|
CONTRIBUCIÓN AL CONOCIMIENTO DE LA BIOLOGIA Y
ECOLOGIA DE MACROBRACHIUM TENELLUM (SMITH) (CRUSTACEA, DECAPODA,
PALAEMONIDAE)
Trabajo recibido el 28 de junio de 1978 y aceptade para su
publicación el 6 de septiembre de 1978.
RAMIRO ROMÁN-CONTRERAS
Universidad Nacional Autónoma de
México, Facultad de Ciencias. Contribución 188 del Centro
de
Ciencias del Mar y Limnologia, UNAM.
Se analiza la estructura de la población del
langostino Macrobrachium tenellum a través de
un año
de capturas en la Laguna de Tres Palos, Gro; se compara el crecimiento de
machos y hembras con base en registros de la longitud del
cefalotórax y
peso de los ejemplares. Se concluye que M. lenellum
alcanza
su mayor tamaño en la Laguna de Mitla, que presenta
características
dulceacuicolas, siguiendo en tamaño los ejemplares de Tres Palos,
y
Chautengo, en las que, las salinidades son comparativamente más
elevadas.
El tamaño y peso máximos registrados
fueron de 132 mm y 28.6 gramos respectivamente, siendo los ejemplares
machos
los más desarrollados; se establece el rango de fecundidad de la
especie, se analiza la proporción de sexos y se establecen los
periodos
de reproducción y pesquería del recurso.
Population structure of the fresh water prawn
Macrobrachium tenellum is analized throught a year of
captures from Tres Palos coastal lagoon; it compares the growth of males
and
females on registers about the total lenght, cephalotorax lenght and
weight of
the specimens.
It concludes that M. tenellum
reaches the biggest lenght in Mitla lagoon which has fresh water
caracteristics, following in size the specimens from Tres Palos and
Chautengo
lagoons, which salinities are higher than Mitla. Size and weight observed
were
132 mm. and 28.6 g respectively, being the males the more developed.
Growth
rates and lenght are compared with other species; fecundity rates,
reproduction
and fisheries periods of the resource are suggested.
La importancia económica que reviste el grupo de los Paleménidos dulceacuicolas especialmente el de los llamados "langostinos", ha originado en los últimos años el inicio de investigaciones que tienden principalmente hacia el mejor entendimiento de las condiciones óptimas de reproducción, crecimiento y cultivo de aquellas especies susceptibles de ser aprovechadas como alimento para el hombre. En México, los Palemínidos están representados tanto en la vertiente del Golfo de México como a lo largo del Pacifico. En el primero, las especies reportadas son: M. acanthurus, M. olfersii y M. carcinus entre otras; mientras que en la costa del Pacifico se encuentran M. americanum, M. digueti, M. acanthochirus, M. ol. fersii y M. tenellum, de la cual se ocupa el presente trabajo. La importancia del estudio de esta especie estriba en el escaso conocimiento que de ella se tiene; pero además, porque es una especie de importancia comercial, si bien no alcanza las tallas de M. americanum o M. carcinus. El objetivo principal de este trabajo es dar a conocer los resultados obtenidos a través de un año de observaciones de campo, en relación a la abundancia relativa, hábitat, longitud y peso de los individuos, crecimiento, periodos de reproducción, fecundidad y problemas asociados con las poblaciones explotadas de M. tenellum en las lagunas de Chautengo, Mitla y Tres Palos, Gro, México. ANTECEDENTESAunque la especie se encuentra ampliamente distribuida desde Baja California hasta el Norte del Perú, no ha sido suficientemente estudiada por lo que la literatura concerniente es más bien escasa. Uno de los trabajos en que se menciona M. tenellum es el de Holthuis (1952), sobre la revisión de la familia Palaemonidae. En México, Ramírez Granados (1952) señaló la presencia de Palemínidos en las lagunas de Tres Palos y Coyuca, Gro., sin indicar la especie en particular; se infiere, por la abundancia actual del recurso en las lagunas mencionadas, que pudiera tratarse de M. tenellum, aunque no existen otros datos que confirmen esta suposición. Rodríguez de la Cruz (1965 y 1968) realizó trabajos importantes para el conocimiento de los Palemínidos de México, en los que hizo mención de M. tenellum en los ríos Mulegó, Baja California y Presidio, Sinaloa; Stuardo et al. (1974) registró más recientemente poblaciones de la especie en las lagunas de Apozahualco, Mitla, Chautengo, Coyuca y Tres Palos, Gro. AREA DE ESTUDIO
LAGUNA DE TRES PALOSLa Laguna de Tres Palos se localiza en la "Costa Chica" del estado de Guerrero, tiene una superficie aproximada de 50 km2 y se ubica entre los ríos, Papagayo y Sabana (Yañez et al., 1976), aproximadamente a 29 km. al Suroeste del Puerto de Acapulco, a los 16116' y 16049' de latitud norte y 99'39' a 99'46' de longitud oeste (Fig. 1 y 2). LAGUNA DE CHAUTENGOTiene una superficie de aproximada de 34 km 2 se ubica entre los W35' a W38'de latitud norte y 99003' a 99009' de longitud oeste (S.R.H., 1975). Recibe la aportación de los rios Nexpa y Copala durante las épocas de escurrimiento. La profundidad máxima es de 5.7 m. (Castellanos, 1975), durante el periodo de lluvia (Fig. 3). LAGUNA DE MITLATiene una superficie aproximada de 32 a 36 km 2 Se ubica al Nordeste del Puerto de Acapulco, entre los W59' a 17005' de latitud norte y 100' 14' a 100'25' de longitud Oeste. La profundidad media es de 2.9 m y se orienta paralela a la costa (Fig. 4). CARACTERISTICAS DE LOS SITIOS DE COLECTAEn la laguna de Tres Palos las profundidades de colecta variaron desde 0.3 m hasta 1.5 m. El tipo de fondo observado, fue arenoso-compacto, limo-arcilloso con restos de Polimesoda sp y lodos con abundancia de detritus. En los fondos arenosos profundos la captura de langostinos fue escasa, mientras que en fondos someros con abundancia de detritus y entre las raíces de mangles y lirios acuáticos, la captura de juveniles fue abundante. La mayoría de los ejemplares adultos, sin embargo, se colectó siempre en áreas abiertas. Como, fauna acompailante se obtuvieron algunos peces llamados comúnmente "charritas" (Cichlasoma trimaculatum), "cuatetes" (Galeichthys caerulescens), "puritos" ( Phylypnus lateralis) y ocasionalmente algunos peneidos. El rango de salinidad para esta laguna varió entre 1.2% en el mes de octubre (al terminar la época de lluvias) y 2.6% para el mes de mayo, que se registra como época de secas; y la temperatura de 30°C en el mes de marzo a 34.5°C en el mes de mayo. Estos datos dan una idea de las condiciones hidrológicas y corresponden a valores obtenidos durante las fechas de colecta exclusivamente. 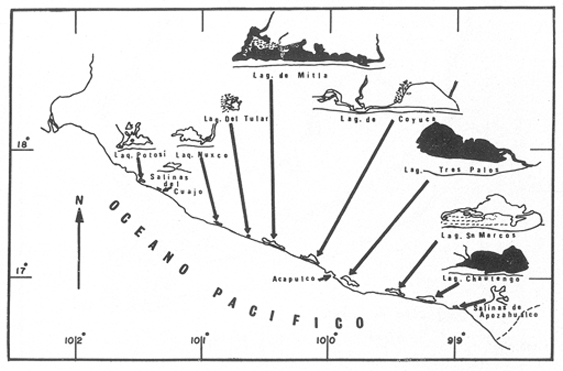 Fig .1. Lagunas Costeras del Estado de Guerrero. Situación geográfica de las lagunas estudiadas. 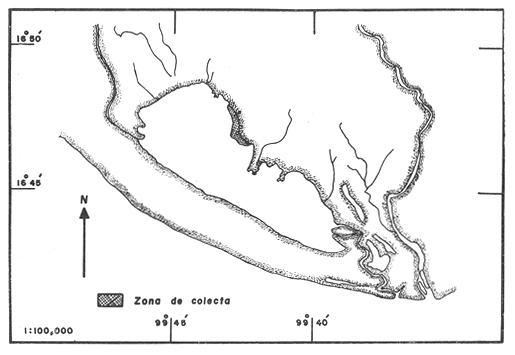 Fig. 2. Zona de colecta en la laguna de Tres Palos. 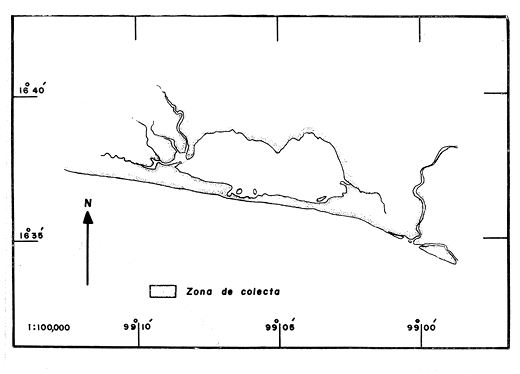 Fig. 3. Zona de colecta en la laguna de Chautengo. 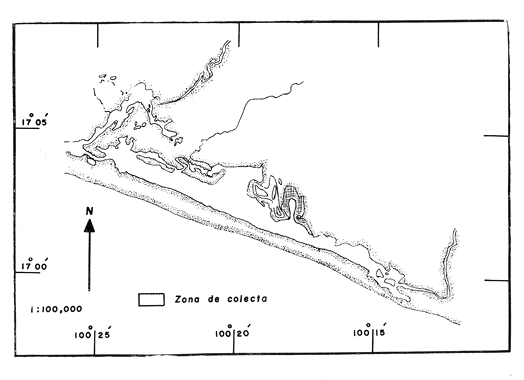 Fig. 4. Zona de colecta en la laguna de Mitla. En la laguna de Chautengo los individuos fueron colectados: a). En fondos arenosos con gran cantidad de restos vegetales a profundidades, de 30-40 cm., en la desembocadura de un arroyo con aguas turbias por el arrastre, en el que se encontraron lirios acuáticos (Elchhornia sp.), tules (Thypha latifolia) y otros vegetales emergidos; y b). En fondos fangosos a la orilla del manglar, en profundidades de 25 - 30 cm., donde se registró una temperatura máxima de 36°C. En esta zona se capturaron la mayor parte de los individuos machos, mientras que en la primera, un elevado porcentaje correspondió a hembras con huevos. Como fauna de acompañamiento se colectaron peneidos juveniles y alfeidos. En la laguna de Mitla, las colectas se llevaron a cabo en profundidades hasta de 1 metro; esta laguna está rodeada de mangles y carrizos (Arundo sp.) y se observaron algunas aves acuáticas como garzas (Bitbttlcits A. ibis), pato buzo (Phalacrocorax auritus) y pelicanos ( Pelecanus occidentalis). MATERIAL Y METODOS
MATERIAL DE ESTUDIOEl presente trabajo fue realizado sobre un total de 1 249 ejemplares provenientes de la laguna de Tres Palos, colectados durante nueve viajes llevados a cabo entre el 29 de julio de 1974 y el 23 de julio de 1975. 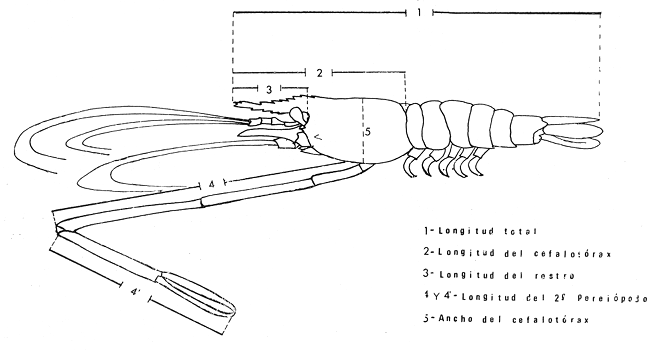 Fig. 5. Esquema de Macrobrachium tenellum que muestra las medidas tomadas. Mediante dos muestreos efectuados en la laguna de Mitla el lo. de agosto y el 2 de septiembre de 1974 se colectaron 400 individuos; y en un viaje efectuado a la laguna de Chautengo el 29 de julio del mismo año, se colectaron 74 ejemplares. De los 1723 individuos, el 74.6% correspondió a machos y el resto a hembras. El ejemplar más grande midió 132 mm., de longitud total, tuvo un peso de 28.6 g., y fue capturado, en la laguna de Mitla. ACTIVIDADES DE CAMPOEl trabajo de campo consistió en colectas diurnas y nocturnas realizadas a bordo de un "cayuco" de aproximadamente 4 m. de eslora; para la captura se utilizaron atarrayas camaroneras de 1.5 y 3 m. de diámetro; también se realizaron colectas a mano durante los meses de mayor abundancia. En el momento de las capturas se registró la temperatura superficial, se midieron profundidades, y se obtuvieron muestras superficiales de agua para determinar la salinidad. Las muestras fueron colocadas en bolsas de polietileno debidamente marcadas y se agregó formaldehído al 10%, para su transporte al laboratorio. ACTIVIDADES DE LABORATORIOLos ejemplares se lavaron con agua corriente durante cinco minutos para eliminar el exceso de formaldehído y se tomaron las siguientes mediciones (Fig. 5) siguiendo los métodos propuestos por Schmitt (1921), Boschi (1969), Carrillo (1968) y Kunju (1968): Longitud total (L.T.), longitud del cefalotórax (L.C.), longitud del rostro, ancho del cefalotárax, longitud del segundo pereiópodo y peso de los individuos. Las hembras ovígeras fueron apartadas y tratadas individualmente. La masa de huevecillos fue separada para su estudio y recuento; los huevecillos se escurrieron y contaron directamente con ayuda de un microscopio de diseción; también se calculó su número por el método volumétrico. Con auxilio de un microscopio Carl Zeiss con objetivo de l0X, ocular de 12.5X y reglilla graduada, se midieron los huevecillos y se observó su estado de desarrollo. RESULTADOS Y DISCUSIÓN
TAXONOMIALos langostinos pertenecen a la Clase: Crustácea Orden: Decapoda Superfamilia: Palaemonidae Familia: Palaemonidae Subfamilia: Palaemoninae Género: Macrobrachium Especie: M. tenellum NOMBRES COMUNESA esta especie de "langostinos" se le conoce en México con los nombres comunes de Acamayas, chacales, piguas, cauques, mayacaxtles, manudas, mayas, huehuentones (Mercado, 1959) o camarones de río. DISTRIBUCIÓN GEOGRÁFICADe acuerdo con Holthuis (1952), la especie vive en aguas dulces y salobres; se conoce su existencia desde Baja California hasta el Norte del Perú. Los estudios realizados señalan su presencia en La Paz, B. C., Los Cocos y Río Presidio, Sin. (Rodríguez de la Cruz, 1968); en las lagunas de Apozahualco, Mitla, Coyuca, Tres Palos y Chautengo, Gro. (Stuardo et al. 1974). Tamnn se reporta en el Rio San Antonio, El Salvador, C. A. (Sánchez, 1975). Todas estas localidades pertenecen a la Provincia Pandmica que comprende desde Isla Cedros, Baja California, hasta Payta, Pero (Keen, 1958, 1971; Olguin, 1968). DIMORFISMO SEXUALEn los Palemónidos existen caracteres morfológicos que hacen posible separar al macho de la hembra a simple vista, cuando se trata de individuos adultos. Según Boschi et al. 1962), se suele dividir a estos caracteres en permanentes y periódicos; entre los primeros se encuentran: a) Pieza suplementaria o appendix masculina en el endopodito del segundo par de pleópodos del macho (Fig. 6). b) Estructura de los coxopoditos del 5o. par de pereiópodos y tamaño de las placas esternaleg de los somitos -12 y 13. c) Crecimiento diferencial del segundo par de pereiópodos y 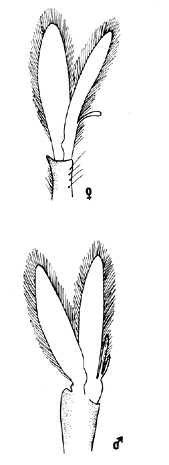 Fig. 6. Diferencias sexuales en lo segundo. pleópodos de M. tenellum. d) Diferencia en la forma y tamaño de las placas tergo-laterales, y desigualdad en el crecimiento del precoxopodito de los pleópodos. CARACTERES SEXUALES PERIÓDICOSDentro de éstos se encuentran: a) Sedas ovígeras no pectinadas que sujetan los huevos durante la época de la incubación. b) Sedas basales no ovígeras y pectinadas que ocupan el margen externo del basipodito en las hembras. c) Sedas basales, externas pectinadas y no pectinadas en el basipodito de los pleópodos lo. al 4o. en las hembras. En Macrobrachium tenellum el macho, alcanza mayor tamaño, que la hembra y es fácilmente distinguible por la longitud de los pereiópodos del segundo par; estos apéndices llegan a tener una longitud mayor incluso, que la longitud total del individuo. Existen ejemplares machos que presentan los segundos pereiópodos de tamaño desigual, generalmente es el pereiópodo izquierdo el de menor tamaño, aunque no siempre sucede así. Este fenómeno ha sido observado por Castro (1966) en ejemplares de Cryphiops caementarius (Molina), quien señalo también que los segundos pereiópodos de las hembras son siempre del mismo tamaño. Esta observación es válida para la especie aquí estudiada. Los, segundos pereiópodos de las hembras no difieren en tamaño y diámetro de las otras patas en el mismo ejemplar. En los individuos machos, a fin teniendo los segundos pereiópodos del mismo, tamaño, que los demás, son más robustos y conspicuos que en las hembras. Los machos adultos de Macrobrachium tenellum presentan el cefalotórax más ancho, que las hembras en relación con él resto del cuerpo. Ling (.1969), señaló que las hembras de Macrobachium rosembergii tienen una espaciosa cámara incubadora bajo el abdomen, formada por la prolongación de las pleuras abdominales y que los poros genitales de las hembras se encuentran en la base de las patas del quinto par; esta observación es válida para Macrobrachium tenellum y todos los palemónidos. ESTRUCTURA Y DINÁMICA DE LA POBLACIÓNAbundancia. La interpretación de las variaciones de la abundancia de la especie, observadas en el período de muestreos, se hizo sobre la base de los datos de la laguna de Tres Palos, principalmente. En el presente estudio se comprobó la abundancia de la especie en el mes de julio, cuando la muestra fue de 137 individuos, hecho que se debió a causas ajenas al azar. En el siguiente mes se procesó un máximo de 200 ejemplares; la abundancia en los meses siguientes continuó con valores altos, ya que en octubre se pudo obtener nuevamente una muestra de 200 individuos; la abundancia disminuyó considerablemente en el mes de diciembre y probablemente en noviembre en que se colectaron sólo 52 ejemplares. La falta de muestreos en los meses de enero y febrero no permite precisar con certeza la abundancia de la especie, pero aparentemente continuó baja; en el mes de marzo en que continuaron los muestreos, se pudo obtener nuevamente una muestra de 200 ejemplares aunque con mayor dificultad que en los casos anteriores. La duración de la pesquería se extiende aparentemente a gran parte del año, con una disminución considerable del recurso en los meses de invierno, a mediados del periodo de secas. Mientras no se establezcan con exactitud aspectos generales de la biología de la especie, como la duración del periodo de reproducción, la variación de las poblaciones, y movimientos migratorios dentro de la laguna y/o hacia los tributarios vecinos, no será posible predecir con certeza las fluctuaciones de la pesquería. Resulta interesante observar la relación que existe entre la disminución de la salinidad y la temperatura con la abundancia del recurso (Fig. 7). Se observó que de julio de 1974 en adelante, la salinidad disminuyó hasta un valor de 1.2%, en el mes de octubre, lo que coincidió con el incremento de individuos en vías de reproducción. El segundo aumento poblacional sé registró entre marzo y junio de 1975, periodo que corresponde- al reclutamiento de juveniles y adultos jóvenes, respectivamente. Parece ser que este aumento en la población es previo a la emigración de los adultos de mayor tamaño en los fondos someros descubiertos; este hecho, combinado con el crecimiento de los adultos jóvenes, determina el auge de la pesquería entre los meses de julio, agosto, septiembre y octubre de cada año. 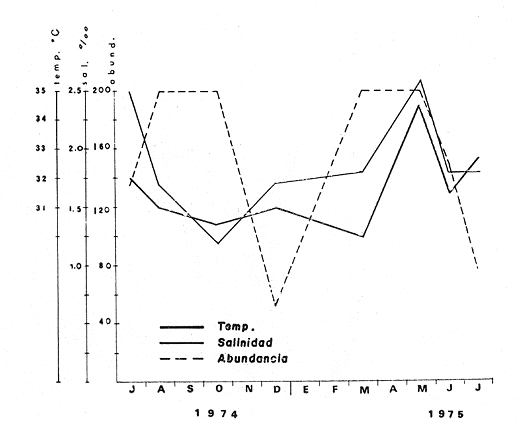 Fig. 7. Relación entre la temperatura, salinidad y abundancia de langostino en la laguna de Tres Palos. Tallas. El resultado de las distintas mediciones sobre el total de la población y separadamente en machos y hembras de todas las lagunas, se presenta en las figuras 8 a 12. En cada caso los valores de los muestreos mensuales se graficaron sin suavizar con objeto de presentar las características reales de la población. Longitud total. La figura 8a. ilustra el comportamiento de la población en base al total de muestreos efectuados. En general, las curvas reflejan un comportamiento que en un ciclo anual corresponde a grupos de edad que se reemplazan cíclicamente. En la muestra del 31 de julio de 1974, la moda, comprendida entre las clases 105-110- mm, representa el 23%, mientras que el 74.42% corresponde a valores comprendidos entre 90 y 100 mm de L. T. El rango de tamaños varió entre 42 y 124 mm, esta última medida, no volvió a registrarse en ninguno de los muestreos posteriores de esta laguna. Es importante señalar en este muestreo, la presencia de individuos juveniles y ejemplares inmaduros de menos de 50 mm de longitud total. 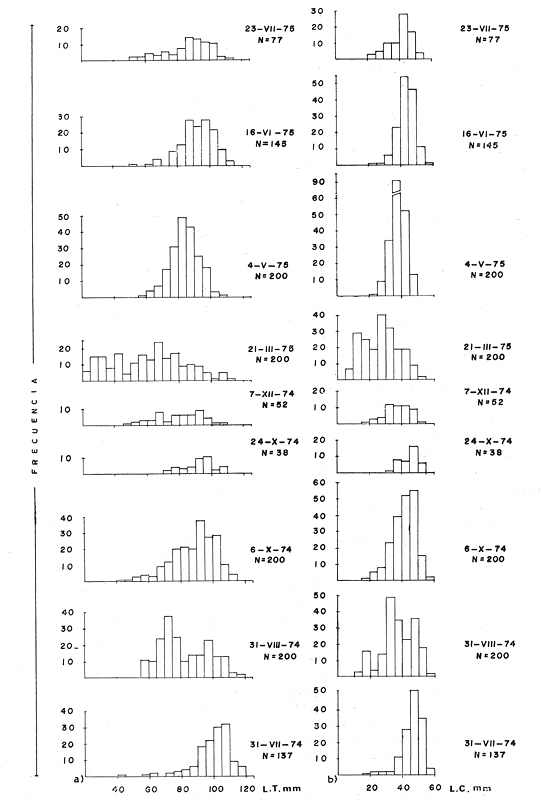 Fig. 8. Polígonos de frecuencia, de la longitud total (L.T.) (a) y longitud del cefalotórax (L.C.) (b) Laguna de Tres Palos. La gráfica correspondiente al segundo muestreo es bimodal con rangos máximos que varían entre 70-75 y 95-100 mm. y representan el 19 y 11%, respectivamente. El 97.5%, de la muestra corresponde a las clases comprendidas entre los límites de 55 a 110 mm., mientras que el rango de tamaños fue menor que en el mes anterior y varió entre 56 y 116 mm. La presencia de dos valores modales el mismo muestreo, sugiere la existencia de grupos diferentes, debido al ingreso a la población de ejemplares de menor tamaño. Es claro que la longitud de los individuos que forman las clases menores de los meses anteriores, han aumentado de tamaño desplazando los valores modales a las clases más altas. Paralelamente se registró el ingreso de individuos juveniles de las clases 40-45 y 45-50 mm., como resultado del desove del mes de julio y subsiguientes. La gráfica del tercer muestreo presenta la moda comprendida en la clave. 90-95 mm.; la clase modal representa el 18.5%, de la muestra total. Aquí el 89.5%, se encuentra entre los límites de clase de 70 a 110 mm mientras que el total de la población varió entre 45 y115 mm. Es claro que la longitud de los individuos que forman las clases menores de los meses anteriores, han aumentado de tamaño desplazando los valores modales a las clases más altas. Paralelamente se registró el ingreso de individuos juveniles de las clases 40-45 y 45-50 mm., como resultado del desove del mes de julio y subsiguientes. La muestra del 24 de octubre, pese al reducido número de ejemplares, refleja el rango de tamaños de los porcentajes predominantes dos semanas antes. La gráfica del quinto muestreo presenta frecuencias análogas. La moda está comprendida en la clase 90-95 mm. y representa el 17.3% de la muestra total; el rango varía de 46 a 107 mm. En este muestreo se observó nuevamente la presencia de ejemplares juveniles y, en general, una heterogeneidad de tamaños que podría ser representativa de los siguientes meses y que se acentúa en el mes de marzo. La disminución de la abundancia de las clases más altas, está relacionada con la pesca excesiva de los meses anteriores y con el desplazamiento de los adultos. En la curva del mes de marzo, la moda está comprendida en la clase de 65-70 mm. de longitud y representa el 12% del total; las clases restantes, sin embargo, no presentan porcentajes significativamente más bajos que los registrados en las últimas clases; la gráfica presenta una amplia variación de tamaños que van desde 20 hasta 112 mm. En este caso la colecta se llevó a cabo en las primeras horas de la noche y se registró un alto porcentaje de individuos juveniles, de tallas no registradas en los muestreos anteriores. En el séptimo muestreo el rango de tamaños se redujo considerablemente con la desaparición de las clases juveniles. La moda representa el 24.5% de la muestra y se encuentra en la clase 80-85 mm, mientras que el 91.5% se agrupó entre los límites de 70 y 100 mm. El rango de tamaños de 56 a 106 mm, se redujo considerablemente respecto del mes de marzo; el desplazamiento de la moda del muestreo anterior (6570 mm.), a la clase 80-85 mm, sugiere un incremento en la talla de los individuos más jóvenes o migraciones a lugares dulceacuícolas o zonas más someras. En la penúltima muestra se registraron 2 valores modales en las clases de 85-90 y 95-100 mm., que representaron el 19.3% de la muestra total; indudablemente ambos corresponden a un solo valor, ya que el 85.9% del total se encuentra ubicado entre los límites de clase de 80 y 110 mm. En este caso, vuelve a observarse el desplazamiento de los valores altos de la población, lo que significa un aumento en la talla de los individuos o el ingreso de adultos en época de reproducción. Durante este mes se observó un reclutamiento escaso de juveniles, probablemente como remanentes de los desoves anteriores. El último muestreo que se realizó en el mes de julio de 1975, presenta un rango y características similares a las del mes anterior y primer muestreo (julio de 1974). Es posible que la similitud respecto de este último fuera mayor si la muestra hubiera tenido por lo menos, el doble de los ejemplares procesados. Longitud del cefalotórax. Con objeto de constatar si las variaciones observadas al agrupar a los ejemplares de acuerdo a su longitud total corresponden a los resultados de la longitud del cefalotórax, se han representado en la misma figura los polígonos correspondientes. En primer lugar, se constató que las curvas construidas con estos datos (Fig. 8b) siguen prácticamente a las de la longitud total, pero el rango se reduce considerablemente con la consiguiente simplificación de las gráficas. La aparición de juveniles fue observable en la misma época que en el caso de la longitud total, excepto en el mes de agosto en que aparece un 9.5% de juveniles al ser considerada la longitud total. Este error se debe indudablemente a causas diferentes al azar. No hubo ningún ejemplar que sobrepasara los 68 mm. de L.C., y en general los valores modales variaron en los distintos meses entre las clases 30-35 y 45-50 mm. Longitud del cefalotórax en machos y hembras. Los polígonos de frecuencia de la longitud del cefalotórax en machos y hembras, señalan una marcada diferencia debido al mayor número de machos en todas las muestras. De la serie de polígonos de las figuras 9a y 9b, se infiere el ciclo biológico de la especie en cuestión. El valor modal del primer muestreo (julio de 1974), que se encuentra entre 45 y 50 mm de L.C., se sostiene durante los meses siguientes (agosto, septiembre y octubre) que se consideran los de mayor abundancia, pero se desplaza negativamente como consecuencia de la desaparición de individuos de talla comercial en la zona a principios del invierno. Esta situación se prolonga hasta el mes de marzo aproximadamente, en que se registra un alto porcentaje de individuos juveniles; a partir de este mes, los individuos incrementan su tamaño hasta alcanzar nuevamente en el mes de junio, la talla comercial señalada con anterioridad. Pesos. Los valores de peso en gramos (Fig. 10) registrados en los distintos muestreos, presentan variaciones que corresponden en gran medida al tamaño de los individuos; sin embargo, no puede asegurarse que exista correlación entre el peso y la longitud. En el polígono del mes de julio de 1974 se observan las características de una curva normal; la moda se encuentra entre las clases de 10-14 g representando el 48.1% de la muestra total; el rango varió de 0.5 a 21.2 g. En el mes de agosto los valores modales se han desplazado negativamente hasta la clase que representa el 36% de la muestra; el rango total varió entre 1.8 y 20.7 g. La predominancia de valores de peso bajo se debe al ingreso a la población de individuos jóvenes que no han alcanzado su talla y peso definitivos. En el mes de octubre se observan dos modas comprendidas entre los 4-6 y 10-12 g y representa cada una el 20.5% del total. El muestreo correspondiente al mes de diciembre presenta un histograma similar al anterior. En el mes de marzo de 1975, la moda se encuentra en la clase 0. 1 - 3.0 g; esta observación se interpreta como el resultado del reclutamiento de individuos juveniles a la población, llegando a representar el 44.5% de la muestra total. En el hecho, el 91.5% se encontró entre los límites de clase de 0. 1 y 8.0 g siendo valores muy inferiores a los encontrados durante los meses de julio, agosto y octubre de 1974. En los meses de mayo, junio y julio de 1975, las modas y porcentajes predominantes de la población se desplazaron hasta alcanzar una estructura ligeramente similar a la de julio de 1974. El análisis anterior nos hace suponer que se llevan a cabo ingresos masivos a la población de juveniles y adultos jóvenes durante la época de engorda. El peso de los juveniles abarca generalmente sólo las clases de 0. 1 a 2.0 g. CRECIMIENTOSin información y observaciones en condiciones controladas sobre el desarrollo larvario de M. tenellum, no se puede dar una interpretación adecuada del crecimiento, aunque es de interés intentarlo utilizando el conocimiento alcanzado en otras especies. Los datos más conocidos corresponden a M. rosembergii, uno de los langostinos de mayor tamaño registrado. En esta especie, los huevos eclosionan después de aproximadamente 19 días de incubación (Bardach et al., 1972; Wickins y Beard, 1974) pasando por 12 estados larvales de los 35 a los 55 días siguientes; después de este tiempo, se transforman en juveniles que miden entre 5 y 6 mm de longitud total; éstos tienen movimientos migratorios 2 o 3 meses después hasta alcanzar las aguas dulces. 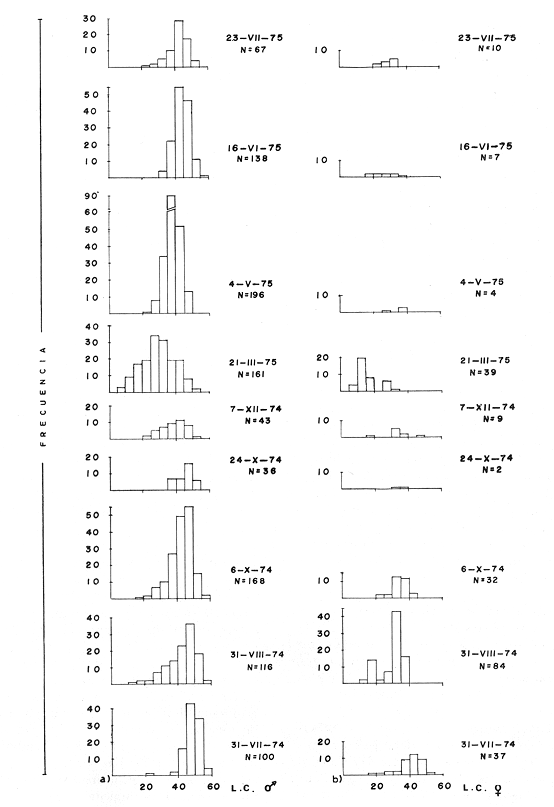 Fig. 9. Polígonos de frecuencia de la longitud del cefalotórax (L.C.) en machos (a) y hembras (b). Laguna de Tres Palos. 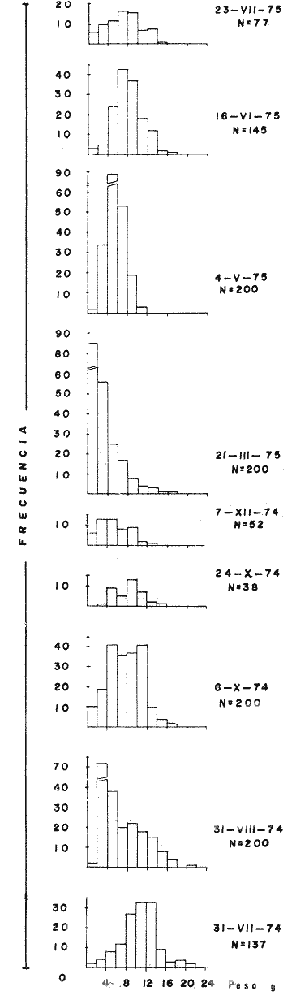 Fig. 10. Polígonos de frecuencia del peso de las muestras de la laguna de Tres Palos. En Macrobrachium americanum los juveniles de metamorfosis reciente, después de 67 días de vida larvaria, medían entre 7 y 9 mm de longitud total y es probable que exista la misma relación de crecimiento en M. tenellum. Rajyalakshmi y Ranadhir (1969) reportaron el ingreso de juveniles de Macrobrachium malcomsonii a la pesquería en tamaños de 8 a 30 mm y en el periodo de pesca alcanzaron hasta 80 mm de longitud. La talla máxima de M. malcomsonii es de 180 mm con tallas promedio que van de 81 a 100 m de longitud total. En el caso, de M. tenellum el tamaño máximo fue de 132 mm de longitud y se registró en la laguna de Mitla; mientras que el tamaño mínimo de ingreso a la población, en la laguna de Tres Palos, fue de 20 mm de longitud total durante el mes de marzo, que como se ha mencionado corresponde al periodo de reclutamiento. Un aspecto complementario de importancia en la determinación del crecimiento de los langostinos juveniles y adultos, es el número de mudas que un ejemplar puede experimentar en un periodo anual, aunque este fenómeno está limitado por las condiciones ambientales predominantes. Bardach et al. (op. cit.) han señalado que los juveniles de M. rosembergii mudan cada 4 a 6 días, pero la frecuencia depende de la edad de los individuos y de la cantidad y calidad del alimento que ingieren. Las hembras sexualmente maduras mudan antes del apareamiento. En condiciones de laboratorio, Ling (1969) observó que las hembras maduras ponen huevos hasta dos veces en cinco meses, lo que sugiere que pueden desovar 4 o 5 veces durante un año en condiciones naturales. Arana (1968) señaló que la frecuencia de mudas en hembras de la especie estudiada por él fue de 40 a 45 días y mencionó el caso de hembras con huevos que después de la eclosión mudan y son fecundadas nuevamente. En Macrobrachium tenellum no se han efectuado observaciones sobre este aspecto. PROPORCION DE SEXOSLas observaciones realizadas al respecto señalan gran diferencia en la proporción de sexos durante el periodo de estudio, en el que predominaron en todos los casos los especímenes machos (Fig. 13; tablas 1 y 2). Boschi et al. (1969) han señalado para Artemesia longinaris Bate, que el hecho de que predomine uno de los sexos sobre el otro se debe a la selección que ejercen las redes de captura sobre los individuos de talla mayor, que en el caso de M. tenellum son los machos. Sin embargo, esta observación no es válida en el presente caso, ya que si bien las dos primeras muestras fueron tomadas de capturas comerciales, las siguientes fueron obtenidas directamente a mano o con atarraya de menor abertura para evitar el error. Los machos son los individuos que alcanzan las mayores tallas y los muestreos realizados sin implementos selectivos como la atarraya camaronera, señalan en todos los casos, la predominancia de individuos del sexo masculino. 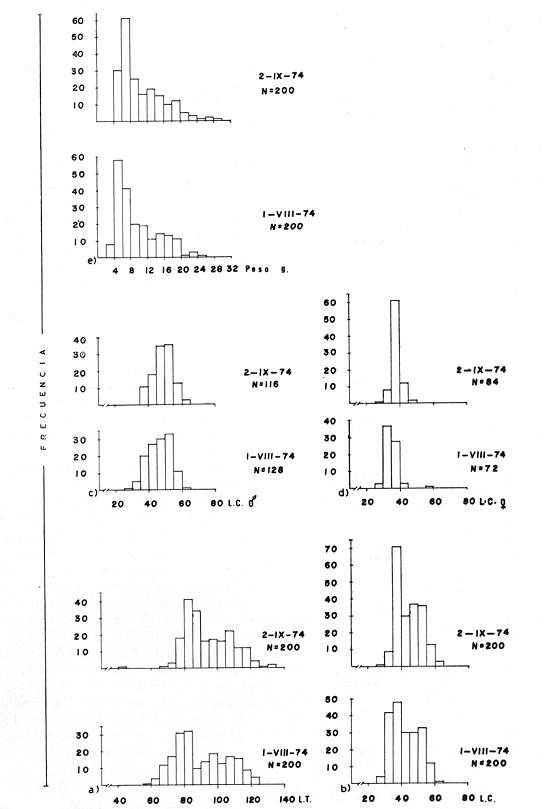 Fig. 11. polígonos de frecuencia de L.T., L.C. de las muestras totales, en machos y hembras y peso de las muestras de la laguna de Mitla. 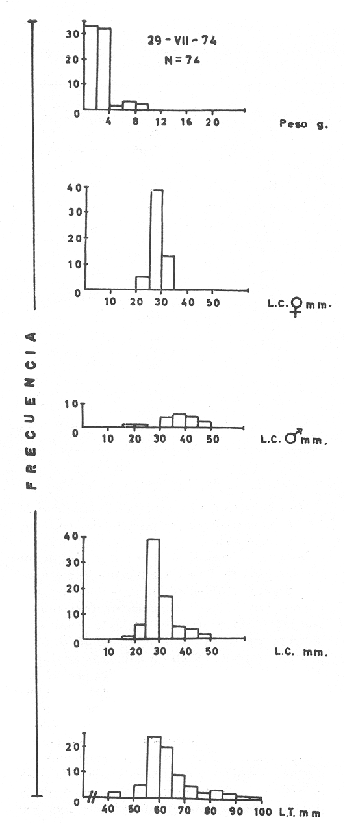 Fig. 12. Polígonos de frecuencia de L.T., UG. y peso de la muestra de la laguna de Chautengo. La disparidad en la proporción de machos y hembras se acentuó durante los meses de mayo, junio y julio en que los individuos alcanzan el mayor desarrollo. La proporción aproximada a 1:1 se observó a fines del mes de agosto, justamente cuando se eleva el número de hembras ovígeras. Sin embargo, en el siguiente muestreo (octubre), se registró nuevamente marcada diferencia en la proporción de sexos, que se mantuvo más o menos constante hasta el mes de marzo del siguiente año. 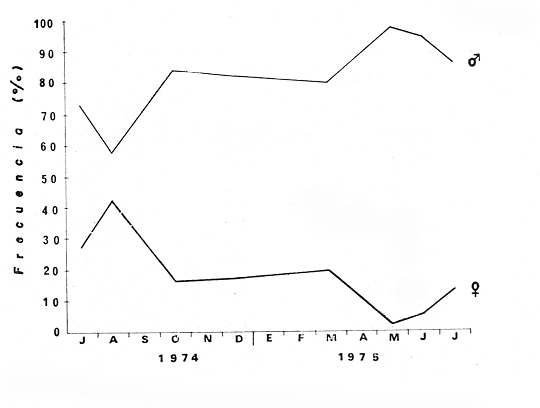 Fig. 13. Frecuencia de sexos de las muestras de la laguna de Tres Palos. 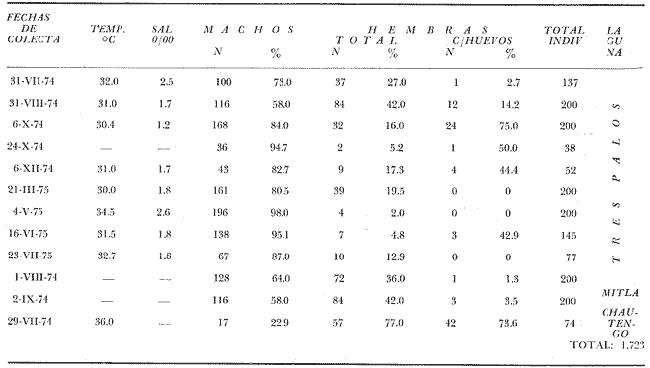 TABLA 1 MUESTRAS DE MACROBRACHIUM TENELLUM INDICANDO LOS TOTALES CAPTURADOS, RELACIÓN DE SEXOS, FECHAS DE CAPTURA Y VALORES DE TEMPERATURA Y SÁLINIDAD 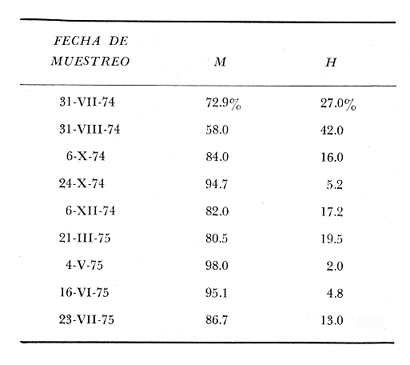 TABLA 2 PROPORCIÓN DE SEXOS DE MACROBRACHIUM TENELLUM EXPRESADA EN PORCIENTO, DE LAS MUESTRAS PROVENIENTES DE LA LAGUNA DE TRES PALOS REPRODUCCIÓNBajo este aspecto es importante considerar la madurez, fecundidad, el periodo de reproducción y su duración. Madurez y fecundidad. Los huevecillos de M. tenellum son de forma ligeramente elíptica; en las muestras estudiadas variaron de 443 a 617 micras. Los huevecillos se encuentran unidos por medio de secreciones de las glándulas de cemento que se localizan en la base de los pleopodos y urópodos (Rodríguez de la Cruz, 1968). Al hacer la medición de huevecillos bajo el microscopio, se observó que están unidos por medio de finos ligamentos que hacen posible separar la freza casi completa de los pleópodos de la hembra sin que los huevos se desgranen. La freza se encuentra dispuesta en la región abdominal de la hembra y fuertemente adherida a las setas de los pleópodos que, la cubren, desde la primera hasta la cuarta pleura; en ninguna ocasión se encontró más atrás. En los huevecillos de. las hembras colectadas se observaron diferentes estados de desarrollo, desde aquéllos con el vitelo uniformemente distribuido, hasta los que presentaron un embrión bien desarrollado con ojos perfectamente visibles. 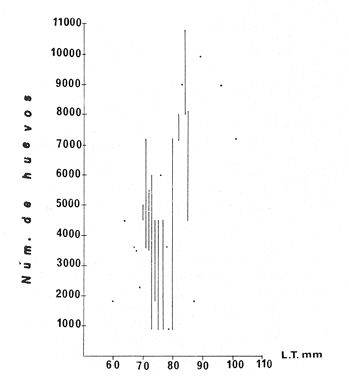 Fig. 14. Fecundidad de M. tenellum de acuerdo al tamaño de las hembras. 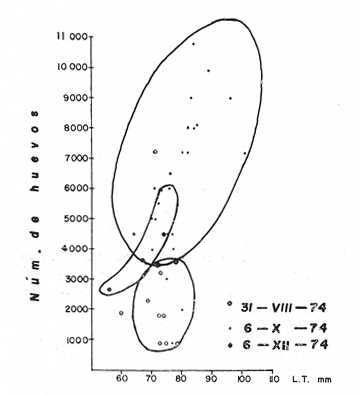 Fig. 15. Fecundidad de hembras de M. tenellum del 31- VIII-74; 6-X-74 y 6-XII-74 en la laguna de Tres Palos. 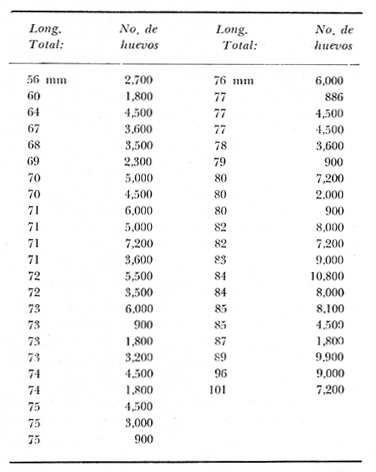 TABLA 3 RELACIÓN ENTRE LA LONGITUD DE LAS HEMBRAS Y EL NÚMERO DE HUEVOS DE LAS MUESTRAS DE LA LAGUNA DE TRES PALOS. Los huevecillos frescos son de color amarillo claro brillante en los primeros estadios; más o menos anaranjados en estado intermedio; verde oliváceo cuando están avanzados en su desarrollo y casi gris cuando están listos para eclosionar. Los primeros estadios larvales ocurren dentro del huevo como característica de los Carídeos de suprimir las formas larvales libres (Rodríguez de la Cruz, 1968; Castro, 1966). El número de huevecillos de M. tenellum no está en relación con la longitud total de las hembras, sino que existe amplia variación con rangos que fluctúan entre 900 y 10 800 unidades (Fig. 14; Tabla 3); aún en individuos de la misma talla, el número de huevos varía considerablemente. En la figura 14 se observa que el rango de 900 a 6 000 unidades está sostenido principalmente por hembras que fluctúan entre 70 y 80 mm de longitud total. El número, de huevos varió de acuerdo a las fechas de colecta registrándose en el mes de octubre el mayor número de unidades por hembra (Fig. 15). El tamaño de las hembras ovígeras fue de 56 a 101 mm de longitud total para la laguna de Tres Palos y de 51 a 69 mm para la laguna de Chautengo. Las hembras recién desovadas se diferencian fácilmente de las otras porque presentan el abdomen más ancho y las pleuras del primero al cuarto segmentos abdominales extendidas a manera de cuchara proporcionando mayor volumen a la región ventral. Periodo y duración de la reproducción. Las hembras con huevos fueron capturadas durante el periodo de junio a diciembre, registrándose el porcentaje. Más elevado durante el mes de octubre con el 75%, del total de las hembras colectadas (Fig. 16; tabla l). Existe un periodo de reproducción bien definido que coincide con la baja de salinidad y la abundancia de detritus acarreado por los afluentes de agua dulce. Sin embargo, la presencia de juveniles y la diversidad de tallas registradas en la mayor parte de los muestreos, sugiere que la especie es capaz de reproducirse a lo largo de gran parte del año. Como se discutió en el inciso de abundancia, existe un periodo en que la disminución de la salinidad y de la temperatura determina la iniciación del perecido reproductivo y en, especial, los movimientos migratorios de los adultos hacia las áreas de reproducción. 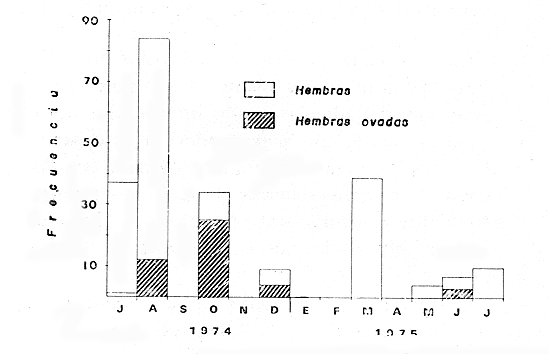 Fig. 16. Total de hembras capturadas en, el periodo de julio de 1974 a julio de 1975, en la laguna de Tres Palos frecuencia de hembras ovadas. 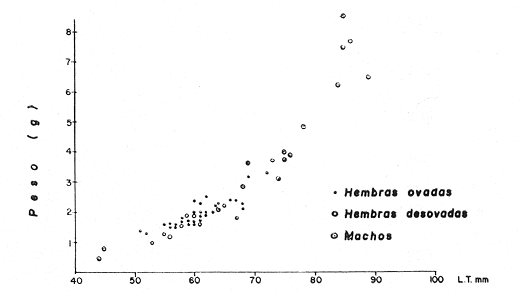 Fig. 17. Correlación entre la L.T. y peso de la muestra de la laguna de Chautengo. Respecto de esta migración, se ha sugerido que presenta dos modalidades de acuerdo a las condiciones de la laguna, si se abre al mar o presenta características dulceacuícolas constantes. El primer caso corresponde a la laguna de Chautengo en que existe una población pobre de langostino durante eln periodo de lluvias y los adultos bajan por los ríos Nexpa y Copala a reproducirse; las formas larvales permanecen en la zona hasta alcanzar las etapas juveniles y de adultos jóvenes. Cuando la laguna abre su boca al mar y aumenta la salinidad en todos los sectores, tanto juveniles como adultos jóvenes remontan aguas arriba. Comparativamente, en esta laguna los langostinos no alcanzan las tallas registradas en las otras, probablemente debido a los altos gradientes de salinidad; es en esta laguna donde las hembras comienzan a reproducirse a más temprana edad y donde se registraron las longitudes más bajas de machos adultos (Fig. 17). En el caso de las lagunas con salinidades bajas durante todo el año como Tres Palos y Mitla, las poblaciones de langostino permanecen en ellas durante todo el año. Esta afirmación se desprende de los muestreos efectuados en Tres Palos, donde ocurren variaciones que están ligadas directamente al periodo de reproducción. En consecuencia, la aparición y predominancia de juveniles en una época, y de adultos en otras demostrada por la presencia de hembras con huevos, indica una alternativa que está regida por un ritmo biológico interno. Los pescadores locales mencionan que los langostinos emigran hacia las aguas profundas durante los meses de menor abundancia y salen en lugares someros a desovar del mes de julio en adelante. Si efectivamente ocurre así, deberá demostrarse con capturas en distintas zonas y mediante el empleo de diversas artes de pesca, pero en especial, por el método de marca recaptura de individuos adultos. No se descarta la posibilidad de que al terminar periodo de reproducción, los adultos busquen protección entre la vegetación de las orillas de la laguna, por lo menos en aquellas zonas que no sufren las consecuencias de la desecación; entre los trabajos consultados, no hay estudios en ambientes salobres similares. EPIZOOS Y PARÁSITOSEn un alto porcentaje de individuos adultos se encontraron unas pequeñas "puestas" de consistencia coriácea, que se presentan como cadenas dobles o triples sobre el exoesqueleto del animal, estos organismos no fueron identificados pero presumiblemente corresponden a huevecillos de insectos acuáticos. Entre los parásitos descritos de Macrobrachium existen algunos isópodos del género Probopyrus, cuya distribución ha sido delimitada por Richardson (1905). Entre las especies conocidas se encuentran P. oviformis, P. panamensis, P. bithynls y P. papaloapanensis, la última descrita por Rioja (1948) de la vertiente del Golfo de México. Este tipo de parásitos se registró ocasionalmente en las muestras de laneostino obtenidas de la laguna de Tres Palos y serán descritos en una publicación por separado. El género Probopyrus no parece ser exclusivo de Macrobrachium sino que también se encuentra sobre otros palemónidos. DEPREDACIÓNAunque no se realizaron en este trabajo, observaciones especiales sobre este aspecto, en la literatura se mencionan como depredadores de los langostinos algunas aves, peces, anfibios y ocasionalmente mamíferos (Hartmann, 1958; Hernández, 1960; Castro, 1966; Bahamonde y Vila, 1971). Las observaciones realizadas sobre depredadores de langostino en las costas de Guerrero son muy generales. En el estudio llevado a cabo por Ramírez Granados (1952) en la laguna de Tres Palos, se mencionó la presencia de restos de crustáceos en el contenido estomacal de peces de las familias Mugilidae, Eleotridae y Gerridae, pero no se identificó la especie de los crustáceos; sin embargo, es probable que tales restos pertenecieran a Macrobrachium tenelum Recientemente Yañez- Arancibia et al. (1976) han reportado restos de Macrobrachium tenellum en el contenido estomacal de Galeichthys caerulescens,, un bagre comúnmente- conocido como "cuatete". Respecto a las aves se cita a las gaviotas "gris" y "blanca" (Larus modestus y L. dominicanus), al "huaco" (Nycticorax nycticorax), las garzas "grande" y "chica" (Ardea egretta y A. candidisima) y al "Martín pescador" (Chloroceryle americana) como depredadores de los langostinos. Bahamonde y Vila mencionan la depredación sobre Cryphiops caementarius de Casmerodius albus egretta, Leucophoyx thula thula, Anas cyanoptera y Phalacrocorax auritus entre las aves; y a la "ranagrande" Calyptocephalella sp entre los anfibios. Algunas de las especies mencionadas, principalmente peces y aves, habitan actualmente en las lagunas del Estado de Guerrero, y probablemente ejercen su acción depredadora sobre Macrobrachium tenellum. ConclusionesEl análisis de los resultados permite obtener algunas conclusiones relacionadas con la biología y ecología de la especie. 1. Macrobracium tenellum no presenta variabilidad en cuanto a las características morfológicas de los individuos adultos, a excepción de la longitud del segundo periodo. 2. M. Tenellum alcanza hasta 132 mm de longitud total, 63 mm de longitud cefalotórax y 28.6 g de peso como valores máximos. 3. Comparativamente los individuos machos alcanzan mayores tallas que las hembras en el mismo periodo de crecimiento. 4. La proporción de sexos está siempre a favor de los machos; a excepción de la laguna de Chautengo, en la que el número de hembras fue mayor que el de los machos, en ninguna ocasión se alcanzó la proporción 1: 1. 5. Macrobrachium tenellum se reproduce durante el período de salinidad más baja en áreas de poca profundidad y es probable que existan migraciones relacionadas con esta función. 6. La mayoría de las hembras ovígeras caen dentro del rango de 70-80 mm de longitud total, con un número de huevecillos que fluctúa entre 900 y 6 000 unidades, observación que permite señalar la fecundidad de la especie. 7. El número de huevecillos de Macrobrachizím tenellum no está en relación con el tamaño de las hembras. 8. El aumento de la población coincide con la disminución de la salinidad en la laguna de Tres Palos y probablemente en las otras lagunas. 9. La duración de las pesquerías en las lagunas de Mitla y Tres Palos abarca desde el mes de junio hasta los meses de septiembre y octubre, periodo que coincide con los meses de mayor abundancia y talla de los langostinos. 10. El periodo de reclutamiento de los individuos jóvenes a la población abarca los meses de invierno (diciembre, enero, febrero) y parte de la primavera (marzo y principios de abril). 11. Macrobrachium tenellum alcanza los valores máximos de peso y longitud en la laguna de Mitla; valores medíos en la laguna de Tres Palos y mínimos en la laguna de Chautengo, lo que parece estar en relación directa con los gradientes de salinidad observados. AgradecimientosA José Stuardo B. por su valiosa ayuda en la elaboración de este trabajo, a Enrique Boschi por sus, útiles sugerencias y revisión final, y a Alfredo Martínez Guerrero por su constante apoyo. Este estudio fue financiado parcialmente, por Contrato de Estudios No. OC-E-03-74, celebrado entre la Comisión del Río Balsas, de la S.R.H. y el Centro de Ciencias del Mar y Limnología de la UNAM. LITERATURAARANA, M. F., Experiencias sobre el cultivo del langostino Macrobrachititpi americanum Bate en el noroeste de México. Sinip,. FAO/CARPAS Acuacult. Amér. Lat. CARPAS/6/74/SE/ 1974. 1-9. 19: BAHAMONDE, N. e I. VILA, Biol. Pesq. Sinópsis sobre. la biología del camarón de río del Norte. Chile. 1971, 3-60. (5): BARDACH, J. E., J. H. RYTHER Y 0. W. Mc. LANEY, Aquaculture. The farming and husbandry of freshwater and marine organisms. Inc John Wiley and Sons. . Nueva York. 1972. 632 p. BOSCHI E. E. y V. ANGELESCU, Descripción de la morfología externa e interna, del langostino con algas aplicaciones de índole taxonómica y biológica. Bol. Inst. Biol. Marina, Mar del Plata, 1962. 1-73, 1: BOSCHI, E. E. Crecimiento, migración y ecología del camarón comercial Artemesia longinarís Bate. FAO Físh. Rep., 1969. 833-846. (57): 3 CARRILLO V. F., Fish. Rep., Morfología. de Macrobrachíúm acanthurus (Wiegmanú) en el Estado de Veracruz, México. FAO 1968. .415-425. (57): 2 CASTELLANOS, L., Convenio Comisión del Río Balsas, Sub programa de Hidrología. Informe Final de la 2a. Etapa del Programa de Uso de la Zona Costera de Michoacán y Guerrero. S. R. H. y Centro de Ciencias del Mar y Lininología, UNAM México, 1975. 44 p., 63 tablas, 46 figs. CASTRO, C., Est. Qceanol, El camarón de río del Norte Cryphiops caementarius (Molina). Chile, 1966. 11-19. 2: HARTMANN, G., Pesca y Caza. Apuntes sobre la biología del camarón de río Cryphiops caementarius (Molina). Palaemonidae, Decapoda. Lima 1958. 17-28. 8: HERNÁNDEZ, J. E., Pesca y Caza. Contribución al conocimiento del camarón de río Cryphiops caementarius (Molina). Lima. 1960. 84-106., 10: HOLTHUIS, L. B., The subfamily Palaemoninae. A general revision of the Palaemonidac (Crustacea, Decapada, Natantia) of the Americas. II. Allan Hancock Foundation Pub. Occ. Pap. 1952. 1-396. 12: KEEN, A. M., Sea Shells of Tropical West America. Stanford Univ. Press., U.S.A., 1958b. 624 págs. 1 Sea Shells of Tropical West America. Stanford Univ. Press., U.S.A., 1971. 1064 págs. KUNJU, M., Fish. Rep., Some aspects of the biology of Solenocera indica Nataraj. FAO 1968. 467-485. (57): 2 LING, S. W., Fish. Rep. The general biology and development of Macrobrachium rosembergii (De Man). FAO 1969. 589-606. (57): 3 MERCADO, P., Bol. Piscic. Rural S.I.C., Proyecto para una estación rústica dedicada al cultivo de los langostinos. México, 1959. 6-9. (5-6): 9 OLGUÍN, P. M., Fish. Rep., Estudio de la biología del camarón café Penaeus californiensis Holmes FAO 1968. 331-356. (57): 2 RAJYALAKSHMI, T. y M. RANADHIR, Fish. Rep., The commercial prawn Macrobrachium malcomsonti (H. Milne Edwards) of the river Godavary, a discussion on the trend and caracteristics of the population during 1963-1966. FAO 1969. 903-921. (57): 3 RAMIREZ, G. R., Rev. Soc. Mex. Hits. Nat. Estudio ecológico preliminar de las lagunas costeras cercanas a Acapulco, Gro. 1952. 199-218. 13: RICHARDSON, H., Bull. A menograph on the isopods ofNorth America. U.S. Nat. Mus. 1905. 1-727. 54: RIOJA, E., An, Inst. Biol. Univ. Nal. Autón. México, Un nuevo, crustáceo isópodo parásito de la familia de los Bopíridos del Papaloapan. 1968. 169-174. (19): 1 RODRÍGUEZ DE LA C., M. C., An. Inst. Nal. Invest. Biol. Pesq. Contribución al conocimiento de los Palemónidosde México. II Palemónidos del Atlántico y Vertice Oriental de México con descripción de dos especies nuevas. S.I.C. México, 71-112 1: Fish. Rep., Contribución al conocimiento, de los Palemónidos de México. III. Palemónidos del Golfo de California, con notas sobre la biología de Macrobrachitim aniericanum Bate. FAO 1968. 373-380. (57): 2 SÁNCHEZ. C., Minist. Agric, y Gan. Desarrollo de juveniles del camarón de río Macrobrachium tenellum (Smith) en estanques de arcilla y concreto. El Salvador, C. A. Inf. Téc. 1975. 1-15. (2): 2 SCHMITT, W. L., The marine decapod crustacea of California. Univ. Calif. Publ. Zool., 1921. 1-470. 23: Informe de la Comisión Interdisciplinaria a la laguna de hautengo, S.R H., Gro. Sría. Rec. Hidr. Dir. Acuacult. Méx. 1975. STUARDO, J., A. MARTÍNEZ, A. YÁÑEZ-ARANCIBIA J. A. WEINBORN y J. Ruiz, An. Centro Cienc. Del Mar y Limnol. Univ. Nal. Autón, México. Informe mimeografiado. Prospe,cción de los recurSos biológicos, y pesqueros del sistema lagunar de Guerrero y, en parte, del litoral rocoso de Michoacán. 1974. 500 p. WICKINS, J. F. y T. W. BEARD. Aquaculture. Observations on the breeding and growth of the giant freshwater prawn Macrobrachium rosembergli (De Man) in the laboratry. 1974. 159-174. 3: YÁÑEZ-ARANCIBIA, A., J. CURIEL-GOMEZ y V. LEYTON-VARGAS, An. Centro Cienc del Mar y Limnol. Univ. Nal. Autón. México. Prospección Wológica y ecológica del bagre marino Galeiclithys, caerulscens (Guntherg) en el sistema lagunar costero de Guerrero, México (Pasces: Arfidae). 1976. 125-180. (1): 3
|