|
OSTEOLOGÍA COMPARADA DE DOS ESPECIES DE
ATERÍNIDOS ATHERINOMORUS STIPES (MÜLLER Y IROSCHEL) Y COLEOTROPIS
BLACKBURNI SCHULTZ. (PISCES: ATHERINIDAE)
Trabajo recibido el 31 de marzo
de 1977 y aceptado para su publicación el 13 de junio de
1977.
LILIA J. RUIZ
Universidad de Oriente, Venezuela, Núcleo de
Nueva Esparta, Isla Margarita.
ALEJANDRO YÁÑEZ-ARANCIBIA
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología, Laboratorio de
Ictiología y Ecología Estuarina. Contribución 72 del
Centro de Ciencias del Mar y Limnología, UNAM
El estudio osteológico de
Atherinomorus stipes (Tacnomembrasinae) y de
Coleotropis blackburni (Menidiinae), determina el mejor
criterio para precisar el estatus taxonómico de las dos especies en dos
subfamilias diferentes.
Atherinomorus stipes se diferencia
fundamentalmente de Coleotropis blackburni por presentar
cráneo lateralmente expandido, epiótico sin cresta, prevomer con
dientes, cuatro huesos en la serie suborbital, borde anterior del hocico casi
recto, procesos ascendentes del premaxilar anchos y truncados en su borde
superior, premaxilar con dientes en bandas viliformes reflejados sobre la cara
externa, proceso coronoides del dentario corto, endopterigoides dentado,
presencia del primer par de infrafaringóbranquiales, dos hileras de
radiales pectorales menor número de vértebras caudales, origen de
la primera dorsal muy por delante del origen de la anal.
The osteologic study of Atherinomorus
stipes (Taenomenbrasinae) and Coleotropis
blackburni (Menidiinae) provides the best criterion to establish the
taxonomic classification of two species in two different subfamilies.
Atherinomorus stipes differs from
Coleotropis blackburni because it has a laterally expanded
skull, an epiotic without crest, prevomer with teeth, four bones in the
suborbital group, anterior edge of the snout almost straight, wide and truncate
premaxillary ascendent proceses in the upper adge, premaxillaries with teeth
and villiforme edges reflected on the external face, short coronoide process of
the dentary, endopterigoides with teeth, presence of the firts pair of
infrapharyngeobranchiales, two rows of pectoral rays, fewer caudal vertebrae,
origin of the first dorsal farther over the origin of the anal fin.
Existe muy poca literatura sobre la osteología de Atherinidae. Starks (1899) se refiere a una comparación parcial del endoesqueleto de las familias Atherinidae, Mugilidae y Sphyraenidae, consideradas anteriormente como pertene cientes al suborden Percesoces. Hollister (1937) estudió el esqueleto caudal del mismo suborden y Gosline (196la) hizo estudios sobre el esqueleto caudal de los representantes de los Mugiliformes, Beloniformes y Cyprinodontiformes, con el objeto de presentar nuevas consideraciones sobre sus relaciones y posición taxonómica, concluyendo en separar a los Atherinidae del orden Mugiliformes y crear el orden Atheriniformes. Greenwood et al. (1966) en un estudio sobre la clasificación de los peces óseos actuales y considerando los criterios anteriores, proponen el superorden Atherinomorpha. Recientemente, Gosline (1971) también considera la separación de Atherinidae del antiguo orden Mugiliformes, así como también Rosen (1973). En las costas de Venezuela, la familia Atherinidae está representada por seis especies, a saber: Atherinomorus stipes, (Müller y Troschel), Allanetta harringtonensis (Goode), Melanorhinus microps (Poey), Xenomelaniris brasiliensis (Quoy y Gaimard), Coleotropis blackburni Schultz y Adenops analis Schults (Cervigón, 1966, 1973 y 1975). Su posición taxonómica se ha señalado sobre la base de diferencias merísticas y morfométricas. Las dos primeras especies han sido ubicadas por Schultz (1948) en la subfamilia Taenomembrasinae y las cuatro últimas en la Menidiinae. Este autor hace una revisión de seis subfamilias y describe nuevos géneros y especies, considerando algunos caracteres osteológicos, tales como procesos maxilares y número de vértebras. Por otra parte, señala que para precisar el estatus y las relaciones de los diferentes géneros de Atherinidae, es necesario hacer algunos estudios especialmente sobre osteología; de esa manera, ha encontrado que algunos de los géneros descritos por Jordan y Hubbs (1919) no se encuentran relacionados siendo su estatus taxonómico poco preciso. En realidad la taxonomía moderna supraespecífica en los peces se basa fundamentalmente en estudios de osteología comparada (Greenwood et al.,1966; Greenwood et al., 1973; Nybelin, 1973; Rosen, 1973). No existe ningún trabajo sobre la osteología de las especies de Atherinidae de Venezuela por lo cual fueron seleccionadas dos especies, una de cada subfamilia, para estudios osteológicos y su comparación. En la taxonomía moderna son de suma importancia los estudios osteológicos por las relaciones filogenéticas de muchas estlucturas óseas (Greenwood et al., 1966; Gosline, 1961a y b, 1965 y 1971). Las especies consideradas en este estudio tienen la siguiente distribución geográfica. Atherinomorus stipes, desde el sur de Florida hasta Brasil y el Golfo de México, y Coleotropis blackburni, en el Pacífico de Panamá y, en el Atlántico, desde el Golfo de Venezuela hasta Brasil. En las costas de Venezuela, A. stipes es frecuente en las orillas de las playas y en las lagunas litorales con manglares. Por otra parte C. blackburni, se encuentra asociada a playas abiertas y batidas, de aguas limpias y fondos arenosos, normalmente cerca de rocas y acantilados (Cervigón, 1973). De las seis especies de Atherinidae marinas conocidas para Venezuela, C. blackburni se diferencia claramente porque el origen de la primera dorsal está situado por detrás del origen de la anal; además, por el número de radios de la anal que es superior a todas las demás especies. MATERIAL Y MÉTODOS
MATERIAL ESTUDIADOLos ejemplares estudiados provienen de colectas realizadas en la Laguna La Restinga (Edo. Nueva Esparta, Venezuela). Atherinomorus stipes, se colectó dentro de la laguna y Coleotropis blackburni a orillas de la playa. Se seleccionaron 30 ejemplares de cada especie, comprendidos entre 28 y 81 mm de longitud estandar (LE). La colección fue separada en clases y se analizó la osteología de los ejemplares de cada clase para observar la evolución de la osificación, sobre todo de la región caudal, y la posible variación de forma y estructura de los huesos. Algunos ejemplares fueron observados preservados para precisar ciertas uniones por ligamentos. Para el estudio osteológico el esqueleto fue separado en regiones, las cuales fueron examinadas al microscopio estereoscópico y descritos los huesos que las constituyen para Atherinomorus stipes. Al final de la descripción de cada región se señalan las diferencias que existen en dicha región para Coleotropis blackburni. Los dibujos fueron realizados con cámara clara y están basados en un ejemplar de 69.5 mm de LE de Atherinomorus stipes y un ejemplar de 78.0 mm de LE de Coleotropis blackburni. La nomenclatura utilizada para los huesos fue la siguiente: Cráneo, D'viller (1958); Aparato branquial, Nelson (1969); Cintura pectoral y pélvica, Chapman (1944); Región circumorbital, Smith y Baley (1962); Esqueleto caudal, Monod (1968). METODOLOGÍALos ejemplares fueron fijados en formol al 10% previo al aclarado y teñidos por el método enzimático (Taylor, 1967), para poner en evidencia sus estructuras óseas. Una vez aclarados y coloreados fueron medidos (LE) y contadas sus vértebras, radios y espinas de las aletas. Reactivos utilizados1. Alizarina 2. Hidróxido de potasio 3. Agua destilada 4. Peróxido de hidrógeno al 3% 5. Borato de sodio 6. Glicerina 7. Tripsina 8. Cristales de Timol Soluciones utilizadas1. Agua destilada saturada con borato de sodio 2. Solución al 1% de hidróxido de potasio 3. Solución buffer enzimática. Mezcla de 3 partes de la solución saturada de borato de sodio con 7 partes de agua destilada. 4. Mezclas de glicerina e hidróxido de potasio: 20 : 80; 40 : 60; 70 : 30; 90 : 10; y finalmente glicerina pura. Desarrollo1. Los especímenes fueron fijados en formol al 10% y dejados en esta solución durante 15 días. La solución se neutraliza agregando una cuchara de borax por litro de solución. 2. Se sacaron los especímenes de la solución fijadora y fueron lavados con cambios repetidos de agua hasta que el formol desapareció. 3. Se blanquearon los especímenes en una solución al 20%, de peróxido de hidrógeno y al 80% de la solución de hidróxido de potasio. Debido a que esta reacción es acelerada por la luz, el material fue colocado en un envase transparente debajo de una lámpara. Las cavidades del cuerpo fueron abiertas para que penetrara la solución. El aclarado se continuó hasta que las áreas obscuras llegaron a ser pálidas, el tiempo varió dependiendo de la pigmentación de los ejemplares. 4. Se colocaron los especímenes en la solución buffer con un volumen 40 veces mayor que el volumen de los peces. Un mínimo de 200 ml de la solución es satisfactorio para ejemplares pequeños. 5. Se agregó la enzima. Una cantidad de más o menos 0.45 g de enzima es suficiente para 400 ml de solución. Se mezcló la enzima sin hacer espuma. El material se dejó a temperatura ambiente. 6. El tiempo de digestión varió. La solución digestiva se cambió cada ocho días. El aclarado de los especímenes se nota primero en los bordes del cuerpo y en las partes delgadas como la pared abdominal, pedúnculo caudal y aletas. El ejemplar pierde rigidez hasta ser muy flexible. 7. Se preparó la solución colorante mezclando una pequeña cantidad de alizarina en suficiente solución de hidróxido de potasio; se sumergieron los especímenes en esta solución y se agregó alizarina hasta alcanzar un color púrpura o la intensidad de color deseada. Los especímenes se dejaron en la solución coloreada hasta que los huesos estuvieron adecuadamente teñidos. 8. Se sacaron los especímenes de la solución colorante. Fueron lavados en agua destilada, evicerados, y se les quitaron las escamas. 9. Se lavaron los especímenes en agua destilada y se colocaron nuevamente en la solución digestiva hasta que todas las áreas de tejido hubieron desaparecido. 10. Luego de la digestión los especímenes fueron trabajados en solución de glicerina e hidróxido de potasio hasta llegar a glicerina pura. Se agregaron unos pocos cristales de timol a la glicerina final para evitar el desarrollo de microorganismos. RESULTADOS
OSTEOLOGÍA DE ATHERINOMORUS STIPES (MÜLLER Y TROSCHEL)
NEUROCRÁNEO
A. Región occipitalLa región occipital está formada por cuatro huesos, un supraoccipital dorsal, un basioccipital ventral y dos occipitales laterales, en posición lateral (Fig. l). Supraoccipital. La porción media anterior se proyecta hacia adelante situándose por debajo de la unión posteromedial de los frontales. La cresta supraoccipital es de pequeño tamaño, y no se proyecta más allá del extremo posterior del cóndilo occipital. Occipital lateral. No descansa sobre el basioccipital, se extiende hacia la parte superior para unirse con su homólogo del lado opuesto, debajo de la cresta supraoccipital; forma junto con el basioccipital el foramen magnum. Cada occipital lateral presenta un cóndilo de articulación para la primera vértebra precaudal. Basioccipital. No muy pronunciado hacia su parte posterior, se articula por su borde anterior con los proóticos y por su borde dorsal con los occipitales laterales; forma junto con los huesos anteriores parte de la vesícula ótica donde se aloja el otolito de mayor tamaño. La porción anterior al cóndilo del basioccipital se curva en forma pronunciada hacia abajo. 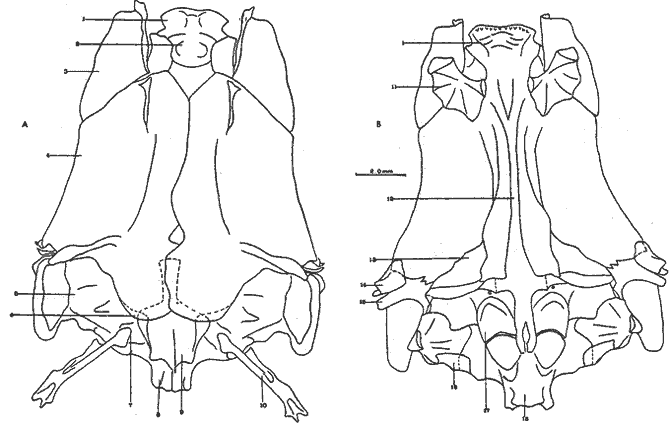 Fig. 1. Neurocraneo de Atherinomorus stipes. A. Vista Dorsal. B. Vista ventral. 1. Prevomer. 2. Etmoides. 3. Nasal. 4. Frontal. 5. Pterotico. 6. Parietal. 7. Epiotico. 8. Occipital lateral. 9. Supraoccipital. 10. Postemporal. 11. Etmoides lateral. 12. Parasfenoides. 13. Pleurosfenoides. 14. Dermosfenotico. 15. Esfenotico. 16. Intercalar. 17. Prootico. 18. Basioccipital. B. Región etmoidalLa región etmoidal comprende el etmoides, el prevómer, los dos nasales y dos etmoides laterales. Etmoides. Es un hueso delgado, de forma discoidal, el cual descansa sobre la porción antero dorsal del prevómer. Los bordes laterales se articulan con el etmoides lateral (existe una cuña cartilaginosa entre este hueso y el etmoides lateral). Prevómer. Es de forma triangular, su extremo posterior puntiagudo se articula con el parasfenoide; su porción anterior es ensanchada; su borde ventral anterior cóncavo presenta una hilera de dientes. En su extremo anterior en su superficie dorsal descansa la cabeza del premaxilar. Presenta a ambos lados de su extremo anterior una escotadura donde descansa el extremo anterior del autopalatino. Nasal. Es de forma triangular, su borde posterior se une fuertemente al borde anterior del frontal. En la superficie interna presenta un pliegue que forma un canal, el cual se continúa con un pequeño pliegue del frontal. La superficie ventral anterior descansa sobre el lagrimal y la superficie ventral posterior sobre la parte anterior al etmoides lateral.Etmoides lateral. Es un hueso de forma irregular situado debajo del nasal y parte anterior del frontal; presenta una superficie articular en su porción posterior interna con el vómer y una externa con el lagrimal; su borde interno anterior, se articular al etmoides, C. Región temporalFormada por dos frontales, dos pleurosfenoides, dos parietales, un basisfenoides y el parasfenoides; los orbitosfenoides están ausentes. Frontal. Los frontales cubren la mayor parte de la superficie dorsal del cráneo, la parte posterior cubre parte del supraoccipital; los bordes laterales externos de la parte posterior se continúan con los esfenóticos; en la unión del frontal y esfenótico se articula el dermosfenótico. En su superficie postero dorsal externa presenta un pliegue que forma un canal que se continúa con un canal en la región lateral externa del Pterótico. Su superficie lateral externa correspondiente a las órbitas, presenta una fuerte concavidad ventral para alojar el gran ojo que prescnta esta especie. Pleurosfenoides. Laminar, curvado en su borde externo, situado en la porción lateral anterior de la base del cráneo; se articula al proótico por su borde dorsal y por su borde lateral externo anterior al frontal. El Pleurosfenoides no se relaciona con el basisfenoides. Parietal. Laminar cubierto por su borde anterior por el frontal; cubre parte de la superficie dorsal de la porción lateral posterior externa del supraoccipital y parte del epiótico y pterótico. Basisfenoides. Bien desarrollado, situado en la base del cráneo; presenta en su parte media un proceso amplio dirigido hacia la región anterior, el cual se articula al parasfenoides; por sus bordes laterales y posterior se articula a los proóticos. Parasfenoides. El parasfenoides se articula por su extremo anterior con el prevómer; la estructura alar del parasfenoides se articula entrelazándose fuertemente con dos prolongaciones dirigidas ventralmente desde los proóticos; su extremo posterior bifurcado no se prolonga más allá del tercio anterior basal del basioccipital. D. Región ótica.Está formada por un proótico, un epiótico, un pterótico, un esfenótico y un intercalar u opistótico de cada lado del cráneo. Proótico. Los proóticos se encuentran situados en la región ventrolateral anterior de la parte posterior del neurocráneo, uniéndose en su parte media para formar la base de la caja craneana junto con el basioccipital; presentan en su parte interna anterior una cavidad donde se aloja el otolito, más pequeño; por sus bordes posterior se unen al basioccipital y por su borde superior al occipital lateral, para formar juntos la ámpula que aloja el otolito de mayor tamaño. Epiótico. Es un hueso con la parte anterior de forma triangular con su base cóncava situada entre la porción posterolateral externa del supraoccipital y el pterótico; en su lado dorsal presenta una superficie de articulación donde descansa la porción anterior del proceso superior del postemporal. Pterótico. Presenta en su borde externo una estructura laminar doble, la cual forma un canal que se continúa con el frontal; se articula por su borde interno con el proótico y el occípital lateral; por su parte anterior se articula al esfenótico. Esfenótico. Situado en la parte antero lateral de la parte anterior de la caja craneana, cubierto por el extremo posterolateral externo del frontal; presenta el proceso postorbitario dirigido hacia abajo y hacia atrás. Intercalar. Laminar, adosado al occipital lateral, entre la sutura con el pterótico, presenta una proyección engrosada, en la cual se articula, por ligamento, el proceso inferior del postemporal. E. Serie orbital o circumorbitalLa serie orbital es incompleta, no existen huesos supraorbitales; está formada por cuatro huesos, los cuales constituyen la serie suborbital. El primer hueso de la serie suborbital es el lagrimal y el último es el dermosfenótico (Fig. 2). El lagrimal o antorbital (Gosline, 1965). Se articula por su parte dorsal con el nasal y por su parte anterior con el etmoides lateral. Su extremo anterior protege la porción anterior del maxilar. El segundo tiene forma alargada articulándose por su borde anterior con el lagrimal y por su borde posterior con el tercero de la serie; descansa sobre el borde externo, en la porción media del maxilar. El tercero de la serie es de pequeño tamaño, descansa sobre la superficie externa, posterior del maxilar, y se articula por su parte posterior con el extremo distal del articular. El sexto o dermosfenótico es angosto, alargado y presenta un surco en su porción media; se articula por su extremo superior con la porción posterior externa del frontal y el esfenótico. El cuarto y el quinto de la serie están ausentes. La tendencia evolutiva es hacia la reducción del número de huesos suborbitales. 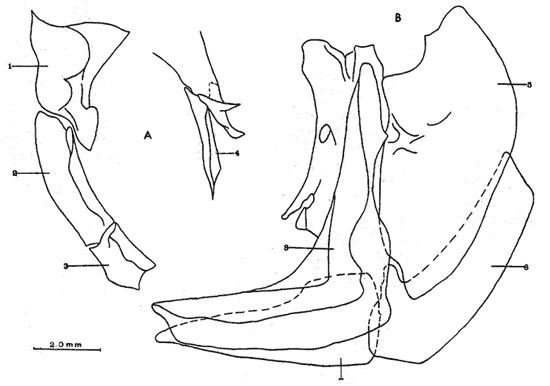 Fig. 2. A. Serie orbital de Atherinomorus stipes. B.Serie opercular de Atherinomorus stipes. 1. Lagrimal. 2.Primer suborbital. 3.Segundo suborbital. 4.Tercer suborbital (dermosfenótico). 5.Opérculo. 6.Subopérculo. 7.Interopérculo. 8. Preopérculo. F. Serie opercularLa serie opercular está formada por cuatro huesos de membrana, el opérculo, subopérculo, interopérculo y peropérculo; aunque este último no es referido como miembro de la serie opercular, Hubbs (1919) (Fig. 2). Opérculo. Es el hueso más grande de la serie opercular, tiene forma de una placa subcuadrada, curvada a lo largo de su borde dorsal y anterior, presenta en el lado interno del borde anterior una concavidad donde se articula el cóndilo posterior del hiomandibular. Subopérculo. Es de forma alargada en sentido anteroposterior y presenta un proceso en el extremo superior de su borde anterior; su superficie supero dorsal externa se encuentra cubierta por el margen inferior del opérculo. Interopérculo. Es alargado en sentido antero posterior, su extremo anterior aguzado se articula con el extremo posterior del articular y su borde posterior ensanchado se articula con el borde anterior del subopérculo. Su superficie externa está cubierta por la porción inferior del preopérculo; su superficie interna descansa sobre el epihial y parte del ceratohial. Preopérculo. Está formado por dos porciones, una porción que se prolonga hacia la parte superior y una hacia la parte anterior, formando ambas porciones ángulo recto. Presenta un pliegue a todo lo largo del borde interno del hueso que forma un canal, el cual se continúa con el tercer suborbital. G. Radios branquiostegosExisten seis radios branquiostegos, los dos anteriores, delgados y más cortos que los posteriores, están adheridos en la parte posterior del borde inferior en la escotadura del ceratohial; dos en la superficie externa de la parte posterior ensanchada del ceratohial y los dos posteriores en la superficie externa del epihial (Fig. 4). 2. BRANQUIOCRÁNEO
A. Región oromandibularComprende una porción superior constituída por dos palatinos, dos cuadrados, dos ectopterigoides, dos endopterigoides, dos metapterigoides, dos premaxilares y -dos maxilares; y una porción inferior constituída por dos dentarios, dos articulares y dos angulares (Fig. 3). Autopalatino. Presenta una porción anterior adelgazada y una porción laminar expandida; el extremo posterior de la región anterior se articula al prevómer, su borde ventral descansa sobre un canal de la porción anterior del maxilar, su borde lateral interno se une al borde extermo del nasal, su borde lateral externo se articula al etmoides lateral. El extremo posterior de la porción laminar se articula al borde externo del ectopterigoides. Cuadrado. Tiene forma triangular; el borde ventral del hueso presenta un largo, proceso dirigido posteriormente, el cual cubre parte del simpléctico. Ectopterigoides. Es de pequeño tamaño, alargado y delgado. Se articula con la parte posterior del autopalatino, por su borde anterior; cubre con su borde interno el borde ventral del endopterigoides; se extiende debajo a más de la mitad del borde antero ventral del cuadrado. Endopterigoides. Es de gran tamaño, extendiéndose ampliamente al piso de la órbita; su borde posterior se une al borde anterior del metapterigoides. Es delgado y lleva en su superficie ventral media una hilera discontínua de pequeños dientes. Su borde externo se curva hacia abajo para articularse al ectopterigoides, cuadrado, simpléctico y metapterigoides. Metapterigoides. Es alargado y laminar, su borde posterior se articula al borde ventral de la porción inferior de hiomandibular y al proceso lateral externo, quedando entre ambos huesos un orificio. Premaxilar. Con procesos ascendentes ensanchados, truncados en su borde superior, situados en un canal formado por dos procesos originados en la porción anterior del maxilar. La porción proximal del hueso, presenta un parche de dientes dirigidos hacia adelante y varias hileras de dientes viloformes sobre su superficie inferior externa. Maxilar. La cabeza del maxilar presenta en su parte posterior un proceso dirigido hacia adelante, el cual se articula con el prevómer y autopalatino y un pliegue o extensión laminar en su parte anterior, el cual forma un canal para recibir el proceso ascendente del premaxilar. La parte posterior del hueso descansa sobre la superficie externa del premaxilar. El maxilar y premaxilar sobrepasan el extremo posterior del dentario y en su totalidad forman el borde de la boca. Dentario. El borde anteroventral del hueso muy curvado; presenta hileras de dientes a todo lo largo de su borde dorsal y un parche de dientes en la porción anterior externa. El proceso coronoides no está pronunciado hacia arriba y hacia atrás, su borde posterior recto; presenta un canal en su porción ventral y un conducto en su parte media, el cual desemboca en la parte posterior y donde se aloja el extremo anterior del articular. Articular. La mitad superior del hueso es laminar y se prolonga hacia adelante en un largo proceso puntiagudo, el cual se articula a la porción interna media del dentario. La mitad inferior es engrosada, con una superficie de articulación en la mitad posteroventral del hueso; su extremo posterior presenta en su superficie dorsal una concavidad donde se aloja el cóndilo del cuadrado. Angular. Es un pequeño hueso, articulado fuertemente al extremo posteroventral del articular, debajo de la concavidad de articulación con el cuadrado. 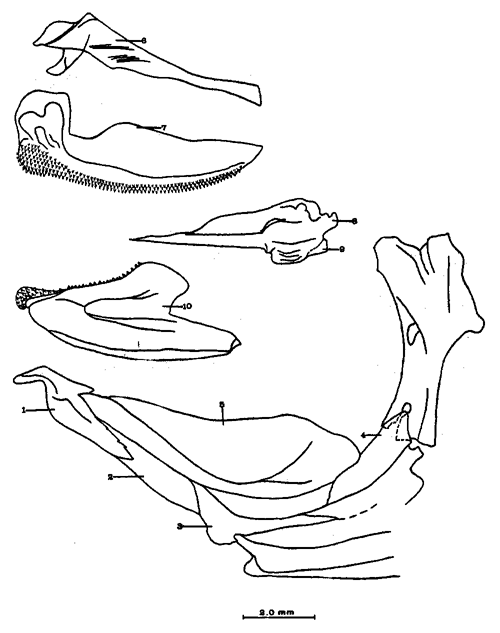 Fig. 3. Branquiocraneo de Atherinomorus stipes. Región oromandibular. 1. Autopalatino. 2. Ectopterigoides. 3. Cuadrado. 4. Metapterigoides. 5. Endopterigoides. 6. Maxilar. 7. Premaxilar. 8. Articular. 9. Angular. 10. Dentario. B. Región hioideaEstá formada por el hiomandibular, interhial, simpléctico, epihial, ceratohial, hipohial inferior, hipohial superior, a cada lado de la región branquial, y el parahioides situado en la región media ventral (Fig. 4). Hiomandibular. La parte anterior o cabeza del hiomandibular presenta en su parte superior dos cóndilos de articulación, el anterior se articula al esfenótico y el posterior al pterótico; en el borde lateral interno presenta un tercer cóndilo de articulación para el opérculo. La mitad inferior del hueso, denominada tallo, es maciza en su porción media, con un pequeño pliegue en su cara externa, el cual se relaciona con el preopérculo, y una extensión laminar en su borde externo, el cual presenta un proceso donde se articular el metapterigoides; la superficie ventral de la porción maciza del tallo, se articula al simpléctico e interhial. Simpléctico. Presenta una porción posterior expandida cuyo borde dorsal se une al borde ventral del metapterigoides, y el borde ventral está cubierto en su parte anterior por el cuadrado y en su parte posterior por el preopérculo; su extremo anterior adelgazado se articula a la superficie ventral del tallo del hiomandibular. Interhial. Es de pequeño tamaño, de forma cilíndrica, articulado por su superficie ventral a un proceso de la porción posterior del epihial. Epihial. Es de forma subtriangular, alargado, está unido fuertemente al ceratohial por un puente óseo. Ceratohial. La mitad proximal del hueso es angosta y la mitad distal es abruptamente ensanchada; el extremo anterior del hueso presenta dos procesos, uno superior corto y uno inferior largo, entre los cuales se articulan los hipohiales. Hipohial inferior. Es de forma semicónica, con su extremo puntiagudo situado entre los dos procesos anteriores del ceratohial; su extremo anterior presenta en su parte superior interna una concavidad. Hipohial superior. Es de pequeño tamaño, de forma cónica con su base situada en una concavidad de la parte superior interna del hipohial inferior. Parahioides. Es laminar, alargado y ensanchado posteriormente, situado en posición vertical debajo de los basibranquiales. En su borde ventral presenta dos láminas dirigidas oblícuamente hacia afuera, dejando entre ellas un canal. La parte anterior del hueso es angosta y engrosada con dos superficies de articulación de las cuales parte un ligamento a la superficie interna de cada hipohial inferior; presenta en la superficie dorsal anterior un proceso laminar, el cual se dirige oblícuamente hacia atrás para articularse a la parte posterior del segundo basibranquial y a los procesos dirigidos ventralmente del primer par de hipobranquiales. 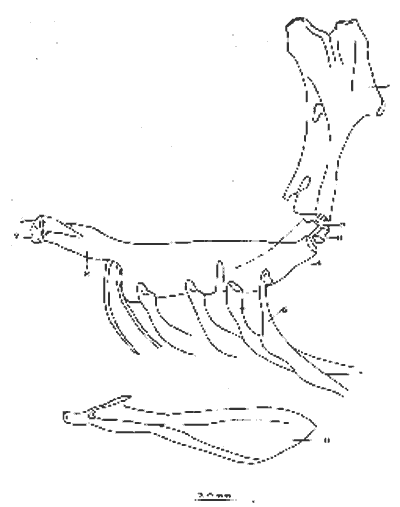 Fig. 4. Branquiocráneo de Atherinomorus stipes. Región hioidea. 1. Hiomandibular. 2. Simpléctico. 3. Interhial. 4. Epihial. 5. Radios branquiostegos. 6. Ceratohial. 7. Hipohial superior. 9. Parahioides. C. Aparato branquialLos arcos branquiales están formados por elementos endocondrales, los cuales incluyen un basihial, tres basibranquiales, tres pares de hipobranquiales y cinco pares de ceratobranquiales en la región ventral; cuatro pares de epibranquiales, y cuatro pares de infrafaringobranquiales. Además comprende elementos dérmicos, los cuales están representados por placas dentarias, adheridas o fusionadas a los elementos endocondrales que los sostienen, son la excepción de las placas dentarias infrafaringeales situadas sobre los ceratobranquiales y los suprafaringeales asociados con los infrafaringobranquiales (Fig. 5). Basihial. De forma triangular y cubierto en su superficie dorsal posterior por una placa dérmica, laminar, con pequeños dientes aislados. El basihial no se prolonga más allá de la articulación de los hipohiales. Basibranquiales. Existen cuatro basibranquiales. El primero, segundo y tercero presentan placas dérmicas dentadas fusionadas a la superficie dorsal; sobre el extremo posterior del tercer basibranquial, existe una pequeña placa dentada no fusionada a este elemento, la cual se extiende lateralmente cubriendo parte del extremo posterior del tercer par de hipobranquiales. El primer basibranquial presenta un proceso laminar ventral dirigido hacia la parte anterior, el cual se relaciona con los hipohiales. Hipobranquiales. El primer par de hipobranquiales presenta en la región ventral anterior un proceso dirigido hacia abajo, el cual se relaciona con su homólogo del lado opuesto y constituye junto con la porción posteroventral del primer basibranquial superficie de articulación del parahioides; se articulan por su extremo anterior a la región anterolateral del segundo basibranquial. El primer par de hipobranquiales presentan una placa dérmica dentada en la región media dorsal fusionada al elemento endocondral. El segundo par se articula a una escotadura de la región anterior del tercer basibranquial; presenta una placa dérmica dentada. El tercer par es el más pequeño, presenta en la región ventral un proceso dirigido hacia la parte posterior; se articula por su borde interno a la región posterolateral del tercer basibranquial. Ceratobranquiales. De los cinco pares de ceratobranquiales los cuatro anteriores son alargados; el primero, segundo y tercero se articulan por su extremo anterior al extremo posterior de los tres primeros hipobranquiales, respectivamente; el cuarto y quinto par se relacionan por su extremo anterior al tercer basibranquial. El cuarto par presenta pequeñas placas de forma oval con proyecciones en forma de dedo a lo largo de toda su superficie dorsal. El quinto par de ceratobranquiales presentan en la porción media anterior de la región ventral, una extensión laminar dirigida hacia abajo; presenta en su superficie dorsal un par de placas dérmicas dentadas, de forma triangular, las cuales se fusionan al elemento endocondral. Epibranquiales. El primer par de epibranquiales se articula por su extremo anterior a la región posterolateral externa del primer par de infrafaringobranquiales; el segundo par se articula al segundo par de infrafaringobranquiales; y el tercero y cuarto par a la parte posterior del tercer y cuarto infrafaringobranquiales fusionado. Cada epibranquial presenta un proceso dirigido hacia la parte posterior, el cual constituye la superficie de articulación para el siguiente. Infrafaringobranquiales. En Atherinomorus stipes existen cuatro pares de infrafaringobranquiales. El primer par es de pequeño tamaño, angosto y curvado; su extremo dorsal se articula al proótico, conectando el aparato branquial al cráneo. El segundo par presenta en su superficie ventral una placa dérmica dentada de forma alargada. El tercer y cuarto infrafaringobranquial se fusionan y están cubiertos por una placa dentada expandida. Suprafaringeales. Hay tres pares de placas suprafaríngeas dentadas. El primer par o par anterior, se fusiona al segundo par de infrafaringobranquiales; el segundo par se fusiona al tercer y cuarto par de infrafaringobranquiales; el tercer par se articula a la parte posterior del segundo par y está situado sobre la región anterior media de los epibranquiales tres y cuatro. Branquiespinas. La hilera interna del primer arco, las del segundo y parte del tercero, se presentan como pequeñas láminas triangulares con diminutos dientes en su margen superior. Las branquiespinas del cuarto par de arcos branquiales se fusionan formando placas extendidas transversalmente sobre la superficie dorsal del ceratobranquial, la placa más interna cubre parte del extremo posterior del quinto par de ceratobranquiales. 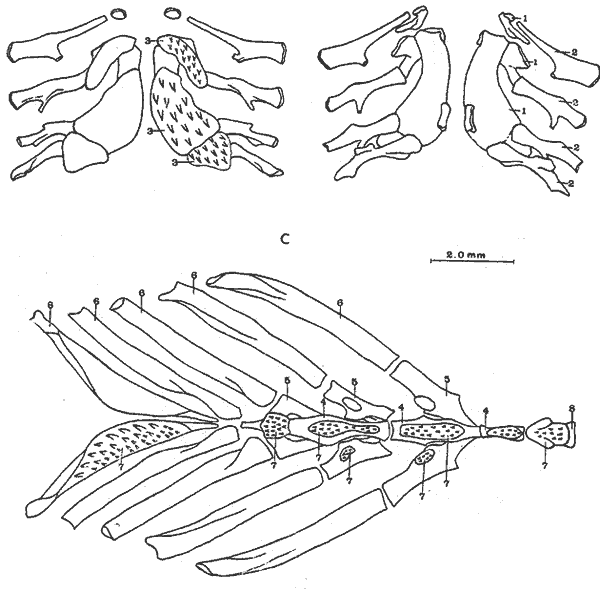 Fig. 5. Branquiocráneo de Atherinomorus stipes. Parte superior de la región branquial. A. Vista ventral. B. Vista dorsal. C. Parte inferior de la región branquial, vista dorsal. I. Infrafaringobranquiales. 2. Epibranquiales. 3. Placas faríngeas superiores. 4. Basibranquiales. 5. Hipobranquiales. 6. Ceratobranquiales. 7. Placas faríngeas inferiores. 8. Basihial. 3. CINTURA ESCAPULARLa cintura escapular está compuesta de una serie de huesos que conectan al cráneo con la aleta pectoral. Un postemporal, un supracleitrum, un postcleitrum, el cleitrum, una escápula, un coracoides y cuatro pterigióforos; a este último se articulan los radios de la aleta pectoral (Fig. 6). Postemporal. Presenta en su porción anterior dos ramas, una superior que se articula al epiótico y una inferior que se une al opistótico o intercalar. Supracleitrum. Tiene forma de escama oval cuyo extremo anterior se articula con la superficie interna de la parte posterior del postemporal y su extremo posterior descansa sobre la superficie anterior externa del cleitrum. Cleitrum. Constituido por varias láminas, las cuales le dan forma compleja; el cleitrum se une por el borde postero superior de la lámina interna a la escápula y por su borde postero inferior al coracoide. Se relaciona por su extremo inferior con su homólogo en la línea media ventral del cuerpo. Postcleitrum. Alargado, laminar formado por una porción superior curvada, la cual se articula a la superficie interna de una corta extensión de la parte superior del cleitrum; se dirige hacia la línea ventral del cuerpo para articularse a la porción inferior, la cual es ensanchada y se articula por su superficie interna con la primera y segunda costillas pleurales. Escápula. Laminar de forma subrectangular o casi rectangular, se articula por su borde superior y anterior con el cleitrum y por su borde ventral con el coracoides, quedando un orificio entre ellos. En su borde posterior se articula dos de los cuatro pterigióforos. Coracoides. Laminar de forma rectangular, alargado en su eje súpero inferior, se articula al cleitrum por su borde ántero superior y ántero inferior, quedando entre ellos una abertura. Actinosteos o radiales pectorales. Existen dos hileras de radiales pectorales o actinosteos, la hilera proximal está formada por cuatro elementos de forma cuboidal, los dos anteriores situados en el borde dorsal de la escápula y los dos posteriores unidos al coracoides; la hilera distal consta de doce elementos óseos de pequeño tamaño, situados en el extremo proximal de los lepidotrichia de la aleta pectoral. 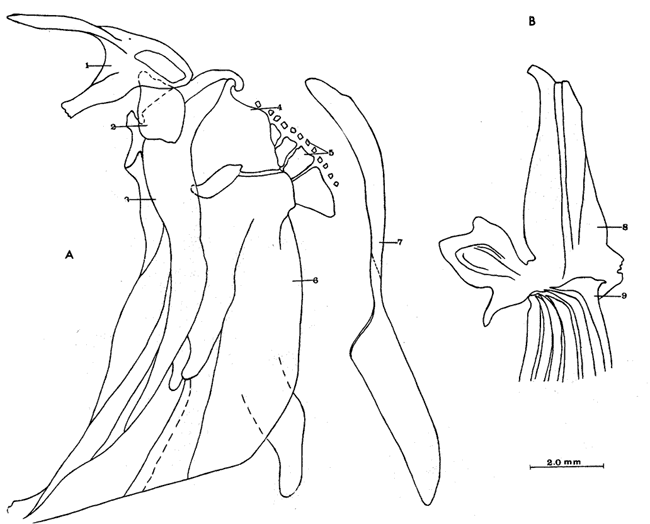 Fig. 6. A. Cintura escapular de Atherinomorus stipes. B. Cintura pélvica de Atherinomorus stipes. 1. Postemporal. 2. Supracleitrum. 3. Cleitrum. 4. Escápula. 5. Radiales pectorales. 6. Coracoides. 7. Postcleitrum. 8. Basipterigios. 9. Espina. 4. CINTURA PÉLVICALa cintura pélvica está formada por un par de basipterigios más o menos separados en su parte anterior interna y unidos en su parte posterior interna por una extensión laminar que emite cada hueso y que se sobreponen una a otra. En el extremo posterior externo presenta un corto proceso, el cual se articula con el extremo posterior de la cuarta costilla pleural (Fig. 6). 5. COLUMNA VERTEBRALLos especímenes examinados en A. stipes presentan de treinta y seis a treinta y ocho vértebras, raramente treinta y nueve incluyendo el urostilo; de las cuales de diecisiete a veinte son precaudales y de dieciocho a diecinueve, son caudales raramente veinte (Fig. 7). Todas las vértebras precaudales llevan parapófisis las cuales se proyectan ventralmente del extremo anterior del centrum. Las parapófisis se expanden a partir de la tercera vértebra, presentando en su superficie posterior, una concavidad para articular las costillas pleurales. Las parapófisis emiten dos procesos ventrales que se unen para formar el arco hemal y la espina hemal a partir de la primera vétebra caudal. Las espinas neurales se presentan en todas las vértebras excepto en la primera donde es rudimentaria, en las siguientes ocho o nueve vértebras las espinas neurales son expandidas y achatadas lateralmente, formando crestas neurales rectangulares las cuales cubren toda o casi toda la porción dorsal de las vértebras, para luego angostarse y desarrollarse como espina hacia las vértebras posteriores. Existen dieciseis pares de costillas pleurales, el último par muy reducido. La primera costilla pleural está sobre la tercera vértebra y está articulada por ligamento a la cintura escapular por medio del postcleitrum, fijándola firmemente al esqueleto axial. El extremo libre de la cuarta costilla, está unido firmemente a la parte posterior de los basipterigios de la cintura pélvica. Las costillas pleurales de las últimas vértebras precaudales son más cortas. Las epipleurales se presentan a partir de la primera vértebra. La primera articulada por ligamento al borde interno superior del postcleitrum, las dos primeras pegadas directamente a las parapófisis y las restantes a las costillas pleurales. 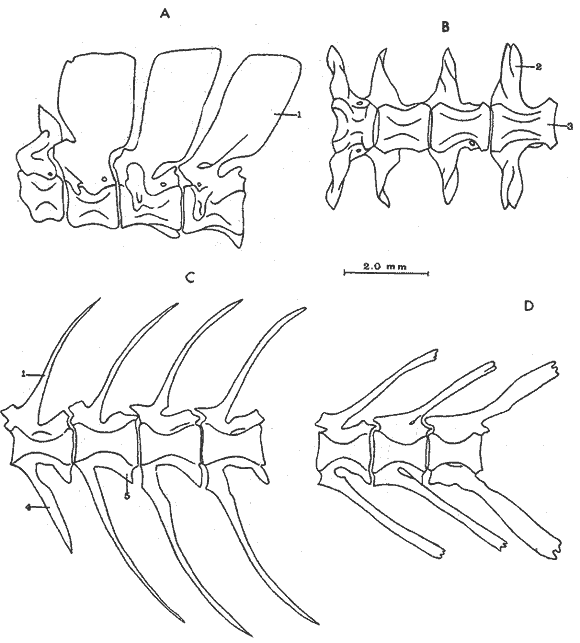 Fig. 7. Columna vertebral de Atherinomorus stipes. A. Primeras vértebras Precaudales, vista lateral. B. Primera vértebras precaudales, vista ventral. C. Primeras vértebras caudales, vista lateral. D. Últimas vértebras caudales, vista lateral. 1. Espina neural. 2. Parapofisis. 3. Centrum vertebral. 4. Espina hemal. 5. Proceso posteroventral. 6. ALETAS DORSALESLa primera aleta dorsal presenta de cuatro a seis pterigióforos, el primero es de forma triangular y expantdido hacia la parte anterior. El número de espinas es generalmente de cuatro a seis (Fig. 8). La segunda dorsal presenta una espina y nueve radios, soportados por diez pterigióforos. Los pterigióforos se proyectan hacia abajo entre las espinas neurales. Entre la primera y segunda dorsal existen cuatro ó cinco supraneurales, situados a lo largo de la línea media dorsal. 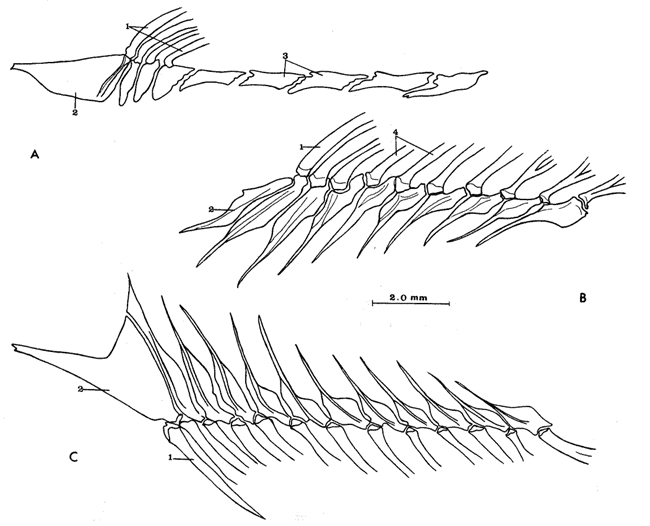 Fig. 8. A. Primera aleta dorsal de Atherinomorus stipes. B. Segunda aleta dorsal. C. Aleta anal. 1. Espinas. 2. Pterigióforos. 3. Supraneurales. Radios. ALETA ANALLa aleta anal presenta una espina y once ó trece radios, los cuales están soportados por doce ó trece pterigióforos. El primer pterigióforo es semitriangular y expandido hacia la parte anterior de la línea media ventral del cuerpo, los restantes son considerablemente largos disminuyendo de tamaño hacia la parte posterior (Fig. 8). 8. ESQUELETO CAUDAL
Caracteres diagnósticosUn Parhipural ventral. Tres Hipurales; dos dorsales y uno ventral en peces de más de 50 mm de LE, notándose claramente las líneas de sutura. Cuatro Hipurales; tres dorsales y uno ventral en peces de menos de 50 mm de LE. Dos epurales en todos los especímenes examinados. Complejo hipural. El complejo hipural de Atherinomorus stipes está constituído por los siguientes elementos (Fig. 9). a) Un centrum preural o Urostilo reducido, su extremo posterior obtuso descansa sobre la base de los hipurales dos y tres fusionados. b) Un parhipural, cuya base engrosada se articula a la porción ventral del centrum preural; en la región anterior presenta la hipurapófisis la cual cubre la región basal del primer hipural. El parhipural corresponde al primer hipural de Hollister (1931). c) Un uroneural, el cual descansa sobre el extremo posterior del centrum preural y el cuarto hipural; el uroneural se fusiona parcialmente a estos huesos en ejemplares adultos. d) Hay cuatro hipurales, uno en la parte inferior del esqueleto caudal y tres en la superior. En ejemplares de 28 mm de LE, están separados para luego fusionarse en ejemplares adultos y formar dos placas, una ventral y una dorsal, pero las suturas son incompletas. El hipural uno, se nota que una vez estuvo separado, por presentar un orificio en su parte basal. e) Dos epurales. Existen dos epurales en todos los especímenes examinados, el anterior es más largo. No se observaron centrum urales libres en ejemplares juveniles. El último centrum completo o centrum preural II carece de espinas neurales presentando sólo la porción basal del arco neural ensanchada. El centrum preural III presenta la espina neural bien desarrollada. Todos los hipurales dependen de un solo centrum como en los Perciformes. 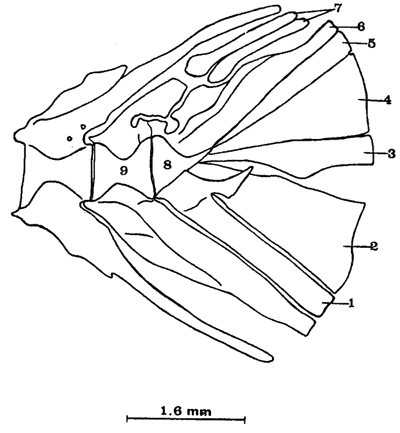 Fig. 9. Esqueleto caudal de Atherinomorus stipes. 1. Parhipural. 2-5. Hipurales. 6. Urodermal. 7. Epurales. 8. Centrum preural I. 9. Centrum preural II. OSTEOLOGÍA DE COLEOTROPIS BLACKBURNI SCHULTZ
1. NEUROCRÁNEO
A. Región occipitalEl proceso anterior del supraoccipital se encuentra expandido lateralmente y la cresta se proyecta hacia atrás a nivel de la primera espina neural. Los cóndilos de los occipitales laterales están más desarrollados. La porción anterior del basioccipital es más bien recta ventralmente (Fig. 10). B. Región etmoidal.El borde posterior del etmoides se encuentra bordeado por los frontales. El prevómer presenta dos extensiones laminares a ambos lados de su porción anterior; carece de dientes. El nasal es de forma semi cuadrada y de menor tamaño, su borde posterior se relaciona con el frontal pero no se encuentra fuertemente unido a éste, presenta un canal en su borde anterior interno (Fig. 10). C. Región temporalSuperficie dorsal del frontal recta, con canales a ambos lados de la porción media de cada hueso. El pleurosfenoides se encuentra articulado por su borde interno con el basisfenoides. El basisfenoides se encuentra articulado por sus bordes laterales externos al pleurosfenoides y proótico; presenta su borde posterior libre, que dando una abertura entre este hueso y los proóticos. Los procesos posteriores del parasfenoides se extienden hasta casi la base del cóndilo del basioccipital. Los parietales de mayor tamaño y extendidos (Fig. 11). 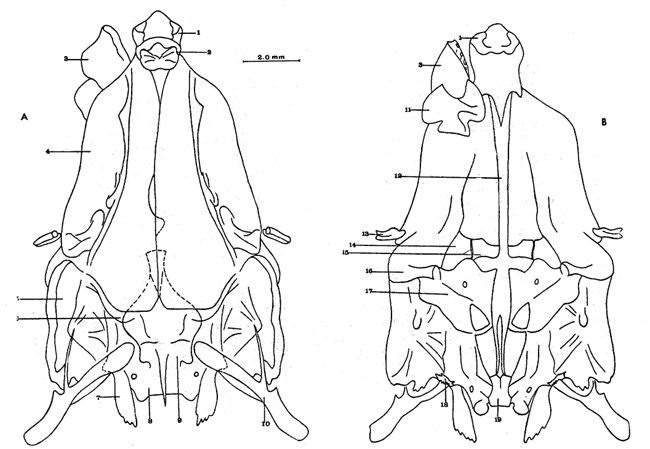 Fig. 10. Neurocráneo de Coleotropis blackburni. A. Vista dorsal. B. Vista ventral. 1. Prevómer. 2. Etmoides. 3. Nasal. 4. Frontal. 5. Pterático. 6. Parietal. 7. Epiótico. 8. Occipital lateral. 9. Supraoccipital. 10. Postemporal. 11. Etmoides lateral. 12. Parasfenoides. 13. Dermosfenótico. 14. Pleurosfenoides. 15. Basisfenoides. 16. Esfenótico. 17. Proótico. 18. Intercalar. 19. Basioccipital. D. Región óticaLa porción o estructura laminar doble del pterótico, se extiende hacia la parte posterior del cráneo. Epiótico con un proceso horizontal, delgado, extendido hacia la parte posterior del cráneo. El proceso lateral del esfenótico (proceso postorbitario) presenta una extensión laminar horizontal, cuya superficie dorsal anterior está cubierta por la región posterolateral externa del frontal. El intercalar está situado sobre el borde externo del occipital lateral y cubre parte del pterótico (Fig. 10). E. Serie circumorbitalLa serie circumorbital está formada por tres huesos, en lugar de cuatro, el segundo de la serie es en forma de escama laminar, alargada con un conducto en su superficie dorsal situado sobre la superficie externa posterior del maxilar; el dermosfenótico es el último de la serie, se articula al esfenótico por su extremo superior (Fig. 11). F. Serie opercularEl opérculo se caracteriza por presentar en la región anterosuperior de la superficie externa, un proceso puntiagudo dirigido hacia el hiomandibular. El interopérculo presenta una escotadura en su borde dorsal externo (Fig. 11). 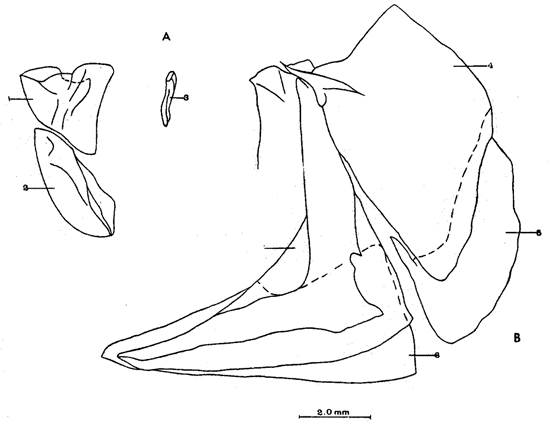 Fig. 11. A. Serie orbital de Coleotropis blackburni. B. Serie opercular de C. blackburni. 1. Lagrimal. 2. Primer suborbital. 3. Segundo suborbital (dermosfenótico). 4. Opérculo. S. Suboperculo.6. Interopérculo. 7. Preopérculo. 2. BRANQUIOCRÁNEO
A. Región oromandibularEl autopalatino carece de la porción adelgazada anterior presente en Atherinomorus stipes; su porción posterior se adelgaza hacia el extremo posterior, articulándose al cuadrado, el borde superior del hueso se encuentra articulado en toda su extensión al endopterigoides. El ectopterigoides está ausente. El endopterigoides es de forma subrectangular, laminar, con un proceso puntiagudo en su ángulo superior derecho, el cual descansa sobre una pequeña extensión laminar, con un proceso puntiagudo en su ángulo superior derecho, el cual descansa sobre una pequeña extensión laminar dorsal del autopalatino; el borde externo del hueso se curva hacia abajo para artícularse al cuadrado, al simpléctico y al metapterigoides; el borde interno del hueso se articula al parasfenoides; carece de dientes en su borde ventral. El metapterigoides es de forma triangular, con su vértice posterior articulado al proceso lateral del hiomandibular. Los procesos ascendentes del premaxilar en forma de placas triangulares cortas, de base ancha, en su borde ventral cóncavo presenta dos hileras de dientes alargados y puntiagudos (en los dos tercios anterior del hueso); el extremo posterior del hueso es ensanchado y su borde redondeado, y no sobrepasa el extremo posterior del dentario; los premaxilares no se unen en su porción media. El proceso interno de la cabeza del maxilar más alargado, y el pliegue externo menos pronunciado, el extremo posterior del hueso sobrepasa el extremo del premaxilar. Los procesos coronoides del dentario elevados y ensanchados con su borde cóncavo. El articular tiene forma subtriangular con su borde anterosuperior bordeando parte del proceso coronoides del dentario (Fig. 12). 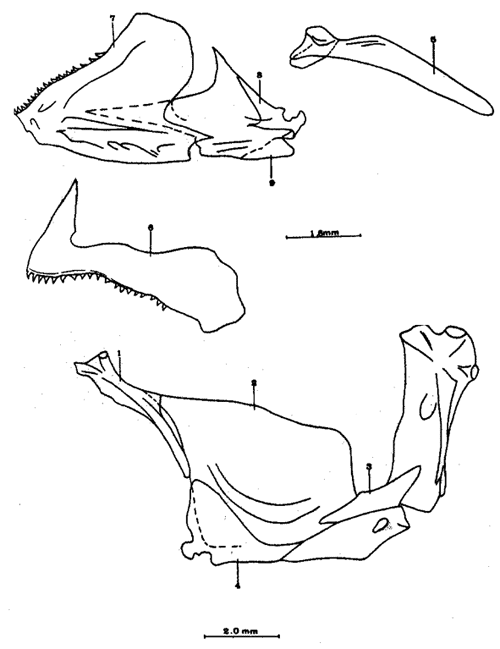 Fig. 12. Branquiocranco de Coleotropis blackburni. Región oromandibular. 1. Autopalatino. 2. Endopterigoides. 3. Metapterigoides. 4. Cuadrado. 5. Maxilar. 6. Premaxilar. 7. Dentario. 8. Articular. 9. Angular. B. Región hioideaHiomandibular con el tallo ensanchado y aplanado, el foramen hiomandibular mucho más desarrollado; la proyección lateral externa está cubierta en su mayor parte por el metapterigoides. El simpléctico es alargado con su porción posterior ensanchada, presenta un foramen en su parte media superior; su porción anterior adelgazada presenta un surco en su superficie externa; el borde externo inferior del hueso está cubierto por el cuadrado anteriormente y por el preopérculo posteriormente. El conjunto epihial, ceratohial e hipohiales es menos alargado y más ensanchado que en Atherinomorus stipes. El Parahioides presenta sólo en su extremo posterior, en el borde dorsal y ventral, pequeñas láminas triangulares dirigidas perpendicularmente, las cuales se unen por ligamentos a los cleitra de la cintura escapular (Fig. 13). C. Aparato branquialEl basihial prolongado muy hacia adelante de los hipohiales; carece de placa dentaria en su superficie dorsal. El primer basibranquial es de tamaño reducido y carece de proyección ventral, no presenta placa dérmica dentada. No existen placas dentarias basibranquiales en Coleotropis blackburni. Los hipobranquiales son de forma irregular y no disminuyen significativamente de tamaño en los arcos posteriores. El cuarto par de ceratobranquiales presentan en su borde dorsal anterior una placa dérmica dentada fusionada al elemento endocondral; las placas de ambos arcos se relacionan en la línea media por su borde interno anterior y cubren la porción anterior del quinto par de ceratobranquiales. El primer par de infrafaringobranquia les está ausente; el tercer y cuarto par están fusionados, formando una placa con su extremo anterior adelgazada y expandida posteriormente, la región dorsal anterior de la placa está cubierta por la región anterior del segundo infrafaringobranquial. La primera placa faríngea superior es de forma triangular y cubre sólo el extremo anterior del segundo infrafaringobranquial; la segunda placa suprafaringeal se fusiona a los infrafaringobranquiales tres y cuatro y la tercera se relaciona al borde posterior de la segunda placa (Fig. 14). Las branquiespinas de la hilera interna del primer arco y las dos hileras del tercer y cuarto arco se presentan como pequeños parches individuales con proyecciones como dedos cubiertos de pequeños dientes en su extremo distal. 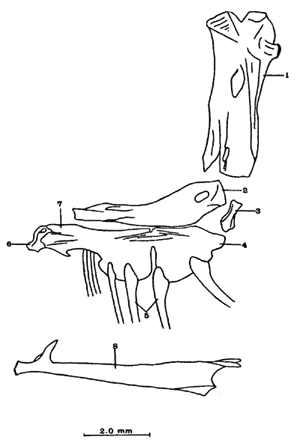 Fig. 13. Branquiocraneo de Coleotropis blackburni. Región hioidea. 1.Hiomandibular. 2.Simplectico. 3.Interhial. 4.Epihial. 5.Radios branquiostegos. 6.Hipohial superior. 7.Ceratohial. 8. Parahioides. 3. CINTURA ESCAPULAREl postemporal presenta en su porción anterior tres ramas en lugar de las dos que presenta en Atherinomorus stipes, la tercera rama es adelgazada y dirigida hacia la porción superior externa del cráneo, situándose sobre la estructura laminar doble del pterótico. El supracleitrum es de tamaño pequeño, de forma subromboidal con su extremo inferior alargado, presenta en su borde interno medio una superficie de articulación a una pequeña protuberancia de la parte superoanterior de la superficie externa del cleitrum; la superficie externa superior del hueso está cubierta por el extremo posterior del postemporal. El cleitrum está formado por dos láminas las cuales se unen por su borde anterior y superior. Del margen posterior del supracleitrum se extienden dos delgados postcleitras, el segundo de los cuales tiene forma de Y con el proceso anterior atenuado, y articulado al superior por el extremo del proceso posterior. La escápula tiene forma triangular con una abertura en su porción media. El coracoides tiene forma triangular con su borde externo curvado, presenta en su superficie interna media un proceso puntiagudo, laminar, dirigido hacia la región anterior interna, el cual se articula con el cleitrum; en el borde posterosuperior del hueso se articulan los dos radiales pectorales posteriores. La cintura escapular, presenta sólo una hilera de radiales pectorales (Fig. 15). 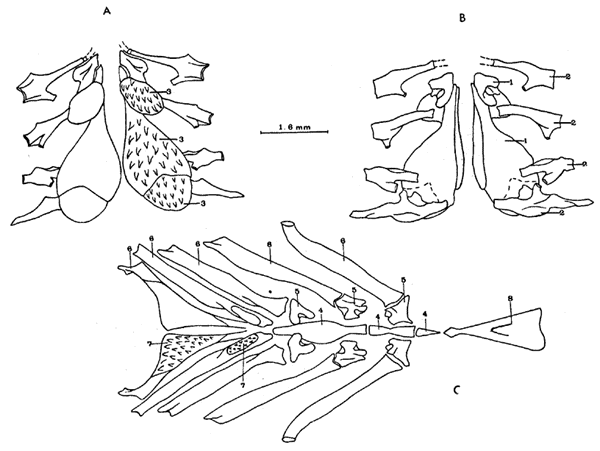 Fig. 14. Branquiocráneo de Coleotropis blackburni. Parte superior de la región branquial. A. Vista ventral. B. Vista dorsal. C. Parte inferior de la región branquial, vista dorsal. 1. Infrafaringobranquiales. 2. Epibranquiales. 3. Placas faríngeas superiores. 4. Basibranquiales. 5. Hipobranquiales. 6. Ceratobranquiales. 7. Placas faríngeas inferiores. 8. Basihial. 4. CINTURA PÉLVICALos basipterigios tienen forma triangular, presentan en su porción postero lateral una extensión laminar de forma triangular dirigida hacia la pared del cuerpo. En la porción interna del borde posterior, presenta un largo proceso laminar en forma de dedo (Fig. 15). 5. COLUMNA VERTEBRALLos especímenes examinados presentan de cuarenta a cuarenta y una vértebras incluyendo el urostilo, de las cuales de catorce a quince son precaudales y de veinticinco a veintiseis son caudales. Las parapófisis de las vértebras precaudales anteriores están expandidas horizontalmente. Existen doce costillas pleurales. Las espinas neurales de las primeras vértebras forman crestas neurales semitriangulares (Fig. 16). 6. ALETAS DORSALESLa primera dorsal presenta de dos a cuatro espinas, generalmente tres. El primer pterigióforo es alargado, laminar, situado en sentido anteroposterior y sostiene las dos primeras espinas (Fig. 17). Le segunda dorsal presenta una espina y ocho ó nueve radios, soportados por pterigióforos de forma alargada y puntiagudos en su parte inferior. 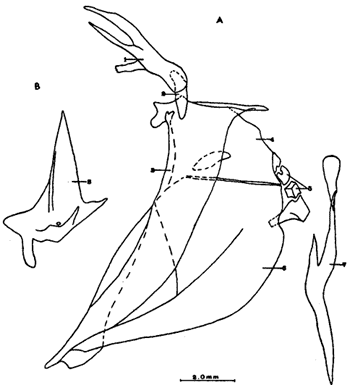 Fig. 15. A. Cintura escapular de Coleotropis blackburni. B. Cintura pelvica de Coleotropis blackburni. 1. Postemporal. 2. Supracleitrum. 3. Cleitrum. 4. Escápula. 5. Radiales pectorales. 6. Coracoides. 7. Postcleitrum. 8. Basipterigio. 7. ALETA ANALEn la aleta anal existe una espina y veintitrés ó veinticuatro radios, soportados por pterigióforos. Todos los pterigióforos tienen forma alargada, dirigidos hacia la porción media del cuerpo, y disminuyen de tamaño hacia la parte posterior (Fig. 17). 8.ESQUELETO ANALEl esqueleto caudal sigue la misma modalidad que Atherinomorus stipes. Se observó un mayor desarrollo en proporción al cuerpo. Los epurales son más alargados (Fig. 18). DISCUSIÓNEs evidente que Atherinomorus difiere significativamente de Coleotropis en muchos caracteres osteológicos sobresalientes, los cuales se señalan como caracteres diagnósticos para cada especie. De esta forma se fundamenta la ubicación de Schultz (1948) de las dos especies en subfamilias distintas. Atherinomorus difiere en algunos caracteres de los señalados por Rosen (1964) y por Greenwood et al. (1966) para definir el orden Atheriniforme y para el superorden Atherinomorfa, respectivamente. Starks (1899) hace un estudio del esqueleto de varios representantes de la familia Atherinidae, Mugilidae y Sphyraenidae, los cuales estaban agrupados bajo el suborden Percesoces; señala como característica común, los epióticos desarrollados en largos procesos, delgados, los cuales se dividen en filamentos más o menos como cerdas; señala a la vez que hay pocos caracteres internos de los cuales difieren estas familias, como un grupo de otros Acanthopterigii. Sin embargo, Atherinomorus carece de proceso epiótico; este mismo carácter ha sido observado por Gosline (1962) para el género Pranesus, señalando que contrariamente a los Polynemidae y Sphyraenidae, los Atherinidae representados por Pranesus el cual está incluído en la Subfamilia Taenomembrasinae al igual que Atherinomorus, (aparentemente) diverge del tronco basal de los Polynemidae, Sphyraenidae y Mugilidae. 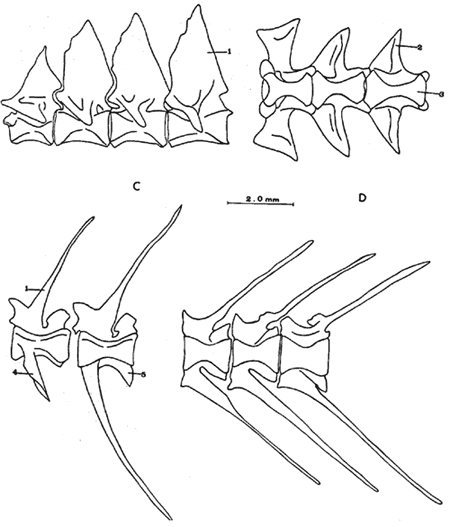 Fig. 16. Columna vertebral de Coleotropis blackburni. A. Primeras vertebras precaudales, vista lateral. B. Primeras vertebras precaudales, vista ventral. C. Primeras vertebras caudales, vista lateral. D. ultimas vértebras caudales, vista lateral. 1. Espina neural. 2. Parapéfisis. 3. Centrum vertebral. 4. Espina hemal. 5. Proceso posteroventral. NEUROCRÁNEOEl neurocráneo en Atherinomorus stipes es mucho más alto y ancho que en Coleotropis blackburni. La superficie dorsal es amplia con una concavidad a ambos lados en la región que forma la órbita; en C. blackburni, la superficie dorsal es lisa. Existe una mayor osificación del cráneo en A. stipes que en C. blackburni.  Fig. 17. A. Primera aleta dorsal de Coleotropis blackburni. B. Parte anterior de la segunda aleta dorsal. C. Parte anterior de la aleta anal. 1. Espinas. 2. Pterigióforos. 3. Suprancurales. 4. Radios. Los frontales forman la mayor parte de la superficie dorsal del cráneo cubriendo parte de los parietales y supraoccipital, los cuales se encuentran situados en la parte posterior; en la parte anterior lateral, se sitúan los expandidos nasales, los cuales se articulan fuertemente al borde anterior de los frontales, y en la parte media anterior se encuentra el etmoides. En C. blackburni los nasales son de menor tamaño y no están articulados fuertemente a los frontales. El epiótico de A. stipes, carece del proceso posterior, el cual está presente en C. blackburni. 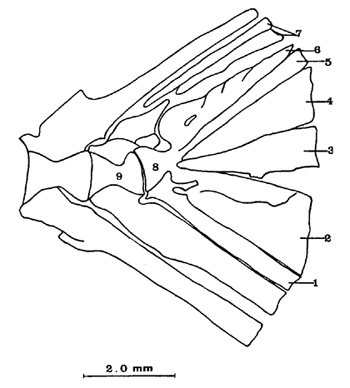 Fig. 18. Esqueleto caudal de Coleotropis blarkburni. 1. Parhipural. 2-5. Hipurales. 6. Urodermal. 7. Epurales. 8. Centrumpleural 1. 9. Centrum pleural II. Las órbitas en Atherinomorus stipes están ampliamente desarrolladas para alojar los ojos de gran tamaño que presenta esta especie. El parasfenoides es ligeramente cóncavo en su parte anterior, mientras que en Coleotropis blackburni se dirige abruptamente hacia la parte anterior. El prevómer presenta una hilera de pequeños dientes en Atherinomorus stipes. Serie circumorbital. En los peces primitivos el ojo está completamente rodeado por una serie de pequeños huesos que puede ser referido como serie circumorbital. La serie circumorbital se divide en una serie supraorbital, que consiste de los huesos situados dorsal a la órbita y según Smith y Baley (1962), no presentan canal sensorial; y una serie de huesos los cuales son atravesados por la rama infraorbital del sistema de la línea lateral. Entre los teleósteos es generalmente seis el número de huesos en la serie suborbital, aunque en algunos peces los huesos tienden a la reducción y en otros se han dividido incrementando el número de elementos. La tendencia de los supraorbitales es hacia la reducción, pérdida o fusión, volviéndose rígida la serie dorsal de la órbita. La concha subocular es definida por Smith y Baley (1962) como una lámina la cual se extiende hacia adentro, desde los huesos suborbitales en ciertos peces, y forma parte de la pared de la órbita. Su presencia o ausencia ha sido usada como carácter taxonómico a nivel de familia y puede servir como un indicador de la historia filética. Así podría pensarse que la pérdida de infraorbitales en Atherinidae y la persistencia junto con la lámina subocular en Mugilidae y Sphyraenidae, representa diferentes tendencias de significado filogenético. La pérdida de huesos infraorbitales puede ser un modelo secundario de Atherinidae o podría confirmarse la separación en Atheriniformes de Greenwood et. al. (1966). En los caracteres señalados para el neurocráneo de Atherinidos, Rosen (1964) menciona que los huesos infraorbitales están representados solamente por el primero (lagrimal) y el último (dermosfenótico), excepto en Melanotaenidae, en los cuales existe un tercer elemento adicional unido firmemente a la superficie ventral del lagrimal. En Coleotropis existe el tercer elemento señalado por Rosen (1964), pero no firmemente unido al lagrimal; en Atherinomorus existe además un cuarto elemento que corresponde al tercero de la serie suborbital. Greenwood et al. (1966) al definir el superorden Atherinomorfa, señala como carácter lo mencionado anteriormente por Rosen (1964). Radios branquiostegos. El número de radios branquiostegos en las dos especies estudiadas, confirman lo. señalado por Hubb (1919)... "el número de radios branquiostegos de los Acanthopterigii y grupos relacionados, es generalmente cuatro más dos, o cuatro más tres"... Gosline (1967), señala que la reducción en el número de radios branquiostegos está más bién relacionado con el acortamiento de la mandíbula inferior en asociación con el desplazamiento hacia adelante de la articulación cuadrado-mandibular. Tal es el caso para las dos especies estudiadas. BRANQUIOCRÁNEOArcos branquiales. Casi toda la superficie oral de los arcos branquiales de Atherinomorus stipes están cubiertos por placas dérmicas dentadas, lo cual representaría un carácter primitivo de este género. Algunas placas están fusionadas a su soporte endocondral, y otras no. El número de placas dérmicas dentadas es reducido en Coleotropis blackburni. Placas dentarias de forma y tamaño regular cubren los basibranquiales de Atherinomorus stipes. En Coleotropis blackburni las placas dentarias están ausentes en el basihial y basibranquiales y presenta un par de placas en el cuarto par de hipobranquiales. Greenwood et al. (1966), al definir el superorden Atherinomorpha señala dos placas faríngeas superiores como característica para el esqueleto branquial. Sin embargo, existen tres en las especies examinadas; aunque las dos posteriores están articuladas entre sí, es evidente su individualidad. También señala el citado autor que el primer par de infrafaringobranquiales está presente sólo pocas veces, pero en forma rudimentaria. Según Rosen (1964), el primer par de faringobranquiales se pierden en los Atherinidae y sólo se presenta como una diminuta osificación en Melanotaenia. Sin embargo, en Atherinomorus stipes se encuentra bien desarrollado y une el aparato branquial al cráneo. El tercer y cuarto infrafaringobranquiales se fusionan y forman una sola placa ósea, cubiertos por el tercer par de suprafaringeales. Los infrafaringeales forman dos placas triangulares situadas encima del quinto par de ceratobranquiales. En Coleotropis, el primer par de infrafaringobranquiales está ausente. CINTURA ESCAPULAREn Atherinomorus stipes existen dos hileras de radiales pectorales, en tanto que, en Coleotropis blackburni existe solo una. No se ha encontrado en la literatura ninguna indicación de la presencia de una segunda hilera de radiales pectorales en la cintura escapular de estos peces. Sin embargo, Berg (1940) señala su ausencia o presencia como un carácter taxonómico en el orden Clupeomorfa. Esta característica podría constituir un carácter taxonómico más dentro de la familia Atherinidae, y las relaciones evolutivas entre los grupos si se interpreta la pérdida como un carácter más especializado. ESQUELETO CAUDALEl esqueleto caudal de Atherinomorus stipes es del tipo estegural según Monod (1968), el cual es característico de los teleósteos superiores en la clasificación de Gosline (1971); en donde el par de uroneurales anteriores se fusionan con el elemento del arco neural del centrum preural I. En Atherinomorus stipes el elemento del arco neural está reducido y no se observa centrum ural durante el desarrollo. El resultado de la fusión da origen a una cresta convexa sobre el uroneural y toda esta estructura ha sido llamada estegural por Monod (1968). Esta estructura se fusiona con los elementos del centrum del esqueleto caudal. En Coleotropis blackburni el esqueleto caudal sigue la misma modalidad de Atherinomorus stipes. ConclusionesCARACTERES DIAGNÓSTICOS DE ATHERINOMORUS STIPES1. Cráneo ensanchado lateralmente. 2. Epiótico sin cresta. 3. Prevomer con pequeños dientes en una hilera en su borde ventral anterior. 4. Un lagrimal, dos suborbitarios y el dermosfenótico en la serie suborbital. 5. Borde anterior del hocico recto o casi recto. 6. Pliegues de los ángulos de la boca fusionados en el extremo posterior de la mandíbula. 7. Borde del premaxilar recto o ligeramente cóncavo, extremo posterior no extendido o dilatado, más bien aguzado y no alcanza el extremo posterior del dentario. Proceso ascendente del premaxilar ancho y truncado o recto en su borde superior, cortos, no alcanzan el espacio interorbital, unidos en la línea media. Dientes en bandas viliformes reflejados sobre la cara externa. Borde lateral del premaxilar recto. 8. Maxilar y premaxilar sobrepasan el proceso coronoides del mandibular. 9. Proceso coronoides del dentario corto, no alcanza el extremo posterior del hueso y su borde posterior recto. Porción anterior de la mandíbula con parches de dientes en su borde externo superior. 10. Endopterigoides dentado. 11. Primer par de infrafaringobranquiales presentes. 12. Dos hileras de radiales pectorales. 13. Vértebras de 17 a 20 precaudales y de 18 a19 caudales. 14. Espina hemal no expandida. 15. Origen de la primera dorsal muy por delante del origen de la anal, segunda dorsal a nivel del quinto radio de la anal. 16. Cavidad del cuerpo se extiende justo enfrente del origen de la anal. 17. Ano situado en el abdomen a nivel del extremo posterior de los radios de las aletas pélvicas, muy por delante de la base de la aleta anal. CARACTERES DIAGNÓSTICOS DE COLEOTROPIS BLACKBURNI1. Cráneo no ensanchado lateralmente. 2. Epiótico con cresta. 3. Premover sin dientes. 4. Un lagrimal, un suborbitario y el dermosfenótico en la serie suborbital. 5. Borde anterior del hocico un poco cóncavo. 6. Pliegues de los ángulos de la boca fusionados cerca de un tercio de la parte distal del premaxilar y dentario. 7. Borde del premaxilar cóncavo. Su extremo posterior dilatado y curvado en su borde extremo. Proceso ascendente de base ancha, triangular, corto, no alcanza el espacio interorbitario; no se encuentran unidos en la línea media. Dientes alargados en doble hilera. 8. Maxilar y premaxilar no sobrepasan el extremo posterior del proceso coronoides. 9. Proceso coronoides del dentario expandido y curvado en su extremo. 10. Endopterigoides sin dientes. 11. Opérculo con un proceso en la región superoanterior de la superficie externa, dirigido hacia al hiomandibular. 12. Glosohial prolongado hacia adelante a partir de los hipohiales. 13. Primer par de infrafaringobranquiales ausentes. 14. Una hilera de radiales pectorales. 15. Vértebras de 14 a 15 precaudales y de 25 a 26 caudales. 16. Espinas hemales no expandidas. 17. Origen de la primera dorsal por detrás del origen de la anal a nivel del cuarto radio. 18. Cavidad del cuerpo extendida justo enfrente del origen de la anal. 19. Ano situado entre el extremo posterior de los radios pélvicos y la base de la aleta anal. 20. Pedúnculo caudal recto y ensanchado. 21. Región ventral del cuerpo lateralmente aplanada. LITERATURABERG, L. S., Classifation of Fishes both Recent and Fossil. . Russian and English lithoprint, 1947. Ann Arbor, Michigan Edwards, J. W. 1940 517 CERVIGON, F., Los Peces Marinos de Venezuela. Fundación La Salle Ciencias Naturales, Caracas, Monogr. 1966 1-436 11 1 Los Peces Marinos de Venezuela. Complemento III. Contribuciones Científicas. Univ. Oriente Núcleo Nueva Esparta, Porlamar, Venezuela, 1973 16-19 4 Los Peces Marinos de Venezuela. Complemento IV. Contribuciones Científicas. Univ. Oriente Núcleo Nueva Esparta, Porlamar, Venezuela, 1975 3-45 5 CHAPMAN, W. M., J. Morph., The osteology of the Pacific deepbodied anchovia, Anchoa compressa. 1944 311-329 74 D'VILLERS, C., Traité de Zoologie, Le crané des poissons. In: Grassé, P. - P. 1958 551-687 13 1 EATON, T. H., J. Morph., Skeletal supports of the median fins of fish. 1945 193-212, 5 figs. 76 GOSLINE, W. A., Copeia, The perciform caudal skeleton. 1961a 265-270 3 Smithsonian Miscellaneous Collections, Some osteological feature of modern lower teleostean fishes. 1961b 1-42, 8 figs. 9 diagr 142 3 Pacific Science, Systematic position and relationship of the percesocine fishes. 1962 207-217, 3 figs. 16 Copeia, Teleostean phylogeny. 1962 207-217, 3 figs. 16 Copeia, Reduction in branchiostegal ray number. 1967 237-239, 1 figs 3 Functional Morphology and Classification of Teleostean Fishes. The University of Hawaii press, 1971 208, 29 figs. GREENW00D, P. H., R. S. MILES Y C. PATTERSON, Interrelationships of Fishes. Zoological Journal of the Linnean Society, Academic Press, London. 1973 (1) 536 53 GREENWOOD, P. H., E. D. ROSEN, S. H. WEITZMAN y G. S. MYERS, Bull. Am. Mus. Nat. Hist., . Phyletic studies of teleostean fishes, with a provisional classification of living form 1966 350-361, 9 figs. 131 4 HOLLISTER, G., Zoologica, Caudal skeleton of Bermuda shallow water fish. II. Order Percomorphi, Suborder Percesoces: Atherinidae, Mugilidae, Sphyraenidae. 1936 265-279, 14 figs. 22 JORDAN, D. S. y C. L. HUBBS, Leland Stanford Jr. Univ. Publ. Univ. Ser., Studies in Ichthyology A monograpliic review of the family of Atherinidae or Silverside. 1919 1-87, 12 láms., 42 figs. MONOD, T., Mem. Inst. Fr. Afr. Noire, Le complexe urophore des poisons téléostéens. 1968 1-705 81 NELSON, G. J., Bull. Am. Mus. Nat. Hist., Gill arches and the phylogeny of físhes, with notes on the classification of vertebrates. 1969 479-552, 92 láms. 141 4 NYBELIN, O., Interrelationships of Fishes. Comments on the caudal skeleton of actinopterygians. In: Greenwood et al. Zoological journal of the Linnean Society, 1973 369-372, 1 fig., 1 lám. 53 1 ROSEN, D. E., Bull. Am. Mus. Nat. Hist., Bull. Am. Mus. Nat. Hist., The relationships and taxonomic position of the halfbeaks, killifishes, silverside, and their relatives. 1964 217-268 127 5 Interrelationships of Fishes. Interrelationships of higher euteleosteans fishes. In: Greenwood et al. Zoological journal of the Linnean Society, 1973 397-519, 129 figs. 53 1 SCHULTZ, L. P., Proc. U. S. Nat. Mus., A revision of six subfamilies of atherine fishes, with descriptions of new genera and species. 1948 1-48 98 SMITH, C. L. y R. M. BALEY, J. Morph., The subocular shelf of fishes. 1962 1-18, 3 láms. 110 STARKS, E. C., Proc. U. S. Nat. Mus., The osteological characters of the fishes of the suborder Percesoces. 1899 1-10, 3 láms. 22 TAYLOR, W. R., TAYLOR, W. R., Proc. U. S. Nat. Mus., An enzyme method of clearing and staining small vertebrates. 1967 1-7 122 3596
|