|
AN ANALYSIS OR GORGONIAN COMMUNITY IN A REEF CALCAREOUS
PLATFORM ON THE CARIBBEAN COAST OF MEXICO
Trabajo presentado en el "Third
International Coral Reef Symposium", Miami Florida, mayo 1977. Recibido el 2 de
febrero de 1978 y aceptado para su publicación el 28 de febrero de
1978.
ERIC JORDAN D.
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología.
Contribución 166 del Centro de Ciencias del Mar y Limnología,
UNAM.
Un cambio notorio en la composición de
gorgonáceos de una plataforma calcárea (entre la comunidad del
fondo y la de los cabezos de coral) en la laguna arrecifal de Puerto Morelos,
Quintana Roo, México, es estudiado. El cambio es significativo respecto
a densidad, composición, diversidad y equitabilidad. Aunque estas
especies son o pueden ser oportunistas, la consistencia del cambio es muy alta
para un medio ambiente homogéneo y altamente protegido. Se propone que
tormentas y nortes puedan ser una de las causas de este cambio.
A conspicuous
change in the composition of gorgonaceans in a shallow calcareous platform
-between the bottom and coral heads-, on the reef lagoon of Puerto Morelos,
Quintana Roo, Mexico, is studied. The change is found to he significative
in terms of density, species composition, specific diversity and equitability.
Even considering that the species studied are (or maybe) opportunistic,
the consistency of the change in conditions of apparent physical homogeneity
and high protection is important. The proposition, that storms and extremly
rough weather are one of the main causes of this change, is formulated
and discussed trough the paper. The study of the ecology of the gorgonians in the West Indies has been curiously limited, since this is a very conspicuous component of the coral reef community. It has been only recently (with the exception of Cary's work - 1914, 1918) that studies in this field have been initiated, mainly with study of the zonation of gorgonians by Kinzie (1973). Several authors have since contributed valuable information on species composition, distributional aspects and general ecological structure (Opresko, 1973; Goldberg, 1973; Preston and Preston, 1975). However, there is not much information on whether a given especies or ssociation of gorgonians is related to a stable or unstable environment. The purpose of this present study was to analyse the change in the composition of a gorgonian fauna on a calcareous platform, in a shallow and apparently homogeneous environment, inside a reef lagoon. STUDY AREAThe study area is located in a reef lagoon, several hundred meters north of the dock at Puerto Morelos in the state of Quintana Roo, México, on the northeast part of the coral reef barrier of the Yucatán península. The lagoon is about 600 m wide, has a maximum depth of 6 m and the bottom is covered by a thick growth of Thalassia testudinum. About 400 m shoreward from the reef barrier a calcareous platform is found in the lagoon bottom. This platform is about 400 m long and 40-45 m wide, the long axis parallel to the reef. The platform is slight1y elevated from the lagoon bottom and has a depth of 5 m (Fig. 1) .  1. Aerial view of the study area, dark patch in front of the right side of town, is the calcareous platform. Over the entire platform exists a relatively abundant scleractinian and gorgonian fauna. At the south and, the scleractinians have formed big coral heads (Montastrea annularis) which actually have surface arcas without living coral (Fig. 2) . These coral heads have an average height of 2.0 m and mean arcas of about 35 m2 The rest of the platform also has scleractinians but they are smaller, the largest ones being no more than 30 cm high and with total arcas of less than 0.4 m2 . The most abundant species in this part of the platform are Dichocoenia stokesi and Siderastrea siderea. In this work I have divided the platform into two distinct zones: the "coral heads" zone and the "bottom" zone. Of the total arca of the platform the coral head zone is about 1,600 m2 and the bottom zone is about 15,000 m2. In each of these two zones a clearly distinct gorgonian fauna is present. 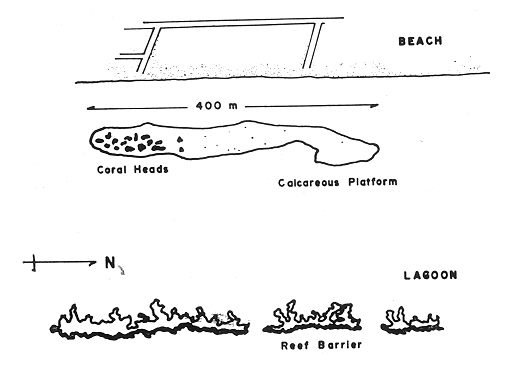 2. Diagramatic sketch of the study area. Coral heads concentration zone is show. METHODSFifteen coral heads having a total arca of 500 m2 were studied. The heads were divided in sections from 2 m2 to 4 m2 and the number of colonies of each gorgonian species was recorded. The identity and relative cover of scleractinians was also registered. Twenty five transects of 2 m by 10 m were laid at random across thebottom zone, with basic count squares every 3.3 m. Diversity was estimated according to Shanon's index (Pielou, 1966; Lloyd et al., 1968) and expressed in bits /individual. Equitability was measured by the J' value of Pielou (1966) , as were the cumulative diversities. RESULTSA total of 21 gorgonian species was registered in the study arca, of which 17 species were found on the coral heads and 15 on the platform bottom. of the 17 species found on the coral heads only 11 were common to both the bottom zone and coral heads zone. Six species were present only on the coral heads and four species only on the bottom. A total of 1,064 colonies were counted on the coral heads, while on the bottom only 687 colonies were recorded. There was, therfore, a relatively low population density in both zones i.e. coral heads = 2.13 Col/m2 , bottom 1.37 col/m2 A single dominance was evident on the coral heads (Gorgonia flabellum = 46.14% of individuals), while two species dominated the bottom (Pseudopterogorgia americana = 23.72% and Pterogorgia anceps = 20.23%. Briareum asbestinum and Plexaura flexuosa were the only important species common to both zones (roughly 15%, see table l). The diversity value for the coral heads zone was 2.45 and for the bottom zone 2.94. The significance of the difference was tested using 11 t" for diversities (Poole, 1974). It was found that they were significatively different (p < 0.01) . It can also be see in table 1, that J' was different (coral heads 0.59, bottom 0.75). These results show that the coral heads zone and the platform zone are, in fact, different in composition, density, diversity and equitability. DISCUSSIONIt has been shown that the species composition of the gorgonians in both zones is different. This is quite interesting since the area studied was very small and the hydrological regime relatively constant and homogeneous, at least in the lagoon zone where the platform is located. So, the question arises, why a relatively homogeneous gorgonian population was not found in both zones, I will now discuss some possible reasons for the difference in species composition. According to Kinzie (1973), substratum is one of the main factors to explain distribution and abundance of gorgonians. He found that specific diversity depends on substratum diversity, while the abundance is relatos to the availability of substratum. I find that there is a difference in the substratum of both zones studied. The coral heads have relatively large areas of their surface without living coral. These coral head areas show different grades of surface condition, plus a more complex spatial framework than does the bottom, which due to its flat and unperturbed surface is relatively homogeneous. The lower density on the bottom can perhaps be explained by the findings of Cary (1914) who related bottom surface texture to planula implantation. Therfore, it is not the total area of rock surface that is important for the planula implantation, but rather the cracks and depressions in the rock. The bottom zone is not just a flat surface but it is covered in most places with a thin layer of sand that shifts with currents. In fact most gorgonians grow on the sides of the small scleractinians or in cracks and small fissures without sand. Because of this, availability of substratum appears to the higher on the coral heads. The accumulative area-diversity curves (Fig. 3) show that the bottom zone curve rises faster and peaks much sooner than does the curve of the coral heads zone. This suggests that the gorgonians on the bottom find their substratum more homogeneous than do the gorgonians of the coral heads (Gleason, 1922) - It is also apparent that the gorgonians of both zones, find the availability of substratum patchy (MacArthur, 1969). This patchiness is probably more pronunced at the bottom, than it actually appears to be. This was because the size of the basic sampling area of the bottom transects was a bit too large, due to the fact that bare rock patches were very common. 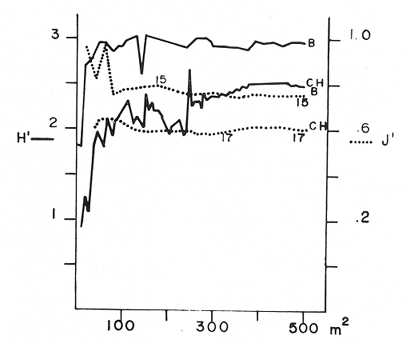 3. Cumulative area-diversity curves: Solid line show the Shannon diversity index. The dotted lines the equitability component. The numbers along the equitability curves are the number of species. CH stands for coral heads and B for the bottom. 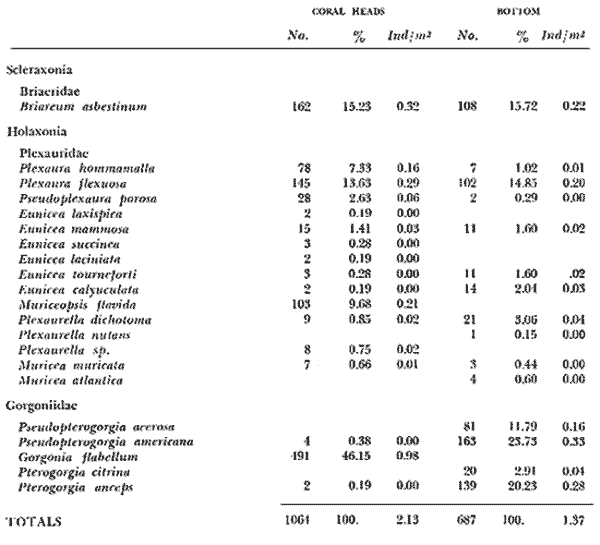 TABLE 1 PERCENTAGE AND DENSITY OF OCTOCORALS FOUND AT THE CORAL HEADS AND BOTTOM ZONES Regarding the diversity values, it can be seen that they are relatively high when compared with those found by Goldberg (1973) and Kinzie (1973). The values are similar in magnitud to other reef areas than the lagoon zone. However my values are lower than those reported by Preston and Preston (1975) for Puerto Rico. Because of the generally high diversity found, I assume that environmental conditions are basically good in the study area. However, the difference in the gorgonian diversity between the coral heads and the bottom may suggest that environmental conditions are more limiting in the coral heads than at the bottom (Margalef, 1970; Slobodkin and Sanders, 1969). That is the coral heads zone is subjected to more stress than the bottom, zone. Moreover, equitability is lower in coral heads than on the bottom. This suggests that the strees in the bottom, zones is more predictable -or less stressfull physiologically- than in the coral reads zone. This may explain why the diversity is lower in the coral heads zone, which has more diverse substratum than in the bottom zone. The most apparent difference between the two zones is that of depth. There is a maximum range of just 3.5 m between the highest coral head and the platform bottom. According to Stoddart (1969) and Kinzie (1973), the most important factors in. relation to gorgonaceans and depth are: temperature, light penetration and water movement. I do not think that temperature is a limiting factor for the coral heads and the bottom. zones, since there is almost no stratification in the water column and there is a constant movement of water along the lagoon that ensures mixing. Regarding light penetration, I think that even if there is a difference in the amount of light reaching the coral heads and the bottom, this is a high1y reflective surface and probably the amount of light available in any point in the water column of the study area, is between certain limits homogeneous enough to be of no selective value. This point is specially so when we see that all the species found in both zones are shallow water species (Goldberg, 1973; Kinzie, 1973) . With respect to water movement, oscillations and currents are important. I have no cuantitative data concerning the strength of currents nor the force of the passing waves. The distinct distributional limits of the most important gorgonian species in the two zones of the platform, led me to use them as indicators of the degree of water movement. Figure 4 Figure 5 There are, with the exception of Kinzie's work (1973), very few data on the relation between the degree of water movement and species distribution in the West Indies. Goldberg (1973), Opresko (1973), Preston and Preston 1975), collected some general information on the topic, but I found that I could not use their results to establish a relation between species and energy levels. Other authors like Bayer (1961), Chamberlain (1966), González-Brito (1970, 1972) and Rees (1973), gave mostly bathymetric data. The majority of studies on currents have been concerned with the orientation of colonies (Theodore and Denizot, 1965; Barham and Davies, 1968; Wainwright and Dillon, 1969).  6. View of one of the big coral heads, a smaller one in the background All of the important species found on the coral heads were considered by Kinzie (1973) to be able to stand strong surge movements. In this present study, Gorgonia flabellum and Muriceopsis flavida did not appear in the bottom zone, while Plexaura homomalla and Pseudoplexaura porosa were rare at the bottom. On the other hand Pseudoterogorgia americana, Pterogorgia anceps and Pseudoterogorgia acerosa, wich were dominant or important species at the bottom. zone (see Table 1) are rare or do not exist on the coral heads. In order to, determine if the gorgonian species on the coral heads showed a significant relation with depth, the abundance of the most important species in the coral heads zone was correlated to the height and area of the coral heads. A positive correlation (r = 0.81, p < 0.01) with coral heads, and no correlation with the area (r = 0.30, p < 0.0 1) , was found for Gorgonia flabellum. A similar trend was found for Muriceopsis flavida, Pseudoplexaura porosa and Plexaura homomalla. Although the correlation is barely significant in the best case the probabilities are low (p < 0.05- 0.0 l). I think for that reason that the trend shown cannot be ignored. Moreover, the consistency of the octocoral composition in the coral heads and the bottom zones is clear (Figs. 4-5). It is very interesting that with Briareum asbestinum and Plexaura flexuosa, which are the only important common species, an inverse effect is observed, i.e. the relation with the area is significative for Briareum asbestinum, and no correlation with height or area for Plexaura flexuosa was found. It is important to point out that the dominant species of the bottom Pseudoterogorgia spp (Kinzie, 1973) are typícal of low energy water movement and the those species are not present at the coral heads. In normal conditions waves and currents are small in the study area. Only in bad wheather, such as during strong "Nortes" or storms when waves and currents reach their maxima, can a condition of stress arise. Other points that support this hypothesis are (a) colony sizes are on the average smaller in the coral heads than in the bottom. (b) and Gorgonia flebellum colonies present an orientation of the blade according to the south-north movement of the current. Which of the two factors (surge or current) is more important, is hard to say on the basis of the present information. However, I believe that both factors acting together are important.  7. View of the bottom of the calcareous platform, showing an almost bare area. In summary, strong water movement is a very possible stress factor in relation to the overall diversity and equitability of gorgonians on the coral heads. Water movement may also be a determining factor in the composition of the gorgonians on the coral heads, while the lower density of gorgonians at the platform zone is due to the fact that available substratum is scarce. AgradecimientosAKNOWLEDGMENTSI wish to express my appreciation to R. S. Nugent (formerly with UNESCO) and also to E. Boschi, C. Kensler of UNESCO and J. Sarukhan and A. Lot of UNAM who kindly reviewed and criticized the manuscript. Thanks are also given to Y. Hornelas and F. Medina for their help in data processing. L. Castañares and R. Ibarra colaborate in the sampling process. LITERATURABARHAM, E. y J. DAVIES Gorgonians and water motion studies in Gulf of California. Underwat. Nat. 1968 24-28 3 5 BAYER, F. M. The Hague The shallow Water Octocorallia of the West Indies Region Martinus Nijhoff 1961 373 p. CARY, L. R. Observations upon the growth rate and ecology of gorgonians Publs. Carnegie Instn. 1914 79-90 182 ---, The Gorgononaceae as a factor in the formation of coral reefs. Publs. Carnegie Instn. 1918 343-362 213 CHAMBERLAIN, C. Some octocorallia of Isla Lobos, Veracruz, Mexico Brigham Young Univ. Stud. (Geol.) 1966 47-54 13 GLEASON, H. A. On the relation between species and arca, Ecology 1922 18-162 3 GOLDBERG, W. M. Bull. Mar. Sci. The ecology of the coral -octocoral communities- off the southeast Florida coast: Geomorphology, species composition and zonation. 1973 465-488 3 23 GONZÁLEZ-BRITO, P. Una lista de los octorales de Puerto Rico. Caribb. Jour. Sci. 1970 63-69 1 10 ---, Octocoralarios de las aguas someras del Golfo de Cariaco, Venezuela Caribb. Jour. Sci. 1972 171-177 34 12 KINZIE, R. A. Bull. Mar. Sci. The zonation of West Indian gorgonians. 1973 93-155 1 23 LLOYD,J. J., H. ZAR, J. R. KARR On the calculation of information theoretical measures of diversity Am Midl. Nat. 1968 257-272 2 79 MACARTUR, R. H. Patterns of communities in the tropics. Biol. J. Linn. Soc. 1969 19-30 1 MARGALEF, R. Perspectives in Ecological Theory. University of Chicago Press. London 1970 111 p. OPRESKO, D. M. Fla. Bull. Mar. Sci. Abundance and distribution of shallow water gorgonians in the arca of Miami. 1973 535-558 3 23 PIELOU, E. C. The measurement of diversity in different types of biological collections. J. Theoret. Biol. 1966 131-144 13 POOLE, R. W. An Introduction to Quantitative Ecology. Mc Graw Hill Inc. New York. 1974 532 p. PRESTON, E. M. y J. L. PRESTON Bull. Mar. Sci. Ecological structure in a west Indian gorgonian fauna. 1975 248-258 2 25 REES, J. T. Shallow water octocorals of Puerto Rico: Species account and corresponding depth records. Caribb. jour. Sci. 1973 57-58 2 13 SLOBODKIN, L. B. y H. L. SANDERS On the contribution of environmental preditability to species diversity. Brookhaven Symp. Biol. 1969 82-95 22 STODDART, D. R. Ecology and morphology of recent coral reefs. 1969 433-498 44 THEODOR, J. y M. DENIZOT Vie et Milieu Contribution a l'étude des gorgones (1) : A propos de Forientation d'organismes marins fixes vegetaux et animaux en fonction du courant 1965 237-241 16 WAINWRIGHT, S. A. y J. R. DILLION Biol. Bull. Mar. Lab., Woods Hole On the orientation of sea fans (genus Gorgonia). 1969 130-139 136
|