note that these regions of maximum stability, especially cingulum and sulcus are the most dynamic of the cells because they are related to the flagellar movements, outlet of the pusulae, phagocytocis, etcetera. According to the autor's experience the sulcus of any species is a fixed structure with variation on a very narrow scale. The tabulation of the cingulum is also fixed in all the genera with one and very striking exception: Pyrophacus. Let us take a look first at the situation in the three orders of thecate dinoflagellates. In the Prorocentroidea the theca is generally described as consisting of two valves, each one formed of a single plate, which imprison in the ventral extreme a small plate which has two pores through which the flagella emerge. Nevertheless careful dissections show that the small flagellar plate is truly made up of several quite small ones up to eight in number. A prolonged and meticulous investigation of many thecae of various species is still necessary. At present we do not know wheter the number of plates is fixed in the genus or at least in each species. In any case the Prorocentridae are, with a maximum number of 10 plates, the simplest thecate we know. The Dinophysioidea are also bivalvar, but much more complicated since each valve is constituted of several plates. Besides there are in them, clearly constitued the four regions also present in the Peridinioidea: epitheca, hypotheca, cingulum and sulcus. A general and detailed examination of the Dinophysioidea shows us two apparently opposite characteristics: extreme morphologic diversity and, at the same time, a structural plan common to all the genera, with only relatively minor differences. If we take as an example any species of Dinophysis we see that epitheca and hypotheca, are each formed of four main plates: two major or dorsal and two ventral. However, in the epitheca, very close to its ventral edge, one or two small plates delimit a kind of pore, constituting the apical series (Balech, 1976). As seen the left sulcal list has two separable parts which correspond to the left and right ventral hypothecal plates. Each one of them is formed by a narrow body, a kind of rod which supports a part of the list. But the left plate is advanced with respect to the right. The right sulcal list is a simple continuation of the right posterior cingular list and, therefore, an integral part of the right dorsal plate. The sulcus is formed also by four principal plates but only two of them, the left and the right lie at both sides of the sagittal plane. One of the other two is anterior and the other is posterior. The latter is bigger and the most characteristic. It usually has the form of an axe with rather a wide handle; in front and to the left is the part which represents the blade of the axe, of a variable shape, high or low, not very or very protruded, but always more or less clearly discernable. At least in some species those four plates surround a fifth plate, membranous, very slender and fragile, which the author calls medium sulcal. This was evident in very few species but it is possible that it exist in all of them: its transparency and fragility make it difficult to see it and, besides, it is certainly easy lost due to loosing, caving in or destruction. 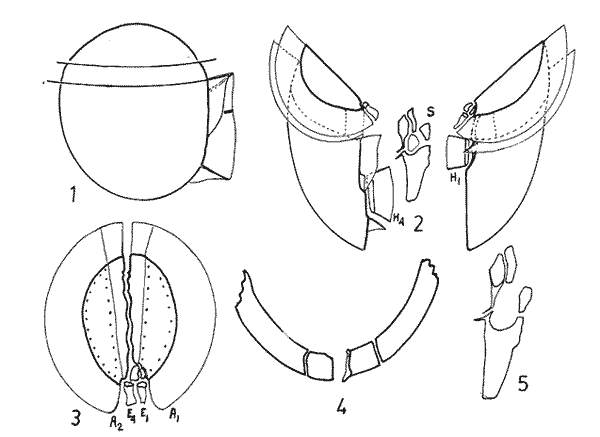 Fig. 1. Structure of Dinophysis. 1, right lateral view; 2, the two moieties; 3, apical view of the epitheca with plates somewhat separated; 4, cingular plates; 5, sulcal plates S = sulcal plates. Note in figure 4 the plates H, and H4 which form the left sulcal list. These structures are repeated even in the genera of completely different forms, with only minor variations. In Heteroschisma the apical region becomes more complicated and is formed by three plates. But the biggest difference is that the left ventral hypothecal has a very wide body which forms an important par of the left hypothecal valve. 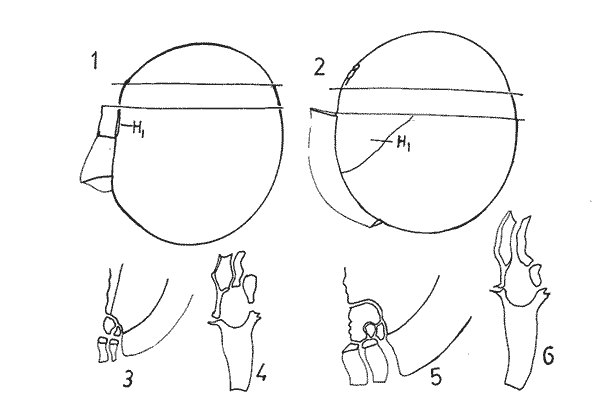 Fig. 2. Heteroschisma compared with Dinophysis. 1. left lateral view of Dinophysis, an 2, the same of Heteroschisma. Note in Heteroschisma the highly developed body of H<1>, and the seemingly lack of the second rib (R2) in theleft sulcal list; 3, E<1>, E<4>, the two apical plates and the ventral end of E<2> of Dinophysis; 4, sulcal plates of Dinophysis; 5, E<1>, E<4> the three apicalplates and the ventral end of E<2> in Heteroschisma; 6, sulcal plates of Heteroschima. In Metaphalacroma we see that the apical region is formed by four relatively robust plates and which apparently have not the left ventral epithecal plate. It seems clear that this apparently missign plate is the first apical. Ornithocercus and Histioneis have a tabular structure which does not differ from. that of Dinophysis. The same can be said of Oxyphysis, in spite of the fact that from its aspect, it might be mistaken for a Peridinioidea (Nie, 1942; Norris, 1969; Tai and Skogsberg, 1934). Let us go now to another two morphologically very different genera of Dinophysioidea. In Amphisolenia the epitheca and the cingulum constitute a very small head in the anterior extreme of a very thin and narrow body. Nevertheles the tabular structure remains unchanged. The sulcus forms the ventral part of a long and very narrow neck and consequently the S.a. and S.d. plates have lengthened a lot forming a pair of long rods. The S.p. has the general form, that we see in other genera with, however, a striking difference: the plate seems to have revolved 180º on its axis and therefore the blade of the axe is directed towards the right instead to towards the left. 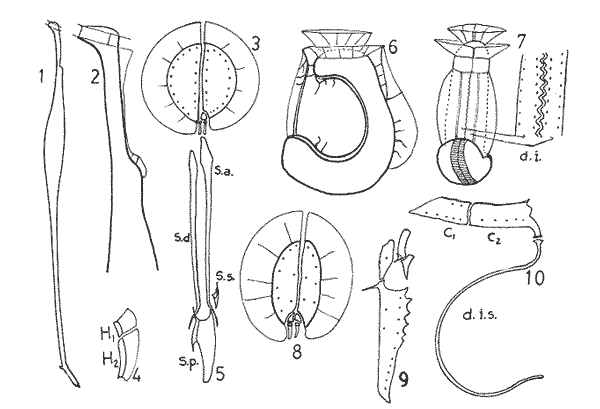 Fig. 3. Structures of two genera of Dinophysoidea which greatly differ in shapefrom Dinophysis; 1-5, Amphisolenia; 1, rigt ventral view; 2, detail of the anterior region of the theca; 3, epithecal plates; 4, the plates which form the left sulcal list; 5, sulcal plates. 6-10 Citharistes; 6, right lateral view;7, dorsal view showing the pair of extra plates or dorsal intercalaries(d.i.); 8, epithecal plates; 9, sulcal plates; 10, the two cingular plates and the intercalary (side view) of the left valve. The strangest of the genera, Citharistes, very rare and with only two species has, nevertheless, a sulcus that does not differ significantly from that of Dinophysis. But it is the only genus which presents a major tabular difference. in the dorsal region of the hypotheca, separating the two main plates, appear another two extra plates. However summing up, the most stricking character of the Dinophysoidea is that in spite of the morphologic differences there is an amazingly constant structural plan through out the whole order. The situation is very different, almost the inverse, in the Peridinioidea. At least many of the genera do not differ fundamentally in their forms (the differences of this type can be greater within a genus than those which separate them from other genera) but, on the contrary, no definite tabular plan for the order is found except a negative one, there is no bivalvar structure. The number of cingular plates, for example, varies from 3 to 12 (even up to 16) and that of the sulcals is also very variable, and the variations of form, type and disposition of the same, are even greater. Let us see now the application of the general rule already established, of the relative importante of the different tabular regions. According to it Balech separated (1959) the genus Scrippsiella from the marine Peridinium, although its general tabulation, this is epithecal and hypothecal, is the same, the second genus has six cingular plates, and the first only three and one transitional. Although this has been contested for some years it was finally accepted by the majority of specialists. It is interesting to stress that their sulcal structures are crearly different and lastly that the important studies of cysts by Wall and Dale showed that those of both genera differ (Wall and Dale, 1968). 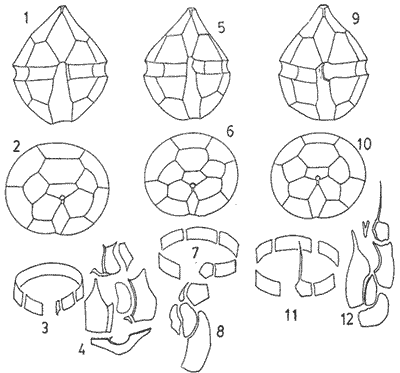 Fig.4. 1-4.Protoperidium. 1,ventral view;2, epithecal plate pattern in apical view; 3, cingular plates and T; 4, sucal plates with T. 5-8. Scrippsiella. 5, ventral view; 6, epithecal plates in apical view; 7, cingular plates and T; 8, sucal plates and T. 9-12, Ensiculifera. 9, ventral view; 10, epithecal plate pattern; 11, cincular plates and T; 12, sulcal plates and T. Note similarities in shape and in epithecal formulae and the keydifferences in cingular and sucal structures. More recently the author separated, for the same reasons, the freshwater Peridinum (Peridinium s. str.) from the marine ones which now have the name of Protoperidinium (Balech, 1974) . Another genus which presents peculiar problems is Gonyaulax which is a complex of forms with at least four epithecal plate patterns. However the rest of the theca, including sulcus, seem to give it sufficient unity. Recently Balech (1977) studied, among others an interesting Arctic species generally called Gonyaulax catenata. Its epitheca has 7 precingulars, a small X-plate of the apical groove and 4 dorsal intercalaries, two of them very small. The differences are too great to allow us to maintain it in Gonyaulax. But is is not a Protoperidium either because it has 6 cinculars, its hypotheca is like that of Gonyaulax and the sulcus resembles that of the latter genus, but with a difference not to be overlooked; its S.d.p (right posterior sulcal) develops a great deal and finishes beyond the right end of the cingulum displacing to the left the S.d.o., a plate which is always connected with the cingulum in Gonyaulax. Peridiniella sphaeroidea has the same type of tabulation (except that there are three anterior intercalaries instead of four). Therefore catenata had to be transferred to Peridiniella. This is another example of the importance of the detailed study of tabulation. Another interesting fact about P. catenata it that two very different forms of it are found: the typical one, short and which forms chains, and the other long which aparently does not form chains. But they are exactly the same in all the other details. It looks like in this species there are two different generations. 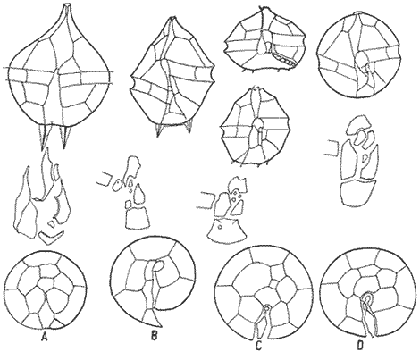 Fig. 5. In vertical rows: A, Protoperidinium pyrum, B, Gonyaulax spinifera, C, Peridiniella catenata and D, Peridiniella sphaeroidea. in horizontal rows, from above, thecae in ventral view (there are two forras of Peridiniella catenata, short and long); middle row, sulcal plates; below, epithecal plate-patterns. It seems convenient to bring to the attention of the specialists that some differences of tabular formulas are due simply to differences of interpretation. Therefore we must try to fix as well as possible, what the series are. In the epitheca this is relatively simple because thase plates that border the small poreplatelet are called apicals. The one exception currently admitted to this rule is the tiny plate, called X, which closes the so-called apical groove of same genera such a Protoperidinium. Althought it would truly be the first apical it is considered a part loosened from Po and therefore the first apical is that which is immediately behind it in the ventral region. In the majority of the Gonyaulax the first is directly connected with Po but in some of the tamarensis-group the connection is made by a slender filiform prolongation and finally in others all contact is lost. We have clearly marked an evolutionary series, and therefore from the practical point of view the plate ought to go on being called first apical. Every plate which borders the cingulum but does not touch Po (with the aforesaid exception) is called precingular and those which touch neither Po nor the cingulum. are intercalary. However the analysis of the hypotheca, is much more complicated and it is not always easy to say where the sulcus ends, which are the antapicals and the intercalaries. We must, however try to normalize as far as possible, the nomenclature to establish through the formulas the real affinities. The sulcus introduces a very special factor here and in some way it can be used in the hypotheca as a reference as we use Po in the epitheca. Thus we call antapical those plates which border the sulcus without being connected with the cingulum (a better name would be "presilla" since some of them are not antapical at all) . An intercalary one, in the contrary, is that which, as in the epitheca, touches neither the cingulum nor the sulcus. Let us apply this criterion to the genera Gonyaulax and Goniodoma. Graham (1942) wrote that the latter has no near relatives and it forms by itself a relatively isolated family. Althoug it is true that, Goniodoma is clearly different from other genera the author does not agree with Graham. It is said that it differs from Gonyaulax by having 5 postcingular instead of 6, no intercalaries and 3 antapical plates instead of one. A deeper analysis shows that the first poscingular of Gonyaulax is always small, narrow, frequently rather sunken in the sulcus and following the species it may be considered a real postcingular or a left anterior sulcal. In Goniodoma there is a similar plate but it is called, with no apparent reason sulcal. The most curious thing is that this plate of Goniodomna is more clearly poscingular than the homologous one of Gonyaulax because in its anterior extreme it has a membranous part which completes the posterior cingular list. 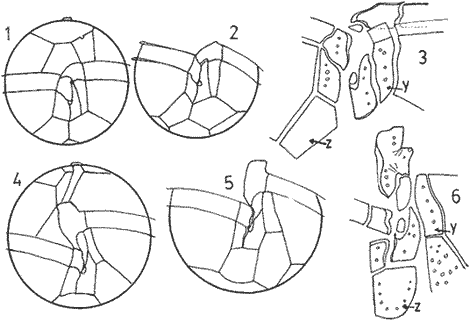 Fig. 6. 1-3, Goniodoma sphaericum: 1, ventral view; 2, slanting hypotheca in ventral posterior view; 3, plates of the ventral region. 4-6, Gonyaulax spharoidea, same explanatíon as for Goniodoma. In both general y and z are homologous plates but they are generally considered to, belong to, different series. The so-called first antapical is exactly equivalent to that which in Gonyaulax is called intercalary. If we apply the aforsaid rule to both genera that plate ought to be called first antapical. Finally in Goniodoma there is a plate which continues the sulcus slanting towards the right and which is considered a third antapical. In this case we face a difficultad decision but it is not unreasonable to consider it as a posterior sulcal; one of the arguments for so doing is that its Position recalls the S.p. of Gonyaulax and of other genera. Besides if it were not a sulcal it would probably be the only genus in which there is no posterior sulcal plate. If we acknowledge these reasonings Gonyaulax and Goniodoma both have 6 cingulars, 6 poncingulars, two antapicals and a rather similar sulcus, all of which indicates that they are two genera very close to each other. The study of the cingular and sulcal plates shows that Protoperidinium is much more related to the Diplopsalis-group and even to the Podolampidae than to Peridinium s. str. and to Scrippsiella. The ephiteca of the Diplopsalis-groups is somewhat simpler than that of Protoperidinium. It generally has three apicals and one or two intercalaries. But its cingulum and its sulcus are of the same type as those of Protoperidinium. 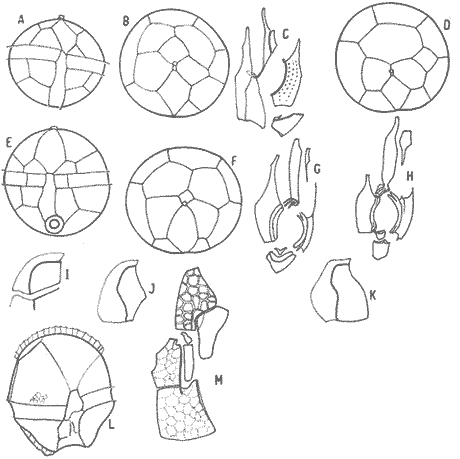 Fig. 7. A-C and I, Protoperidinium (Archaeperidinium) porosum. A, ventral view, B. epithecal plate pattern; C, sulcal plates; 1, plate l. D, H and K, Diplopeltopsis granulosa; D, apical view; H, sulcal plates; K, plate 1. E-G and J, Protoperidinium (?) imitatio. E, ventral view; F, epithecal plate-pattern; G, sulcal plates; J, plate l. Note that the epithecal plate-pattern is that of Archaeperidinium but 1 and the sulcal plates are clearly like those of Diplopeltopsis. L and M, Peridinium willei; L, ventral view; M, sulcal plates. As to confirm this relation I found, a short time ago, a new species which I had to ascribe, on behalf of its general plate-pattern to the subgenus Archoeperidinium of the genus Protoperidinium. However, the location is doubtful and certain details would make it be included in the genus Diplopsalopsis. As in every species of the Diplopsalis-group the four principal sulcal plates forra an "annulus" almost circular and very strong. Also is in all of them the l" ' is narrow anteriorly and holds a very wide sulcal fin strongly bent towards the right and which does not continue in the first antapical (in Protoperidinium it has not got such a spread and it always continues with a list of the antapical) . Another characteristics is that the sulcals have a very strong internal reinforcement which tends to exfoliate. To finish with the plate-pattern we are obliged to make a brief reference to the genus Pyrophacus which seems to be the almost inevitable exception to every rule. Its tabular variation are surprising. The type species, P. horologium has five apicals, 9 precingulars and the same number of cinculars and postcingulars, 3 antapicals and a true posterior intervalary. The other species, bigger, P. steinii, has seven apicals, 12 to 13 precingulars, the same number of postcingulars (exceptionally 14) and a variable number of intercalaries, from 3 to 11. However there are two series that keeps steady in the genus: the antapicals and the sulcus as can be seen in figure 8. 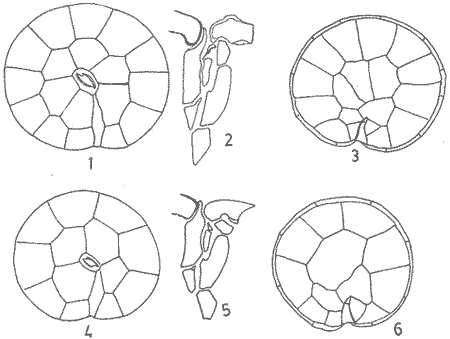 Fig. 8. 1-3, Pyrophacus steinnii; 1, epitheca; 2, sulcal plates and extreme of the last cinguiar plate; 3, antapical view of the hypoteca. 4-6, Pyrophacus horologium, same explanations Let us conclude with a few reflections on some tabular details. The covering of the plates follows, at least in Protoperidinium and in the Diplopsalis-group, as it does in Peridinium, a fixed order that can be classified as centrifugal from the cingulum towards both extremes and from the dorsal to the ventral region. The second observation is that which refers to the so-called trichocyst-pores. The author can not agree with this name. He believes that the fundamental role of the pores of thes theca is not to facilitate the explusion of the trichocysts. Contractions and a certain rotation of the protoplasm make it difficult for this to me the primal aim of the pores. This is more evident in the case of the cingular plates in which the pores are situated precisely in front of the axial plane of the transversal flagellum, that is to say, in the worst place to expel trichocysts. We are prone to believe that in this case they serve to connect the protoplasm with the flagellum by means of fine filaments. Of course this is only a hypothesis. 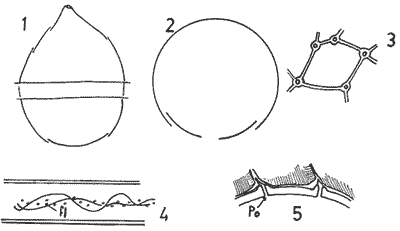 Fig. 9. 1, scheme chowing plate coverings in Protoperidinium, right side view; 2, the covering of cingular plates, apical view; 3, part of the reticulum with poroes in the reticular junctions; 4, part of a cingular plate showing the relationship of the pores with the transverse flagellum (fl); 5, scheme of a section of a general plate through pores (P.o) and ridges As for the pores of the other plates their real function seems to be to facilitate the exchange of substances with the medium and to facilitate the connection of the cytoplasm with the external layer of the theca. The pores frequently open to the outside in the middle of a small protuberance like a crater, and from these protuberances there often extend irregular lines, more or less strong, according to the age of the theca which tend to form a reticulum. These external formations and their growth can be fulvilled only by the protoplasm through the pores. Otherwise entire plates, not perforated would make the processes of absorption and excretion very slow and difficult, and therefore they would prevent the normal cellular metabolism. REMARKS ABOUT THE FIGURESAll the figures are somewhat schematic since they are intented to show structures, not to identify species, magnifications were not calculated. Symbols for plates are those in general use. LITERATURABALECH, E., Bull., Two new genera of dinoflagellates from California, Biol. 1959. 195-23. (2), 116 El género Protoperidinium Bergh, 1881 (Peridinium Ehr., partim). Rev. Mus. Arg. C. Nat. "B. Rivadavia", Hidrobiología, 1974. 1-79. (1): 4 Notas sobre el género Dinophysis (Dinoflagellata), Physis, Sec 1976. 183-193. (91): 35 Cuatro especies de Gonyaulax sensu lato, y consideraciones sobre el género. Rev. Mus. Arg. C. Nat. "B. Rivadavia", Hidrobiología, 1977. 115-136, 3 láms. (6): 5 GRAHAM, H. W., Studies in the morphology, Taxonomy and Ecology of the Peridiniales. Carnegie Inst. Wash. Public., 1942. 1-129. 542: NIE, D., Sinencia Dinoflagellates of the Hainan Region. 7. On the thecal morphology of Ornithocercus thurni Schmidt) Kofoid & Skogsberg. 1942. 23-28. (1-4). 14 NORRIS, D. R., Bull. Thecal morphology of Ornithocercus magnificus (dinoflagellates) with notes on related species. Mar. Sci. 1969. 175-193. (1): 19 WALL, D. B. DALE J. Paleont. Quaternary calcareous dinoflagellates (Calcionellideae) and their natural affinities. 1968. 1935-1408. (6): 42
|