|
ANÁLISIS DE LA COMUNIDAD DE UNA LAGUNA COSTERA EN LA
COSTA SUR OCCIDENTAL DE MÉXICO
Trabajo recibido el 27 de
marzo de
1978 y aceptado para su publicación el 6 de septiembre de
1978.
Ernesto A. Chávez
Escuela Nacional de Ciencias Biológicas,
Laboratorio de Ecología Marina, Apartado Postal 42-186,
México
17, D. F.
La comunidad analizada está compuesta por 97
especies de las cuales 72 son peces, 24 son invertebrados (15
crustáceos, seis moluscos, dos medusas y un poliqueto) y
además
se registró una tortuga marina. Las especies más abundantes
resultaron ser el camarón café (Penaeus
californiensis), el camarón blanco (P.
vannamei), una sardina (Lile stolifera),
anchovetas (Anchoa spp) y varias especies de mojarras
(Gerreidae). La información disponible
permitió hacer un bosquejo de las características del nicho
de
siete de las especies más abundantes, mostrándose alguna
evidencia acerca de los mecanismos que operan en la segregación de
tales
nichos en la comunidad. La abundancia relativa de las especies se
describió a base de los modelos de MacArthur, la serie
logarítmica y la serie log-normal, encontrándose un ajuste
más razonable con esta última. A juzgar por la
composición
de la fauna estudiada, es evidente que se trata de una comunidad
inestable
dominada por especies con estrategia r y controlada por los factores
ambientales.
Los registros de la abundancia de cada especie por
estación sirvieron de base para un análisis de afinidad
entre
inventarios elaborándose dendrogramas mensuales, cuyos resultados
fueron
sintetizados en un diagrama de enrejado que representa las relaciones
entre
unas estaciones y otras, globalmente consideradas. Se determinó la
diversidad de la comunidad mediante el índice de Shannon; los
resultados
mostraron valores mensuales que fluctúan entre 2.35 y 3.91 bits
por
individuo. Aunque no existe un patrón claramente definido de
variación estacional de la diversidad, parece notarse cierta
tendencia a
variar en relación inversa con la salinidad.
El espectro de la diversidad, analizado a
través de la integración sucesiva de inventarios
mostró
cierta analogía con respecto al análisis de afinidad entre
las
muestras hecho a través de los dendrogramas. Se puso en evidencia
un
incremento brusco en la diversidad durante la integración de las
cinco
primeras muestras; posteriormente el aumento fue menos notable.
Las variaciones estacionales de los valores de
diversidad no reflejan características estructurales de la
comunidad
sino diversos grados de estabilidad del biotopo. Se ofrecen algunos
resultados
que permiten suponer la existencia de una relación inversa entre
la
diversidad y las condiciones de fuerte dominancia por parte de alguna
especie.
The community studied is composed by 97 species of
which 72 are fishes, 24 are invertebrates (15 crustaceans, six mollusks,
two
medusae and a polychaete); and a marine turtle. The most abundant species
are
the brown shrimp (Penaeus californiensis), white
shrimp
(P. vannamei), a sardine (Lile
stolifera), anchovies (Anchoa spp) and
several
species of mojarras (Gerreidae) . The information
available
permited to sketch the niche of the seven most abundant species, showing
evidences upon operating mecanism leading to niche segregation in the
community.
Relative abundance of species was described by the
broken stick model, logarithmic and log nonnal series; the data fitted
the best
with the latter one. Based upon the composition of the studied fauna, it
is
concluded that this is an unstable community dominated by strategics and
controlled by environmental factors.
With the data showing abundance of each species per
station a cluster analysis was done obtaining monthly dendrograms, whose
results were summarized in a trellis diagram representing the
relationships
among the stations trough all the year round samplings Diversity of
community
was measured. through the Shannon index; resulting values lie around 2.35
and
3.91 bits per individual. Although there is not a seasonal variation
pattern
clearly defined of diversity values, there seems to be an inverse
relationship
between this and salinity variations.
Diversity spectrum, analyzed through succesive
integration of inventaries showed some analogies respecting the cluster
analysis. Diversity increased abruptly through the integration of the 5
first
samples, after this, the values did not increase so rapidly as at the
former
situation.
Seasonal variations of heterogeneity values do not
reflect structural characterístics of community, but diverse
stability
degree of biotope. Some of the data lead to supposse the existence of an
inverse relationship between diversity and the conditions of strong
dominance
by one or a few species.
El presente trabajo, aunque promovido originalmente con el propósito específico de determinar la época de entrada de postlarvas de camarón a las lagunas Oriental y Occidental, Oaxaca, sirvió como punto de apoyo para la obtención de resultados de mayor alcance, algunos de los cuales se consideran como antecedentes imprescindibles para el entendimiento de los procesos que regulan la dinámica de la comunidad en general así como la supervivencia y migraciones de las especies de camarón en lo particular. La etapa inicial consistió en hacer una prospección que sirvió de base para planear los muestreos sucesivos en lo que se refiere a número de estaciones, naturaleza de los registros y su frecuencia. Los datos obtenidos como resultado de los muestreos tienen por objeto caracterizar el habitat de las comunidades bióticas con las que interactúan mutuamente, y pueden explicar, bajo ciertas circunstancias, sus fluctuaciones y cambios estacionales. Se ha procedido al estudio de la comunidad a través de la macrofauna bentónica tal como puede estimarse mediante el uso de dos redes empleadas, un chinchorro playero de 10 m de largo y una red de puertas de prueba, de las que el primero fue el más utilizado. Uno de los aspectos que más llamó la atención en el transcurso del estudio y del examen de los datos estadísticos, fue la precaria situación que prevalece en esta región y sin embargo, es indudable que, el Estado de Oaxaca con su litoral tiene buenas posibilidades de desarrollo pesquero, con base en la explotación de los recursos de dos biotipos radicalmente distintos siempre y cuando nazca de una correcta planificación. Al primero de tales biotipos pertenecen las pesquerías de mar abierto, que en la actualidad se encuentran representadas por la captura de camarón (el cual se encuentra ya, cerca de su nivel máximo de pesca) y algunos peces valiosos, como la corvina, pargo y bocadulce, que con mucha frecuencia son extraídos durante los lances de arrastre camaronero, y que por lo tanto forman parte eventual de esa captura. El barrilete, atún y jurel se encuentran también en la lista de recursos importantes; sin embargo, la falta de barcos adecuados para su pesca, por una parte y por otra, la carencia de cierta infraestructura básica en la zona, han impedido en gran medida el desarrollo de pesquerías litoral y pelagica que permitan al Estado de Oaxaca elevar su raquítica economía. El otro biotipo se refiere a las lagunas litorales del Estado: Lagunas Superior e Inferior, Occidental y Oriental y gran parte del Mar Muerto. En estos biotipos de alta producción biológica donde el desarrollo de las pesquerías (que en mayor o menor grado, en la actualidad son del tipo artesanal) ofrece también buenas posibilidades de incremento. Dentro de las lagunas litorales del Estado. se encuentran especies como el camarón, sábalo, robalo, lisa y mojarra. De menor importancia, ya que su pesca está en relación con el consumo local, se pueden mencionar el macabí, chiro, sardina, etc. Es evidente que la dificultad de transportar el producto desde los sitios de pesca hasta los centros de consumo, es una de las principales razones por las cuales sus pesquerías se encuentran en un estado de desarrollo incipiente. Los aspectos comprendidos en este trabajo se basan fundamentalmente en los resultados del análisis de las muestras obtenidas mediante el uso del chinchorro playero y la red de puertas, pues aunque el objetivo original del muestreo biológico perseguía determinar la abundancia local de camarón, se considera que ha permitido lograr una representación adecuada de las especies más conspicuas de la comunidad estuárica de la que aquel forma parte, aunque no se soslaya el sesgo inevitable que implica el pretender evaluar la abundancia de peces con el uso de los métodos aquí empleados, de acuerdo con las ideas de Kjelson y Colby (1976), quienes aportan evidencias acerca de la ineficiencia y sesgo en las estimaciones de la abundancia de peces que se capturan en ambientes de estuario con varios tipos de redes. De igual manera, la información analizada permite afirmar que los elementos más importantes son los planctófagos, nivel trófico representado por varias especies de anchoveta (Anchoa spp) y una sardina (Lile stolifera), que también ingieren cantidades importantes de animales bentónicas (véase Tabla l). Otros peces que también resultaron ampliamente representados fueron las mojarras, miembros de la familia Gerreidae, de distribución típicamente tropical y cuya base alimenticia está constituida principalmente por animales bentónicos. Es pertinente tener en cuenta que la importancia de algunas de tales especies radica en las posibles relaciones tróficas que las concatenen con el camarón, principalmente como depredadores de éste. Además, debido al régimen omnívoro de estos crustáceos, las fases larvarias de las especies de peces principalmente, o sus restos, pueden entrar a formar parte de la dieta del camarón, y de esta manera cierran un ciclo de interrelaciones tróficas. OBJETIVOS1. Conocer la composición de la comunidad representada por los macroinvertebrados y peces que comparten el biotopo con los camarones; establecer los cambios estacionales en su composición y diversidad. 2. Analizar los cambios en la composición y diversidad de la comunidad en relación con el régimen hidrográfico o con la variación de alguno de sus componentes. MATERIAL Y MÉTODOSPara el desarrollo del trabajo se contó con una embarcación con motor fuera de borda; la temperatura se registró con termómetro y termonoconductímetro; el oxígeno disuelto se determinó mediante el uso de un equipo portátil Hach para el análisis de aguas; la salinidad se registró, al principio con un densímetro y posteriormente con el conductímetro; la transparencia se midió con el disco de Secchi. Los registros de los tres primeros factores se hicieron en la superficie y en el fondo, excepto en aquellas ocasiones en que no había profundidad suficiente para poder tomar ambas muestras. Las muestras de fauna de macroinvertebrados y peces fueron tomadas con un chinchorro plano de 10 metros de largo por dos de alto con copo de 1.5 metros; la luz de la malla era de un centímetro tanto en las alas como en el copo. Dadas las dimensiones de este arte de pesca, las muestras resultaron selectivas, pues los peces grandes y muy móviles no quedaban atrapados en la red; en cada estación de muestreo se hacían dos lances abarcando ambos un área de 750m² aproximadamente. 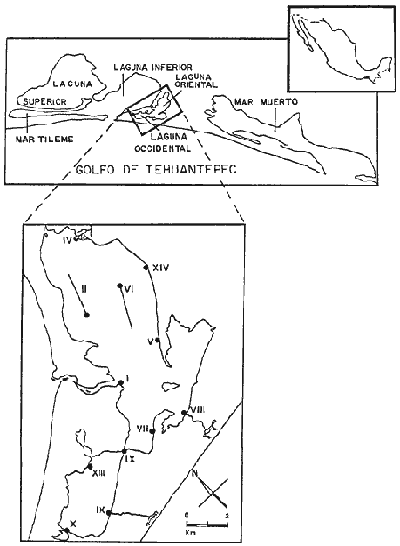 Fig. 1. Localización de las lagunas Oriental y Occidental, en Oaxaca en el Pacífico Sur de México. En las estaciones II y VI, la primera por lo fangoso del fondo y la segunda por su profundidad, el muestreo se hizo mediante el uso de una red de puertas de prueba, abarcando un área de arrastre similar a la de las muestras tomadas con la otra red. El análisis estadístico de los datos se realizó mediante la aplicación de métodos convencionales como son las regresiones lineal y logarítmica, así como de otros más elaborados conocidos colectivamente como métodos de análisis multifactorial. Los primeros se utilizaron para establecer relaciones entre variables. En cambio, los métodos de estadística multifactorial se utilizaron fundamentalmente para el análisis de la comunidad. Debido a la cantidad de información que se manejó y al gran número de cálculos que se hicieron, fue indispensable disponer de los recursos de computación habiéndose utilizado, esencialmente, los sistemas IBM 360 y HP 2100. AREA DE ESTUDIO
DATOS FISIOGRÁFICOSLas lagunas Oriental y Occidental (Fig. l), forman parte de un complejo estuárico de cuerpos lagunares costeros y marismas que se inician hacia su extremo occidental en las inmediaciones de Salina Cruz, Oax., y terminan en la costa chiapaneca, en el Mar Muerto a la altura de Tonalá. De este grupo de lagunas, resaltan por su tamaño las conocidas como Superior e Inferior en el extremo occidental del sistema, y el Mar Muerto en el extremo oriental del mismo. Las lagunas Oriental y Occidental son dos pequeños cuerpos lagunares de origen deltaico que comunican con la laguna Inferior por el estero de Otates que desemboca a la laguna Oriental cerca de su extremo suroeste y por el estero Lagartero que desemboca en el extremo norte de la misma. La comunicación de estas dos lagunas con el Mar Muerto se realiza mediante una serie de marismas que las unen durante los meses de lluvia y que interrumpen la comunicación durante la época de sequía; ésta a su vez, coincide con un descenso en el nivel del mar. Por todas estas características., se les puede considerar dentro del tipo de marismas de playa deltaica formadas por sedimentación terrígena diferencial, según la clasificación propuesta por Lankford. Una descripción más detallada del área de estudio ha sido presentada en un trabajo previo, así como una relación acerca de la posición de las estaciones de muestreo (Chávez et al, 1977). Sin embargo, dada la naturaleza de este trabajo, se considera imprescindible reproducir el área geográfica, así como el mapa donde se muestra la posición de las estaciones de muestreo (Fig. l). ANÁLISIS DE AFINIDAD ENTRE LAS MUESTRASCon base en las listas de los elementos componentes de la macrofauna capturada con el chinchorro playero, se ha hecho un análisis que permite asociar unas estaciones con otras, en función de sus elementos componentes, y permiten inferir sobre las posibles diferencias en la estructura de la comunidad, así como las características más evidentes de su distribución espacial. De acuerdo con la observación de dichas listas, y tomando en cuenta el tipo y dimensiones de los organismos incorporados en el análisis, así como el tamaño del área de estudio, se postula que en las lagunas está representada una sola comunidad y que las diferencias locales que pueden registrarse son debidas tanto a defectos en la eficiencia de las artes de muestreo utilizadas, como a heterogeneidad tanto en la composición del habitat como en la estructura de la comunidad. Se consideran bien identificadas las estaciones que por su localización están más expuestas a los cambios estacionales del entorno, como son la IV y la XI, que manifestaron, la primera, una marcada tendencia hacia el carácter limnético y la segunda a la hiperhalinidad. 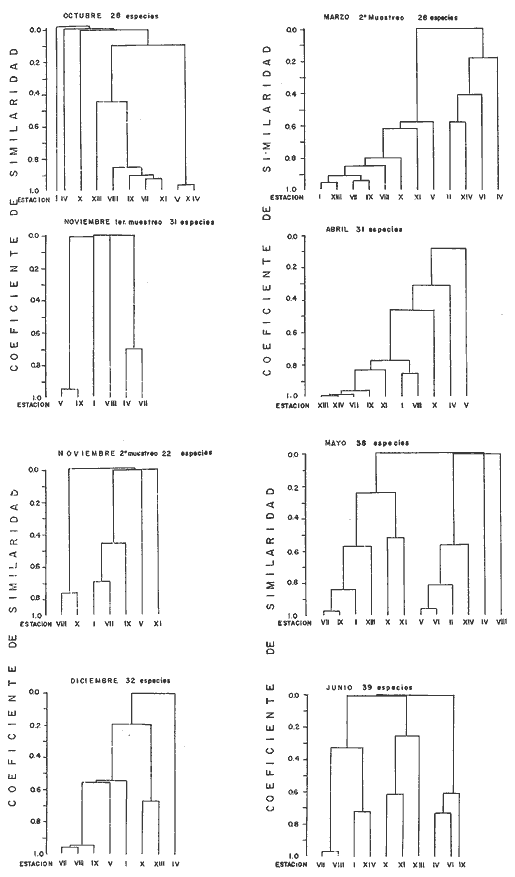 Fig. 2 A y B. Dendrogramas que expresan la afinidad entre las muestras de la comunidad expresada a través del coeficiente de similaridad. 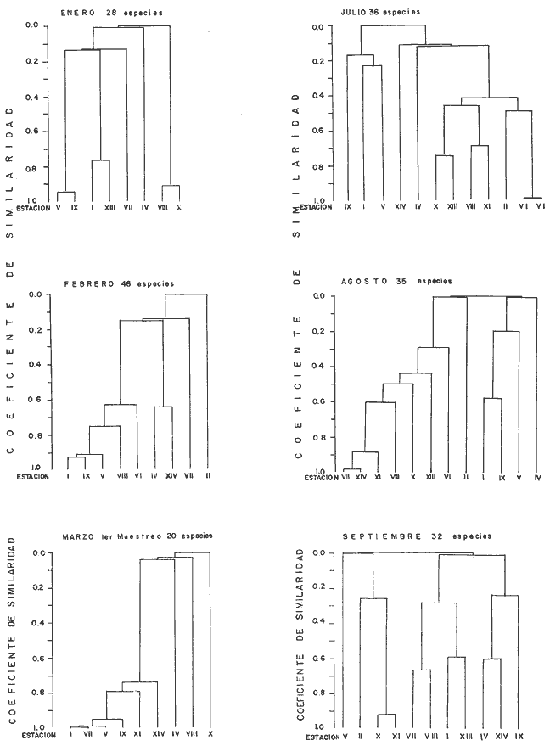 A continuación se procedió al análisis de los datos aplicando un método comúnmente usado en taxonomía numérica (véase Sokal y Sneath, 1963), que consiste en la elaboración de dendrogramas. Un dendrograma es una manera de representar jerárquicamente un conjunto de características en función de la expresión cuantitativa de sus cualidades, muchas de ellas comunes entre sí; el grado de afinidad existente entre las partes de dicho conjunto determina una mayor o menor proximidad en su representación gráfica. Existen varios procedimientos para la elaboración de un dendrograma, y lo complicado de su análisis determina que sólo se puedan realizar, en la práctica, con el auxilio de las computadoras electrónicas (Margalef, 1974), y tal es el caso del presente análisis, para el cual se utilizó un programa de los publicados por Goldstein y Grigal (1972), el cual tiene la ventaja de tener algunas subrutinas que permiten seleccionar varias opciones de análisis; la opción escogida utiliza el coeficiente de similaridad; en el transcurso del análisis, compara sucesivamente unos inventarios con otros a base de sus cualidades (las especies) , ponderadas por el número de individuos de cada una. Los datos que se utilizaron para alimentar el programa son los inventarios mensuales de cada estación, con el número de individuos registrado de cada especie, de tal manera que el análisis se repitió tantas veces como muestreos se hicieron; es decir, se efectuó catorce veces cuyos resultados se han consignado en los dendrogramas correspondientes (Fig. 2). Sin embargo, de su observación surge la incertidumbre ya que, en general, la mayor parte de las estaciones resultan relacionadas con un coeficiente de similaridad mayor de 0.5; por lo tanto, se decidió hacer una síntesis de tales resultados agrupándolos en una matriz de asociación en la cual se dispuso por una parte, el número de veces en que dos estaciones fueron objeto de muestreo en la misma fecha y por otra, el número de veces en que aparecen asociadas con afinidad mayor de 0.5. Esto sirvió de base para determinar, la proporción de frecuencia de afinidad entre los distintos pares de estaciones analizados, concentrando estos resultados en un diagrama de enrejado (Fig. 3), donde se aprecia que las estaciones XI y XII fueron las más afines a lo largo del estudio, con un valor de afinidad de 0.88; además con las estaciones VII, VIII y X ambas mantienen un grado de afinidad comprendido entre valores de 0.60 y 0.80. DATOS ACERCA DE LA ESTRUCTURA DE LA COMUNIDADSegún Cody y Diamond (1975), la manera según la cual los recursos de una comunidad se distribuyen entre las especies, así como los patrones de abundancia espacial y temporal de éstas es su estructura, "en particular si las especies están incorporadas a las comunidades de manera no aleatoria, y si la estructura fina de tales agrupaciones está determinada por el ambiente físico y biológico, los patrones de la estructura de la comunidad deben ser reproducibles, independientemente del conjunto de especies del que forman parte". Por lo que se refiere a la comunidad de las lagunas, en el diagrama de la figura 4 se muestra lo que indirectamente pudiese interpretarse como una evidencia de la estructura y cambios estacionales de la misma representados por la composición relativa de las especies dominantes, donde además se observa alguna variación inversa de la abundancia como en el caso de Penaeus californiensis y Lile stolifera, que durante los siete primeros muestreos la manifiestan de manera muy llamativa; de igual manera, Eucinostomus sp parece seguir el mismo tipo de variación que P. californiensis aunque en una proporción considerablemente más modesta. Si bien un bosquejo de esta naturaleza resulta poco informativo, puede ser de alguna utilidad más que nada para mostrar la composición y variación de los elementos más conspícuos, y al mismo tiempo apreciar, de acuerdo con su variación, los efectos que pueden tener sobre algunos de ellos o sobre la comunidad en general, tanto las interacciones debidas a competencia y depredación, como la influencia de los factores ambientales que bien pudiesen, bajo ciertas circunstancias, ejercer un efecto tal que supere o impida que tales interacciones pluri específicas se manifiesten de una manera más nítida, como tal vez sea el caso de, por lo menos, en los tres últimos muestreos donde las relaciones de abundancia se vuelven mucho menos claras, y al mismo tiempo permiten entender el punto de vista de Leigh (1975), quien discute cómo la estructura de una comunidad puede afectar la respuesta de sus poblaciones a la variación en el ambiente. 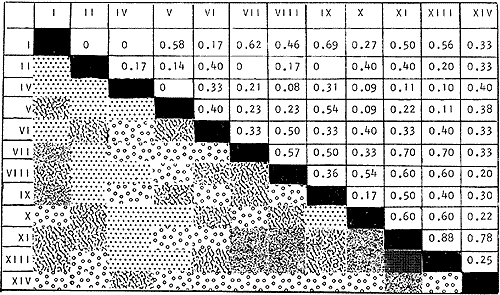 Fig. 3. Diagrama de enrejado que muestra el grado de afinidad entre estaciones, expresado como una proporción de la frecuencia de asociación. Se ha visto que la heterogeneidad hidrográfica a pequeña escala puede influir de manera decisiva sobre la composición de las comunidades planctónicas de modo que éstas se distribuyen en el espacio de manera irregular siguiendo variaciones paralelas a la primera. Esta respuesta de los organismos del plancton tiene su origen en la coexistencia de elementos ecológicamente distintos que al encontrar condiciones favorables aseguran una respuesta instantánea de sus poblaciones a los cambios ambientales (Margalef, 1969). En el transcurso del desarrollo del presente estudio, se encontró una curiosa analogía entre lo expuesto en líneas anteriores y la composición de las muestras en estudio y en especial de los componentes del necton aunque quizás más sutil pues se trata de una substitución parcial de las proporciones entre los elementos dominantes y resulta más notable al tratarse de las densidades absolutas de la población. Es razonable suponer que estos cambios puedan obedecer al mismo patrón señalado para el plancton donde es este último el estímulo que determina la concentración y composición del necton cuyos elementos substituyen con su capacidad de desplazamiento lo que en el plancton sería una tasa de multiplicación elevada y que en ambos casos se traduce en cambios en la densidad de población en un lugar y momento determinados. Por otra parte, esta situación se manifiesta de un modo más o menos claro dentro del ecosistema en estudio, excepto cuando las condiciones hidrográficas se tornan muy extremosas, pues entonces la respuesta ante los problemas de osmorregulación y adaptativos en general adquiere preponderancia sobre los mecanismos que condicionen la búsqueda y selección del alimento. 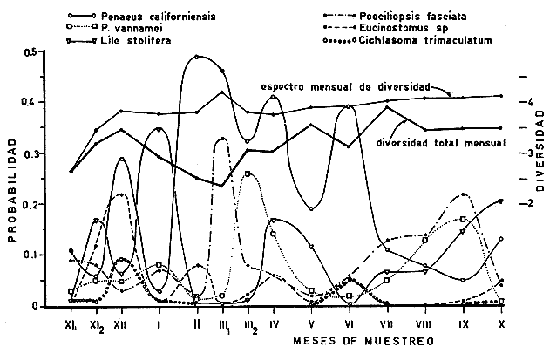 Fig. 4. Abundancia de las especies dominantes expresada en términos de probabilidad de acuerdo con el total de individuos, registrados en cada muestreo. Las dos líneas superiores representaban la diversidad total mensual y el espectro global elaborado mediente la integración sucesiva de éstos. Esta discusión da pie para ofrecer alguna información que pueda contribuir a la definición del nicho ecológico de las especies más abundantes, vertida en gráficas elaboradas a partir de los datos recopilados entre el hábitat durante el desarrollo del presente estudio; esto resulta oportuno si se toma en cuenta parte de lo que afirma Patrick (1975) en el sentido de que los límites del nicho de cada especie, tanto espaciales como temporales están determinados por factores físicos, competencia y depredación. Este tipo de representación gráfica de los datos ha sido utilizado anteriormente en el plancton en diagramas conocidos como Temp.-Sal.-plancton; en este caso, Margalef y González-Bernáldez (1969) lo consideran como un espacio de probabilidad definido, para cada especie, dentro de otro de muchas dimensiones, tantas como factores puedan distinguirse en el ambiente del que forman parte; según lo anterior, la probabilidad de estar presente será tanto mayor cuanto más cerca esté de la concurrencia de condiciones óptimas para cada especie. En las figuras 5 a 9 se presenta la información recopilada, para siete de las especies más abundantes observadas, a saber, Penaeus californiensis, P. vannamei, Lile stolifera, Melaniris guatemalensis, Poeciliopsis fasciata, Eucinostomus sp. (especie aún no descrita) y Ciclilasoma trimaculalum. Los factores ambientales que han servido de marco ecológico son temperatura, salinidad y oxígeno; sin embargo, dada la relativamente escasa información de que se dispuso para elaborar dichos diagramas, con la consiguiente falta de representatividad, se decidió presentar una nube de densidad de población expresada como número de individuos por estación, más que en términos de probabilidad. De la observación de dichos diagramas resulta llamativa la forma en que las especies se distribuyen desigualmente bajo las distintas combinaciones de valores de los factores ambientales, lo cual pone de manifiesto el mecanismo mediante el cual tiene lugar la segregación de nichos en especies cercanas como es el caso de las dos de Penaeus consideradas. En este punto es oportuna la referencia al trabajo de Smith (1972), quien alude al hecho de que la heterogeneidad ambiental tiende a promover la diversidad indirectamente y a través de aumentos en el grado de competencia intra especifica de los elementos de la comunidad. 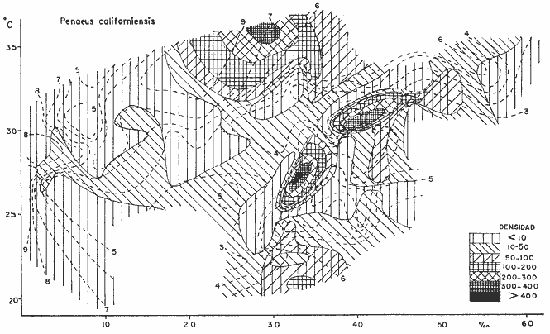 Fig. 5. Condiciones del hábitat representando por la concurrencia de distintos valores de temperatura, salinidad y oxígeno disuelto en relación con las densidades de la población de vannamei. 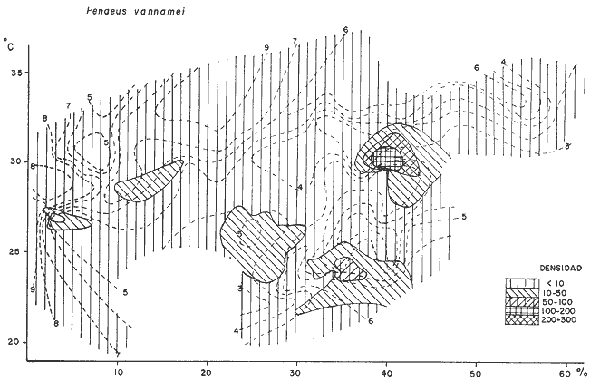 Fig. 6. Condiciones del hábitat, representado por la concurrencia de distintos valores de temperatura, salinidad y oxígeno disuelto en relación con las densidades de la población de vannamei. 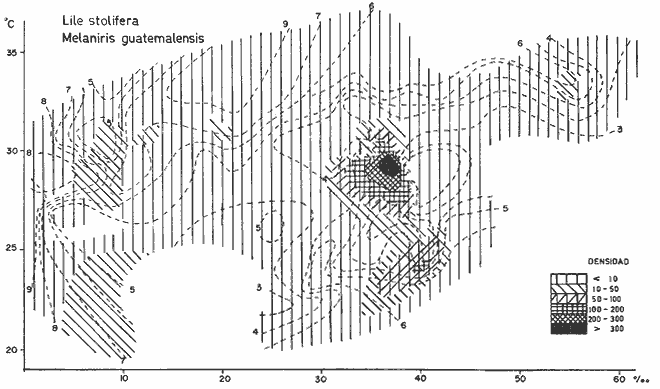 Fig. 7. Condiciones del hábitat, representado por la concurrencia de distintos valores ae temperatura, salinidad y oxígeno disuelto en relación con las densidades de las poblaciones de Lile stolifera y Melaniris guatemalensis. 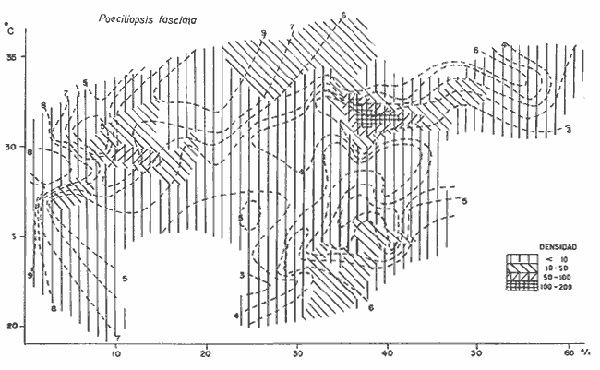 Fig. 8. Condiciones del hábitat, representado por la concurrencia de distintos valores de temperatura, salinidad y oxígeno disuelto en relación con las densidades de la población de Poeciliopsis fasciata. 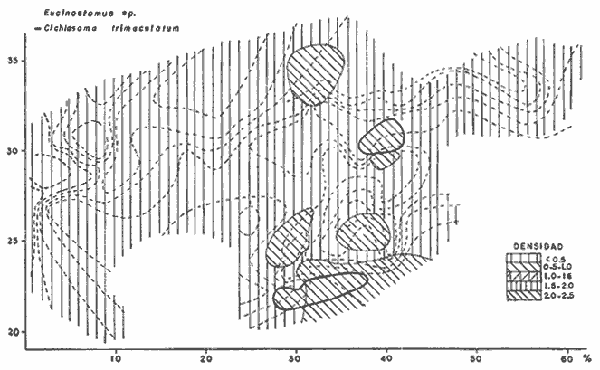 Fig. 9. Condiciones del hábitat representado por la concurrencia de distintos valores de temperatura, salinidad y oxígeno disuelto en relación con las densidades de las poblaciones de Eucinostomus sp. y de Cichlasoma trimaculatum. ABUNDANCIA RELATIVA DE LAS ESPECIES Y MODELOS DE AJUSTECon el interés de lograr entender mejor los mecanismos que determinan la estructura de la comunidad, su estudio se ha enfocado, desde hace algún tiempo, hacia el conocimiento de la abundancia relativa de las especies analizado desde un punto de vista estadístico, pretendiendo con ello tomar en cuenta no sólo la manera como los recursos se distribuyen entre los distintos elementos que la constituyen sino también los posibles mecanismos de interacción de unas especies con otras y con su medio ambiente, que por otra parte, se traduzcan en situaciones tangibles que determinen el grado de dominancia, estabilidad, madurez y sucesión de una comunidad. Como resultado de tal interés se han propuesto varios modelos, los que infortunadamente tienen más valor heurístico que analítico, pues simplemente describen con mayor o menor precisión la abundancia relativa de las especies que componen la comunidad, pero dicen muy poco con respecto a su estructura. Los modelos mejor conocidos y ampliamente discutidos en la literatura son la serie logarítmica, propuesta por Fisher et al (1943), el modelo de la barra fragmentada debido a MacArthur (1957) y el de la llamada distribución log normal de Preston (1948). Los datos recopilados en el presente estudio se utilizaron con la intención de hacer el ajuste de los mismos al modelo más adecuado; por lo tanto, en las líneas que siguen a continuación se hace una breve exposición de los postulados en que se basa cada uno y los resultados del ajuste: a) La Serie Logarítmica. Es simplemente una distribución de frecuencias y aparentemente no se apoya en razonamiento alguno de tipo biológico, aunque uno de sus parámetros α) , pueda tener este significado. La serie es la siguiente: 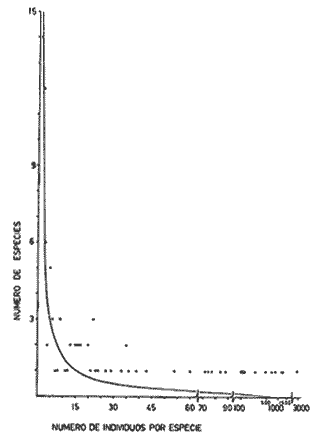 Fig. 10. Distribució n del número y abundancia relativa de las especies y su ajuste a la serie logarítmica (X= 0.9985 y oc = 15.32) Se observa que este ajuste es bueno solo en la parte izquierda de la curva. αX, αX² ÷ 2, αX³ ÷ 3, αX4 ÷ 4, αX5 ÷ 5··· αXn ÷ n donde n1 = Número de especies representadas por un individuo. n = Clase de abundancia. x = Constante relacionada con el número promedio de individuos por especie; depende del tamaño de la muestra y su valor suele ser muy cercano a 0.9. α = n1/x. Es una medida de la diversidad en la colección. Como resultado de la aplicación de los datos de la tabla 1 a este modelo, el valor de las constantes resultó ser de x = 0.9985 y α = 15.32. La línea que describe esta serie se expone en la figura 10 donde puede apreciarse que su ajuste es bueno sólo en la parte izquierda de la misma, pero no lo es en la correspondiente a las especies raras; por lo tanto es poco satisfactorio. b) Modelo, de la línea fragmentada ("broken stick model"). Las inferencias sobre las cuales MacArthur concibió este modelo se basan en la suposición de que los recursos disponibles para la comunidad, representados por una línea o barra, está dividida al azar en s fragmentos y a cada uno se lo considera como el recurso utilizado por una especie, o sea su nicho. De acuerdo con lo anterior, el modelo establece que las Sespecies se dividen el ambiente en S nichos no superpuestos cuyos tamaños relativos se distribuyen aleatoriamente. El único parámetro del modelo es S o sea, el número de especies y si éste se conoce, a partir de él pueden predecirse las abundancias relativas de todos los elementos cuando estos se disponen en orden creciente de abundancia. Por lo tanto, el modelo es el siguiente: Nj ÷ N = 1 ÷ S j/Σ/i = 1 1 ÷ Si + 1 en donde Nj = Es la abundancia esperada de la j décima especie expresada como un por ciento del número total de individuos en la comunidad. El ajuste de este modelo a los mismos datos resultó menos afortunado que el primero, según se aprecia en la figura 11, por lo que se intentó la aplicación del tercer modelo. c) La serie log normal. Esta distribución se basa en la hipótesis según la cual el nicho de cada especie se considera dependiente de una multitud de factores distintos que determinan la amplitud del tal nicho y consecuentemente los recursos de que la comunidad dispone se deben repartir entre las especies de ésta de una manera equivalente a una curva normal, de modo que tanto las especies abundantes como las raras se dispondrán hacia los extremos de la distribución, mientras que la mayor parte del inventario, que está representado por especies de frecuencia intermedia, ocupará la parte central de la curva. Este modelo tiene la peculiaridad de que los intervalos de la distribución son sus logaritmos de base dos y de acuerdo con lo antes indicado, su valor modal se localiza en el intervalo que corresponde al mayor número de individuos. Además, debido a que suele haber muchas especies raras que no aparecen representadas en la muestra la distribución log normal suele aparecer truncada hacia la izquierda y en una comunidad no alterada el valor modal generalmente se localiza en el primer intervalo. En un trabajo posterior a aquel en el que se postuló por primera vez este modelo, Williams, (1964) hace un análisis bastante ambicioso sobre la aplicación de esta serie, así como de la logarítmica a distintos grupos de colecciones faunísticas. La función que describe la serie log normal es la siguiente: S = Soe- (a R)² donde: S = Es el número de especies en el R-ésimo intervalo a la izquierda y derecha del valor modal. 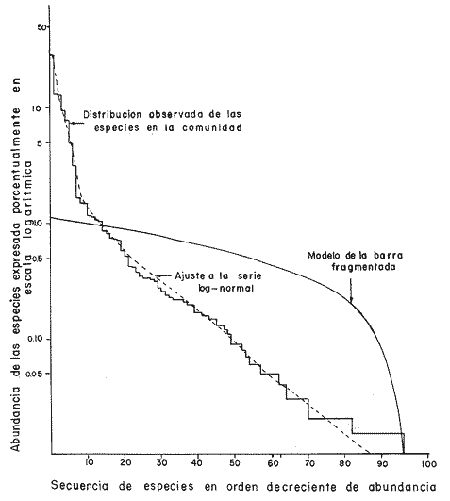 Fig. 11. Número y abundancia relativa de las especies de la comunidad y las distribuciones teóricas esperadas de acuerdo con el modelo de MacArtur y con la log normal. So = Es el número de especies en el intervalo modal. a = Parámetro estimado a partir de los datos. Como resultado de la aplicación de los datos a esta serie se encontró un ajuste más aceptable que en los dos primeros casos, según puede observarse en las figuras 11 y 12. En atención a los patrones que determinan la abundancia y diversidad de las especies, May (1975) hace una revisión analítica del tema, con énfasis en la intención de separar de los modelos de distribución a los aspectos que reflejen algún detalle de la estructura de las comunidades los que tengan un significado simplemente estadístico gobernado por leyes de grandes números. Una de las características por él señaladas y que vale la pena hacer notar, se refiere al hecho de que en los modelos que describen la abundancia de las especies, la distribución log normal refleja el teorema del límite central, mientras que en aquellos casos donde es posible hacer el ajuste de los modelos tales como el de la barra fragmentada, o la serie logarítmica, está implícito algún aspecto de la biología de la comunidad. 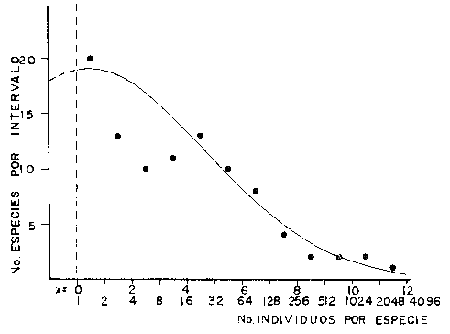 Fig. 12. Relación que describe la abundancia relativa de las especies y su ajuste a la serie log-normal. May (1975), menciona que las distribuciones de abundancia del tipo log normal en comunidades con organismos "oportunistas", como posiblemente sea el caso de las que son objeto del presente estudio, reflejan poco detalle sobre la estructura de la comunidad y que las especies dominantes son simplemente aquellas que recientemente han disfrutado de un valor grande de r, o sea su potencial biótico y por lo mismo, en momentos distintos habrá diferentes dominantes. Por lo que respecta a la peculiaridad observada en la serie log normal de que el valor de a usualmente vale 0.2 y el de y 29 1, es decir, la relación existente entre el intervalo donde el número de especies es máximo, dividido por el intervalo donde se localiza la especie más abundante, May sugiere que en su opinión estas características no tienen mayor significado que el de ser propiedades matemáticas de la distribución log normal. Al respecto de los modelos de la barra fragmentada y la serie logarítmica o geométrica, él opina que son distribuciones características de comunidades relativamente simples cuya dinámica está dominada por algún factor individual, pues en un extremo está el citado en primer término, que es una expresión estadística realista de una distribución intrínsecamente uniformes; y en el otro está la serie logarítmica que con frecuencia expresa estadísticamente el proceso desigual del "niche preemption", o sea, el derecho de ocupar un nicho disponible antes de que otra especie lo haga, del cual la forma ideal es la serie geométrica. Por otra parte, la serie log normal, que probablemente es la más importante, se puede considerar en muchos aspectos como intermedia entre las otras dos. Toda esta serie de modelos, aunque ilustrativos, poco dicen acerca de la estructura de la comunidad, y su validez reside en el carácter esencialmente descriptivo de las distribuciones de la abundancia de las distintas especies representadas en la muestra objeto del análisis. De acuerdo con Margalef (com. pers.), una probable alternativa que tal vez convendría sondear, consiste en tratar de desarrollar un modelo de distribución parecido al de MacArthur, elaborado sobre la base de las mismas premisas, pero derivado a partir de las relaciones depredador-presa. DIVERSIDADEn el estudio de las comunidades, uno de los problemas cuya solución mas ha inquietado a los ecólogos durante los últimos años se relaciona con la búsqueda de posibles leyes que gobiernen la estructura y composición de aquéllas. Entre las vías de investigación que en este sentido se han estado desarrollando destaca el estudio de la variabilidad y abundancia relativa de las especies a través de la medida de su diversidad. El concepto que este término implica según señala Margalef (1974), significa no simplemente el número de especies de la comunidad, sino la complejidad de su red trófica; por otra parte, según señala Pielou (1975), una medición de esta característica no debe sobrevalorarse, pues es simplemente un estadístico meramente descriptivo análogo al intervalo estadístico de una variable expresada cuantitativamente, como la variancia, y consecuentemente los valores son significativamente distintos, pues se supone que cada uno mide la diversidad de la población entera (Poole,1974), la cual, en las comunidades naturales no excede de 5 bits por individuo (Margalef, 1972). Por lo tanto, cualquier medición que se intente hacer al respecto necesariamente debe limitar se topográfica y taxonómicamente, o sea, referirse al espectro de la diversidad, a fin de evitar sesgos importantes en su estimación; además, no debe perderse de vista que las inferencias que se hagan son válidas sólo para la muestra, y cualquier extrapolación que se intente hacer tiene carácter exclusivamente subjetivo. Al mismo tiempo, y a pesar de esta restricción, no debe olvidarse que la cantidad de biomasa de cada nivel trófico guarda estrecha relación con la estructura de la comunidad (Hutchinson, 1975) y ésta a su vez debe reflejarse en su diversidad. Por otra parte, y evocando las palabras de Margalef (1972), "Una de las dificultades para imputarle significado a la diversidad estriba en equilibrar adecuadamente el realismo y la generalidad; así como en la necesidad de observaciones cercanas a los ecosistemas reales y la conveniencia de un modelo abstracto que sea en lo posible, aplicable a un amplio conjunto de sistemas". Existen varias formas de estimar el valor de la diversidad de una taxocenosis, algunas de las cuales son citadas por Margalef (1962), sin embargo, es conveniente que la medición de esta característica por cualquier método que se estime, dé un valor directamente proporcional al número de especies, ponderados por el número de individuos de cada una de las presentes en la muestra. Las unidades de diversidad dependen de la base de logaritmos utilizada; en este caso se usó la base dos y por lo tanto la diversidad se expresa como contenido de información, en bits por individuo, ya que se utiliza el índice de Shanon, ampliamente usado en estudios ecológicos. Este índice se determina con la fórmula siguiente: H = - ΣPi log2 Pi donde: H = Indice de diversidad Pi = Probalidad de aparición de la especie i en la muestra. Los valores de diversidad observados se encuentran en su mayor parte, alrededor de 2 bits por individuo. Sobre la base del marco ambiental, tan variable estacionalmente dentro el cual se desarrolla la comunidad estudiada, se considera a esos valores como indicadores de una comunidad inestable controlada por los factores ambientales. Según Sanders (1968) en un ambiente de esta naturaleza predominan las especies estrategas de la revéase también Southwood, 1976), lo cual es lógico esperar después de observar cómo los cambios estacionales más extremosos puestos de manifiesto en este caso por el influjo de grandes volúmenes de agua dulce al sistema, provocaron un descenso notable en la densidad de población de especies anuales como el camarón así como de algunos elementos conspícuos del fitobentos (Gracilaria sp.). A este respecto, Kushian (1976) ofrece evidencias en el sentido de que la dominancia de peces depredadores o de peces forrajeros es una función de la estabilidad del nivel del agua, lo cual se reflejó en los valores de diversidad por él estimados. LA DIVERSIDAD Y EL ECOSISTEMAAl verter los datos de temperatura, salinidad y oxígeno sobre una gráfica (Fig. 13), se ha podido observar que existe una cierta tendencia de la distribución de este último con respecto al de los primeros, pues las concentraciones más altas del oxígeno (mayores de 5 ppm) se distribuyen principalmente a temperaturas mayores de 30° en aparente contradicción con las características de solubilidad de este gas en el agua en función de la temperatura; sin embargo, es bien sabido que en este tipo de ecosistemas pueden darse condiciones transitorias durante las cuales, por alta producción primaria y escasa turbulencia, pueden ocurrir situaciones de sobre saturación de oxígeno disuelto en el agua; esto ocurrió independientemente de la concentración salina aunque las concentraciones máximas de oxígeno corresponden a valores de salinidad cercanos a 30% y alta temperatura así como a valores intermedios de esta última en ambiente temporalmente limnético (estaciones I y II); por otra parte, valores altos del mismo se localizan también a temperaturas más bajas (menores de 300) cuando las salinidades observadas eran casi limnéticas, así como cuando éstas eran superiores a 29‰ y la temperatura menor de 25°. 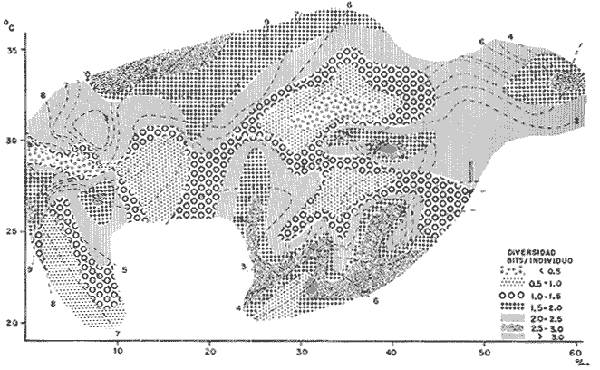 Fig. 13. Distribución de la diversidad en relación con distintas combinaciones de salinidad, temperatura y concentración de oxígeno. Los valores altos de oxígeno registrados junto con salinidades bajas se atribuyen principalmente a intercambio con la atmósfera, pues durante los meses de agosto, septiembre y octubre el río Ostuta descargó grandes volúmenes de agua provocados por las lluvias que cayeron en las partes altas de la región y la descarga hidráulica se efectuó de manera muy turbulenta; en cambio, las concentraciones altas registradas bajo el régimen de mixohalinidad se atribuyen esencialmente a producción biológica, tanto del fitoplancton como del fitobentos. El análisis de la diversidad ha sido desglosado en varios aspectos, dada la naturaleza del tema que se aborda en cada caso. En esta ocasión se trató de encontrar alguna posible relación entre las características del ambiente, determinadas por los tres factores antes mencionados y la diversidad de la comunidad, registrada a partir de las muestras de peces y macroinvertebrados capturados con el chinchorro playero. Para tal efecto se vertieron los datos de diversidad en la gráfica antes discutida, pero agrupando los valores de la misma en intervalos de 0.5 bits, según se observa en esa figura, donde puede apreciarse que prevalecen los valores que caen en el intervalo de 1.5 a 2.0 bits; sin embargo, lo que atrae más la atención son las áreas de diversidad alta, que en parte coinciden con las concentraciones altas de oxígeno bajo las circunstancias señaladas para el mismo, pero además se ponen en evidencia tres manchones aislados de alta diversidad que no se explican fácilmente ya que no se vislumbra una clara relación con los factores ambientales y se sospecha que puede ser un problema atribuible a varias causas, tales como algún posible error de muestreo que por insuficiencia de datos impida mostrar una variación mas uniforme; o por otra parte debido al hecho de que en dicha gráfica están incluidos los datos de todos los muestreos, lo cual representa una complicación adicional pues el tiempo está implícito en ellos y con aquél la variación estacional, porque es razonable también asignarle a ésta el origen de esas manchas aisladas de diversidad alta. 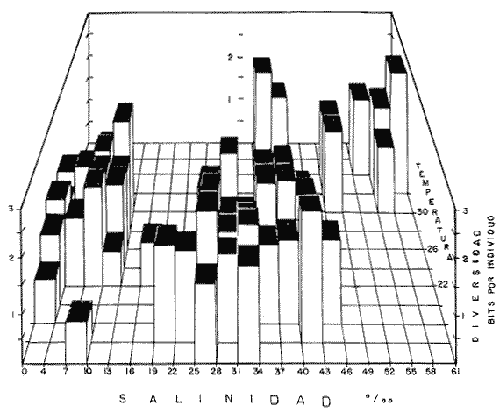 Fig. 14. Gráfica que muestra la posible influencia de la temperatura y la salinidad sobre la diversidad de la comunidad. Con la intención de establecer una relación más evidente entre la influencia del ambiente sobre la comunidad, se hizo comparación gráfica entre el índice de diversidad y dos de los factores ambientales, la temperatura y la salinidad (Fig. 14); el resultado parece no ser muy claro con respecto a la influencia de la temperatura, pues como se hizo notar anteriormente en un estudio similar (Chávez, 1972), los efectos de este factor se reflejan fundamentalmente en la composición faunística y en los ajustes fisiológicos de los organismos a los cambios estacionales de aquél; en otras palabras, los efectos de la temperatura se reflejan en las características biogeográficas de la comunidad en una localidad determinada, pues resultan muy difíciles de detectar aisladamente después de que las especies han superado la etapa larvaria. En cambio, por lo que se refiere a la salinidad, se ponen en evidencia tres grupos más o menos bien marcados; se atribuye como causa probable de esta situación a la presencia de fauna de raigambre limnética en el grupo de especies que se localizan hacia las salinidades más bajas; los inventarios situados hacia la mitad de la escala se atribuyen a aquella parte de la comunidad en la que predominan elementos muy eurihalinos, tanto marinos como estudia y finalmente, el grupo de la derecha contiene especies esencialmente estuarinas, pues se ha observado que las mejores adaptadas a los estuarios son capaces de soportar mejor las condiciones de hiperhalinidad en el agua (Gunter, 1945). 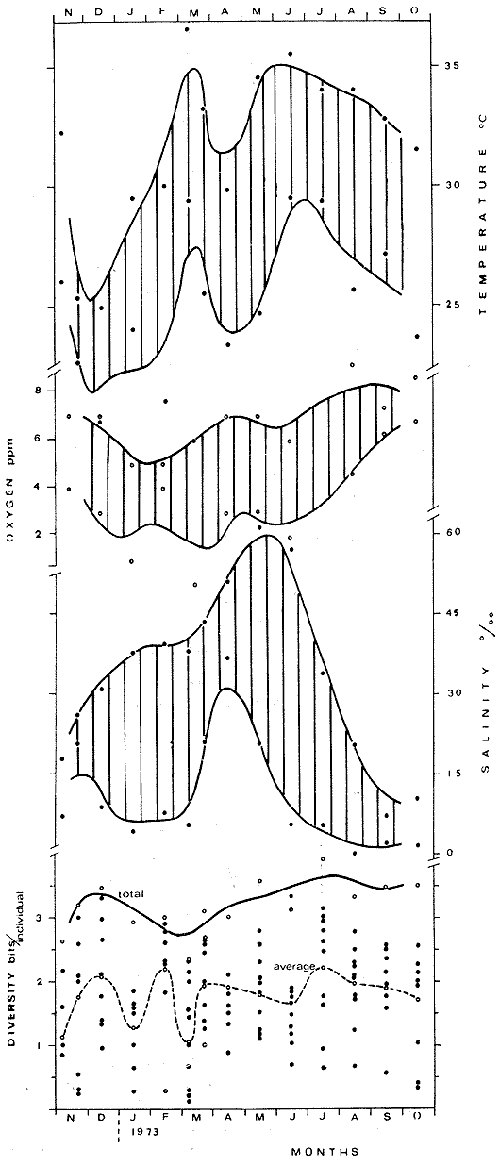 Fig. 15. Variaciones estacionales de los factores ambientales analizados, comparados con los cambios en la diversidad. Las líneas que limitan áreas sombreadas unen valores correspondientes a promedios móviles. Lo mismo se hizo con los datos de diversidad total mensual. 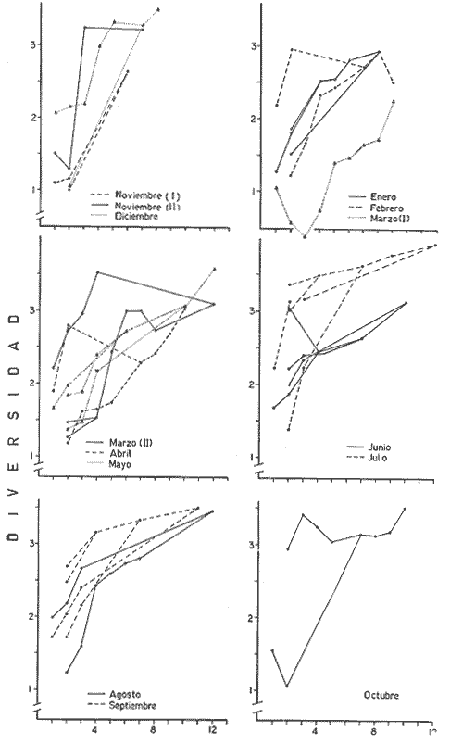 Fig. 16. Espectro mensual de la diversidad. La integración sucesiva de las muestras se hizo con base en los dendogramas de la figura 2. Después de analizar cada uno de los aspectos relacionados con la diversidad y sus cambios, se hace evidente que dentro de la variación estacional en el ambiente, las interrelaciones en los factores que caracterizan al ecosistema resultan claras en su conjunto, pero con modalidades particulares difíciles de dilucidar, determinadas por las características locales del biotopo; a ello se atribuye el hecho de que la única variación estacional nítidamente definida sea la salinidad. Consecuentemente, la variabilidad de la comunidad, representada por el índice de Shanon, muestra cambios a lo largo del año, pero éstos no siguen un patrón estacional bien marcado, según se aprecia en la figura 15, donde se indican las variaciones extremas de los factores ambientales antes discutidos. Por otra parte, la comunidad, cuya variabilidad está representada en esa misma figura por los valores total y medio mensuales no muestran cambios estacionales claros, pues sólo parece vislumbrarse una ligera tendencia oscilatoria con sus valores más bajos observados en la estación seca y los más altos durante la temporada lluviosa. ESPECTRO DE LA DIVERSIDADEl análisis de este espectro se ha llevado a cabo mediante la integración sucesiva de inventarios muestrales de modo análogo al realizado por Margalef (1969), o de grupos de inventarios según el caso, lo cual se ha determinado tomando como base los dendrogramas previamente elaborados; en la figura 16 (a a f) se exponen los resultados y en ellos puede apreciarse un claro incremento en la diversidad conforme se incorporan nuevas listas de especies. Es muy evidente también el paralelismo que se observa entre la composición de los dendrogramas y el espectro de la diversidad, pues al haber realizado el análisis por grupos, se ha puesto de manifiesto una segregación de los valores mostrándose con frecuencia dos grupos por lo menos, con diversidades notablemente diferentes, pues generalmente se presenta uno con tendencia a tener alta diversidad y otro con diversidades bajas. También es muy notorio apreciar que son pocos los valores parciales que superan los tres bits, y los descensos observados cuando se agrupan más de dos inventarios se atribuyen a deficiencias en el tamaño muestral. Finalmente, es evidente en general, un incremento brusco en la diversidad durante la integración de las cinco primeras muestras; posteriormente el aumento resulta menos notable. Otro aspecto sobre el que se ha considerado pertinente llamar la atención se refiere al ordenamiento de los valores de la diversidad observada en las listas de especies de cada campaña, tal y como se ha hecho en la figura 17; esta disposición de los valores guarda una relación con la distribución del número de individuos de las especies componentes de la comunidad y puede tener algún significado con respecto la disposición espacial de la estructura y su relación con los factores limitantes del ambiente, pues al mismo tiempo que indica si existe un hábitat con cierta homogeneidad donde la diversidad se distribuye uniformemente, como en el caso de las muestras del mes de enero, también es posible hacer inferencias sobre un gradiente tanto de la estructura de la comunidad como del ambiente y en este caso se pondría de manifiesto en la distribución resultante de los valores individuales cuya tendencia estará caracterizada por una mayor pendiente. Dadas las características de mobilidad de la fauna objeto del presente estudio se ha considerado un poco difícil el estudio de la variación mensual de la diversidad en cada estación de muestreo, pues no refleja características estructurales de la comunidad en sí misma, sino más bien puede significar estabilidad o inestabilidad del biotipo que determine ser más o menos habitable para la ictiofauna que sobre esta base lo ocupe o abandone. Al examinar los valores de diversidad plasmados en los histogramas de la figura 18, resulta un poco difícil discernir alguna tendencia estacional, y donde se nota con cierta claridad es en las estaciones IV y XIV que muestran variaciones paralelas a las de la salinidad; esto indudablemente guarda una relación estrecha con la influencia limnética, dada la proximidad del río Ostuta cuyo caudal, aunque escaso durante la mayor parte del año tiene algún efecto limitante sobre los organismos que en general son de raigambre marina y que a pesar de su capacidad osmorreguladora rehuyen los cambios violentos de salinidad. Por otra parte, resulta llamativa una relación aparentemente inversa a la antes discutida y que con poca claridad se pone de manifiesto en las estaciones I, VII y IX; es de suponerse que en este caso la reducción en el valor de la diversidad obedezca principalmente a un descenso en el nivel medio del mar que se hizo notorio durante los meses de febrero y marzo, dando como resultado una importante disminución del nivel de las lagunas en las que algunas áreas de cierta extensión permanecieron secas durante más de un mes, particularmente en los extremos sur y sureste de las mismas, al grado de que cerca de un 25%, del fondo de la laguna Occidental se encontraba expuesto al aire. 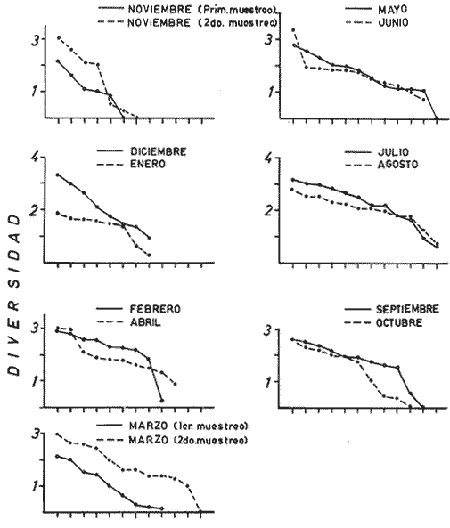 Fig. 17. Arreglo mensual de la diversidad en orden descendiente. 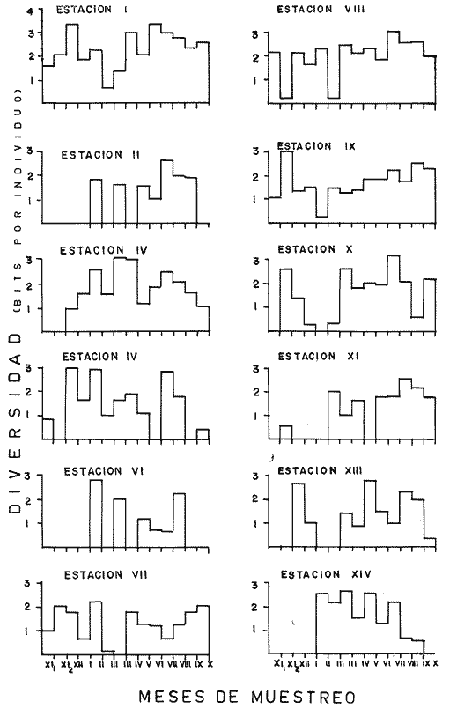 Fig. 18. Variación estacional de la diversidad observada en cada estación de muestreo. Margalet (com. pers.) sugiere sondear la posibilidad de analizar el espectro de la diversidad desde un punto de vista poco tratado; esto es, a partir de la fórmula de Brillouin sumando un segundo término que contenga los valores de diversidad con casi los mismos datos que en el caso de la muestra global, pero con la diferencia de que a una especie común se le suprima un ejemplar y a una rara se le agregue otro; esto es lo que por otra parte se está llevando a cabo en la práctica con la intervención humana en la explotación de recursos o aún incluso en las comunidades sujetas a los efectos de la contaminación ambiental y el análisis de un concepto esencialmente teórico y tradicionalmente implicado en discusiones básicamente académicas, de manera casi inesperada adquiere matices de interés práctico. Esto quizás hiciese vislumbrar un poco de claridad acerca de la estructura de la comunidad que se pondría en evidencia a través de los cambios en el espectro resultante, así como de sus cambios a través del tiempo, pues probablemente pudiesen estar implicados los mecanismos de la sucesión. Al mismo tiempo, parece ser este el mecanismo matemático adecuado para describir el proceso que ha propuesto Connell (1975) sobre la estructura y sucesión de las comunidades; es decir, tratando cada grupo por separado según su similaridad y agregando primero los inventarios con coeficiente más alto hasta llegar a un punto en el que se le agrega el inventario global de un grupo vecino de muestras y así sucesivamente hasta agotar el muestreo. Sin embargo, cuando compara uno sobre el mapa la distribución espacial de las muestras, se observa que con frecuencia dos muestras muy afines se encuentran físicamente muy distanciadas una de otra y esto estimula el espíritu de inconformidad, pues causa la impresión de que se incorpora un factor de ruido; a pesar de todo no fue posible resolver este problema de una manera más satisfactoria. Margalef (1967) ha hecho algún intento por abordar este problema mediante la construcción de una especie de retículo cuya trama tiene una densidad proporcional a la diversidad, pero en ese mismo trabajo el autor citado lo considera poco recomendable y en cambio, más recientemente (Margalef, com. pers.), sugiere la posibilidad de que exista alguna relación entre el índice de diversidad y el logaritmo de la distancia, ya que se trata de distribuciones horizontales; con este punto de vista propone la idea de intentar establecer alguna correlación entre los coeficientes de similaridad o la diversidad con el logaritmo de la distancia. También se hizo el intento de descubrir alguna relación entre la diversidad y los cambios estacionales en la proporción de especies dominantes, tal y como se indica en la figura 4, ya discutida, donde se muestran los valores mensuales de la diversidad total, así como los correspondientes a un pretendido análisis del espectro de la misma a través del tiempo; por lo que se refiere a los primeros, los valores varían dentro de límites no muy amplios y además no se manifiesta algún cambio estacional más o menos claro como sería deseable esperar, pues sólo se hace aparente una tendencia a la disminución observada de enero a marzo y una posterior recuperación con altibajos tendientes a estabilizarse hacia un valor cercano a 3.5 bits. Por lo que respecta a la integración sucesiva de los valores mensuales de la diversidad, se nota un incremento notable en el valor global sólo durante los tres primeros muestreos y posteriormente este aumento sólo es gradual hasta alcanzar un total de 4.12 bits. Con el interés de encontrar alguna relación entre la diversidad y la abundancia de las especies dominantes se hizo la comparación de los valores de ambos de la manera ya indicada en la figura 4; sin embargo, se observó que no es tan fácil lograrlo a causa de la cantidad de procesos que intervienen y consecuentemente se llega a la conclusión de que la posible relación, si existe, no es tan simple y directa como para ponerla en evidencia con este procedimiento, ya que existe mucho ruido en el sistema y cuando más, parece notarse una incipiente relación inversa entre los valores de diversidad mensuales y la abundancia de Penaeus californiensis hacia el final de la época seca y principio de la temporada lluviosa, pero es muy sutil y casi podría pensarse que se trata de una relación casual o fortuita más que de un hecho real y con significación biológica; sin embargo, en el caso de que esta relación tenga sentido biológico real, existe una explicación lógica probable en relación con la diversidad y la distribución de la abundancia entre los distintos elementos componentes de la comunidad, ya que cuando existe una fuerte dominancia por parte de una especie, debe notarse su efecto en una reducción del valor de diversidad, pues bajo similares condiciones de densidad de los individuos en una comunidad al haber muchos ejemplares de una misma especie, el contenido de información por individuo es menor y viceversa.  TABLA 1 ARREGLO TAXONÓMICO DE LAS ESPECIES SE INDICA EL NÚMERO DE INDIVIDUOS CAPTURADOS  CONTINUACIÓN TABLA 1A 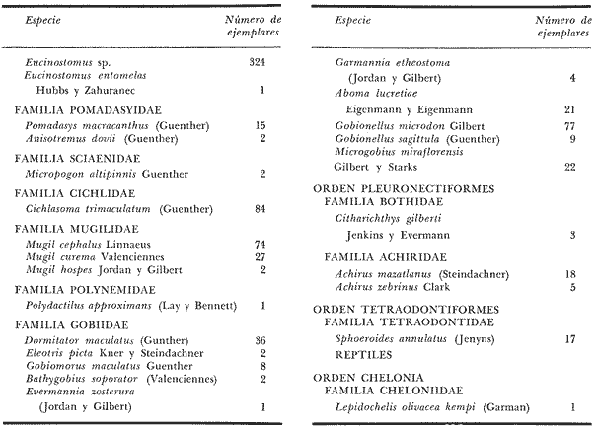 CONTINUACIÓN TABLA 1B AgradecimientosEl presente estudio se llevó a cabo en el Laboratorio de Ecología Marina de la Escuela Nacional de Ciencias Biológicas, originalmente bajo el patrocinio de la Secretaría de Recursos Hidráulicos. Es pertinente mencionar que durante el desarrollo del trabajo de campo de manera constante se recibió colaboración por parte del personal de la Residencia de Recursos Hidráulicos en Ixhuatán, especialmente Román Hernández Azpeitia, quien estaba entonces al frente de la misma, gracias a cuya intervención fue posible resolver problemas logísticos imprevistos que se presentaron en ciertas ocasiones. Rafael Barrera, miembro también de esa Residencia, colaboró en casi todas las campañas de observación. Por lo expuesto en el párrafo anterior, debe entenderse que para la integración de los datos en este trabajo se contó con la participación de varias personas, principalmente las que se mencionan a continuación, miembros todos de la Escuela Nacional de Ciencias Biológicas: María de Jesús, Parra, quien colaboró en la separación de las muestras y en la preparación de tablas y gráficas. La identificación de los peces se debe principalmente a José Luis Castro Aguirre. Asimismo, Federico Bonet Marco se encargó de leer y corregir el manuscrito; gracias a sus meticulosas observaciones y atinadas sugerencias el trabajo mejoró sustancialmente. Vaya pues mi agradecimiento a estas personas, así como a todas aquellas que participaron de un modo más informal y con cuya participación fue posible recopilar la información que ha servido de base para el presente estudio. De igual manera, quisiera aprovechar este conducto para manifestar mi agradecimiento a Ramón Margalef, de la Universidad de Barcelona, quien me permitió pasar una corta temporada en su laboratorio donde hubo oportunidad de comentar con él algunos aspectos, aclarar dudas y organizar el manuscrito. LITERATURACHÁVEZ, E. A. Mem. IV Congr. Nac. Ocean (México): Notas acerca de la ictiofauna del estuario del río Tuxpan y sus relaciones con la temperatura y la salinidad. In: Carranza, J. (Ed.) 1972. 177-199. CHÁVEZ, E. A. M. L. Sevilla y E. Hidalgo, Estudio hidrográfico de dos lagunas costeras de Oaxaca. In: Mem. V. Congr. Nac, Ocean (México): Manrique, F. A. (Ed)). 1977. 201-232. Ecology and Evolution of Communities. The Belknap, Harvard University Press. CODY, M. L. y J. M. DIAMOND (Eds.). 1975. CONNELL, J. H. Some mechanisms producing structure in natural communities: a model and evidence from field experiments. In: Ecology and Evolution of Communities. Cody, M. y J. M. Diamond (Eds.). The Belknap, Harvard Universíty Press. 1975. FISHER, R. A., A. S. CORBERT y C. B. WILLIAMS. J. Anim. Ecol; The relation between the number of species and the number of indivíduals in a random sample of an animal population. 1943. 42-58. 12: GUNTER, G. Studies on marine fishes of Texas. Publ. Inst. Mar. Sci. Univ. Texas, 1945. 1-190. (1) : 1 HUTCHINSON, G. E. Ecology and Evolution of Communities. Variations on a theme by Robert MacArthur. In: Cody, M. y J. M. Diamond (Eds). The Belknap, Harvard University Press. 1975. KJELSON, M. A. y D. R. COLBY. Estuarine Processes, Academic Press. Inc. The evaluation and use of gear efficiencies in the estimation of estuarine fish abundance. In: M. Wiley (Ed). 1976. 416-424. 2: KUSHLAN, J. A. Ecology, Environmental stability and fish community diversity. 1976. 821-825. (4): 57 LEIGH JR., E. G. The Belknap, Harvard University Press. Population fluctuations, community stability, and environmental variability. In: Ecology and evolution of Communities. Cody M. y J. M. Diamond, (Eds.) 1975. MACARTHUR, R. H. Proc. Nat. Acad. Sci. On the relative abundance oí bird species. 1957. 293-295. 43: MARGALEF, R. Comunidades Naturales. Instituto de Biología Marina, Universidad de Puerto Rico. Publ. Especial. 1962. Ecological Essays in Honor of G. Evelyn Hutchinson. Trans Conn. Acad. Arts. Sci., Homage to Evelyn Hutchinson, or why there is an upper limit to diversity. In: Growth by Intussusception. Deevey, E. S. (Ed.). 1972. 211-25. 44: Ecología. Editorial Omega. 1974. -. y F. GONZÁLEZ-BERNÁLDEZ. Investigación Pesquera, Grupos de especies asociadas en el fitoplancton del mar Caribe NE de Venezuela. 1969. 287-312. (1): 33 MAY, R. M. Ecology and Evolution of Communities. The Belknap, Harvard University Press. Patterns of species abundance and diversity. In: Cody, M. L. y J. M. Diamond (Eds.). 1975. 81-120. PATRICK, R. Ecology and Evolution of Communities. Structure of stream Communities In: Cody, J. M. y J. M. Diamond (Eds.) The Belknap. (Harvard University) Press. 1975. 445-459. PIELOU, E. C. Wiley Interscience. Ecological Diversity. 1975. POOLE, R. W. An introduction to quantitative ecology. Mc Graw-Hill. 1974. PRESTON, E. W. Ecology, The commoness and rarity oí species. 1948. 254-283. 29: SANDERS, H. L. Am. Naturalist, Marine benthic diversity: a comparative study. 1968. 243-282. 102: SMITH, F. E. Growth by Intussusception. Ecological Essays in Honor of G. Evelyn Hutchinson. Trans. Spatial heterogeneity, stability, and diversity in ecosystems. In: Deevey, E. S. (Ed.). Conn. Acad. Arts. Sci; 1972. 291-305. 44: SOKAL, R. R., y P. H. SNEATH. Principles of NuTaxonomy. Freeman. 1963. SOUTHWOOD, T. R. E. Theoretical Ecology, Principles and Applications. Blackwell. Bionomic strategies and population parameters. In: May, R. M. (Ed.). 1976. 26-48. WILLIAMS, C. B. Patterns in the Balance of Nature, and Related Problems in Quantitative Ecology. Press. Inc. New York. 1964. 45-58 (1979) (2): 6
|