|
CRECIMIENTO Y ASPECTOS POBLACIONALES DE LA LOBINA NEGRA
MICROPTERUS SALMOIDES LACÉPÉDE EN EL LAGO DE CAMÉCUARO
MICHOACÁN (PISCES: CENTRARCHIDAE)
Trabajo recibido el 1 de febrero
de 1978 y aceptado para su publicación el 28 de febrero de
1978.
MANUEL GUZMÁN-ARROYO
JOSÉ LUIS ROJAS-GALAVIZ
FRANCISCO VERA-HERRERA
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología, Laboratorio de
Limnología y Acuicultura. Contribución 163 del Centro de Ciencias
del Mar y Limnología, UNAM.
En diciembre de 1971, fue colectada una muestra de
lobina negra Micropterus salmoides Lacépede, del Lago
de Camécuaro en el Estado de Michoacán, con base en los datos
biométricos se elaboraron gráficas de distribución de
frecuencias para longitud patrón y peso total. Se estableció la
relación entre longitud y peso obteniéndose: log. W=6.0252
-3.5572 log. L y W=.000000943 L3-5572. Por medio del
análisis de escamas fue determinada la edad y el crecimiento anual,
calculándose para cada anillo su correspondencia con la longitud del
pez, de acuerdo a la relación: Lp = 3.752711 Le + 3.544307,
determinándose la composición de tamaño para cada grupo de
edad.
Para el desarrollo del Modelo de Von Bertalanffy se
obtuvieron las constantes e-k = e
5631=.5694; longitud patrón máxima
teórica L8 = 358.4 mm; peso total teórico W8= 1151.3 g; promedio
del tiempo teórico de inicio del crecimiento To = - 0.803. Con los datos
citados fue ajustada la curva de crecimiento, quedando establecida de la
siguiente manera:
Para longitud patrón; Lp = 358.4
[I-e-.5631 (T + 0.803)]
Para peso total; Wt = 1151.3 [Ie-5631 (T
+ 0.803)] 3.55
Con los valores obtenidos de la curva, fue calculada
la tasa instantánea de crecimiento y el ritmo de crecimiento
específico para longitud y peso. Se determinó también la
tasa instantánea de supervivencia (S = .2,
e-z = e-1.690) y la tasa
instantánea de mortalidad (Z = 1.609).
El máximo ritmo de crecimiento se
registró entre los 2 y 3 años de edad. El factor de
condición (K) fue calculado para los valores observados y
teóricos en relación a sexo y edad.
Se propone el uso de Q (múltiplo de 10) como
constante para facilitar los cálculos del factor de condición y
su tasa instantánea de incremento.
In December
1971 a sample of largemount black bass Micropterus salmoides
Lacépede was collected from Camecuaro Lake, State of Michoacan.
On the basis of the biometric data, graphs of distribution of frequencies
for standard length and total weight were made. The relation established
between length and weigth was: log. W = 6.0252 3.5572 log. L y W = .000000943
L3..5572. By means of analysis the age and annual growth
were determined. Each ring was compared with fish lenght and the following
relation was determined: Lp = 3.752711 Le - 3.544307; determining the
composition of sixe for each age group. In order to
develop the Von Bertanffy model the constants e-K = e-5631=
.5631 were obtained; maximum, theoretical standar length Loo = 358.4
mm; maximum theoretical total weight W8 = 1151.3 g; average theorical
time of growth commencement T. = - 0.803. With the data cited above
the growth curve was fitted in the folliwing manner: total weight
Wt = 1151.3 [I-e-.5631 (T+ 0.803)] standard length
Lp = 358.4 [I-e-.5631 (T + 0.803)]3.55
With the values
obtained from the curve the instantaneous growth rate and the specific
growth rhythm, for length and weight were culated. Also, the instantaneous
survivial rate (S = .2, e-z = e-1.609) and the
instantaneous morality rate (Z = 1.609). were determined. The maximum.
growth rate was registered between 2 and 3 years of age. The condition
factor (K) was calculated for the values observed and theorically in
relation to sex and age. The use of
Q (multiple of 10) is proposed as a constant in order to facilitate
the calculation of the condition factor and the instantaneous increment
rate. El crecimiento y la mortalidad, como parámetros poblacionales son de gran interés en el análisis de las pesquerías, permiten conocer las características de una población en un momento dado y junto con otros factores como fecundidad, reclutamiento, vulnerabilidad, captura, etcétera, permiten integrar modelos predictivos, sobre un recurso pesquero determinado (Galicia, 1976). El crecimiento se expresa fundamentalmente, como la variación de una dimensión cualquiera del individuo o de la población, en función del tiempo. Dicha función corresponde a una curva asintótica, por lo cual es una magnitud suceptible de aumentar gradualmente cada vez con una mayor lentitud, hasta aproximadamente a su máximo (Margalef, 1974), el crecimiento se suele presentar por una curva que corresponde a dimensiones medidas en tiempos sucesivos sobre una muestra de la población, es por eso que la curva de crecimiento es una característica del individuo y lo es de la población. El modelo de Von Bertalanffy es muy usual en el estudio del crecimiento de poblaciones de peces y otros organismos, tiene la ventaja de que está respaldado por un razonamiento biológico. La ecuación del modelo se basa en una hipótesis fisiológica, puesto que considera que el crecimiento en peso es el resultado de la diferencia entre factores anabólicos y catabólicos considerados como proporcionales a la superficie y al peso respectivamente. En una ecuación de crecimiento son características deseables, que el trabajo necesario para ajustarla a los datos observados no sea muy laborioso; que el número de las constantes sea mínimo; que de esas constantes deriven de razonamientos biológicos y que si se extrapola hasta edades más allá de las observadas lleve a resultados razonables (Gullard, 1971). Mortalidad y supervivencia, factores opuestos, indican cómo evoluciona la población en cuanto al número de sus componentes, la mortalidad de dinámica poblacional pesquera, se considera como total, natural y por pesca, y es uno de los factores sustractivos de la poblacional. La mortalidad natural deriva de la interacción entre el medio ambiente y la población, relaciones de predación-presa o medio ambiente-organismo, son reflejados por medio de este factor. La mortalidad por pesca es la acción humana sobre la población y dicha acción puede ser cuantificada para un mejor aprovechamiento del recurso. ÁREA DE ESTUDIOEl Lago de Camécuaro, ubicado en el Municipio de Tangancícuaro, del estado de Michoacán, entre los 19°52' de latitud Norte y los 102°11' de longitud Oeste, a una altitud de 1,750 m sobre el nivel del mar. Este lago se encuentra en un Parque Nacional establecido por decreto Presidencial, el 8 de marzo de 1941 (González y Sánchez 1961). De acuerdo a García (1973), el clima es (A) C (W1) (W) a (e) g, definido como un clima de tipo semicálido, con temperatura promedio anual de 18.5°C, el mes más cálido es mayo con 23.3°C y el mes más frío es diciembre con 14.7°C, la precipitación anual es de 900.6 mm, siendo la época de lluvias en verano. La superficie del lago se estima en 1.5 ha, con una profundidad máxima de 5-6 m; las riberas están conformadas en su mayor parte por "ahuehuetes" o "sabinos" (Taxodium mucronatum) . El lago se forma por una serie de manantiales exteriores y subacuáticos, esto hace que sus aguas sean muy cristalinas. En el fondo se encuentran 2 tipos de vegetación, algas filamentosas y Chara sp. Los troncos sumergidos y raíces que caracterizan los fondos forman un habitat ideal para la lobina negra. El lago drena a través del Río de las Adjuntas. MATERIAL Y MÉTODOS
MATERIAL DE ESTUDIOEl material consta de 29 ejemplares de lobina negra o huro Micropterus salmoides (Lacépede), colectados en el Lago de Camécuaro, Michoacán, durante los días 26 y 27 de diciembre de 1971. Por la circunstancia de que este lago es un Parque Nacional en el cual esta estrictamente prohibido pescar, fue necesario un permiso especial de pesca para que se autorizara la captura de 30 ejemplares (uno de los cuales fue descartado) a pesar del pequeño número consideramos representativo de la población de lobina, dado el tamaño pequeño del embalse y de que la población no es muy grande. Se escogió el mes de diciembre por ser el más frío y cuando presumiblemente se forma un anillo anual de crecimiento por la reducción del metabolismo, característico de esta época. COLECTASEn virtud de que no estaba permitido el uso de redes de ningún tipo y de que el uso de la caña de pescar no permite seleccionar las tallas grandes de las pequeñas, se decidió por el uso de arpón de liga mediante buceo libre, con la ventaja de seleccionar los ejemplares grandes. La pesca se realizó por la mañana hasta el mediodía, capturando 16 y 14 ejemplares en cada uno de los días. BIOMETRÍAComo un procesamiento primario con cada ejemplar se realizaron las siguientes observaciones y mediciones: a) Longitud patrón (mm) del extremo distal de la mandíbula inferior hasta la base de la cubierta escamosa de la cola. b) Longitud furcal (mm) del extremo distal de la mandíbula inferior hasta la escotadura furcal de la cola. c) Peso total (g) . d) Peso eviserado (g) . e) Sexo. f) Estado sexual. g) Estado de replexión del estómago. h) Muestra de escamas (20 aproximadamente, tomadas abajo, de la línea lateral y posterior a la aleta pectoral izquierda. PREPARACIÓN Y LECTURA DE ESCAMASLas escamas se colocaron en una caja de petripequeña, en una solución suave de jabón, durante 12 horas, posteriormente se limpiaron con un pincel de cerdas finas y amoniaco al 10%, se montaron y etiquetaron en un portaobjetos en series de 8 escamas escogidas de las 20 de la muestra. De las 8 escamas se escogió a su vez la escama más clara de acuerdo al método presentado por Sokolov y Wong (1974), con el auxilio de un microscopio estereoscópico (Zeiss) y un ocular micrométrico se realizaron las mediciones de cada anillo de la escama, que corresponde a cada marca anual, la medición se realiza desde el focus hasta cada anillo y por último hasta el borde de la escama. RESULTADOS
DISTRIBUCIÓN DE FRECUENCIA PARA LONGITUD Y PESOLos datos de longitud patrón y peso total, fueron organizados en distribución de frecuencias. Longitud: valor promedio 300 mm, valor máximo 4-95 mm y valor mínimo 272 mm, se elaboró la gráfica de la distribución de frecuencias junto con otros parámetros poblacionales (Fig. 1A) definiéndose una distribución polimodal, correspondiendo las diversas modas a los grupos de edad. Peso: valor promedio 521.8 g, valor máximo 203.0 g, valor mínimo 430.0 g, en la figura 1B se encuentra la distribución de frecuencias para peso, así como otros parámetros poblacionales, asimismo la curva presenta una distribución polimodal. RELACIÓN ENTRE LA LONGITUD PATRÓN Y EL PESO TOTALLa relación entre la longitud patrón y el peso total concuerda con un modelo logarítmico que puede expresarse por medio de la ecuación: Log. W = a Log. L + b (1) Que en su forma exponencial se expresa como: W = a L a b (2) 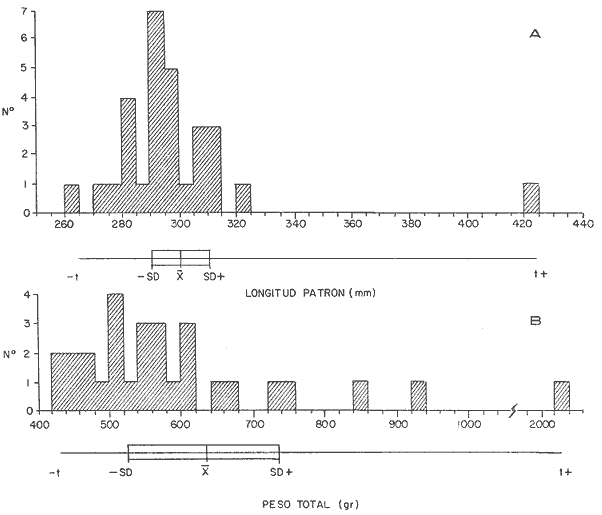 Fig. 1. Distribución de frecuencias: A Longitud patrón; B Peso total. Los valores encontrados para dicha relación son: Logarítmico: log. W -6.0252 + 3.5572 log. L Exponencial: W = .000000943 L3.5572 Cuya representación gráfica se presenta en la figura 2. Con base en los valores obtenidos de esta ecuación, se calcularon los valores teóricos, para el peso total (Tabla 1) . RELACIÓN ENTRE LA LONGITUD PATRÓN Y LA LONGITUD DE LA ESCAMAExisten diversos métodos para el cálculo, del ritmo de crecimiento, el cual podemos definir como el incremento en longitud del ejemplar en cada año de su vida. Un método sencillo es el de la proporción directa: Ln = Sn/S x L (3) 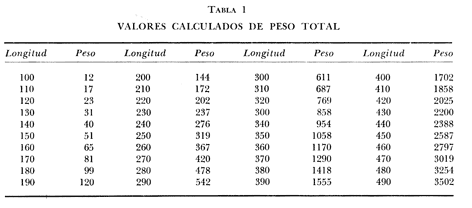 TABLA 1 VALORES CALCULADOS DE PESO TOTAL. 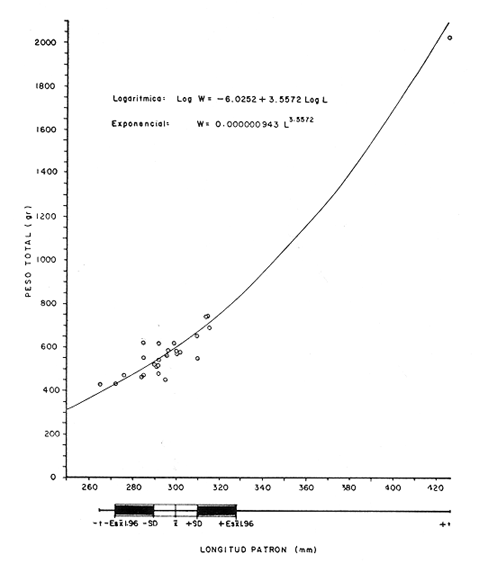 Fig. 2 Regresión longitud patrón / peso total. Donde Ln es la longitud del pez a la edad n; Sn es la longitud del anillo correspondiente a la edad n; S es la longitud total de la escama proporcional a L (longitud patrón del pez) . Para el presente trabajo se utilizó el método estadístico, mediante una regresión líneal del tipo: Y = aX + b (4) Relacionando la longitud patrón con el tamaño de la escama, a partir de los valores de la ecuación obtenida, se calcularon las longitudes patrón correspondientes a cada anillo. Lp = 3.752711 Le + 3.544307 Con un coeficiente de correlación de: r = + .8180 El tamaño de la escama fue expresado en unidades arbitrarias, la figura 3 nos muestra gráficamente esta relación. LONGITUD PATRÓNUtilizando los valores de la ecuación anterior, fue calculada para cada año la longitud patrón de cada una de las escamas, agrupando estos datos en distribución de frecuencias se elaboró la figura 4. Igualmente para cada una de las edades, se obtuvieron los valores estadísticos básicos, expresados en la tabla 2. 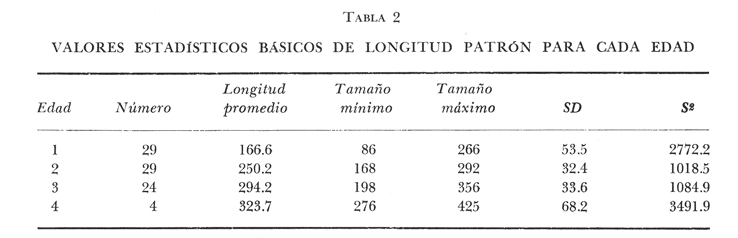 TABLA 2 VALORES ESTADÍSTICOS BÁSICOS DE LONGITUD PATRÓN PARA CADA EDAD. 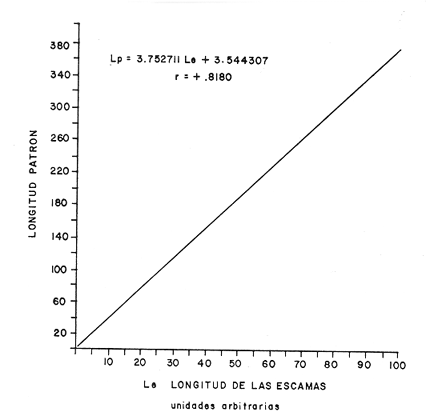 Fig. 3. Relación tamaño de la escama / longitud patrón. 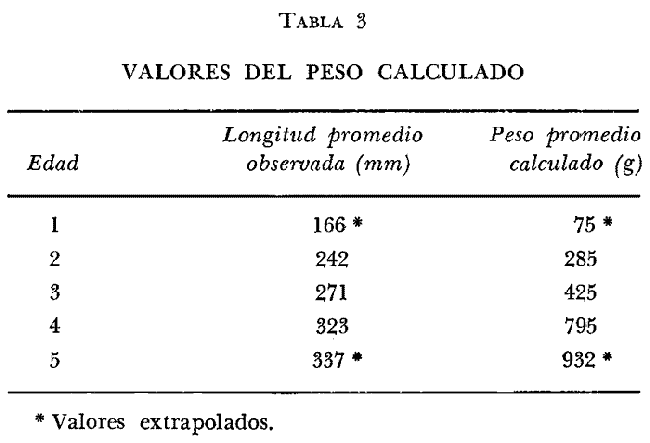 TABLA 3 VALORES DEL PESO CALCULADO. PESO CALCULADOAsimismo, se realizó el procesamiento respectivo a fin de obtener el peso calculado para cada una de las edades (Tabla 3). CRECIMIENTO EN LONGITUD Y PESO DE ACUERDO AL MODELO DE VON BERTALANFFY
Regresión lineal (n + 1/n) Para obtener las diversas constantes, es necesario obtener los valores de la regresión lineal; Longitud patrón a la edad n + 1, con longitud patrón a la edad n, la ecuación obtenida es: Ln + 1 = a Ln + b (5) Ln + 1 = .569490 Ln + 154.39 Con un coeficiente de correlación de r = + .99 el desarrollo gráfico de esta ecuación se conoce como modelo de Ford-Walford. (Fig. 5) . Determinación de la constante KTal y como se menciona en el inciso de la regresión lineal, e-k corresponde a la pendiente de dicha regresión: e-K = e -.5631= .5694 Por lo tanto el valor 1 - e-k corresponde a .4305. Determinación de las constantes L8 y W8 Para llegar a obtener el modelo de Von Bertalanfyy se requiere primero determinar ciertas constantes como es L (infinio) y W8, correspondíentes respectivamente a esta población: 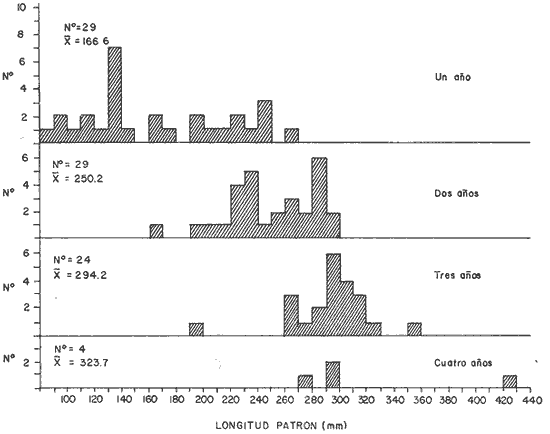 Fig. 4. Frecuencia de la longitud patrón en relación con la edad. 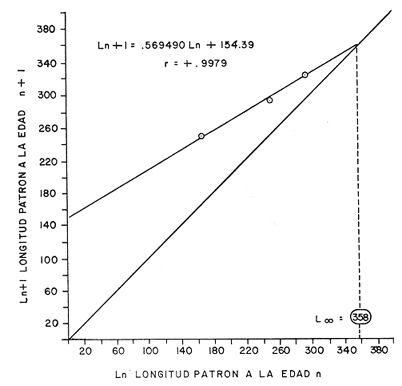 Fig. 5. Regresión longitud patrón a la edad n/n +1 (Modelo de Ford-Walford).  Fórmula (1) Donde L (infinio) es la longitud máxima teórica alcanzada para la población; Li es el valor de la intercepción entre la relación longitud patrón a la edad n, con la longitud patrón a la edad n + 1 (como se describió en el inciso referente a la regresión lineal) . Substituyendo la fórmula correspondiente tenemos: 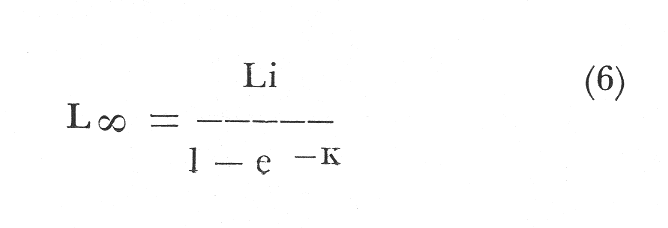 Fórmula (2) Conocido el valor L (infinio) calculamos el de W (infinio), de acuerdo a los valores obtenidos en la regresión-longitud peso; por lo tanto: L = 358.4 mm = W8 = 1151.3 gr Determinación de To
Otra de las constantes es To, que es el tiempo teórico en el cual el organismo inició su crecimiento. Este valor se obtiene para cada una de las edades mediante la fórmula; To = t + 1/e-k Log. e (L8 Lt)/L8 (7) Donde t es la edad a la que queremos saber el valor de To, y Lt es la longitud observada a la edad t; e -k y Loo, son valores ya obtenidos anteriormente. Substituyendo, para cada una de las edades los factores de la ecuación descrita se obtuvo el valor promedio para To; To = - 0.0803 Ajuste a la curva de crecimiento de acuerdo al modelo de Von BertalanffyLa expresión matemática es: Para longitud: Lp = L8 [1-e - K (t - to )](8) Y para peso: Wt = W8 [I-e - K (t - to)]b (9) Donde n equivale a la potencia en que el crecimiento de la longitud es proporcional a la del peso y se obtiene a partir de la regresión longitud patrón / peso total y corresponde al valor exponencial de la misma: W = a Lb, en este caso b. Substituyendo los valores de la ecuación tenemos: Para longitud: Lp = 358.4 [ 1 -e - .5631 (t + 0. 803)] Para peso: Wt = 1151.3 [I-e -5631 (t + 0.803) ] 3.55 Las constantes e -k y To son iguales tanto para longitud como para peso, en el caso del peso se incluye el exponente 3.55, que corresponde a la potencia, de la regresión longitud patrón / peso total (Fig. 2). Los valores obtenidos al desarrollar el modelo son los que se presentan en la tabla 4. Con estos datos se elaboraron las figuras 6 y 7 que representan gráficamente el crecimiento en longitud y en peso respectivamente.  TABLA 4 VALORES DE LONGITUD PATRÓN Y PESO TOTAL A PARTIR DEL MODELO DE VON BERTALANFFY. Tasa instantánea de crecimiento La tasa instantánea de crecimiento en longitud y peso, de acuerdo a Gulland (1971) está dada por la relación: Para longitud: TCL = Lf-Li/Tf-Ti (10) Y para peso: TCW = Wf-Wi/Tf-Ti (11) Donde Li y Wi, es la longitud y el peso al inicio de un periodo y Lf y Wf, es la longitud y el peso al final del mismo. Tf, es tiempo final y Ti tiempo inicial. Cuando este periodo de tiempo se refiere a edades mensuales o anuales, es llamado tasa instantánea y cuando se refiere al periodo que comprende, por ejemplo la vida de un organismo, se considera como tasa total.  TABLA 5 VALORES DE LA TASA INSTANTÁNEA DE CRECIMIENTO 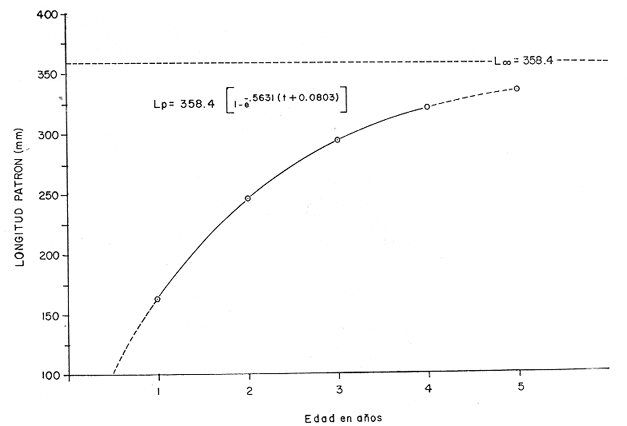 Fig. 6. Crecimiento de longitud. 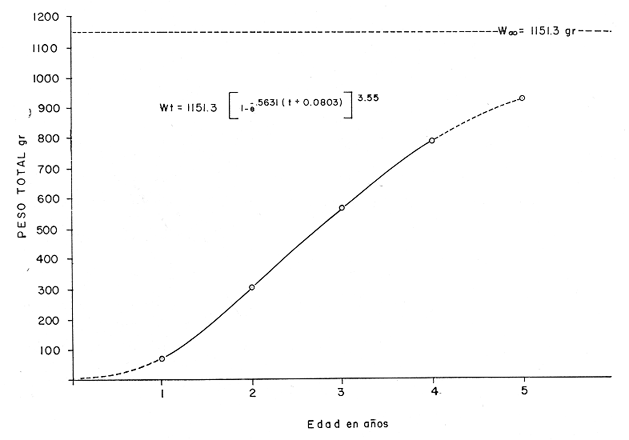 Fig. 7. Crecimiento en peso. Con base en los valores obtenidos a partir del Modelo de Von Bertalanffy se derivan los valores de tasa instantánea de crecimiento (Tabla 5), los cuales permiten elaborar la gráfica de la tasa instantánea de crecimiento. (Fig. 8-A). Ritmo de crecimiento específico Ritmo de crecimiento específico propuesto por Frost y Brown (1970), es similar a la tasa instantánea de crecimiento, la diferencia radica en el uso de logaritmos en la ecuación; y se expresa como porcentaje.  TABLA 6 VALORES DEL RITMO DE CRECIMIENTO ESPECÍFICO. 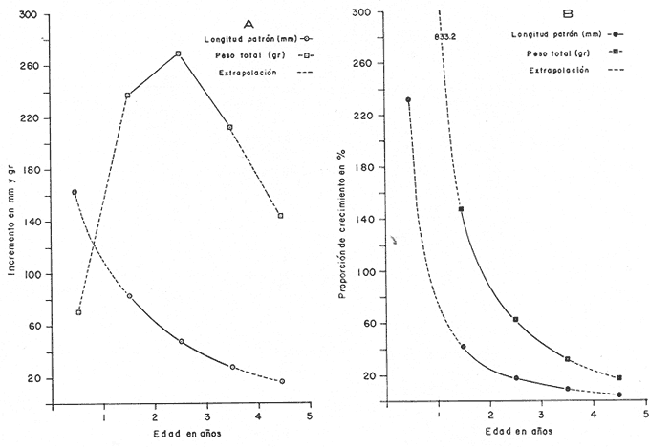 Fig.8. A. Tasa isntantánea de crecimiento; B. Ritmo de crecimiento específico. Para longitud patrón: CeL = (Log. e Lf) - (Log. e Li)/Tf-Ti X 100 (12) Para peso total: CeWt= (Log.w Wf)-(Log. e Wi)/Tf-Ti X 100 (13) Por lo tanto, con base en los valores del modelo de Von Bertalanffy, es posible derivar los del ritmo de crecimiento específico (Tabla 6), cuya representación gráfica corresponde a la figura 8B. MORTALIDAD Y SUPERVIVENCIA
Tasa instantánea de supervivencia (S) También llamado coeficiente instantáneo de supervivencia. Se considera supervivencia a la fracción de la población que sobrevive en un periodo dado. Gulland (1971), la describe como: S e-z (14) Y Ricker (1975) S = Nt + 1 / Nt (15) Mientras que Márquez (1974), la expresa como: Sx = Nx / No (16) Donde No, es el número de individuos a la edad inicial y Nx es el número de individuos a la siguiente edad, Sx, es la sobrevivencia a la edad X. La expresión de Ricker la transformamos en la de Gulland, de la siguiente manera: S = Log e Nt + 1 / Nt = e-z (17) Y lo mismo, sucede con la de Márquez: Sx = Log e No / Nx = e-z Esto es el logaritmo natural (log e) de la fracción superviviente. Para poder derivar los valores de supervivencia se requiere aplicar los valores de la estructura de edad de la muestra (Tabla 7). 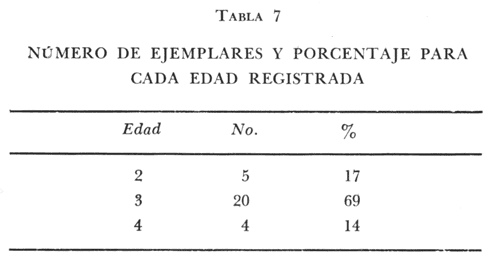 TABLA 7 NÚMERO DE EJEMPLARES Y PORCENTAJE PARA CADA EDAD REGISTRADA Así por ejemplo la tasa instantánea de supervivencia de la edad 3 a la edad 4 es: S = .2, e-z = e-1.609 Y en porcentaje es el 17% de supervivencia, ver figura 9. Para calcular el número de sobrevivientes a una edad X, Márquez (1974), usa la expresión: Nx = No. Sx (19) Multiplicando S o e-z, por el número inicial se obtiene el número sobreviviente. 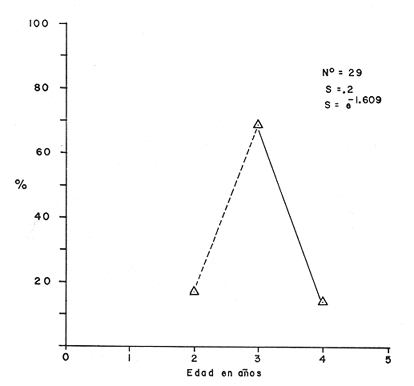 Fig. 9. Curva de supervivencia de 3 a 4 años. Tasa instantánea de mortalidad (Z) Dado que la mortalidad es el inverso de la supervivencia tenemos de acuerdo a Ricker (1975) : Z = - Log e S (20) Por lo cual, Z es igual al logaritmo natural (signo cambiado) de la fracción sobreviviente. O bien es posible calcularla directamente si se aplica: Z = - (Log e Nt + 1 - Log e Nt) (21) o Z = - Log e Nt + 1 / Nt (22) Así por ejemplo para la edad 3 a 4 el valor de Z será de 1.609 y en porcentaje el 83%, Varley ed. (1974), expresa la mortalidad como k, representada por la siguiente ecuación: k = Log Nt - Log Nt + 1 (23) Donde Log Nt, es el logaritmo del número inicial, menos Log Nt + 1 que es el logaritmo de la edad siguiente. De acuerdo a esta ecuación tenemos que, por ejemplo, el valor K para la edad 3 a 4 será de 0.69897. DATOS BIOLÓGICOS ADICIONALES Sexo y estadio sexual La estructura sexual de la muestra fue analizada en número y porcentaje correspondiente, expresándose los resultados en la tabla 8 de acuerdo a la escala de Nikolsky (Ricker 1971). 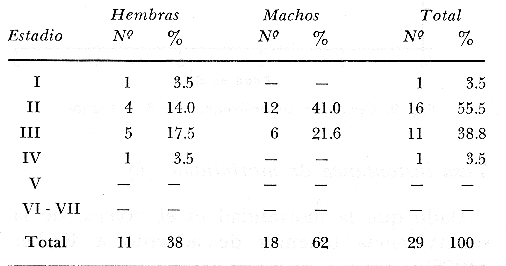 TABLA 8 VALORES DE LA ESTRUCTURA SEXUAL DE LA MUESTRA DE ACUERDO A LOS ESTADIOS PROPUESTOS POR NIKOLSKY Factor de condición (K) Utilizando los valores observados y los calculados para el modelo de Von Bertalanffy, se calculó el factor de condición de acuerdo a la fórmula de Fulton (Lagler, 1975; Ricker, 1971 y 1975; Weatherly, 1972). K = W / L3 (24) Utilizando una modificación, mencionada por algunos autores (Frost y Brown 1970), la cual consiste en multiplicar el resultado de la ecuación anterior, por un múltiplo de 10, para que su resultado sea cercano a 1 y evitar el uso de decimales, a esta constante se propone llamarla Q, y para el caso del presente estudio será: Q = 105 Quedando la ecuación (24) como: K = 1W / L3 Q, K = W / L3 105 (25) 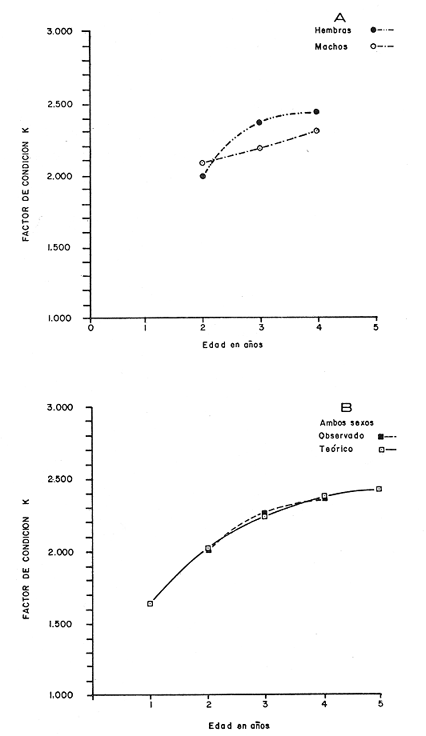 Fig. 10. Incremento del factor de de condición: A. Con la edad a sexo. B. Con la edad, observada y teórica. Los valores obtenidos para el factor de condición K de acuerdo a la edad y sexo se presentan en la tabla 9 y su representación gráfica en las figuras 1OA y B. Otro aspecto que se ha desarrollado dentro del concepto de (K) factor de condición es la tasa instantánea de incremento del factor de condición, y el ritmo de incremento específico en relación con la edad. Substituyendo los parámetros de la ecuación (10 y 11) tenemos: Ki = Kf - Ki / Tf - Ti (26) De la cual es posible derivar los valores de la tasa instantánea de K (Tabla 9) cuya expresión gráfica se presenta en la figura 11. La misma ecuación hace posible también, obtener la tasa total de incremento del factor de condición. 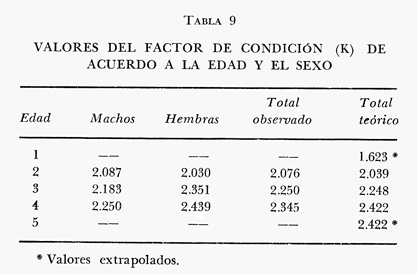 TABLA 9 VALORES DEL FACTOR DE CONDICIÓN (K) DE ACUERDO A LA EDAD Y EL SEXO. 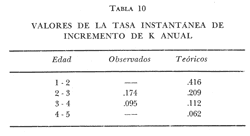 TABLA 10 VALORES DE LA TASA INSTANTÁNEA DE INCREMENTO DE K ANUAL. 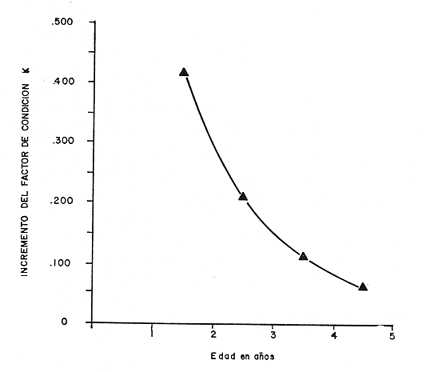 Fig. 11 Tasa instantánea de incremento del factor de condición (k) CONCLUSIONES1. El crecimiento de la lobina negra (Micropterus salmoides) en el Lago de Camécuaro, se ajusta adecuadamente al modelo de Von Bertalanffy en el cual se basó este trabajo. 2. Los parámetros obtenidos para el desarrollo de la ecuación fueron los siguientes: 2.1. Constante e -k = e- .5631 = 0.5694 2.2. Longitud patrón máxima teórica L8 = 358.4 mm. 2.3. Peso total máximo teórico W8 1151.3 g. 2.4. Promedio tiempo teórico de inicio del crecimiento To = - 0.803. 3. Las ecuaciones del modelo quedaron ajustadas: 3.1. Para longitud patrón. Lp = 358.4 [I-e - .5631 (T + 0.803)] 3.2. Para peso total. Wt = 1151.3 [I-e - .5631 (T + 0.803)] 3.55 4. De acuerdo a la tasa instantánea de crecimiento obtenida, el máximo ritmo de incremento en talla se lleva a cabo entre el año 2 y 3 de edad, etapa previa a la madurez sexual. 5. La tasa instantánea de sobrevivencia y mortalidad de la edad 3 a la edad 4 fue respectivamente: S .2 1 e-z = e -1.609 Z 1.609. lo que equivale a una sobrevivencia del 17% y una mortalidad del 83% en esta etapa. 6. El mayor porcentaje de la muestra (55.5%) quedó incluido dentro del estado sexual tipo II 7. El factor de condición K se incrementa con la edad y su representación gráfica muestra una tendencia asintótica. 8. El valor máximo de la tasa instantánea de incremento del factor de condición (K) se presenta en la edad 1 a 2. RECOMENDACIONESSe recomienda capturar los ejemplares de lobina negra (Micropterus salmoides) del Lago de Camécuaro que posean una talla promedio. de 300 mm (edad 3-4 años mediante el método de pesca de red agallera selectiva, con base en las siguientes consideraciones: A. Estos ejemplares inician una etapa de crecimiento lento y su conversión alimentaria es cada vez más elevada. B. Los ejemplares habrán realizado su primer desove, con lo cual han contribuido al incremento de la población. En el caso que se mantenga la actual reglamentación de veda a la captura, la población de esta especie muy posiblemente se verá afectada, ya que se estará favoreciendo la predominancia de ejemplares adultos que consumirán mucho alimento y crecerán poco, lo cual además plantea una competencia desfavorable para los ejemplares juveniles y depredación sobre las tallas menores determinando esta situación una baja en la eficiencia del desarrollo de la población. AgradecimientosAGRADECIMIENTOSLos autores quieren hacer patente su agradecimiento a las siguientes personas: Enrique Boschi, por su inapreciable auxilio en el procesamiento, de datos, Craig B. Kensler, por sus comentarios y supervisión del trabajo, a Leandra Salvadores por su ayuda en la preparación del material, Enrique Rubio, por su ayuda y supervisión del procesamiento de datos. LITERATURAFROST, W. E. y M. E. BROWN The Trout. Collins The Fontana New Naturalist London 1970 316 p GALICIA, X. R. Crecimiento del "Camarón Azul" (Penaeus sty1irostris) y "Camarón Café" (Penaeus californiensis) en la zona de Puerto Peñasco S. I. C., S. P., I. N. P. Son. Mem. Simp. Biol. Dinam. Pobl. Camarones, Guaymas Son. México. 1976 189-210 2 1973 GARCÍA, E. Modificaciones al Sistema de Clasificación Climática de Koppen. Inst. Geogr. Universidad Nacional Autónoma de México México 1973 246 p. GONZÁLEZ, A. y U. M. SÁNCHEZ Los Parques Nacionales de México Inst. Méx. Rec. Nat. Renov. México 1961 149 p. GULLAND, J. A. Manual de Métodos para la Evaluación de Poblaciones de Peces Ed. Acribia, Zaragoza 1971 164 p. LAGLER, K. F. Freswater Fhisery Biology. WM. C. Brown Co. Pub. IOWA. 1975 421 p. MARGALEF, R. Ecología Ed. Omega Barcelona España 1974 951 p. MÁRQUEZ, M. R. Observaciones sobre mortalidad total y crecimiento en longitud de la lisa (Mugil cephalus) en la Laguna de Tamiahua, Ver., México. Inst. Nal. Pes. INP/SC 1974 1-15 3 RICKER, W. E. Methods for Assessment of fish Production in Fresh Waters IBP. Handbook 1971 348 p. 3 ---, Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Bd. Canada 1975 382 p. 191 SOKOLOV, V. A. y R. M. WONG Prog. Inv. y From. Pesq. México/ PNUD/FAO, CEPM Programa General para las Invstigaciones de los peces pelágicos del Golfo de California. 1974 51 p. 3 VARLEY, M. E. Population Dynamics Block-B nits-6-9. The open University Press. 1974 100 p. WEATHERLEY, A. H. Growth and Ecology of Fish Populations Academic Press London 1972 293 p.
|