|
EVALUACIÓN POBLACIONAL DE PLEXAURA HOMOMALLA (ESPER)
EN LA COSTA NORESTE DE LA PENÍNSULA DE YUCATÁN.
(OCTOCORALLIA)
Trabajo recibido el 12 de mayo de 1977 y aceptado para su
publicación el 13 de junio de 1977.
ERIC JORDÁN
Universidad Nacional Autónoma de
México. Centro de Ciencias del Mar y Limnología
RICHARD S. NUGENT
Experto de la UNESCO en Biología Marina.
Contribución 76 del Centro de Ciencias del Mar y Limnología,
UNAM.
Se estudió la abundancia y distribución
de la población de Plexaura homomalla, en la zona
arrecifal de la parte norte del estado de Quintana Roo, México, frente
al Mar Caribe. El propósito fue determinar las posibilidades de explotar
racionalmente el recurso, sin dañar ni a la población explotada
ni a la comunidad arrecifal. Se encontró que la población es
relativamente abundante. Se proponen métodos de cosecha conservadores,
mientras se obtiene la información básica para entender la
dinámica poblacional de la especie. Se analiza también la
posibilidad de usar transplantes para incrementar el volumen
explotable.
The abundance and distribution of the population
of Plexaura homomalla was studied in the coral reef zone of
the Caribbean Sea, to the north of the state of Quintana Roo, Mexico. The
object of the study was to determine the possibilities to rationally exploit
the coral resource, without damaging the resource of the coral reef community.
The population was found to be relatively abundant. Methods were proposed to
harvest conservatively whilst basic information is obtained to better
understand the population dynamics of the coral species. The possibility of
using transplants to increase the exploitable volume has also been
analysed.
La importancia económica de las prostaglandinas en el campo farmacéutico ya ha sido establecido por numerosos autores, y es resumido por Pike (1974) . Es en este contexto que Plexaura homomalla es muy importante ya que es la única fuente natural de estas substancias, que tiene valor comercial, debido a la alta concentración encontrada en individuos de esta especie. Es aún más importante, debido a que P. homomalla solamente existe en la región zoogeográfica del Caribe (West Indies), no encontrándose en ningún otro mar tropical del mundo. Dada la importancia de dicho recurso, la protección y explotación racional del mismo, se consideran sumamente importantes. Siendo el objetivo de este trabajo mostrar la forma en que se realizó un inventario poblacional piloto de esta especie, y plantear algunas consideraciones sobre su explotación (Jordán, 1977b). ÁREA DE ESTUDIOEl área de estudio se encuentra localizada en la zona noreste de la península de Yucatán, entre Punta Nizuc y un lugar situado a 12 km al sur de Tulum, Quintana Roo, México (Fig. 1) , realizándose el trabajo en la zona arrecifal de la costa, cubriendo una distancia aproximada de 120 km lineales, entre septiembre de 1975 y septiembre de 1976. Esta área fue seleccionada como el sitio inicial de un proyecto general que pretende cubrir todo el litoral mexicano en el Mar Caribe. 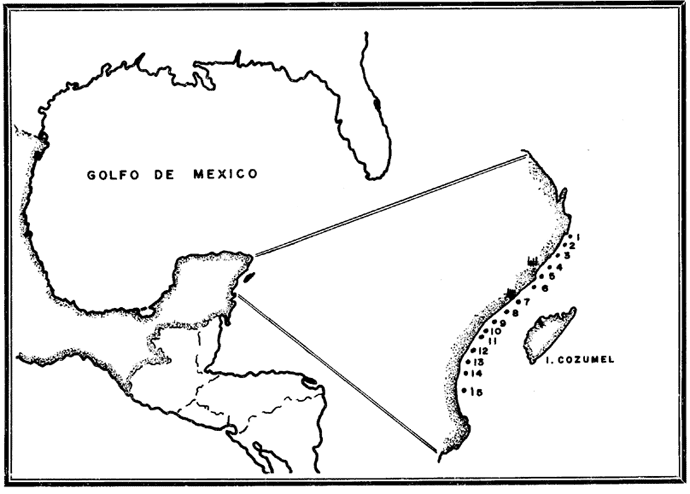 Fig. 1. Area de estudio. Los números corresponden a las unidades estudiadas: 1. Nizuc. 2. Bajo Finduvet. 3. Punta Petempich. 4. Puerto Morelos. 5. Punta Brava. 6. Punta Maroma. 7. Playa del Carmen. 8. Xcaret. 9. Pamul. 10. Yakul. 11. Akumal. 12. Cemuyil. 13. Xelha. 14. Tulum. 15. Tulum-12. MATERIAL Y MÉTODOSEl estudio está basado en unidades de muestreo intensivo formados por transectos de 10 m de ancho exactamente, que corren desde la rompiente arrecifal hasta la parte anterior de la laguna o canal arrecifal, cubriendo lo que corresponde al arrecife posterior, estas líneas tienen una longitud variable que va desde menos de 50 m hasta más de 160 m. El número de transectos por unidad es de 10, aunque pueden ser menos dependiendo de las condiciones locales, estando separadas entre sí por 90 - 100 m, tratando de cubrir 1 kilómetro de arrecife lineal por cada unidad básica. Dentro de cada transecto se realiza un minucioso conteo y medición de las colonias. Esta medición consiste en tomar la altura a partir de la ramificación primaria y después determinar los diámetros mayor y menor de la colonia vista desde su parte superior. En esta medición se incluyen todas las colonias presentes desde unos cuantos centímetros hasta más de 1 m de altura. Entre cada unidad básica queda un espacio de 8 a 12 kms dependiendo de las condiciones locales. Este espacio es estudiado cuidadosamente pero sin incluir el muestreo cuantitativo, a menos que la diferencia entre los estimadores de dispersión de las unidades limitantes sea importante o en el caso de que la abundancia y distribución subjetiva también sean diferentes; en tales casos se incluyen una o más unidades extra, de hecho se estudiaron 14 zonas, en vez de las once planteadas originalmente. Cuando la abundancia subjetiva de la población es muy pequeña o el área poblada muy restringida, en vez de utilizar las líneas paralelas de 10 m de ancho, se utilizó una sola línea y se midieron colonias en una franja de 6 m de ancho, 3 m a cada lado de la línea. En estos casos la colocación de las líneas fue hecha de tal forma que se tiende a estimar la abundancia máxima de la población. Para poder conocer la biomasa teórica de Plexaura homomalla, fue necesario desarrollar estimadores de volumen que se pudieran utilizar a partir de las medidas básicas de las colonias, terminando por utilizar índices de cobertura similares a los empleados por los botánicos (Müeller-Dombois, 1974). Los índices son de dos tipos: uno que se aplica a colonias en las que la relación del diámetro mayor (D) y el diámetro menor (d) es menor que 2, (D+d/4)² II · h · 10³ índice de cobertura denominado C, para colonias de forma redondeada típicas de zonas protegidas. Mientras que las colonias de zonas más expuestas, tienden a una forma más bien aplanada o de candelabro cuya relación D/d es igual o mayor que 2 representados por D·d·h/2 X 10-³, índice de cobertura tipo R. Asignando, mediante el índice de cobertura, un peso dado a cada tamaño y considerando el número de colonias de un tamaño dado en cada zona, puede deducirse la biomasa global para cada unidad de estudio. Con objeto de establecer el grado de relación del volumen teórico dado por los índices de cobertura (tanto R como C), y la biomasa real de las colonias medidas, se colectaron especialmente 237 colonias previamente medidas, de las cuales se obtuvo el peso húmedo y el peso seco en gramos. El peso húmedo de la colonia se obtuvo pesando directamente después de la colecta, mientras que el peso seco se obtuvo después de dejar las colonias colgadas al sol por más de 7 días, a estas mismas colonias se quitaba la porción basal y tronco por debajo de la primera ramificación, con objeto de no alterar el rango de índices. Los datos de biomasa teórica estimados a partir de los índices de cobertura, consideran un rango de variación natural y son estimados con un intervalo de confianza del 95%. Simultáneamente a los conteos, se determinó la abundancia subjetiva, la cual considera el grado de dificultad que tiene el buzo colector para encontrar y medir colonias de esta especie, así como la concentración colonial relativa, datos que los autores consideran de mucha importancia, ya que la protección racional del recurso depende en alto grado de estos dos factores. También se realizaron estudios sobre la viabilidad de transplantes en pequeña escala, para lo cual se cortaba una pequeña rama terminal de la colonia "madre" de 10 a 15 cm de longitud, la cual era colocada entre dos pedazos de madera embebidos en botes llenos de cemento en forma tal que ejercieran presión entre sí, fijando al transplante. Una vez que el transplante quedaba bien asegurado, el bote se enterraba en el sedimento, quedando sólo la parte superior con los maderos en el agua, asegurando así un substrato firme (Schroeder, 1974). Se transplantaron 4 lotes de 15 a 50 transplantes cada uno, habiéndose utilizado el mayor, para determinación de concentración de prostaglandinas en transplantes. Para la realización de este trabajo se utilizó principalmente buceo libre, y cuando las condiciones locales o el tipo de trabajo así lo demandaron, se utilizó equipo SCUBA. RESULTADOS Y DISCUSIÓN
CONSIDERACIONES GENERALESUno de los hechos más sobresalientes en el comportamiento de la población de Plexaura homomalla en la zona de estudio es la acentuada distribución de parche que presenta con cambios muy fuertes de densidad local, y las variaciones de acuerdo al tipo, de medio ambiente, en la curva de distribución de alturas. La zona óptima de desarrollo para esta especie está localizada entre la meseta arrecifal y la laguna arrecifal, de acuerdo a la zonación de Goreau (1959), en lo que sería el arrecife posterior. Dentro del marco de distribución se encontró que las abundancias "fuertes" están estrechamente relacionadas con. un desarrollo considerable de la zona de Acropora palmata, situada después de la rompiente y con la presencia de macizos de Montastrea annularis y un fondo duro aún cuando esté superficialmente cubierto por Thalassia y/o arena; es importante sin embargo mencionar que ésto último se refiere principalmente a colonias, mayores. La disponibilidad de un substrato duro para la implantación de larvas (Kinzie, 1974; Theodor, 1967) por un lado, y el probable efecto de corrientes y oleajes extremados (Barham y Davies, 1968; Rield, 1964) producen un efecto múltiple en la posición relativa de las colonias de todos tamaños que quedan muy heterogéneamente distribuidas. Este hecho hace que para el objetivo de este trabajo la expresión de la densidad poblacional por área determinada carezca de un significado real, sobre todo si se utilizan las áreas pequeñas comunmente utilizadas para inventario poblacional bentónico. En este caso, se consideró más apropiado expresar la densidad poblacional por secciones arrecifales de área variable (Jordan, 1977a) . Incluyendo en este concepto las variaciones locales normales que se encuentran en un arrecife y que no son por naturaleza forzosamente parecidas a ninguna otra sección arrecifal. Aún cuando en un principio se determinó establecer diferencias de densidad poblacional por zona en el arrecife posterior, el estudio mostró que esta diferenciación es poco significativa ya que las mismas zonas tienen muy poca definición, debido a una fuerte integración específica en los supuestos límites de estas zonas, por lo cual se consideró más apropiado expresar los valores por región arrefical posterior global. Debido a que las diferencias estructurales del arrecife son muy fuertes a lo largo de la zona de estudio, la población de Plexaura homomalla no presenta un gradiente hasta ahora reconocible de densidad y distribución, sino que existe en algunas zonas y en otras no. Dentro de las unidades en que la especie existe, la abundancia y concentración local está directamente relacionada con la disponibilidad de substrato y la protección a marejadas y oleajes muy fuertes. El área de las secciones arrecifales estudiadas va desde un mínimo excepcional de 70 m² hasta 10 000 m² dependiendo de la anchura del arrecife posterior, de la existencia o no de cabezas de coral dentro del piso lagunar, y de la concentración colonial relativa. La información obtenida a partir de los conteos fue utilizada en forma global por unidad, obteniéndose en primer lugar curvas de distribución de altura, asignando de acuerdo a los picos dominantes una posición en la población general, mediante el área total que cubre la sección arrecifal específica; posteriormente se establecía la proporción de colonias de una altura dada (entre 5 cm y 120 cm) por unidad. Independientemente de las variaciones locales en la distribución, se encontró que los picos de mayor abundancia de la población son rela tivamente constantes entre las diferentes unidades y que sólo son cambiados o desplazados en condiciones ambientales con mayor stress para la especie. Estos picos corresponden a las alturas de 50, 25, 30 y 75 centímetros, en orden de importancia numérica. Considerando las estimaciones de Opresko (1974) en relación con nuestros datos de biomasa la edad de estas colonias puede ser de aproximadamente 8 años para h = 25 cm, hasta más de 30 años para h = 75 cm. De ser ésto así, la figura 2 representa la curva de distribución de la población en un período mayor de 40 años. 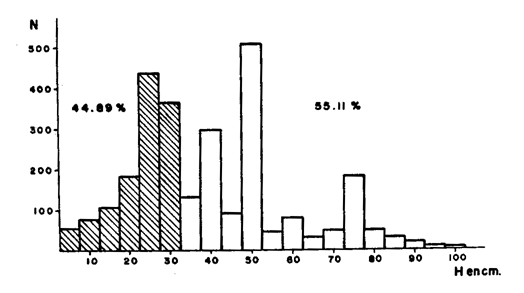 Fig. 2. Curva general de distribución de altura de la especie, en la zona de estudio. En zonas de muy alta energía (marca y oleaje) la densidad poblacional puede mantenerse muy alta (hasta 235 colonias, por un área de 38 m²), pero el pico de 75 cm desaparece por completo y el de 50 se ve muy reducido. Además, es importante considerar que en estos ambientes las colonias son de forma aplanada predominantemente, debido precisamente al alto nivel de energía (Rees, 1972) , considerado por nosotros como índice de cobertura R en los cálculos de biomasa, lo que las coloca automáticamente en la parte menos precisa de la curva de relación de biomasa, llegando a ser éste el único caso en que se encuentre una alta densidad poblacional con una relativamente baja biomasa de las poblaciones. Sin embargo, en forma global la proporción numérica de colonias chicas con relación a grandes, es alta, del orden del 45% de las primeras contra 55% de las segundas, lo que plantea la posibilidad de que a nivel poblacional exista una fuerte tasa de reclutamiento. Un hecho interesante que se observó es que la proporción de colonias chicas de esta especie es mucho mayor que la de las grandes en la zona posterior a la rompiente, o sea en la zona de alta energía y lo contrario sucede en zonas menos agitadas. Se puede descubrir en ésto un efecto selectivo, producido por la ola sobre colonias mayores que presentan mayor resistencia y por lo tanto más fáciles de derribar en la zona de alta energía, mientras que el menor tamaño de las colonias menores en la parte posterior podría deberse más bien a interrelaciones de tipo biológico, como competencia por substrato y cubrimiento por algas. Con objeto de establecer la equivalencia entre el índice de cobertura y la biomasa real, se extrajeron un total de 237 colonias, las que fueron previamente medidas en el agua y posteriormente pesadas (obteniéndose peso húmedo y peso seco) . De algunas de estas mismas colonias se estableció la relación biomasa-esqueleto para las diferentes formas del organismo. En este caso, la biomasa como peso seco incluye las espículas y en el anterior el esqueleto, habiéndose eliminado el disco basal y el tronco primario por debajo de la primera ramificación, que son básicamente esqueleto axial. Un punto importante a considerar es que el uso de los índices de cobertura supone, para dar una máxima definición, que la densidad de las colonias de un volumen dado es igual o similar entre sí, sin embargo, esto no, sucede y la variación en el número y grado de ramificación es alto, variando fuertemente la densidad media de las colonias, lo que produce un decremento en la eficiencia del índice de cobertura. Los resultados muestran que la relación entre el índice de cobertura (o sea la biomasa teórica a partir del volumen estimado) y la biomasa real es moderada, con una dispersión no muy alta, al 5% de error, con valores entre el 68 y el 76%, (Fig. 3), mientras que la relación entre la altura de la colonia y la biomasa real es más pobre y más dispersa, entre 52% y 70%, (Fig. 4) ; sin embargo, es definitivamente superior a estimaciones basadas en la sola densidad poblacional, sobre todo considerando que las variaciones en el tamaño y forma de las colonias es muy alta y que la distribución de parche de éstas es muy acentuada. 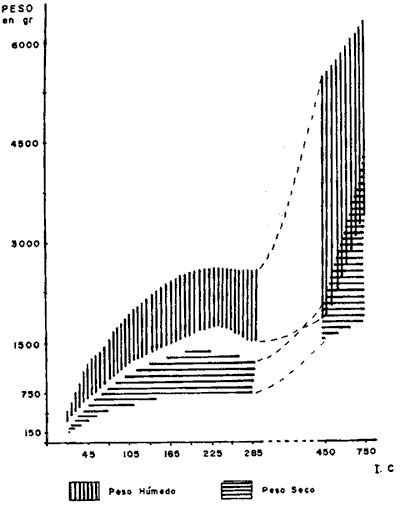 Fig. 3. Curva general de distribución de biomasa en la zona de estudio. 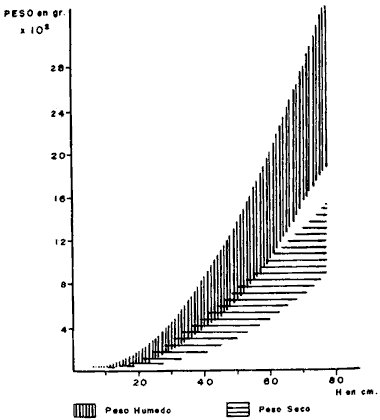 Fig. 4. Relación entre altura y peso seco. Un hecho interesante es que hay una relación directa entre el índice de cobertura y la altura, a pesar de las fuertes variaciones en la densidad colonial. EVALUACIÓN DEL RECURSO Y VOLUMEN DE COSECHAEn la evaluación del recurso y estimación del posible volumen de explotación, se han utilizado tanto las estimaciones poblacionales como las de biomasa; sin embargo, es elemental considerar la forma de cosecha para llegar a una estimación global adecuada. Puede decirse que en términos generales la ocurrencia de esta especie es común y aún dominante en algunas áreas de la zona estudiada; sin embargo, la variabilidad poblacional local es muy alta lo que impide encontrar o definir gradientes. Respecto a la abundancia del recurso es más difícil valorarla actualmente, ya que es un término relativo a la cantidad de materia prima necesaria por unidad de tiempo, que se determina en alto grado por el nivel de eficiencia de la industria proyectada. Para datos concretos sobre los valores y volúmenes determinados ver Jordán, 1977b. METODOLOGÍA DE COSECHAPara determinar qué forma de cosecha es la más adecuada, se partió del tipo de explotación sugerido a la Upjohn Co. por Kinzie (1974), el cual establece que deben podarse colonias mayores de 30 cm y del 50% al 75% de dicha colonia, siempre y cuando no se expusiera el eje axial, cuando su diámetro fuera mayor de 1.5 cm de diámetro. Sin embargo, esta proposición no fue basada en estudios específicos sobre Plexaura homomalla, sino sobre octocorales en general (Kinzie, 1973) . Se observó en este estudio que, de acuerdo a la curva de distribución de la población general (Fig. 2), un 55% de las colonias de esta especie son mayores de 30 cm, y por lo tanto representarían el stock cosechable, mientras que un 45% de la población en número de colonias no sería cosechado hasta que alcanzaran alturas mayores de 30 cm. Quedando así un amplio margen de recuperación para la población. Sin embargo, estos datos corresponden a una estimación basada en la sola densidad poblacional colonial. Se ha demostrado en este estudio que existe una variabilidad considerable en estas poblaciones, en base a lo cual sería erróneo utilizar sólo densidades coloniales; por lo tanto se decidió establecer la relación de explotación con valores de biomasa. Encontrándose que 94% de la biomasa total de la población estimada corresponde a ese 55% explotable, de acuerdo a los criterios de densidad poblacional colonial, pero al 45% no explotable corresponde solamente un 6% de esta biomasa total (Fig. 5) . 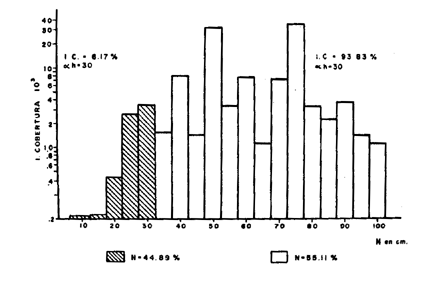 Fig. 5. Relación entre Índice de cobertura y peso seco. Así, si se sigue un criterio de densidad poblacional, en sólo la primera cosecha se reduciría tan drásticamente la población individual que sería muy difícil que la población recuperara su estado anterior. Esto se debe al hecho de que las colonias son conjuntos de individuos, no individuos en sí. La disposición de estos individuos o pólipos en la colonia es de una sola capa de espesor, siendo igual esta condición desde la base hasta las puntas. En la fotografía se observa que la mayor superficie se encuentra en las zonas terminales debido a la amplia ramificación, por lo tanto el número de pólipos es mucho mayor precisamente en las zonas de la colonia sujetas a explotación. Ahora bien, Kinzie (1974) aconseja podar del 50% al 75% de la colonia: de hacerse así se estaría reduciendo un porcentaje similar o mayor del total de pólipos, afectando de esta manera el potencial reproductor sexual de esta especie, ya que existe una relación directa entre el número de pólipos y el número de larvas que al fijarse a un substrato adecuado darán origen (si sobreviven) a una colonia nueva. En base a estos resultados, lo conveniente será podar un porcentaje menor (25%), de cada colonia, y mejor aún podar principalmente colonias de gran volumen y densidad que se encuentren sobre los picos de 50 y 75 cm o más de altura, lo cual aparte de dar un mayor margen a la población explotada, tiene un efecto positivo como el mismo Kinzie señala, ya que, debido a la reducción del volumen de la colonia, la posibilidad de que la colonia sea derribada y muera por abrasión con la arena, debido al efecto mecánico de olas de tormenta o marejada fuerte, es mucho menor. De hecho, ésta es considerada por numerosos autores como la mayor causa de mortalidad (Cary 1918, Kinzie 1973) de gorgonáceos. Si la abundancia y distribución de la especie fuera la adecuada, sería conveniente diseñar una metodología de cosecha de tipo revolvente. Esto es, empezar en un sitio de la costa y seguir el litoral hasta el final, a un ritmo tal que no se volviera a tener que explotar el primer sitio hasta haber transcurrido de 7 a 10 años en base a la información actual. Claro que ésto depende de la magnitud del volumen de materia prima necesario para la industria proyectada, pero de ser posible, debe utilizarse este método ya que es el más favorable ecológicamente. CONSIDERACIONES ECOLÓGICASDurante el desarrollo del presente estudio se empezaron a plantear los problemas a resolver y sus metodologías correspondientes, aplicando un énfasis especial en las causas de la mortandad natural de la especie y su relación con la explotación racional del recurso. Entre los factores que pueden afectar la población, aparte del antes mencionado, está el efecto de predadores (Bayer, 1961), en particular Cyphoma gibbosum, no tanto por la propia predación directa, sido debido a que al comer el coenénquima (Theodor, 1967; Kinzie, 1974; Silveira, 1976) expone el eje de la colonia (Fotogr. l), lugar adecuado para la implantación de epibiontes, ya sean algas filamentosas (Fotogr. 2) (en cuyo caso se impide la regeneración) o hidrocorales como Millepora alcicornis, en cuyo caso, tarde o temprano, la colonia morirá irrevocablemente (Fotogr. 3) . Sin embargo Cyphoma no parece ser un predador específico de Plexaura homomalla, lo que disminuye la presión de predación.  Foto 1. Ovúlidos del género Cyphoma predando sobre una colonia (le Plexaura homomalla observándose la denudación de las ramas por los predadores.  Foto 2. Algas filamentosa que impide la regeración del tejido de Plexaura homomalla. Un factor muy importante es que aún cuando los hechos antes mencionados no son abundantes, el efecto de la poda bien puede incrementarlos, de ser así quizás el más peligroso sea la implantación de algas filamentosas, sobre todo si las condiciones ambientales no son, en un momento dado, muy adecuadas para Plexaura homomalla. Los autores han observado que en zonas con alto contenido de sedimentos en suspensión y fuerte movimiento de agua, la incidencia de colonias afectadas por algas filamentosas es mucho mayor que en ambientes más tranquilos. DESARROLLO EXPERIMENTAL DE LOS TRANSPLANTESEn cuanto se refiere a los medios para incrementar la población artificialmentel los transplantes fueron realizados en 4 lotes y se escogieron las localidades arrecifales de Puerto Morelos y Punta Petempich para la experimentación. Aunque el manejo de esta especie es muy delicado, debido a que aparentemente es muy sensible a la manipulación, los resultados fueron altamente exitosos, un 92% de los transplantes sobrevivió. En la fotogr. 4 se puede observar un transplante recién efectuado y en la fotogr. 5 se observa otro aproximadamente al mes y medio; se aprecia un engrosamiento del tejido en la parte basal, lo que significa que la colonia está formando su propio disco basal, con lo cual se considera que el transplante ha tenido éxito.  Foto 3. Millepora alcicornis extendiendose sobre el esqueleto axial de Plexaura homomalla.  Foto 4. Transplante a la semana de haber sido fijado  Foto 5. Transplante al mes y medio. Se observa el inicio de la formación del disco basal. Todos los lotes fueron plantados cerca de las colonias madres con objeto de disminuir el efecto de la manipulación. Aun cuando los resultados son muy alentadores, la tasa de crecimiento de estos transplantes es considerablemente baja, siendo la tasa de regeneración de estos transplantes muy baja también; de no darse el tiempo necesario para la formación de colonias relativamente grandes (mayores de 30 cm) según Opresko (1974) una colonia de 25 cm tardaría aproximadamente ocho años en alcanzar esa talla. Por otro lado, no se sabe qué proporción de transplantes sobreviviría en escalas masivas, cuando la manipulación fuera mucho más fuerte, debido a la necesidad de transplantes a distancias considerablemente mayores de la colonia madre. Es también importante tomar en cuenta que se desconoce el efecto que pueda tener un incremento considerable de esta especie en la comunidad arrecifal y en sí misma. Es probab le encontrar problemas serios para los transplantes si se traspasa una masa crítica máxima de la población, con respecto a la comunidad de la que forma parte, ya que se pueden disparar mecanismos de autorregulación comunitaria que de una forma u otra acaben con esta "nueva población". Existen también razones para preocuparse del efecto que la población artificialmente incrementada tenga en la comunidad. Es además necesario considerar los argumentos de orden económico, directamente dependientes de la demanda de las prostaglandinas como se plantea en los antecedentes. Un punto de considerable importancia es el averiguar si la concentración de prostaglandinas en los transplantes es similar a la encontrada en la colonia madre, y sobre todo si dicha concentración se mantiene tanto en los transplantes como en las colonias madre después de tres o cuatro cortes en la misma colonia, como sería el caso de una explotación a largo plazo. Conclusiones1) Se encontró que la zona óptima de desarrollo para Plexaura homomalla es somera, agitada pero protegida de oleaje y marejada muy fuerte, características de la zona posterior arrecifal, cuando existe una barrera bien formada. 2) Que la formación de grandes macizos de Acropora palmata y Montastrea annularis después de la zona de rompiente, pueden indicar zonas en que Plexaura homomalla se encuentre en condiciones tales que la densidad poblacional sea alta. 3) Que la forma de la colonia sufre un cambio conforme el grado de energía en el movimiento del agua aumenta. Este cambio va de forma de arbusto a forma de candelabro. 4) La utilización de índices de cobertura, mostró ser un elemento muy útil en la evaluación poblacional (de la biomasa), obteniendo una precisión considerable en relación a la magnitud del área por evaluar. 5) Puede considerarse que en la zona de estudio, las poblaciones de Plexaura homomalla, se encuentran en un estado de desarrollo normal, activo, no observándose alteraciones en la comunidad arrecifal. 6) A pesar de tener una población abundante, es necesario considerar seriamente la utilización de transplantes y medios de semicultivo, con objeto de aumentar el stock existente. La importancia de este proceso es mayor conforme la demanda industrial se incremente. 7) Que el máximo cosechable por colonia, no debe exceder al 25% de la biomasa de la colonia mayor de 30 cm, mientras no exista información suficiente para establecer la dinámica poblacional de la especie. 8) Aun cuando predadores y epífitas no parecen ejercer un efecto regulador sobre la abundancia de la población, si la poda de las colonias no se hace cuidadosamente, puede incrementarse el número y efecto de epibiontes implantados sobre el eje, con las consecuencias planteadas en el texto. AgradecimientosLos autores agradecen a las siguientes personas su ayuda y colaboración en los diferentes aspectos de este trabajo. A F. Bayer por su desinteresada ayuda en la iniciación de este trabajo. A R. Kinzie por sus consejos y comentarios. A A. Laguarda-Figueras por su constante apoyo y a A. Ayala-Castañares sin cuya ayuda este trabajo no se habría podido realizar. A P. Moreno por su invaluable ayuda a todo lo largo del desarrollo de este estudio. A los señores L. Castañares y R. Ibarra en el trabajo de campo. LITERATURABARHAM, E. y J. DAVIES, Underwat. Nat., Gorgoníans and water motion studies in the Gulf of California. 1968 24-28 5 3 BAYER, F. M., The shallow water octocorallia of the West Indian region. Martinus Nijhoff The Hague Holanda 1961 373 CARY, L. R., Carnegie Inst. Wash. Publ., Observations upon the growth rate and Oecology of Gorgonians. 1914 79-90 182 Publs. Carnegie Inst. The gorgonacea as a factor in the formation of coral reefs. 1918 341-362 213 GOLDBERG, W. M., Bull. Mar. Sci., The ecology of the coral-octocoral communities of the southeast Florida coast. Geomorphology, species composition and zonation. 1973 465-488 23 3 GOREAU, T. F., Ecology, The ecology of Jamaican coral reefs: I-Species composition and zonation. 1959 67-90 40 1 GRIGG, R. W., Ecology and population of the gorgonians Muricea california and Muricea fruticosa. Ph. D. Dissertation, Univ. of Cálifornia, 1970 260 JORDAN, D. E., . Variaciones en el desarrollo estructural del sistema arrecifal del NW del Caribe Mexicano. IV Congreso Internacional de Ecología Tropical. Panamá. 1977 a Contrato de Estudios Ecología y evaluación de las poblaciones de Plexaura homomalla y su explotación racional con el fin de preservar el recurso. Informe Final Convenio de Colaboración entre Consejo Nacional de Ciencia y Tecnología (CONACYT) y Centro de Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 1977b 56 KINZIE, R. A., Bull. Mar. Sci., The zonation of West Indian gorgonians. 1973 93-155 23 1 Stu. Trop, Oceanogr. 12 Plexaura homomalla; The biology and ecology of a harvestable marine resource. In: (Bayer, F. M. Ed.). Prostaglandins from Plexaura homomalla: Ecology, Utilization and Conservation of a Major Medical. Marine Resource A Symposium. University of Miami Press. 1974 MULLER-DOMBOIS, D. y H. ELLENBERG, Aims and Methods of Vegetation Ecology. John Wiley & Sons P. 1974 OPRESKO, D. M., Bull. Mar. Sci., Abundance and distribution of shallow water gorgonians in the area of Miami, Fla., 1973 535-558 23 3 Stu. Trop. Oceanogr. 12, Recolonization and regrowth of a population of the gorgonian Plexaura homomalla. In: (Bayer, F. M. Ed.) Prostaglandins from Plexaura homomalla: Ecology, Utilization and Conservation of a Major Medical Marine Rosource A Symposium. University of Miami Press. 1974 PIKE, J. E., Stu. Trop. Oceanogr. 12, The prostaglandins: their history, development and potential. In: (Bayer, F. M. Ed.) Prostaglandins from Plexaura homomalla: Ecology, Utilization and Conservation of a Major Medical Marina Resource A Symposium. University of Miarni Press. 1974 REES, J. T., Jour. exp. Mar. biol. ecol., The effect of current on growth form in an octocoral. 1972 115-123 10 RIELD, R., Helgolander wiss Meresunters, Die Erscheinungen der Wasserbenegung und ihre wirkung auf sedentarien im Mediterranean felslitoral. 1964 155-186 10 SCHROEDER, R. E., Stu. Trop. Oceanogr. 12, Observations on the vegetative culture of Plexaura homomalla in the Cayman Islands. In: (Bayer, F. M. Ed.) Prostaglandins from Plexaura homomalla: Ecology, Utilization and Conservation of a Major Medical Marine Resource A Symposium. University of Miami Press. 1974 SILVEIRA, F. L. y T. VAN'T HOF , . Contr. Zoology (Amsterdam) submitted in 1976 (en prensa). Regeneration in the gorgonian Plexaura flexuosa. 1976 THEODOR, J., Vie et Milieu, Contribution a l'etude des gorgones (7): Ecologie et comportement de la planula. 2a 18 291-301 18
|