|
DIAGNOSIS DE LA PESQUERÍA DE CAMARÓN DEL
GOLFO DE TEHUANTEPEC, PACÍFICO SUR DE MEXICO
Trabajo
recibido el
27 de marzo de 1978 y aceptado para su publicación el 6 de
septiembre de
1978.
Ernesto A. Chávez
Escuela Nacional de Ciencias Biológicas,
Laboratorio de Ecología Marina, Apartado Postal 42-186,
México
17, D.F.
Con el propósito de determinar el estado de
explotación de la pesquería de camarón en el Golfo
de
Tehuantepec, tanto de las lagunas como de alta mar, se hizo una
síntesis
de la información analizada; se tomó como punto de partida
el
conocimiento de la estructura por edades de las poblaciones en estudio.
La
mortalidad total calculada para el camarón café de alta mar
oscila entre Z = 0.318 y Z = 0.593 hecha sobre la base de una tasa de
cambio
mensual. En aguas interiores no fue posible separar los datos
correspondientes
a cada especie y los valores del coeficiente de mortalidad
instantánea
total, estimados con base en una tasa de cambio semanal fluctúan
entre Z
= 0.05 y Z = 0.22. En este caso se determinaron los valores del
coeficiente de
capturabilidad y mortalidad natural y son los siguientes: q = 0.00046 y M
=
0.096 y M = 0.071.
Se aplicó el modelo de reclutamiento de Ricker
encontrándose los siguientes valores de sus parámetros:
alfa =
5.148 y q = 0.0001656. A partir de las diferencias entre la abundancia de
adultos y la de reclutas se determinó una curva de rendimiento
sostenible. Del examen de los resultados se concluye que el valor del
rendimiento, máximo, aunque es bajo, está aún dentro
de
los niveles de rentabilidad económica.
Con los valores de mortalidad y capturabilidad
previamente estimados se aplicó el modelo de rendimiento de
Beverton y
Holt encontrando que el ajuste para la población de camarón
café de alta mar no es bueno; y en cambio, el que se logró
para
la población de las lagunas es bastante lógico, pues
señala que el rendimiento máximo se alcanza con una
mortalidad
por pesca de F = 0.85 que corresponde a unos 89 pescadores por noche y a
una
edad de captura de unas 15 semanas; es decir, inmediatamente antes de que
los
camarones abandonen las albuferas.
The study was undertaken with the aim to determine
the situation of exploitation of shrimp resources at the Gulf of
Tehuantepec at
west Southern Mexico. Shrimps are fished offshore and at the lagoons.
Catch and
effort data of off shore and inshore fish. ery were used; unfortunately
only
one year data was available for the analysis. Previ ously known growth
parameters for white and brown shrimps were used and age structure was
determined. Total mortality coeficient estimated for brown shrimp
population
lies between Z = 0.318 and Z = 0.593 (estimations are based upon a
monthly
change rate). At inshore waters, estimation of total mortality coeficient
includes both species, whose values, based on a weekly change rate are
between
Z = 0.05 and Z = 0.22. Catchability coeficient and natural mortality
vales
estimated for inshore shrimp populations are q = 0.00046 and M = 0.096
and M =
0.071.
Recruitment was analyzed through the Ricker's model,
whose parameter values are: alfa = 5.148 and q = 0.0001656. A sustainable
yield
curve was obtained for offshore fishery of brown shrimp based upon
differences
between adults and recruits abundances. The maximum sustainable yield
levey
appears to be low, but still within the profitable economic
level.
With known values of growth rate, mortality and
catchabílity coeficient previously estimated, Beverton and Holt
model
was applied in order to determine yield per recruit. The model did no fit
weIl
the offshore data, but the inshore fishery data fit reasonably well,
concluding
that maximum yield can be reached with a fishing mortality of F = 0.85
corresponding to near 89 fishermen per day catching 15 week-old shrimps
as
first age catch; this is, immediately before leaving the
lagoons.
El presente trabajo se realizó como parte de un estudio, más amplio llevado a cabo sobre dos pequeñas lagunas costeras del Estado de Oaxaca, donde se lleva a cabo la explotación de camarón juvenil, aunque con procedimientos que hacen de ella una pesquería poco eficiente. El objetivo original del proyecto era determinar la época de entrada de postlarvas de camarón a las albuferas, lo cual permitió al mismo tiempo, recopilar información sobre la pesquería tanto en las lagunas como en mar abierto. Se ha considerado conveniente determinar el estado de las poblaciones en explotación a fin de contribuir a llenar las numerosas lagunas que existen en el conocimiento de este recurso y poner algunas bases científicas sobre las que se puedan apoyar en el futuro las medidas más recomendables para el manejo racional de la pesquería de camarón en el área de estudio. El esfuerzo de investigación que durante los últimos años se ha estado haciendo en México sobre este recurso ha permitido reunir un cúmulo de datos tal que el conocimiento de la dinámica de las poblaciones de camarón avanza a pasos agigantados, principalmente en el Golfo de California, donde se ha concentrado la mayor parte de las investigaciones debido a que en esa región se captura el 50% de la producción nacional de peneidos. De acuerdo con lo antes expuesto, ya se han logrado estimar previamente los valores de algunos de los parámetros importantes de las tres principales especies de camarón del Pacífico, los que han servido de base para complementar la información obtenida para la realización del presente estudio donde casi no existen trabajos de investigación pesquera. Es de esperarse que el presente estudio de las poblaciones de camarón sirva de estímulo para fomentar la investigación y análisis más profundo de los recursos pesqueros del país con nuevos procedimientos y más información que al ser obtenida de manera continua y permanente permita asegurar un manejo adecuado y racional de los mismos. Los datos que han servido como punto de partida han sido el conocimiento de la estructura por edades de las poblaciones en estudio, determinada por medio de los muestreos, de los datos estadísticos de la empacadora y del conocimiento previo de la tasa de crecimiento, la que se determinó con base en el modelo de Bertalanffy. Los resultados de los muestreos han permitido seña lar la edad a la que los camarones emigran al mar desde las albuferas y esto. ha servido como criterio importante para tomar en cuenta en algunas etapas del análisis (Fig. l). 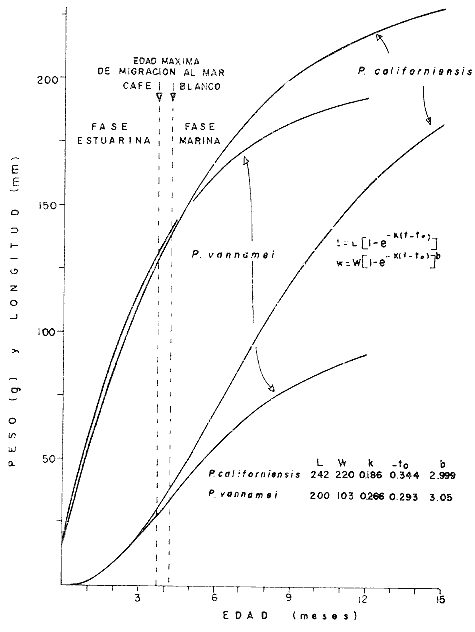 Fig. 1. Descripción gráfica de las tasas de crecimiento de las dos especies de camarón estudiadas. Se indica el tiempo que permanecen en las albuferas como juveniles y en el mar durante su etapa adulta. Los análisis que se llevaron a cabo consisten en cálculos de la mortalidad como antecedente para evaluar el valor de la captura máxima sostenible. MORTALIDADLa estimación de la tasa de decremento del número de individuos en una población a través del tiempo se logra mediante muestreos más o menos rutinarios y permanentes. La evaluación de este parámetro es imprescindible para el entendimiento de la dinámica de una población, producción, rendimiento y manejo racional de cualquier pesquería (Robson y Regier, 1968); sin embargo, la inaccesibilidad de los recursos pesqueros a la observación directa en sus hábitats naturales origina problemas no encontrados en otras situaciones por lo que ello ha dado lugar al desarrollo de una serie de métodos más o menos elaborados tendientes a evaluar las poblaciones y los cambios en su composición. Para fines prácticos, la mortalidad de las poblaciones pesqueras suele diferenciarse en dos grupos principales, mortalidad natural y mortalidad por pesca; aunque, según Nikolskii (1969) las causas principales de la muerte se pueden clasificar más o menos arbitrariamente como causadas por la edad, por la influencia de los factores ambientales, por depredadores, parásitos y enfermedades, por falta de alimentos y por pesca. La mortalidad total de cada especie fluctúa alrededor de un cierto valor, el cual está determinado, a su vez, por otras características intrínsecas de la población, de tal manera que por ejemplo, en aquellas que manifiestan una alta fecundidad la mortalidad también es muy alta; además, cuando se conjugan estas dos características, generalmente existe corta longevidad. La fórmula empleada para determinar la mortalidad es la siguiente: Nt = No e-zt en donde: No = Número inicial de individuos en la población. Nt = Número final de individuos en la población después de un tiempo t. t = Tiempo, en meses en este caso. z = Coeficiente de mortalidad total instantánea. Dicha fórmula describe una función exponencial que corresponde a una curva con pendiente negativa (z). El cálculo del coeficiente de mortalidad total puede hacerse, en especies que viven varios años, a base de una sola muestra representativa y en donde se conozca tanto la composición por edades como la proporción de cada clase de edad. Sin embargo, en especies anuales y de alta mortalidad como es el caso de los camarones, resulta inadecuado intentar el cálculo de esa manera y por lo tanto se establece la necesidad de efectuarlo haciendo estimaciones indirectas de la densidad de población. Esto se llevó en primer término, transformando los datos de maquila expresados en peso, a números de individuos, y en segundo término calculando el número de camarones de cada edad (previamente calculada con el modelo de Bertalanffy) capturados por viaje de pesca. La información de alta mar disponible permitió hacer las estimaciones de mortalidad sólo en la población de camarón café. Con las transformaciones previas se aplicó la fórmula de la mortalidad y se encontraron cuatro valores correspondientes a otros tantos cálculos realizados sobre una base mensual, que oscilan entre - z = 0.318 y - z = 0.593; las curvas que describen se muestran en la figura 2. El análisis de la mortalidad en la pesquería de aguas interiores fue hecho con base en los datos de captura y esfuerzo diario; es decir, siguiendo la declinación de la captura por unidad de esfuerzo, con la particularidad de que incluye a ambas especies en conjunto pues no hubo manera de separarlas. Estos datos se agruparon en periodos semanales comprendidos entre enero de 1972 y febrero de 1973 y a partir del rendimiento promedio diario se hicieron varias estimaciones del coeficiente de mortalidad total (por semana) mediante el cálculo del valor de supervivencia (S = e-z). Los valores obtenidos están expresados a continuación donde se indica también el valor del esfuerzo promedio diario expresado como número de pescadores (f), observado durante cada uno de esos periodos. 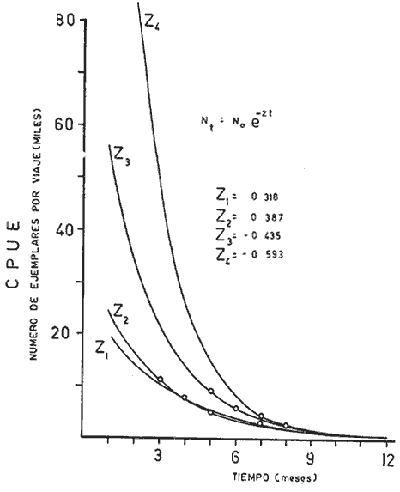 Fig. 2. Tasa de decremento poblacional del camarón café de alta mar con cuatro valores diferentes de coeficiente de mortalidad total (z) estimados a partir de la captura por unidad de esfuerzo. 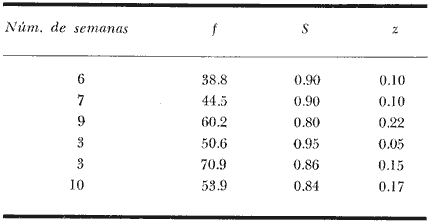 TABLA Como se puede observar, existe una gran variabilidad en el valor de la mortalidad, que no se ajusta estrictamente a cambios paralelos en el esfuerzo de pesca. Esto en primer término, puede ser un síntoma de que la influencia del ambiente sobre la supervivencia es superior a la que ejerce la pesca, y más aún cuando existe además una emigración constante hacia el mar, ya que sus efectos enmascaran las estimaciones de la mortalidad y contribuyen a complicar el problema, cuya solución limita sus posibilidades por causa de la escasa información estadística que se pudo recopilar y por tal motivo se procedió a depurar un poco los resultados obtenidos. Por otra parte, debe recordarse que el coeficiente de mortalidad total tiene dos componentes: la mortalidad total y la mortalidad por pesca, de ellas, la segunda es el producto del coeficiente de capturabilidad por el esfuerzo de pesca; se hizo una estimación de dicho coeficiente encontrándose un valor de q= 0.00046 y al estimarlo fue posible conocer también el de la mortalidad natural que fue de M = 0.09628, correspondiente a una tasa semanal. Este valor de M parece ser algo grande, lo cual se atribuye no sólo a la mortalidad que de por sí es muy alta, sino también a la constante emigración de los camarones hacia el mar, fenómeno que hace aparecer a la mortalidad más grande aún de lo que es. Sin embargo, se trató de llegar a este mismo punto a través de otra vía, análoga a la anterior, solo que en vez del reclutamiento se usó la mortalidad. En pares de datos de captura por unidad de esfuerzo en los que el segundo valor fuese más bajo que el primero se determinó el por ciento de la diferencia, lo cual se llevó a cabo en 26 ocasiones, y el valor medio de esa tasa de mortalidad fue de 0.07; luego se substituyó ese número en la fórmula correspondiente a la tasa de mortalidad natural (D). D= (M/z) (1-S) Los valores de z fueron los mismos que se utilizaron en el caso anterior, encontrando un valor medio de M = 0.071 que es muy similar al determinado anteriormente, por lo cual se consideró adecuado aplicarlo en los cálculos sucesivos. RECLUTAMIENTOExisten varios modelos que establecen una relación entre los adultos y su progenie. Hay pesquerías en las que esta relación es muy clara, tal es el caso del salmón. En el Camarón del Golfo de Tehuantepec es poco perceptible, ya que como se ha visto, existe reclutamiento de manera permanente y por tal motivo el fenómeno referido no se hace evidente; sin embargo, se supone que esto debe ser más factible de determinar en la pesquería de estas mismas especies en el Golfo de California, donde la influencia de los factores ambientales manifiesta una estacionalidad más marcada, lo cual se refleja en el régimen de pesca, que también muestra una tendencia estacional bastante clara en la variación de la captura por unidad de esfuerzo. A pesar de la dificultad antes señalada y con el agravante de contar con información de sólo un año, que limitó aún más las posibilidades del análisis, se hizo el intento de establecer la relación entre la población de camarón adulto y la de reclutas; en este caso se utilizaron datos de la captura por unidad de esfuerzo de camarón café en alta mar. La dispersión de los escasos valores analizados permiten suponer que el modelo que más se ajusta a la relación mencionada es el de Ricker (1954), cuya fórmula se expone a continuación: R= alfa A e-qA en donde R = número de reclutas A = número de adultos alfa = y q son parámetros propios del modelo. Después de hacer el ajuste de los datos se encontraron los valores siguientes: alfa = 5.148 q = 0.0001656 Al substituir distintos valores de A en la ecuación se encontraron los correspondientes de R según se observa en la figura 3; al mismo tiempo, sobre la base de los postulados en que este modelo se apoya, se determinó la curva de rendimiento sostenible, así como el valor donde el rendimiento es máximo. 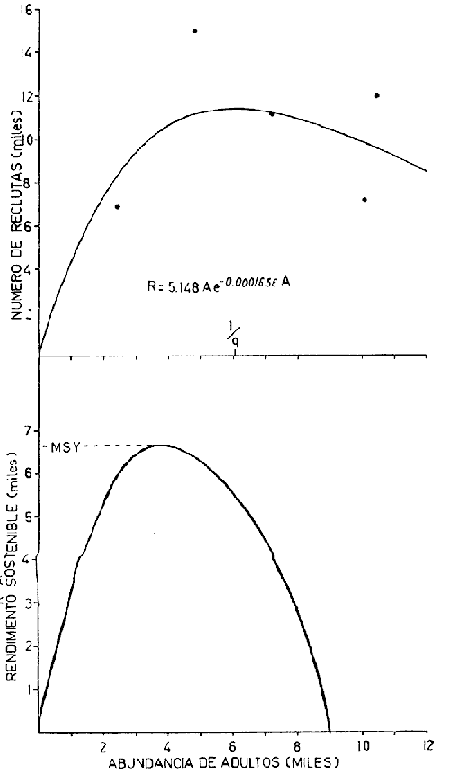 Fig. 3. Curva de reclutamiento descrita por el modelo de Ricker y su ajuste a los datos de captura y esfuerzo de camarón café de alta mar. También se determino la curva de rendimiento sostenible, así como el valor correspondiente al reclutamiento. Es pertinente hacer notar una vez más, que la intención del análisis no sólo de este aspecto, sino de varios de los que se abordan en el presente estudio llevan implícitas mejores intenciones que materia prima suficiente que sirviera de base para las estimaciones de los parámetros de la población, y apoyado en las primeras, se ha tratado de aprovechar al máximo la segunda, no siempre con resultados afortunados; parcialmente se dio esta situación en el caso del reclutamiento, pues se contó con muy pocos valores para el análisis, lo que confiere cierto grado de incertidumbre a la validez de los resultados; sin embargo, parece razonable la idea de que una población de 3,750 individuos reproductores permita mantener un nivel de captura óptimo, dada su alta fecundidad. Considerándose que se explotarán en alta mar sólo los excedentes de ese número y al mantener constante la cantidad de reproductores, los rendimientos que se logren en esas circunstancias serán los máximos posibles, aún a pesar de las conocidas fluctuaciones que año con año sufren estas poblaciones a causa de su susceptibilidad a los cambios ambientales, especialmente las variaciones en la intensidad de la precipitación pluvial. Esta circunstancia establece la necesidad de hacer una evaluación anual de las existencias para determinar, por diferencia, la cuota de captura más adecuada. Es bien sabido que la pesca de camarón en esta región es más bien baja, pero económicamente rentable. A este respecto, es oportuno citar un hecho colateral, en el sentido de que es bien conocido el comentario de los pescadores al señalar que en cada viaje de pesca se debe capturar una tonelada como mínimo para sufragar satisfactoriamente los gastos del mismo; esto ha venido sucediendo desde los inicios de esta pesquería en alta mar, cuando los rendimientos por viaje solían ser mayores aún que en la actualidad. Afortunadamente, durante los últimos años, los biólogos del Instituto Nacional de Pesca han hecho esfuerzos considerables para el conocimiento de las poblaciones de camarón de la región (Lluch, 1974 y 1977; Rodríguez De la Cruz, 1972 y 1976; Cruz Romero y Reyna, 1976; García Gómez, 1976 y Jacquemin, 1976). MODELO DE RENDIMIENTOCon la información analizada hasta ese punto, se consideró conveniente aplicar otro modelo distinto, a fin de detectar el estado actual de la pesquería y al mismo tiempo conocer la captura óptima. El modelo que se optó por aplicar es el de Beverton y Holt (1957) que originalmente fue utilizado con éxito en algunas pesquerías del mar del Norte. Como datos de entrada requiere la tasa de crecimiento de los individuos, y la mortalidad natural. La ecuación de rendimiento es la siguiente: 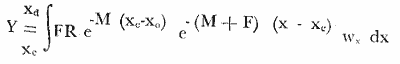 en donde: Y = Rendimiento F = Mortalidad por pesca M = Mortalidad natural x = Edad xo = Edad inicial xc = Edad de captura Ro = Captura de individuos con edad xo Wx = Peso individual a la edad x El cálculo del rendimiento se llevó a cabo con la población de camarón café en mar abierto y con las poblaciones de ambas especies, mezcladas, de las lagunas. Para el primer caso se usó un valor de M = 0. 11 y para el segundo de M = 0.33, mensual en ambos casos, substituyendo en Wx los valores correspondientes a la media ponderada de la tasa de crecimiento de ambas especies en la proporción ya conocida. De acuerdo con los cálculos llevados a cabo substituyendo cada vez un valor distinto de F para todos los valores posibles de x se obtiene un conjunto de valores de Y, observándose que esta función alcanza valores máximos cuando la mortalidad por pesca y la edad mínima de reclutamiento son altos, según se puede observar en la figura 4 que corresponde a la pesquería de aguas interiores. En dicha figura se indica el punto que señala el valor de captura máxima sostenible y el punto que muestra el estado actual de la pesquería; en ella se hace evidente que la explotación, tal como se realiza actualmente, es bastante ineficiente y que podría mejorar sensiblemente aumentando la luz de la malla e intensificando el esfuerzo de pesca para alcanzar niveles óptimos. Se hace notar el hecho de que la predicción es razonable, en el caso de la pesquería de aguas interiores, ya que el valor de la captura óptima corresponde a la edad inmediatamente anterior a la de emigración, lo cual según la predicción corresponde a un valor de mortalidad por pesca de 0.85 aproximadamente éste a su vez a un esfuerzo de unos 89 pescadores por noche de pesca con redes que tengan la malla considerablemente más abierta para que la edad mínima de captura sea casi tres veces mayor que la actual. Para esto último es necesario realizar aún bastante trabajo experimental sobre la selectividad de las redes y así poder estar en condiciones de recomendar el tamaño de luz de malla adecuado. Barrera Huerta (1976) ha hecho un intento en este sentido y aunque es empírico, también propone un aumento en el tamaño de las mallas. 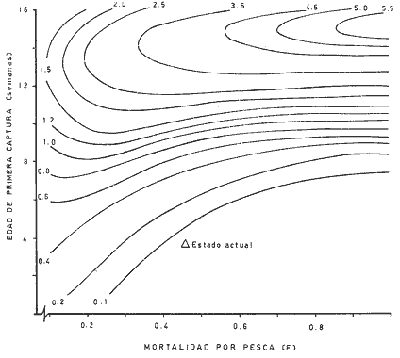 Fig. 4. Isopletas de rendimiento de la pesquería de camarón juvenil de las lagunas Oriental y Occidental (P. californiensis y P. vannamei en proporción de 6.3: 3.7). Mortalidad natural (semanal) = 0.065. Por lo que se refiere a la pesquería de camarón café en alta mar, en la que también se aplicó este modelo de rendimiento, los resultados son muy poco prometedores, y no precisamente al respecto de la mortalidad por pesca óptima de la predicción, que al compararla con la observada previamente manifiesta gran diferencia; el problema realmente radica en que de acuerdo con este resultado, la edad de primera captura debe ser a los trece meses de edad, lo cual resulta absurdo, pues a esa edad sobrevive sólo una fracción mínima de la población. Este problema ya había sido observado al tratar de aplicar el modelo a la pesquería de camarón blanco del Golfo de Campeche (Schultz y Chávez, 1976) en donde los resultados también mostraron un sesgo en el mismo sentido. El valor de M que se aplicó en este caso fue tomado originalmente del trabajo de Lluch (1974), aunque modificado en 0.03. LITERATURABARRERA-HUERTA, R. Algunos aspectos a considerar para la determinación de vedas, apertura y cierre de tapos y reglamentación de la luz de la malla de las atarrayas de pesca de camarón en las lagunas y marismas del estado de Oaxaca Mem. Simp. Biol. Dinam. Pobl. Camarones,. Guaymas (Méx.). 1976. 124-130. 1: Agosto 8 al 13, 1976. BEVERTON, R. y S. J. HOLT. On the dynamics of exploited fish populations. Fishery Investigations. Ser. 1957. 533 p. 11, 19: CRUZ-ROMERO, M. y I. REYNA. Mem. Simp. Biol. Dinam. Pobl. Camarones, Análisis de algunos parametros poblacionales del camarón de alta mar de Salina Cruz, Oax. Guaymas (Méx.). 1976. 387-408, 1: 9 figs. Agosto 8 al 13 1976. GARCIA-GOMEZ, M. Fecundidad del camarón café (Penaeus californiensis) y camarón azul (Penaeus stylirostris), de Puerto Peñasco, y Guaymas, Son. Mem. Simp. Biol. Dinam. Pobl. Camarones, Guaymas (Mex.), 1976. 131-139, 7 figs. 1: Agosto 8 al 13, 1976. JACQUEMIN, P. Estimación de algunos parámetros poblacionales de tres especies de camarón del Pacífico mexicano. Mem. Simp. Biol. Dinam. Pobl. Camarones, Guaymas (Méx.). 1976. 2: Agosto 8 al 13, 1976, LLUCH-BELDA, D. La pesquería de camarón de altamar en el noroeste. Un análisis biológico pesquero. Instituto Nacional de Pesca (S.I.C.). Serie Informativa INP/SI: 1974. 116. LLUCH-BELDA, D .- Diagnóstico, modelo y régimen óptimo de la pesquería de camarón de alta mar en el noroeste de México. Tesis Doctoral. Escuela Nacional de Ciencias Biológicas, I.P.N., México. 1977. 430 p. NIKOISKII, G. V. Theory of fish Population Dynamics. Oliver y Boyd. 1969. RICKER, W. E. Stock and recruitment. J. Fish. Res. Bd. Can. 1954. 559-623. 11: ROBSON D. S. y H. A. REGIER. Estimation of population number and mortality rates, In: W. E. Ricker (Ed.): Methods for Assessment of fich Production in Fresh Waters. Blackwell IBP Handbook 1968. 124-158. 3: RODRIGUEZ-DE LA CRUZ, C. Estudio biológico estadístico sobre la pesquería de camarón del Golfo, de California INP. CPPG. Serie Técnica Núm. 1. 1972. RODRIGUEZ-DE LA CRUZ, C.- Sinopsis biológica de las especies del género Penaeus del Pacífico mexicano. Manto. Simp. Biol. Dinam. Pobl. Camarones, Guaymas (Méx.). 1976. 282-316, 37 figs 1: Agosto 8 al 13 1976. SCHULTZ, L. y E. A. CHÁVEZ. Contribución al conocimiento de la biología, pesquera del camarón blanco (Penaeus setifertu (L.) del Golfo de Campeche, México. Mem. Simp. Biol. Dinam. Pobl. Camarones, Guaymas (Méx.). 1976. 58-73, 27 figs 1: Agosto 8 al 13, 1976.
|