|
DETERMINACIÓN DE ÁCIDOS GRASOS EN UNA NUEVA
ESPECIE DE ALGA DEL GÉNERO NOSTOC
Trabajo recibido el 9 de junio
de 1976 y aceptado para su publicación el 15 de octubre de
1976.
LUCÍA IRENE GONZÁLEZ VILLASEÑOR
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología.
Se determinó la composición de
ácidos grasos por cromatografía de gases en la alga azul-verde
Nostoc sp. La muestra fue aislada de las raíces de
Macrozamia (Cycadaceae) y cultivada en condiciones
experimentales de laboratorio.
Se encontraron un total de seis ácidos grasos:
mirístico, C14-0; palmítico, C16-0; polmitoléico, C16:1;
esteárico, C18:0; oléico, C18:1 y linoléico,
C18:2.
Se observó una alta proporción de
ácido palmítico, pero no se detectó el ácido
linolénico, 18:3, el cual se ha reportado en diferentes proporciones en
la mayoría de las cianoficeas, incluyendo otras especies del
género Nostoc.
The fatty acid composition of a blue-green algae,
Nostoc sp., was investigated by gas liquid chromatography.
The algae was isolated from the roots of Macrozamia
(Cycadaceae) and cultivated in experimental laboratory conditions.
Six fatty acids were found: miristic, C14:0;
palmitic, C16:0; palmitoleic, C16-1; estearic, C18:0; oleic, C18:1 and
linoleic, C18:2.
A major feature in the distribution of the fatty
acids in the studied Nostoc sp., was the dominance of
palmitic acid and the lack of linolenic acid, wich is a fatty acid
characteristic of most Nostoc species.
Las diferencias en la distribución de ácidos grasos entre las distintas especies de algas indica tanto variaciones taxonómicas (Korn, 1964), como variaciones de los niveles de nitrógeno; de ahí que sean dependientes de las condiciones del medio ambiente (Miller, 1962), como de las condiciones experimentales de crecimiento a las que son sometidas cuando se trata de cultivos puros (Holton et al., 1964 y Van Baalen, 1967). Una de las divisiones menos estudiadas son las cianofíceas y algunos datos sobre la composición de ácidos grasos en, ellas han sido, reportados por diferentes investigadores Erwinet al. (1969), Holton et al. (1964 y 1968), Levin et al.(1964), Nichols et al. (1968), Oró et al. (1967), Parker (1965 y 1967), Watanabe (1960) y Watanabe et al. (1968). En este estudio se da a conocer la composición de los ácidos grasos presentes en Nostoc sp., una nueva especie aún no identificada del género Nostoc que se desarrolla sobre las raíces coraloides de una cicadácea del género Macrozamia que crece en las costas de Texas en el Golfo de México (Hoare et al., 1971). MATERIAL Y MÉTODOEsta cianofícea presenta características que la diferencian tanto taxonómicamente como también por la composición en ácidos grasos, con respecto a otras especies del género Nostoc ya conocidas. Como aún no se ha definido taxonómicamente la nueva especie, se presenta en este trabajo como Nostoc sp (Hoare et al., 1971). El alga fue aislada de las raíces de la cicadácea coraloide del género Macrozamia y desarrollada en un medio de cultivo Cg-10 en glucosa al 0.5% (Van Baalen, 1967), ventilado con corriente continua de aire mezclado con CO2 al 1% durante 24 hrs., en la obscuridad, a 39°C. La muestra fue decantada para desechar el medio de cultivo y tratada con metanol durante 30 minutos sometida a agitación y vibración ultrasónica. El metanol fue recuperado por filtración y la muestra secada a 50°C y pesada. Al extracto de metanol se agregó cloroformo y agua; se separó la fase de cloroformo y se lavó con HCL 0.1 N para remover sales inorgánicas, posteriormente se evaporó bajo una corriente de aire seco. Después se saponificó con hidróxido de potasio en metanol y las impurezas fueron removidas por extracción de la solución alcalina con hexano, esta solución fue luego acidificada y los ácidos grasos extraídos con hexano. Los ésteres metílicos fueron preparados con BF3 en metanol al 10% y extraídos nuevamente con hexano. La identificación y determinación de la concentración relativa de los ácidos grasos como ésteres metílicos se llevó a cabo por cromatografía de gases, con un cromatógrafo Hewlett Packard modelo 5710 A con detector de llama ionizada usando hidrógeno y aire, se utilizaron columnas de aluminio de 1.5 m de largo y 3.2 mm de diámetro. El material de las columnas fue FFAP al 5% (Varian Aerograph) sobre Gas Chrom Q de malla 60/80 (Applied Science Laboratories, Inc.) . Se utilizó como gas transportador helio con un flujo de 25 ml/min. La temperatura fue programada de 100 a 280°C con incrementos de 4°C por minuto. El trabajo se llevó a cabo en el Instituto de Ciencias Marinas de Port Aransas, Texas. RESULTADOSCon respecto a la composición de los ácidos grasos determinados en otras cianofíceas nostocales, la especie que se ha estudiado presenta ciertas variaciones cuali y cuantitativas. Un total de seis ácidos grasos fueron identificados: mirístico, palmítico, palmitoléico, esteárico, oléico y linoléico. En la tabla 1 se presenta una comparación de los ácidos grasos de Nostoc sp. y algunos miembros de la familia Nostocaceae. El ácido palmítico 16:0 resultó ser el más abundante, correspondiendo a un 45.9% del total de ácidos grasos, el ácido mirístico, 14:0 se encontró en menor porcentaje y dentro de los ácidos grasos no saturados, el ácido linoléico, 18:2 fue el que se encontró en mayor proporción, lo cual es comparable con los datos de otras especies de Nostoc. El cromatograma de los ésteres metílicos de los estandards y el de los ésteres metílicos aislados de Nostoc sp.se muestra en las figuras 1 y 2. En la figura 3, se muestra a los ésteres metílicos separados en dos grupos, los de cadena simple o saturados con 14, 16 y 18 átomos de carbono y los no saturados con 16 y 18 átomos de carbono pero con 1 y 2 dobles enlaces en su cadena. El ácido linolénico 18:3 no fue detectado en Nostoc sp., sin embargo para otras especies se ha reportado en alta proporción (Watanabe et al., 1968), esto es debido principalmente á las condiciones de cultivo, de Nostoc sp. En experimentos hechos con Chlorogloea fritschii (Holton et al., 1968), se ha demostrado que cuando se somete a diferentes condiciones de crecimiento, ya sea luz u obscuridad, se presenta una variación cualitativa importante en la composición de ácidos grasos; de tal forma que en condiciones de obscuridad hay casi una completa ausencia del ácido linolénico. 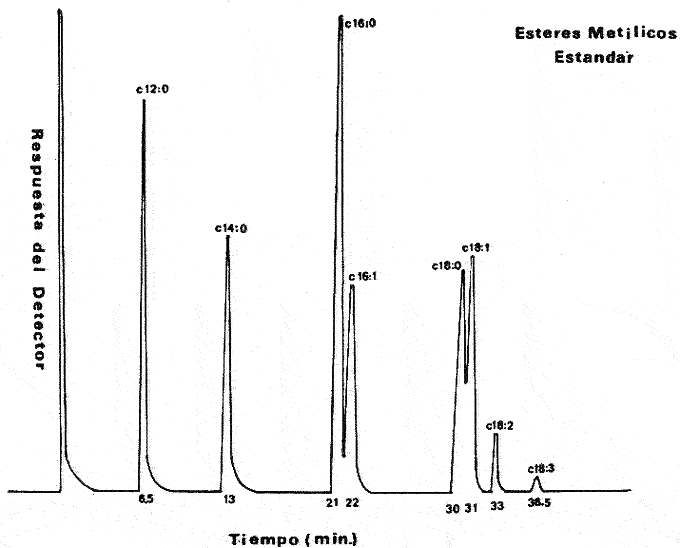 Fig. 1. Cromatograma de los ésteres metálicos de los estándar. 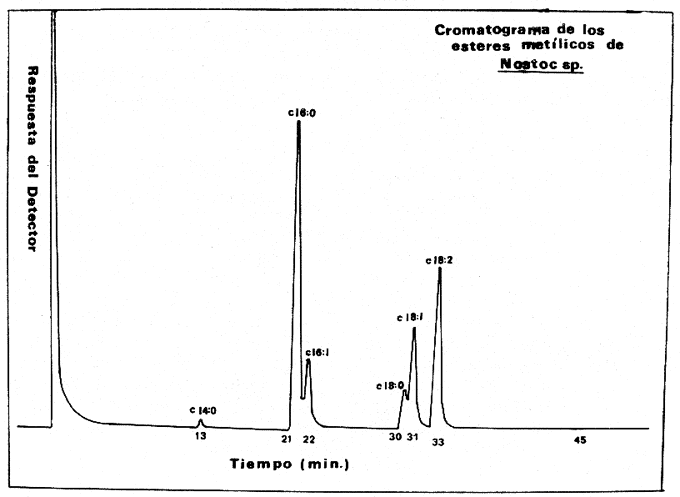 Fig. 2. Cromatograma de los ésteres metálicos aislados de Nostoc sp. 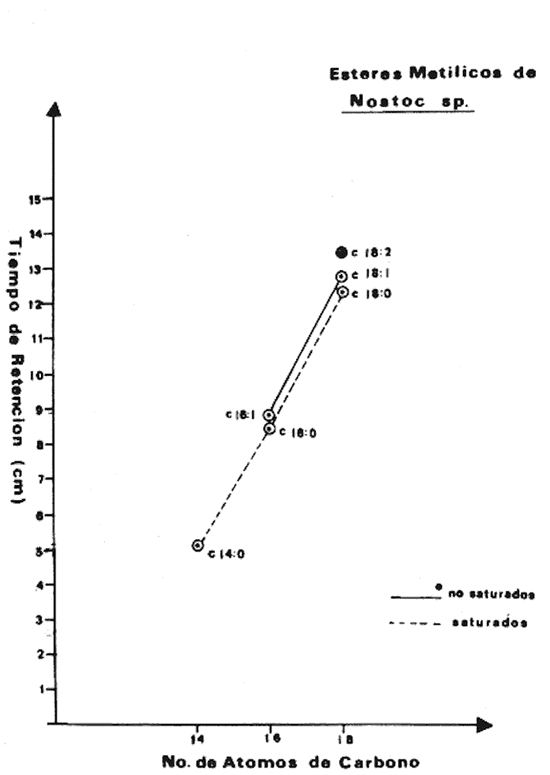 Fig. 3. Cromatograma de los ésteres metálicos separados en dos grupos. DISCUSIÓNLos efectos del medio ambiente, las condiciones de crecimiento y los diferentes requerimientos físico-químicos de las algas influyen en su contenido de ácidos grasos. También la variedad entre ellos puede ser significativa taxonómicamente, sin embargo, es clara la dependencia del vegetal a los factores externos a que se encuentra sometido (Lewin, 1962). Por otro lado, ya ha sido demostrado que cuando los organismos crecen en condiciones de completa obscuridad, desarrollándose heterotróficamente, como es el caso de Chlorogloea fritschii, hay una casi completa ausencia del ácido graso 18:3, lo que sugiere una relación directa entre éste y la actividad fotosintética (Holton et al., 1968); en cambio en condiciones de luz continua, se ha detectado el ácido linolénico el cual se incluye química o físicamente como un requerimiento necesario en la producción de oxígeno durante la fotosíntesis (Holton et al., 1964 y 1968; Watanabe et al., 1968). Por lo tanto puede deducirse que la variedad en la composición de ácidos grasos entre las diferentes especies de Nostoc estudiadas y entre las cianofíceas en general debe atribuirse en parte a las condiciones experimentales de crecimiento y a las condiciones del medio ambiente. También, la presencia de ácidos grasos característicos en ciertos grupos, así como su relación directa con la habilidad bioquímica de sintetizarlos, puede tener algún significado evolutivo en las algas azul-verdes. Por ejemplo, el ácido linolénico no se presenta en bacterias, pero si en algas cianofíceas, a pesar de que morfológicamente se relacionan a las bacterias; sin embargo, en las algas cianofíceas existe ya un aparato fotosintético simple. Esto sugiere que durante los procesos evolutivos, este carácter bioquímico fue transferido en asociación con la maquinaria fotosintética que incluye la producción de oxígeno (Erwin et al., 1964; Holton et al., 1964 y 1968; Kenyon et al., 1960). Por lo tanto, es importante hacer estudios posteriores acerca de estas especulaciones para determinar con claridad el significado entre la diversidad de los patrones de ácidos grasos presentes en las algas cianofíceas. 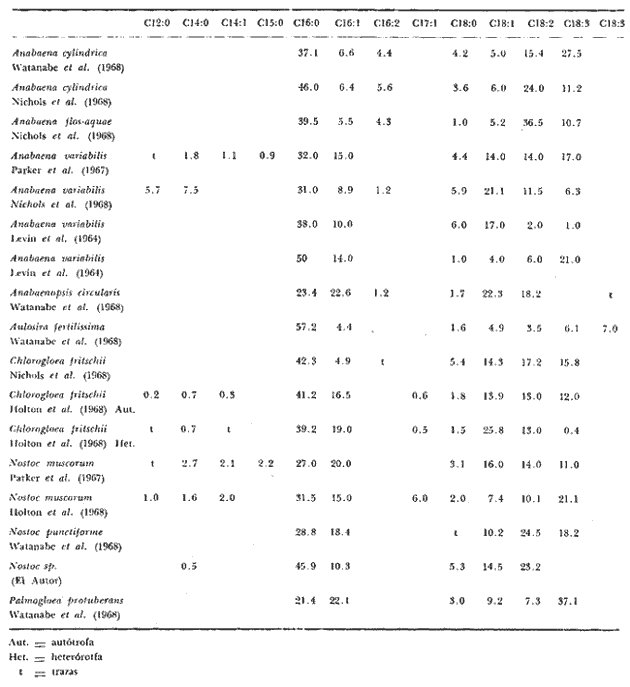 TABLA 1 COMPOSICIÓN DE ÁCIDOS GRASOSOS EN CIANOFICEAS (%), FAMILIA NOSTOCEAE AgradecimientosA Patrick L. Parker, ex Director del Instituto de Ciencias Marinas de Port Arkansas, Texas, por las facilidades prestadas para el uso de instrumentos y equipo en el laboratorio de química, así como por su dirección en la realización de este trabajo. A Chase Van Baalen, del Departamento de Botánica del mismo Instituto, quien donó la muestra de Nostoc para el análisis de ácidos grasos. A Ken Winters, por su ayuda en el manejo del equipo empleado. A Robert Eklund y Paul Carangelo por su colaboración. A Enrique F. Mandelli, por sus valiosos consejos para la elaboración del presente trabajo. Finalmente al Comité Editorial de las Publicaciones del Centro de Ciencias del Mar y Limnología de la UNAM la gentileza de aceptar esta contribución. LITERATURAERWIN, J y K. BLOCH, Science. Biosynthesis of unsaturated fatty acids in microorganisms. 1964 1006-1012. 143 HOARE, D. S., L. O. INGRAM, E. L. THURSTON y R. WALKUP, Arch. Mikrobiol. Dark heterotrophic growth of an endophytic blue-green alga. 1971 310-321. 78 HOLTON, R. W., H. H. BLECKER y T. S. STEVENS Fatty acids in blue-green algae: possible relation to phylogenetic position. 1968 602 KENYON, C. N. y R. Y STANIER Nature Possible evolutionary significance of polyunsaturated fatty acids in blue-green algae. 1970 1164-1165. 227 KORN, E. D. The fatty acids of Euglene gracillis. J. Lipid Res. 352-362 5 LEWIN, R. A., Physiology and biochemistry of algae Academic Press New York 929 LEVIN, E., W. J. LENNAR y K. BLOCH, Biochem. Biophys. Acta Ocurrence and localization of a- linolenic acid containing galactolipids in the photosyntetic apparatus of Anabaena variabilis. 471-474 84 MILLER, J. D. A. Physiology and biochemistry of algae. Physiology and biochemistry of algae. Fats and steroids Lewin, R. A. Academic Press New York 357-370 NICHOLS, B. W. y B. J. B. WOOD, Lipid The ocurrence and biosynthesis of gamma-linolenic acid in a blue-green alga, Spirulina platensis. 1968 46-50 3 ORO, J. T. G. TORNABENE, D. W. NOONER y E. GELPI, Journal of Bacteriology Aliphatic hydrocarbons and fatty acids of some marine and freshwater microorganisms. 1967 1811-1818 93 PARKER, P. L. y R. F. LEO, Science Fatty acids in blue-green algal mat communities. 1965 373-374 148 PARKER, P. L. C. VAN BAALEN y L. MAURER ScienceScience Fatty acids in eleven species of blue-green algae: Geochemical significance. 1967 707-708 155 VAN BAALEN, C. Journal of Phycology Further observations on growth of single cells of coccoid blue-green algae. 1967 154-157 3 3 WATANABE, A., J. Gen. Microbiol. List of algal strains of collection at the Institute of Applied Microbiology, University of Tokyo. 283-292 6 WATANABE, A. y Y. YAMAMOTO, Taxonomy and Biology of Blue-Green Algae. Linolenic acid producing species among the Cyanophyta. In: Desikachary, T. W. University of Madras India 1968
|