|
PROSPECCIÓN BIOLÓGICA DE LA LAGUNA DE
NICHUPTÉ, CANCÚN, Q. R.., MÉXICO: NOTA
CIENTÍFICA
Trabajo recibido el 22 de abril de 1977 y aceptado para
su publicación el 13 de junio de 1977.
ERIC JORDAN
ROBERTO DE LA TORRE
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnologia.
MICHEL ANGOT
Experto de UNESCO en Biología Marina.
Contribución 75 del Centro de Ciencias del Mar y Limnología,
UNAM.
De noviembre 1971 a febrero 1972, se realizó un
estudio de prospección biológica en la laguna de Nichupté,
con el propósito de definir las principales características
hidrológicas del sistema. Se encontró que el intercambio de las
aguas lagunares con el Mar Caribe es bajo y afecta principalmente la parte
norte de la cuenca. La circulación interior es también baja y se
debe principalmente al viento. El efecto de las lluvias y de aguas
subterráneas se deja sentir fuertemente en los gradientes de salinidad y
temperatura. Se realizó un inventario general del bentos y necton
incluyendo fanerógamas marinas, algas, moluscos y peces,
encontrándose que la diversidad de la comunidad es relativamente alta, a
pesar del carácter preliminar de los muestreos.
A biological survey was realized from november 1971 to
february 1972, in the lagoon system at Nichupté (Cancún).
Hidrological data suggest that water exchange between the system and the
Caribbean sea is poor. The water circulation inside the system is also poor and
affects mostly the northern part of the lagoon system. The effect of fresh
water coming from the substrate through "cenotes" and rain is marked during the
season studied. Biologically the lagoon is dominated by Thalassia testudinum.
Marine fanerogams, algae, moluscs and fishes were collected. For the samples
taken H' is relatively high, even if sampling is regard just as
preliminary.
El objeto de la presente nota es facilitar información sobre los resultados de un estudio de reconocimiento efectuado en el Sistema Lagunar de Nichupté (Cancún), Q. Roo, durante 3 meses a finales de 1971 y principios de 1972. Uno de los objetivos principales del estudio era programar un proyecto de investigación y control ambiental marino a largo plazo, encargado de monitorear el impacto humano sobre esta zona de futuro desarrollo turístico. Dicho programa nunca se llevó a cabo, aunque el desarrollo urbano y turístico sí, de ma nera que la información contenida en esta nota, aun cuando poco elaborada, puede ser de utilidad en un futuro cercano, para tratar de entender y luego remediar los efectos del desarrollo de la zona turística sobre el ecosistema lagunar. Este estudio consistió de dos partes: una encargada de determinar las generalidades del comportamiento hidrológico del sistema y otra, iniciar el inventario poblacional, con bentos y necton, además de un análisis de variabilidad natural de las poblaciones de Thalassia testudinum en diferentes localidades del sistema, información que puede encontrarse en Jordán et al. (1972) y que no se menciona en esta nota. MATERIAL Y MÉTODOSDurante el estudio se realizaron 47 estaciones hidrológicas, donde se estimó batimetría, temperatura, salinidad y concentración de oxígeno disuelto en la columna de agua. Para el estudio de intercambio de agua se instalaron tres estaciones de registro continuo (cada 30 minutos) para mareas, velocidad y dirección de corrientes y velocidad y dirección de viento. Para el estudio de circulación se utilizaron cruces de deriva en la laguna y fluoresceína sódica en los canales. Ambos métodos fueron monitoreados tanto desde el aire con avión, como desde la superficie del agua con lancha. A la vez que se realizó el estudio sobre las condiciones hidrológicas de la laguna, se llevó a cabo un inventario poblacional y estudio de la variabilidad de biomasa en cuatro áreas lagunares diferentes y una marina (Jordán, et al. 1972) ; utilizando para ello dos métodos de muestreo, uno cualitativo que correspondió a la recolección de material en viajes de reconocimiento a lo largo de toda la laguna, y otro cuantitativo mediante el método de Petersen (Thorson, 1957), con muestreo al azar, cubriendo en cada estación aproximadamente 5 m² de fondo. También se hicieron colectas de peces en diferentes partes de la laguna como se explica más adelante. Para la recolección de muestras bentónicas se utilizó SCUBA, y para el necton diferentes artes de pesca (chinchorro, atarraya y anzuelo) logrando un total de 21 capturas. La identificación del material se dejó a cargo de personal especializado del entonces Departamento de Ciencias del Mar del Instituto de Biología, UNAM, la flora fue identificada por la Dra. Martha Ortega, los moluscos por el Dr. Antonio García-Cubas y la ictiofauna por el M. en C. Andrés Reséndez, la diversidad (H') fue estimada por medio de la fórmula de Shanon y Weaver y expresada en bits (Pielou, 1975). En las muestras se recogieron también otros organismos que no son considerados aquí por su bajo número o dificultad de identificación específica, entre ellos hubo poliquetos (sabéllidos, serpúlidos) isópodos, decápodos, amfípodos, equinodermos (echinoideos y holoturoideos), así co mo celenterados (hydroidea). RESULTADOS Y DISCUSIÓN
FISIOGRAFíA Y CLIMALa laguna de Nichupté (Cancún) está situada en la parte NE de la península de Yucatán, en el territorio de Quintana Roo, a los 86° 44' de longitud y 21° 31' de latitud aproximadamente. Se encuentra en una región tropical con clima cálido, subhúmedo con lluvias aisladas, clasifi cación AW (Koeppen). El substrato en toda la región es virtualmente carbonato de calcio, mientras que el sedimento dentro de la laguna es lodo aragonítico, probablemente resultado de la desintegración de foraminíferos, codiáceas y algas rojas (Brady, 1972). El sistema está formado por la laguna Nichupté propiamente, una laguna más o menos aislada conocida como Laguna de Bojórquez y dos lagunas pequeñas, la de Somosaya y Río Inglés que se caracterizan por tener numerosos cenotes sumergidos, los cuales aportan cantidades considerables de agua dulce al sistema (Fig. 1) . La profundidad promedio de la cuenca es de 1.5-2 metros y muy rara vez excede de los 3.5 metros, la pendiente de las orillas al fondo es muy suave excepto en los canales. Dentro de la cuenca lagunar se encuentran dos bajos (Bajo Norte al norte y Bajo Zeta al sur) que dividen prácticamente la cuenca en tres partes y que determinan en alto grado el movimiento de la masa de agua lagunar. Dichos bajos tienen una profundidad media de 30 a 40 cm y en algunas zonas llegan a aflorar durante la marca baja, su anchura en ocasiones es de más de 800 m. Los lugares más profundos del sistema están localizados en los canales de comunicación con el mar, el Canal Cancún, al norte de la laguna, frente a Isla Mujeres, el canal Nizuc al sur de ésta y el canal de la Zeta que atraviesa el bajo del mismo nombre, siendo todos ellos de fondo irregular con un promedio de 2.0 a 2.5 m, llegando en ocasiones hasta 5 m de profundidad. El ancho de estos canales también variable, va de 17 a 7 5 m. 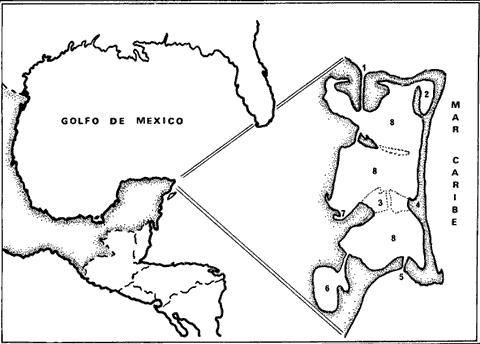 Fig. 1. 1. Canal Cancún. 2. Laguna Bojórquez. 3. Bajo Zeta. 4. Isla Cancún. 5. Canal Nizuc. 6. Río Inglés. 7. Somosaya. 8. Laguna de Nichupté. HIDROLOGÍADurante el estudio hidrológico pudo observarse que las masas de agua lagunar se portan de manera diferente en las tres zonas de la laguna Nichupté, debido probablemente al efecto de los bajos, por una parte y al aporte de agua del subsuelo a través de cenotes situados en el margen occidental en las zonas central y sur de la laguna, por otra. En la época de estudio la precipitación pluvial provocada por "nortes" fue alta, esta agua escurre de la zona de manglares hacia la laguna, también por el lado occidental, acentuando los gradientes de salinidad. Es notoria la cantidad de taninos provenientes de los manglares que es arrastrada a la laguna dando en algunas localidades como la laguneta de Somosaya, un color café rojizo obscuro al agua. Por estos aportes, y los antes mencionados de los cenotes, la laguna presentó características hiposalinas. Llegando la salinidad a 37.5 %o, sólo en la laguneta de Bojórquez, esto en una zona aislada del margen occidental, mientras que en el margen oriental la salinidad promedio fue de sólo 6 - 7°/oo. A pesar de los fuertes aportes de agua dulce no se apreció ninguna estratificación, mostrando que la acción del viento es suficiente para mezclar la masa de agua en sentido vertical, debido a lo somero del sistema. Es probable que solamente en esta época del año la laguna sea hiposalina, ya que la evaporación debe ser alta por acción combinada de vientos constantes e insolación. De hecho la temperatura de la laguna fue sólo 2° C, más alta que la temperatura de las aguas superficiales del Mar Caribe y el rango interno de 3° C (28.1 a 31.0° C) estableciéndose el gradiente en el mismo sentido general que el de salinidad. El contenido de oxígeno disuelto, puede considerarse relativamente bajo en general, ya que el 90% del fondo lagunar está cubierto, por Thalassia testudinum, y los valores medios son de alrededor de 3.0 a 3.5 ml/1, encontrándose el punto de saturación en la mayoría de los casos superior a 4.5 m1 / 1 aumentando conforme la salinidad disminuye. Esto puede deberse, en parte a una concentración de materia orgánica en la masa de agua, producto del arrastre, de la llanura de inundación. En su dinámica el sistema es principalmente afectado por el viento y secundariamente por las mareas. Los vientos dominantes son: S-SE, excepto cuando sopla el "norte", la velocidad promedio durante la época de estudio fue de 2-3 nudos, manteniéndose a veces en velocidades bajas (menos de 1 nudo) por periodos largos, lo que parece ser suficiente para mantener, en general, un movimiento continuo de la capa de agua superficial del este al oeste, lo que probablemente ayuda al proceso de mezcla, ya que la dirección dominante tanto del viento como del agua, es contraria a los gradientes de salinidad y temperatura. Este movimiento fue claramente observado por medio de cruces de deriva. Las mareas en esta zona, son de tipo semidiurno y de muy pequeña amplitud (rango máximo 25 cm en la época de estudio) , el efecto que la marea pudiera tener en el intercambio laguna mar, es reducido por la longitud y sinuosidad de los canales de entrada y el efecto de los bajos. Esto es más notorio en el canal Nizuc que presenta el comportamiento de un canal basculante, limitándose la masa de agua a recorrer el canal de ariba a abajo sin llegar a establecer un intercambio efectivo entre el mar y la laguna. Esto fue claramente observado por medio de marcado con fluoresceína sódica, siguiendo el curso de las manchas, tanto desde el aire como desde el agua. El intercambio entre mar y laguna se realiza básicamente a través del canal Cancún, y dicho movimiento afecta principalmente la parte norte de la cuenca, pero no el resto de la laguna debido a la presencia de los bajos. Se encontró por las observaciones con fluoresceína, que en general la masa de agua lagunar se mueve de la zona central oeste hacia el canal Cancún, registrando posteriormente que el canal de Cancún presenta un tiempo de reflujo (6-8 horas) mayor que el de flujo (3-5 horas) y también velocidades de corriente mayores en el reflujo (1-2 nudos) que en flujo (0.5-1 nudos) lo cual indica un movimiento de agua de sur a norte dentro de la zona central de la laguna. Calculando (en base a 44 horas) el volumen de intercambio entre la laguna y el mar a través de los canales de Nizuc, se encontró que en general el intercambio efectivo era de sólo un 7% del volumen lagunar total calculado en el orden de 68.9 × 106 m³, de este intercambio, el canal Nizuc sólo transporta un 18%, de lo cual se deduce que en la época de estudio, el canal Cancún arrojaba al mar un excedente respecto al canal Nizuc, excedente que es proveniente tanto del escurrimiento de la llanura de inundación, como de los cenotes sumergidos. Esto concuerda con los gradientes de salinidad, temperatura y oxígeno, así como con el movimiento y velocidad de los marcadores (Jordán et al., 1972). BIOTALa laguna se encuentra bordeada en su margen interno por densos manglares (Rhizophora mangle, Avicennia sp.) que unen prácticamente la tierra firme con la Isla Cancún. En el lado occidental estos manglares crecen sobre una llanura de inundación y son a medida que se adentran a tierra continental, sustituídos por una selva baja perenifolia. FanerógamasBiológicamente el sistema se caracteriza por tener el fondo cubierto por Thalassia testudinum, la laguna de Nichupté tiene el fondo formado por sedimentos carbonatados y cubierto por una capa vegetal cuyo principal componente (más de 90%) es Thalassia testudinum. Esta fanerógama marina cubre también el fondo de la laguneta de Bojórquez y parte de los canales tanto internos como los que establecen comunicación con el mar, sin embargo no se encuentra ni en la laguna de Somosaya, ni en la del Río Inglés, donde las salinidades (6-7 %o) son demasiado bajas para permitir su presencia (den Hartog, 1970), sin embargo aquí se encontró (Somosaya) una población de Ruppia maritima, bien representada. La importancia de Thalassia testudinum en diferentes aspectos de la producción tanto primaria como de detritos, en la estabilización de ambientes marinos, en el aporte de substrato para organismos epifíticos, en la protección de etapas larvales y juveniles de muchas especies, etcétera ya ha sido mencionado por muchos autores (den Hartog, 1970; Zieman, 1970; Phillips, 1960; Bernatowicz, 1952; Lot, 1968; Patriquin, 1973; Hornelas, 1975). La cobertura vegetal del fondo varía en densidad, presentando su máximo en los fondos de la cuenca, y su mínimo en los canales o zonas de corrientes, donde se puede encontrar arena desnuda, o una cubierta parcial de Thalassia en pequeños parches con Penicillus capitatus aunque den Hartog (1970) reporta que T. testudinum no es la única fanerógama en las camas de esta especie, dentro de la laguna no se encontró más que Syringodium filiforme en zonas expuestas (sobre los bajos) , donde domina en pequeños parches. Un dato interesante es que la proporción de la biomasa que ocurre bajo la superficie (rizomas y raíces) es estimada entre 55 y 90% del total por den Hartog (1970) y otros. Sin embargo en nuestra área de trabajo los rizomas y raíces comprendieron un 72% en la zona deThalassia marina, en una zona lagunar con Thalassia alternada con parches de arena 48% y en las zonas más densas 44%. Estos dos últimos valores son significativamente más bajos que lo reportado anteriormente. Es probable que el hecho de vivir en un ambiente tan protegido permita un incremento de la superficie fotosintética con respecto a raíces y rizomas, algo similar encuentran Burkholder y Rivero, en Puerto Rico en condiciones aparentemente similares (den Hartog, 1970) . Sin embargo a pesar de que la densidad de follaje es mucho mayor en la laguna que en el mar, la biomasa total es un 38% mayor para Thalassia testudinum en el mar que en la laguna. Algas y fanerógamasLas camas de Thalassia ofrecen un substrato muy importante para algas epífitas (Humm, 1964) y en la zona de estudio se encontraron abundantemente, representadas con 15 familias, 22 géneros y 29 especies (Tabla 1) .  TABLA 1 LISTA DE LAS ESPECIES DE ALGAS Y FANERÓAGAMAS OBSERVADAS  CONTINUACIÓN TABLA 1 De éstas, las epífitas más importantes fueron Melobesia farinosa, Laurencia implicata, L. poitei, Herposiphonia tenella y Ceramium sp, sin embargo el género Fosliella importante alga calcárea, no se encontró en ninguna de las muestras. De las 29 especies, 11 corresponden a cianofitas, las cuales no se encontraron en Thalassia sino en Ruppia maritima en la laguna de Somosaya, es importante señalar que sólo la rodofita Polysiphonia sp se encontró tanto en R. maritima, como en Thalassia testudinum. Otras algas reportadas como importantes para esta comunidad son: Acetabularia, Udotea, Dictyota, Rhipocephalus (Zieman, 1970), que aunque han sido observadas en áreas aledañas no se encuentran en la laguna, ni aparentemente en la zona de influencia de canales, excepto Penicillus capitatus, de hecho con excepción de esta especie y Batophora oerstedi, las macroalgas están muy pobremente representadas. MacrofaunaMoluscos: Los moluscos fueron colectados tanto en la superficie como dentro de sedimento hasta una profundidad de 10 cm. En este tipo de ambiente la acumulación de conchas de épocas anteriores es muy fuerte debido a que no hay fuerzas mecánicas que los cambien de lugar (Ekdale, 1972; Warmke y Abbot, 1961), por esta razón se extrajeron, tanto muertos como vivos, con un total de 24 familias, 34 géneros y 43 especies (Tabla 2) de los cuales 14 familias, 15 géneros y 15 especies corresponden a los moluscos vivos, los cuales constituyen solamente un 22% de la muestra total. Una situación ecológica interesante es el no encontrar a las especies vivas representadas en la laguna fuera de ésta, lo cual es particularmente interesante en Chionecancellata, reportada por Ekdale (1972) como la más abundante en la región. Es sin embargo importante mencionar que la zona de muestreo de Thalassia marina de los autores, no se encuentra en la zona de muestreo de Ekdale (Isla Contoy y sur de Isla Cancún) , que es al norte del Canal Cancún, incluso especies como Tricolia thalassicola y T. affinis características de pastos marinos (Warmke y Abbot, 1961) se encuentran dentro de la laguna, pero no fuera de ella. Brachidontes citrinus, se encontró exclusivamente en zonas de arena, sin Thalassia testudinum.   Una especie muy abundante fuera de la laguna Strombus gigas no fue encontrada en el sistema lagunar, lo mismo puede decirse de Charonia variegata, Cassis tuberosa, Fasciolaria tulipa y otros gasterópodos muy comunes en la región. Del total de moluscos vivos, los pelecípodos son los más abundantes, alcanzando entre Pterya colymbus, Chione cancellata y Brachidontes citrinus un 86% de la muestra viva, lo que arrojó una diversidad relativamente baja H'= 2.36; J' = 0.64, comparándola con otras regiones del Caribe. Peces: Las colectas de peces fueron realizadas en las cuencas norte, central y sur de la laguna de Nichupté, en los canales de Nizuc y Cancún, así como en la laguna de Somosaya y de Bojórquez. La captura, cuyo propósito fue iniciar con un inventario somero, el estudio de los peces de esta región totalizó 158 especímenes los cuales representan 15 familias, 17 géneros y 25 especies (Tabla 3).  TABLA 3 LISTA DE ESPECIES DE PECES OBSERVADOS (*) Esta lista corresponde al informe solicitado por los autores al investigador Andrés Reséndez Medina sobre la colección de peces de este estudio. Estando este trabajo en prensa apareció la referencia de Reséndez (1975). Sin embargo algunos de ellos como el pargo (Lutjanus griseus), la mojarra blanca (Gerres cinereus) y el sábalo (Megalops atlanticus) fueron observados en cantidades mucho mayores a las capturadas, lo cual se puede deber a que las raíces de mangle forman un refugio efectivo respecto a las artes de pesca utilizadas al menos para el sábalo y el pargo (Bohlke y Chaplin, 1968). La mojarra blanca fue observada en la laguna de Somosaya durante un viaje de reconocimiento; sin embargo, el arrastre de aguas de la llanura de inundación por las lluvias, con alto contenido de materia orgánica y por ende poco oxígeno disuelto, puede haber sido la causa de que esta población no se volviera a encontrar a la hora del muestreo. Otros géneros como Sparisoma, Scarus y A canthurus, aunque reportados como pastoreadores de fanerógamas marinas, entre ellas Thalassia testudinum (den Hartog, 1970) son poco comunes, debido probablemente a que el habitat no es el adecuado para las poblaciones adultas (Randall, 1967). Sin embargo Archosargus rhomboidalis, también pastoreador de fanerógamas marinas (Randall, 1967) es la única especie de este tipo de alimentación que está bien representada, ocupando el segundo lugar (18.0% de la captura) en abundancia en el muestreo. La mojarra blanca (Gerres cinereus) fue abundantemente colectada en la cuenca sur y central de la laguna sobre las zonas densas de Thalassia testudinum, concordando con lo mencionado por den Hartog (1970), sobre el hecho de que esta fanerógama forma un campo de protección. El pargo gris (Lutjanus griseus) también representado abundantemente, es común en estas áreas según Longlye y Hildebrand (1941), mientras que el ronco (Haemulon sciurus) que se considera característico de zonas arrecifales (Longley e Hildebrand, 1941) fue el más abundante en el muestreo- (27.8% de la captura total), Randall (1967) encontró que en algunas ocasiones se encuentran pastos marinos en el contenido estomacal de individuos de esta especie., pero no parece que sea parte de su dieta básica. Sphyraena barracuda también se encontró en una proporción relativamente abundante; sin embargo, aunque presente dentro de la laguna, la mayor parte de la captura fue realizada en los canales de intercomunicación con el mar o en el mar. Aunque el propósito de este muestreo no fue de tipo cuantitativo es interesante observar que de 25 especies registradas, Haemulon sciurus (27.8%), Archosargus rhomboiclalis (18.0%), Lutjanus griseus (17.0%) y Gerres cinereus (8.9%), forman el 73% de la captura en número de individuos; sin emabargo, la diversidad promedio de la muestra es H'= 3.31, J'= 0.78, reflejando la alta diversidad reportada para áreas del Caribe por Caldwell y Caldwell (1964), Bullis y Struhsaker (1970), Randall (1967, 1968), Bohlke y Chaplin (1968), Cervigon (1966), Palacio (1974) y otros. AgradecimientosLos autores hacen patente su agradecimiento a las siguientes personas, sin cuya cooperación no habría sido posible la realización de este estudio: Enrique Lozano, Arturo Toledano y Alfonso Vázquez Botello, por su ayuda en el muestreo de campo. Antonio García-Cubas, Martha Ortega y Andrés Reséndez, por la identificación del material. Agustín Ayala-Castañares por su ayuda en el desarrollo y planeación del trabajo así como en la revisión del manuscrito. Alejandro Yáñez-Arancibia por sus consejos y la revisión del manuscrito. A Yolanda Hornelas por su ayuda en la terminación de este trabajo. LITERATURABERNATOWICZ, A. J. Bull. Mar. Sci. Gulf and Caribbean, Marine Monocotyledoneous plants of Bermuda. 1952 334-335 2 1 BOHLKE, J. E. y C. C. G. CHAPLIN Fishes of the Bahamas and adjacent Tropical Waters. Livingston Publ. Co., Wynnewood, Pa., Acad. Nat. Sci. Phila., 1968 771 p., 223 figs., 36 láms. BRADY, M. J., Sedimentology and diagenesis of carbonate muds in coastal lagoons of NE Yucatan. Tesis doctoral, Rice Univ. Houston, Texas. 1972 BULLIS, H. R. y P. J. STRUHSAKER, Quart. J. Fla. Acad. Sci., Fish fauna of the Western Caribbean upper slope. 1970 43 -76 33 1 CALDWELL, D. K. y M. C. CALDWELL, Allan Hancock Atlantic Expedition, Rept Fishes from the Southern Caribbean collected by Velero III, in 1939 1964 1-61 10 CERVIGON, F., Los peces marinos de Venezuela. Fund. La Salle de Ciencias Naturales 1966 1-451 11 1 y 2 DEN HARTOG, C., The Sea - grasses of the world. North Holland Publ. 1970 275 EKDALE, A. A., Ecology and Paleoecology of marine invertebrate communities in calcareous substrates, Northeast Quintana Roo, México. Tesis doctoral. Rice Univ. Houston, Texas. 1972 HORNELAS, Y., Konig, 1805, en tres diferentes áreas geográficas del Golfo de México Comparación de la biomasa, densidad y algunos aspectos morfométricos de la fanerógama marina Thalassia testudinum. Tesis licenciatura, Fac. Ciencias, UNAM, 1975 HUMM, H. J., Sci. Gulf and Caribbean Epiphytes of the Sea-Grass, Thalassia testudinum, in Florida. Bull. Mar. 1964 306-341 14 2 JORDAN, D. E., M. ANGOT y R. DE LA TORRE Estudio Ecológico de prospección de la laguna de Cancún, Quintana Roo. Informe al Banco de México. Inst. Biol., Univ. Nal. Autón. Méx. 1972 36 LONGLEY, W. H. y S. F. HILDEBRAND, Tortugas lab. Papers 5 (Carnegie Inst. Wash., Publ. 535) Systematic catalogue of the Fishes of Tortugas, Florida. 1941 1-331 LOT, H. A., Estudios sobre fanerógamas marinas en las cercanías de Veracruz, Ver. Tesis Fac. Ciencias, UNAM. 1968 PALACIO, F. J., Bol. Museo del Mar, Peces colectados en el Caribe Mexicano por la Universidad de Miami. 1974 1-137 61 PATRIQUIN, D., Jour. Sci. Estimation of Growth rate, Production and age of the marine angiosperm Thalassia testudinum Koning Carib. 1973 111-124 13 1-2 PHILLIPS, R. C., Fla. St. Bd. Conser. Mar. Lab., Professional Pap. Ser., Observations on the ecology and distribution of the Florida Sea - grasses. 1960 1-72 PIELOU, E. C., Ecological Diversity. J. Wiley & Sons, New York. 1975 RANDALL, J. E. Stud. Trop. Oceanogr. Food habits of reef fishes of the West Indies. Miami 1967 665-847 5 Caribbean Reef Fishes. T. F. H. Publ. Inc. Sersey City, N. J. 1968 RESÉNDEZ-MEDINA, A., Zoología Lista preliminar de peces colectados en las Lagunas de Nichupté y Bojórquez, Q. Roo, México An Inst. Biol. Univ. Nal. Autón. México, Ser. 1975 87-100 46 1 THORSON, G., Treatise on Marine Ecology and Paleoecology. Memoir 67. Bothom Communities. The Geological Society of America, 1957 461-534 1 WARMKE, G. L. y R. T. ABBOT, Caribbean Seashells. Livingston. Publ., Col. Narberth, Pa. 1961 ZlEMAN, J. C., Biscayne Bay. The effects of a thermal effluent strees on the sea-grasses and macroalgae in the vicinity of Turkey Point. Tesis doctoral. University de Miami, USA. 1970
|