|
INVESTIGACIONES PRELIMINARES DE LA BIOMASA DE THALASSIA
TESTUDINUM KONIG, EN LA COSTA DEL CARIBE MEXICANO: NOTA CIENTÍFICA
Trabajo recibido el 8 de junio de 1977 y aceptado para su
publicación el 13 de junio de 1977.
RICHARD S. NUGENT
Experto de la UNESCO en Biología
Marina.
ERIC JORDAN
ROBERTO DE LA TORRE
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología.
Contribución 79 del Centro de Ciencias del Mar y Limnología,
UNAM.
Durante los meses de marzo y abril de 1974, se
inició un estudio con el objeto de realizar determinaciones de biomasa
(peso seco), de las hojas de la fanerógama marina Thalassia
testudinum Konig (1805) en el Mar Caribe, en zonas adyacentes a la
Isla Cancún, Quintana Roo, México. Las áreas de muestreo
fueron de 2 tipos, protegidas por arrecifes coralinos y zonas costeras sin
protección; con un total de 6 estaciones. En cada estación se
tomaron cinco muestras de hojas de Thalassla testudinum de
cada una de las siguientes profundidades: 1 metro, 3.5 metros y 7
metros.
El promedio de la biomasa de las hojas (peso seco), de
las muestras obtenidas a 1 metro de profundidad fue de 40.7 g/M2 (rango 25.0 -
60.1 g/M2), para las muestras de 3.5 metros, la biomasa fue de 43.2 g/M2
(rango: 13.1 - 92.8 g/M2). Finalmente para las muestras de las zonas más
profundas, 7 metros, el promedio de biomasa fue de 44.8 g/ni2 (rango 30.2 -
66.6 g/m²), considerando todos los resultados obtenidos el rango de
biomasa se estableció entre 13.1 g/m2 y 92.8 g/m², con un valor
promedio de 42.9 g/M2, peso seco.
Estos valores son considerablemente más bajos
que los obtenidos por otros investigadores en otras regiones del Caribe y zonas
adyacentes.
Dry weight leaf biomass determinations were made on
samples of the marine fenerogam Thalassia testudinum Konig,
collected during March and April, 1974 along the shore of the Caribbean sea
near Isla Cancún, Quintana Roo, México.
Two arcas were sampled: one in a relatively protected
zone behind a fringing coral reef and the other in an open, unprotected coastal
area. Three depths were sampled in each area: 1 meter, 3.5 meters and 7 meters.
Five samples of 0.25 m² were taken at each depth.
All the Thalassia leaves were
washed in a 10 per cent phosphoric acid solution prior to being dried at
105°C.
The mean value for dry weight biomass of leaves
collected at 1 meter depths was 40.7 g/M2 (range: 25.5 - 60.1 g/M2); at 3.5
meters it was 43.2 g/M2 (range; 13.1 92.8 g/m²), and at 7 meters it was
44.8 g/M2 (range: 30.2 - 66.6 g/m²). For the entire study area the mean
biomasa was 42.9 grams dry wt./m² and the range was 13.1 - 92.8 grams dry
Wt./m².
These values are considerably less than those obtained
by other investigators in other regions of the Caribbean and adjacent
arcas.
Existen muy pocos trabajos relacionados con la distribución geográfica, abundancia, biomasa y productividad de los pastos marinos. La necesidad de obtener información básica sobre este aspecto de la ecología marina fue indicado en el International SeaGrass Workshop, convenido en Leiden (Holanda en octubre de 1973. Se recomendó tratar de recabar información sobre este aspecto en forma tal que se aumente el conocimiento sobre estas plantas. Con este propósito en mente, se comenzó un programa para estudiar este aspecto a lo largo de las costas de México, aprovechando generalmente salidas de campo destinadas a otros propósitos, que es el caso concreto de este trabajo. Desde el momento que no existe información alguna sobre Thalassia en la región de Cancún, Quintana Roo, excepto la elaborada por los autores Jordán et al.(1978), la información aquí presentada es útil. El objetivo del estudio sobre Thalassia fue planeado para determinar diferencias entre la biomasa de las hojas de plantas que crecen a distintas profundidades, en zonas protegidas y expuestas en la región de la Isla Cancún. Thalassia testudinum es una de las especies más importantes de los pastos marinos a lo largo de las costas del Caribe y el Golfo de México (den Hartog, 1970). Ha recibido gran atención desde el punto de vista de su biología (Orpurt y Boral, 1964; Tom1inson y Vargo, 1966; Tom1inson, 1969a, 196b, 1972; y Patriquin, 1973), su ecología y distribución (Phillips, 1960; Strawn, 1961; Moore, 1963; Zieman, 1968, 1973, 1974; y Patriquin, 1972). Se ha determinado que Thalassia es un componente importante de los ecosistemas costeros, influenciando estos ambientes de diversas formas (Ferguson Wood et al., 1969). Thalassia contribuye considerablemente en la cadena alimenticia de detritos (Zieman, 1973). La producción de esta especie es elevada (2.3 -5 g/M2/día, peso seco, Zieman, 1973) existiendo muy poca utilización por otros organismos marinos de la planta viva (Ferguson Wood et al., 1969, Zieman, 1973). Zieman (1973) estima que el 90% o más de la energía proporcionada por Thalassia llega a los niveles tróficos superiores a través de detritos. Otro aspecto importante de esta fanerógama marina, con respecto al flujo de energía, es su función como substrato para una gran variedad de algas epífitas (Humm, 1964; Reyes-Vázquez, 1965). Ha sido estimado que en ciertas ocasiones la biomasa de la epífita puede igualar a la biomasa de las hojas de Thalassia (Ferguson Wood et al., 1969). Estas algas epifíticas son una fuente de alimento para organismos que tienen su habitat o utilizan los bancos de Thalassia. Las praderas de Thalassia son importantes como áreas de desarrollo de etapas juveniles de peces e invertebrados y zonas de protección para otros animales marinos (Hoese, 1960; Bruer, 1962; Tabb et al., 1962; Moore, 1963; y Hoese y Jones, 1963). Una comparación de la abundancia y diversidad de organismos marinos en bancos de Thalassia y zonas contiguas revela la importancia de los pastos marinos como subambientes en lagunas costeras y áreas costeras abiertas. (Tabb et al., 1962). Una consideración final es el impacto de las praderas de Thalassia sobre la sedimentación, pues incrementan la acumulación de partículas orgánicas e inorgánicas. (Ginsburg y Lowenstam, 1958; Ferguson Wood et al., 1969; y Scoflin, 1970). Los bancos de Thalassia también ayudan a estabilizar la superficie de los sedimentos previniendo, la erosión. Existen otros aspectos sobre la importancia y función de los bancos de Thalassia en zonas cercanas a la costa, que si bien han sido reconocidos no están bien entendidos. Entre estos aspectos están: su papel en el ciclo de los nutrientes, su contribución al contenido orgánico de los sedimentos por el crecimiento de los rizomas y su descomposición además de sus efectos sobre la química del agua de mar en zonas de poca circulación. Aunque el detalle de estos procesos y sus efectos, aún no son totalmente comprendidos, es un hecho que Thalassia es un componente muy importante en ecosistemas costeros tropicales y subtropicales. ÁREA DE ESTUDIOLa región estudiada se encuentra en la parte Noreste de la península de Yucatán, frente al Mar Caribe, aproximadamente a los 86° 48' de longitud oeste y a los 21° 05' de latitud norte (Fig. 1). El substrato en toda la región es prácticamente carbonatos de calcio. El clima es cálido subhúmedo con lluvias aisladas, AW, según la clasificación modificada de Koppen. El área de estudio (Fig. 1) corresponde a dos zonas, una detrás de una barrera de arrecifes y por lo tanto protegida, al Sur de la Isla Cancún, en el área conocida como Punta Nizuc. Esta zona tiene una profundidad máxima de 8 m y el substrato es de arena calcárea alternándose con bancos arrecifales. En esta zona tanto el oleaje como las corrientes son muy pequeñas. La zona no protegida corresponde al Norte de la Isla Cancún, frente a Isla Mujeres, el substrato es también de arena calcárea, pero no hay corales arrecifales en esta zona. Fuertes corrientes se establecen con dirección Este-Oeste, y no hay protección contra el oleaje. 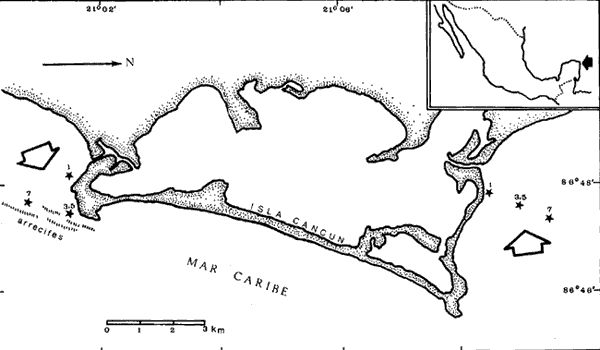 Fig. 1. Área de estudio. Localización de las estaciones de muestreo. MATERIAL Y MÉTODOSSe tomaron en cada una de las seis estaciones cinco muestras de hojas de Thalassia de bancos localizados a 1 metro, 3.5 metros y 7 metros de profundidad. Cada una de las muestras consistía de todas las hojas de Thalassia encuadradas en un área de 0.25 M2, delineadas con un marco de hierro de 0.5 por 0.5 metros. El marco o cuadrante fue colocado al azar en los bancos de Thalassia por buzos autónomos. Dos buzos trabajaron en equipo, uno de ellos cortando las hojas al nivel de la superficie del fondo y con tijeras de poda y el otro buzo colocando las hojas en una bolsa; posteriormente las hojas se secaron al aire y se colocaron en bolsas de plástico marcadas adecuadamente. En el laboratorio cada muestra se lavó con agua potable para eliminar sedimentos y otras substancias indeseables. Las muestras se lavaron en una solución de ácido fosfórico al 10% por espacio de 15 minutos hasta que no se observaron burbujas. El tratamiento ácido tiene por objeto eliminar carbonatos y algas epífitas adhehidas a las hojas de Thalassia. El procedimiento reviste gran importancia si se quiere hacer una determinación correcta de biomasa (Zieman, 1968, 1973). Después de remover las hojas del tratamiento ácido se lavaron abundantemente con agua potable. Las muestras luego se seleccionaron para eliminar otros materiales y remover el resto de las epífitas de las hojas. Las muestras se secaron en horno a 1050 C por 24 horas y se pesaron separadamente con una precisión de 0.1 gramos. RESULTADOS Y DISCUSIÓNLos resultados de esta investigación se presentan en las tablas 1 y 2. Las biomasas expresadas en peso seco de las muestras individuales variaron desde 13.1 g/m² a una profundidad de 3.5 metros para una estación costera no protegida, a 92.8 g/m² a la misma profundidad para una estación protegida. La más amplia variación de biomasa para las cinco muestras de una estación fue determinada a 3.5 metros en una zona protegida (S= 25. 1; X = 60.7). La menor variación fue determinada para una profundidad de 7 metros también de una zona protegida (S= 4.9; X = 37.6). La biomasa media más alta de las seis estaciones fue de 60.7 g/m² a 3.5 metros en la zona protegida, y la media más baja fue de 25.6 g/m² a 3.5 metros en la zona no protegida. El valor medio para todas las muestras recogidas a 1 metro de profundidad fue de 40.7 g/m² (DS = 13.9); para todas las muestras recogidas a 3.5 metros fue 43.2 g/m² (DS = 26.2) y para las obtenidas de 7 metros de profundidad 44.8 g/m² (DS = 11.1). La biomasa media considerando todas las muestras obtenidas de zonas protegidas fue de 45.6 g/m² (DS = 19.6). El valor correspondiente para zonas no protegidas fue de 39.8 g/m2 (DS = 15.8). La media de todas las muestras, treinta en total, fue de 42.9 g/n12 (DS = 17.8). El análisis de varianza indicó que estadísticamente no hay diferencias significativas (P=0.01) entre los valores de biomasa determinados en las seis estaciones. Tampoco existen diferencias significativas (P=0.01) entre las biomasas de Thalassia provenientes de zonas protegidas y no protegidas o de las tres profundidades muestreadas. Las determinaciones de biomasa de hojas de Thalassia (peso seco) registradas en la presente investigación, se comparan en la tabla 3 con los resultados de otros estudios similares. Puede observarse claramente que la biomasa de Thalassia en el área de Cancún es mucho menor que la de otras regiones estudiadas. Los resultados más cercanos a los obtenidos en esta investigación son los determinados en la zona de Biscayne Bay, Florida (Zieman, 1973). Aún así estos son tres veces más altos que los valores registrados en el área de Cancún. Debe sin embargo, aclararse que a excepción de los datos de Zieman, los otros datos deben considerarse artificialmente altos debido a que no se realizó en estas muestras eliminación de carbonatos y algas epífitas. De todas maneras los valores registrados en Cancún son bajos, y es muy difícil definir las razones en un estudio preliminar como este. Sin embargo, otros investigadores demostraron que los dos factores ambientales más importantes para el desarrollo de Thalassia testudinum son la temperatura y la salinidad. Esta planta parece preferir temperaturas en el rango entre 200 C y 300 C (Phillips, 1960 y den Hartog, 1970). Zieman (1974) encontró máximos de productividad a 300 C. El rango de salinidad óptima parece encontrarse entre 24%. y 35%, (Phillips, 1960 y den Hartog, 1970) con un máximo de productividad registrado a cerca de 30%, (Zieman, 1974). 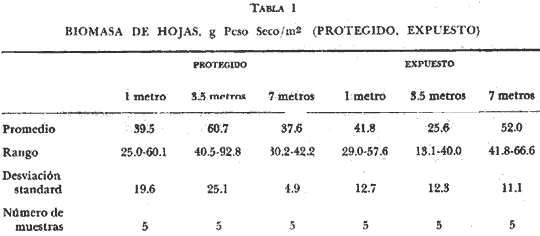 TABLA 1 BIOMASA DE HOJAS, g Peso Seco, /m2 (PROTEGIDO, EXPUESTO) 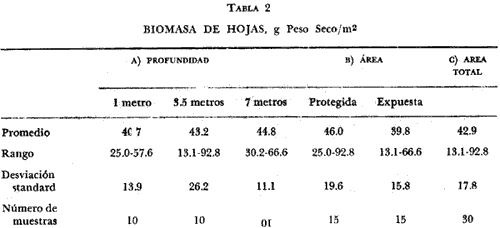 TABLA 2 BIOMASA DE HOJAS, g Peso Seco/m² 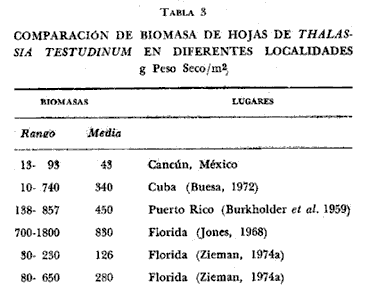 TABLA 3 COMPARACIÓN DE BIOMASA DE HOJAS DE THALASSIA TESTUDINUM EN DIFERENTES LOCALIDADES Otros factores tales como intensidad lumínica, niveles de nutrientes, características de los sedimentos y circulación, influyen sobre la distribución y densidad de los bancos de Thalassia, al presente se conoce muy poco sobre estos aspectos. Es necesario realizar investigaciones intensivas para determinar los factores ambientales responsables de las bajas biomasas de Thalassia en la zona de Cancún. Otras fanerógamas marinas se encontraron compartiendo el habitat de los bancos de Thalassia como Syringodium filiforme y Halodule beaudettes. En todas las áreas donde estos pastos se localizaron se encontraron en pequeña densidad. De acuerdo con den Hartog (1970) ambas especies se encuentran en el Caribe y Golfo de México y son especies subordinadas a Thalassia en aquellas zonas donde Thalassia se desarrolla bien Syringodium filiforme es dominante en aguas sublitorales con salinidades menores de 22%, a 25%. En aguas de mayor salinidad Thalassia no está presente. Halodule beaudettes domina sublitorales solamente donde Thalassia y Syringodium están ausentes o muy escasas. Puede considerarse como una especie de avanzada que estabiliza sedimentos sueltos para luego éstos ser colonizados por Thalassia. Conclusiones1. No se encontró diferencia significativa (P = 0.01) entre las distintas determinaciones de biomasa de hojas de Thalassia testudinum en las seis, estaciones muestreadas. 2. El rango de biomasas determinado varió entre 13.1 g/M2 y 92.8 g/m² (peso seco). El valor medio para toda el área muestreada fue de 42.9 g/m² (peso seco) con una desviación standard de 17.8. 3. Las biomasas de hojas de Thalassia en el área estudiada están muy por debajo de los valores obtenidos en estas áreas del Atlántico-Golfo de México y Mar Caribe. 4. Se encontraron otras fanerógamas marinas entre las frondas de Thalassia, ellas fueron Syringodium filiforme y Halodule beaudettes. En todas las áreas muestreadas el crecimiento de estos pastos fue escaso en comparación con el crecimiento de Thalassia. LITERATURABRUER, J. P., An Ecological Survey of the Lower laguna Madre of Texas, 1953-1969. Publ. Inst. Mar. Sci., Texas, 1962. 153-183. 8: BUESA, R. J., Cont. Inv. Pesqueras Reunión Bal. Trab., Producción Primaria de las Praderas de Thalassia testudium de la Plataforma Noroccidental de Cuba. INP 1972. 101-143. 3: BURKHOLDER, P. R., L. M. BURKHOLDER and J. A. RIVERO, Bull. Some Chemical Constituents of Turtle Grass, Thalassia testudinum. Torrey Bot. Club 1959. 88-93. 86 (2): GINSBURG, R. M. and H. A. LOWENSTAM, The influence of Marine Bottom Communities on the Depositional Environment of Sediments. S. Geol., 1958. 310-318. 66 (3): DEN HARTOG, C., The Sea-grasses of the World. NorthHolland Publishing Co., Amsterdam, 1970. 275 p. FERGUSON WOOD, E. J., W. E. ODUM and J. C. ZIEMAN, Lagunas Costeras. Un Simposio. Influence of Sea Grasses on Productivity of Coastal Lagoons In: Ayala, A. and F. B. Phleger (Eds.) UNAM-UNESCO. 1969. 495-502. HOESE, H. D., Ecology, Juvenile Penaeid Shrimp in the Shallow Gulf of Mexico. 1960. 592-593. 41 (3): HOESE, H. D. and R. S. JONES, Seasonality of Larger Animals in a Texas Turtle Grass Comunity. Publ. Inst. Mar. Sci., Texas 1963. 347-357. 9: Humm, H. J., Bull. Epiphytes of the Sea Grass, Thalassia testudinum, in Florida. Mar. Sci. Gulf and Carib., 1964. 306-341. 14 (2): JONES, J. A., Ph. D. Dissertation, Primary Productivity by the Tropical Marine Turtle Grass, Thalassia testudinum Konig, and its Epiphytes. University of Miami, 1968. 196 p. JORDAN, E., M. ANGOT y R. DE LA TORRE, An. Centro Cienc. del Mar y Limnol. Prospección biológica de la laguna de Nichupté (Cancún), Q. Roo, México: Nota Científica. Univ. Nal. Autón. México, 1978. 5 (1): MOORE, D. R., Bull. Distribution of the Sea Grass, Thalassia in the United States. Mar. Sci. Gulf and Carib., 1963. 329-342. 13 (2): ORPURT, P. A. and L. L. BORAL, Bull. The Flowers, Fruits and Seeds of Thalassia testudinum Konig. Mar. Sci. Gulf and Carib., 1964. 296-302. 11 (2): PATRIQUIN, D. G., Thalassia testudinum. The Origin of Nitrogen and Phosphorus For Growth of the Marine Angiosperm Mar. Biol., 1972. 35-46. 15: Thalassia testudinum Estimation of Growth Rate, Production and Age of the Marine Angiosperm. Konig. Carib. J. Sci., 1973. 111-123. 13 (1-2): PHILLIPS, R. C., Prof. Papers Ser., Fla. Bd. Conserv Observations on the Ecology and Distribution of the Florida Sea Grasses. 1960. 1-72. 2: REYES-VÁZQUEZ, G., Thesis, Studies on the Diatom Flora Living on Thalassia testudinum Konig in Biscayne Bay. Unpublished M. S. University of Miami, 1965. 81 p. SCOFFIN, T. P., J. Sed. Petrol., The Trapping and Bínding of Subtidal Carbonate Sediments by Marine Vegetation in Bimini Lagoon, Bahanias, 1970. 40 249-273. (1): STRAWN, K., J. Wildl. Mang., Factors influencing the Zonation of Submerged Monocotyledons at Cedar Key, Florida. 1961. 178-189. 25 (2): TABB, D. C., D. L. DUBROW and R. B. MANNING, Fla. Bd. Conserv. Tech. Ser., The Ecology of Northern Florida Bay Adjacent Estuaries. 1962. 1-81. 39: TOMLINSON, P. B., Bull. On the Morphology and Anatomy of Turtle Grass, Thalassia testudinum (Hydrocharitaceae). II. Anatorny and Development of the rootin Relation to Function Mar. Sci. Gulf and Carib., 1969 57-71. 19 (1): Bull. On the Morphology and Anatomy of taceae). III. Floral Morphology and Anatomy Bull. Turtle Grass, Thalassia testudinum (Hidrocharitaceae). IV..Leaf Anatomy and Development. Mar. Sci. Gulf and Carib., 1969b. 286-305. 19 (2): Bull. On the Morphology and Anatomy of Turtle Grass, Thalassia testudinum (Hidrocharitaceae). IV. Leaf Anatomy and Development. Mar. Sci. Gulf and Carib., 1972. 75-93. 22 (1): TOMLINSON, P. B. and G. A. VARGO, Bull. On the Morphology and Anatorny of Turtle Grass, Thalassia testudinum (Hidrocharitaceae). I. Vegetative Morphology- Mar. Sci. Gulf and Carib., 1969. 748-761. 16 (4): ZIEMAN, J. C., Thesis. A Study of the Growth and Decomposition of the Sea-grass Thalassia testudinum. Unpublished M. S. University of Miami, 1968. 50 p. Res. Conf. Myrtle Beach S. C., Quantitative and Dinamic Aspects of the Ecology of Turtle Grass, Thalassia testudinum. Recent Advances in Estuarine Research. Proceedings of Second Intl. 1973. Oct. 1973. Aquaculture (in press). Seasonl Variation of Turtle Grass, Thalassia testudinum Konig, with Reference to Temperature and Salinity Effects. 1974.
|