|
DIVERSIDAD Y AFINIDAD DEL FITOPLANCTON EN UN CICLO
NICTEMERAL
Trabajo presentado en el II Simposio Latinoamericano de
Oceanografía Biológica, Cumaná, Venezuela, noviembre 24-28
de 1975. Recibido el 28 de septiembre de 1976 y aceptado para su
publicación el 15 de octubre de 1976.
HERMILO SANTOYO, MARTHA SIGNORET
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología, Laboratorio de
Fitoplancton y Productividad. Contribución 67 del Centro de Ciencias del
Mar y Limnología, UNAM
Se analizan en un punto de la Bahía de
Campeche, Golfo de México, las fluctuaciones de la diversidad de
especies y el grado de afinidad de las poblaciones fitoplanctónicas
durante un ciclo nictemeral. La estimación de la diversidad se hace por
medio de la ecuación de Shannon y Weaver y la afinidad por el
índice de Renkonen.
Se identificaron dos capas diferentes
estructuralmente; la superficial, con valores de diversidad y afinidad poco
constantes y bajos a través del tiempo y la del fondo con valores altos
de ambos parámetros y una composición cualitativa
diferente.
Hubo poblaciones persistentes a través del
tiempo, variando únicamente su densidad numérica; sin embargo, se
registraron cambios en la estructuración de la comunidad
señalando un dinamismo marcado como resultado de la influencia de
factores intrínsecos y condiciones ambientales. La estructura de la
comunidad estuvo afectada principalmente por el comportamiento de las
fitoflageladas y dinoflageladas las cuales efectúan migraciones
verticales durante el ciclo nictemeral.
Les variations de la diversité
spécifique et le degré d'affinité des populations
phytoplanctoniques sont analysés en un point de la Baie de Campeche,
Golfe du Méxique, pendant un cycle nycthéméral.
L'estimation de la diversité se fait d'aprés la formule de
Shannon et Weaver et l'affinité d'aprés l'indice de
Renkonen.
Deux couches différentes en structure ont
été identifiées; une superficielle, avec des valeurs de
diversité et d'affinité basses et irréguliéres
à travers le temps et celle du fond, avec des valeurs
élevées pour les deux paramètres et une composition
qualitative, différente.
Durant le cycle nycthéméral certaines
populations ont persisté dans le temps, variant uniquement leur
densité numérique; par ailleurs ont été
enregistrés des changements dans la structure de la communauté
indiquant un dynamisme marqué, résultat de l'influence de
facteurs internes et des conditions du milieu. La structure de la
communauté a été affectée principalement par le
comportement des phytoflagellés et dinoflagellés, lesquels
effectuent des migrations verticales pendant le cycle
nycthéméral.
Las unidades básicas de una comunidad están representadas por las especies cuyas interrelaciones le otorgan una cierta estructura, la que se ve afectada por diversos factores que alteran las relaciones cualitativas y cuantitativas del conjunto. Una comunidad fitoplanctónica es pues susceptible de cambios en su microambiente, surgiendo numerosas fluctuaciones cuyo resultado es la respuesta integral a las alteraciones del medio, así como a diversos factores intrínsecos propios de las mismas especies u organismos. Considerando la continuidad que existe en esta taxocenosis y el hecho de presentar una tasa de renovación acelerada, un problema desafiante se presenta al considerar la inestabilidad temporal y la irregularidad espacial de las poblaciones fitoplanctónicas en periodos cortos, de lo que se deriva la importancia o interés de un estudio durante un ciclo nictemeral, el cual podrá proporcionar una imagen más o menos clara del comportamiento de los fitoplanctontes así como permitir determinar ciertas características del conjunto que no son dadas por los organismos o especies considerados separadamente. La estimación de la diversidad se ha revelado como un buen auxiliar para el entendimiento del dinamismo de las comunidades por el hecho de dar una imagen de su estructura, la que permite determinar de una manera indirecta el funcionamiento y hasta el grado de eficiencia de éstas (Margalef, 1970). A pesar de que una cierta regularidad en la distribución de individuos en especies ha sido observada por varios autores (Fager, 1963; Margalef, 1957) existen alteraciones en las relaciones cualitativas y cuantitativas del conjunto, las cuales pueden ser evaluadas por medio de los índices de afinidad. En noviembre de 1973 se efectuó un muestreo del fitoplancton durante un ciclo de 24 horas en una estación fija situada en la Bahía de Campeche, frente a la Laguna de Términos (Fig. 1). Los resultados de la composición cualitativa y cuantitativa del fitoplancton fueron presentados por Santoyo y Signoret (1975). En el presente trabajo se aborda el análisis de la diversidad y de la afinidad del fitoplancton en dicho ciclo nictemeral. DIVERSIDADHa sido señalado por diversos autores que la diversidad es un indicador de la complejidad estructural de una comunidad, tomándose como punto de partida cierta regularidad en la distribución de los individuos en especies. Margalef (1957) mostró la utilización de la teoría de la información sobre algunos aspectos en ecología, derivando de ella un índice de diversidad que permite conocer la madurez de un ecosistema dado. Las medidas de información están relacionadas con el principio de incertidumbre siendo que el contenido de información de una comunidad es equivalente a la incertidumbre involucrada en predecir el próximo encuentro al azar de un organismo de una especie con otro individuo de la misma u otra especie; es decir, que una mayor incertidumbre conlleva a una mayor diversidad (Lloyd et al., 1968). 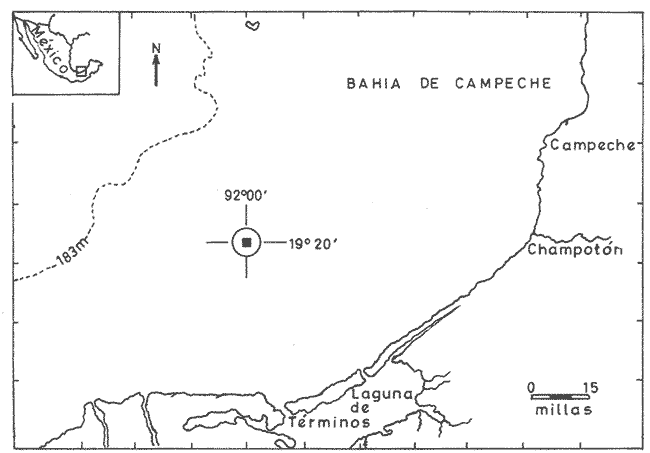 Fig. 1. Área de estudio y localización de la estación de muestreo. Pielou (1966a y 1966b) hizo una evaluación de los diferentes colectivos que pueden estar sujetos a un análisis estructural por medio de los elementos poblacionales y definir por sus características la fórmula adecuada para la obtención del índice de diversidad. De acuerdo al criterio de este autor se utiliza la ecuación dada por Shannon y Weaver (1963) en la que se efectúa una estimación de la diversidad por medio de una muestra de la comunidad en la que se desconoce el valor real del número de individuos pertenecientes a cada una de las especies, postulándose que la comunidad es grande al grado de considerarse infinita y que contiene S especies en proporciones p1, p2, ... Ps de manera que la diversidad o información por individuo está dada por la siguiente función: H'=-sumatoria donde i= 1 hasta s de Pi log2 Pi bits * de información / individuo en la que pi = Ni/N = es la proporción de la i especie en la comunidad. Ni = número de individuos de la i especie. N = número total de individuos de la comunidad. La diversidad de especies obtenida de la manera ya descrita tiene otro componente que es la denominada "equitatividad" o la abundancia relativa de las especies (Lloyd y Ghelardi, 1964) que Pielou (1966b y 1966c) denomina "igualdad" (eveness) asumiéndose teóricamente, para la estimación de esa característica, que todas las especies de un colectivo están igualmente representadas o son equifrecuentes, particularidad que conduce a obtener la diversidad máxima dada por el log2 S (H'max); este valor al ser comparado con la diversidad calculada (H') por medio de la relación: J'= H'/H'max puede ser considerada como una medida del grado de organización de la comunidad (Margalef, 1974). Un incremento en el número de especies y/o una tendencia a una distribución equifrecuente de los individuos en especies conduce a un valor elevado de H' y asimismo al resultado de J´. Es menester hacer la consideración de que los índices de diversidad fueron subestimados, en virtud de que algunos organismos no fueron identificados a nivel específico y sólo quedaron registrados a nivel de grupo, siendo considerados como representativos de una especie para el efecto de los cálculos; tal fue el caso de las dinoflageladas, fitoflageladas, cianofitas y clorofitas (Santoyo y Signoret, 1975) ; ésto da un margen de error ya que los individuos agrupados de esta manera lógicamente pertenecen a más de una especie. Las fluctuaciones nictemerales de la diversidad estuvieron enmarcadas por un valor mínimo de 1.9 bits/individuo y un máximo de 4.3 bits/ individuo (tabla l). De manera general se registró un gradiente de la diversidad de especies al iniciarse el ciclo nictemeral (0618 horas) con los valores mayores en las profundidades de 20 y 30 metros y un núcleo pequeño en la super ficie, disminuyendo hacia las horas de iluminación hasta registrarse los valores menores en las primeras horas de oscuridad en la capa de 0 a 20 metros seguido, posteriormente, de un incremento a partir de la media noche hasta el crepúsculo matutino en donde se identificó una disminución (Fig. 2). El índice de igualdad J' tuvo variaciones considerables con los límites señalados por los valores de 0.47 y 0.94 como mínimo y máximo respectivamente (tabla 1) en los que la riqueza de especies (número de especies) no tiene relación alguna con el valor del índice de igualdad. Efectuándose la correlación (coeficiente r) entre demostrándose que las variables son independientes. Fager y Hasler (1969) obtuvieron resultados similares, sugiriendo que la magnitud de la igualdad en el fitoplancton queda definida en su mayor porción por las 10 a 15 especies más abundantes y que las especies con menores efectivos ejercen una influencia mínima en este componente de la comunidad. 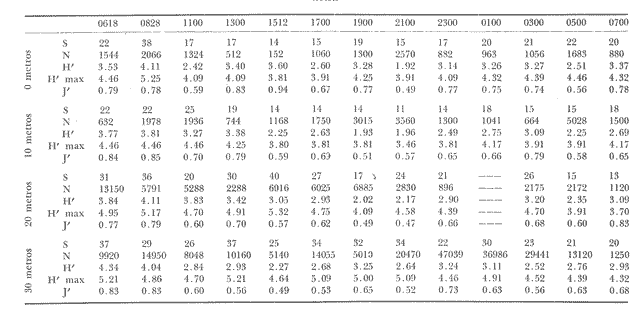 TABLA 1 VALORES DEL NÚMERO DE ESPECIES (S) NÚMERO DE CELULAS POR LITRO (N),INDICE DE DIVERSIDAD (H'). INDICE DE DIVERSIDAD TEÓRICA (H' MAX) E ÍNDICE DE IGUALDAD (J' = H' /H' MAX) AL TRAVÉS DE UN CICLO NICTEMERAL. HORAS 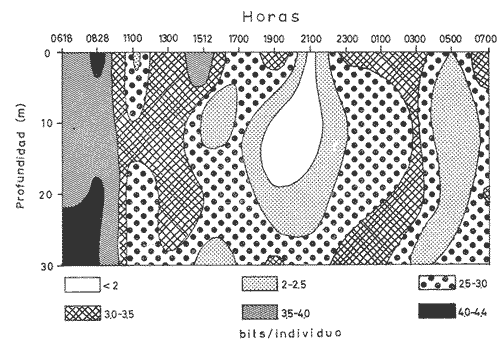 Fig. 2. Variación nictemeral de la diversidad (H') fitoplanctónica. Es interesante hacer notar que el valor máximo de igualdad estuvo dado con un valor de S = 14 y N = 152 células por litro en la capa superficial (1 512 horas) ; esto demuestra lo ya señalado e indica una distribución de los individuos en las S especies casi equifrecuente (Fig. 3) ; en cambio, con un valor máximo registrado de 40 especies (al nivel de 20 metros, a las 2100 horas) el valor de J' fue de 0.57, lo cual se debió a la dominancia del grupo de fitoflageladas en una proporción del 66 por ciento (Santoyo y Signoret, 1975) hecho que influyó en forma definitiva para abatir el índice de igualdad; de manera que a pesar de haberse presentado el máximo de especies en el ciclo no se alcanza el valor más alto de diversidad. La relación del índice de diversidad con el índice de igualdad se evidencia mirando las fluctuaciones del valor de J' al través del tiempo (Figs. 2 y 3) con incrementos en las horas donde hubo alta diversidad (0618-0828 horas) y decrementos en las horas en donde se obtuvieron bajos valores de H' (crepúsculo vespertino y primeras horas de la noche y crepúsculo matutino) . 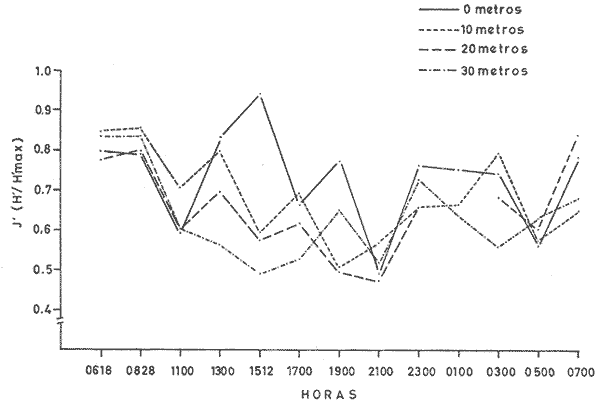 Fig. 3. Variación del índice de igualdad J' (H' / H' max) a diferentes niveles en el ciclo AFINIDADEn el estudio de las comunidades fitoplanctónicas aparecen ciertos aspectos interesantes en cuanto a su estructura que se traducen a la presencia de especies comunes en el tiempo y en el espacio. La afinidad cualitativa y cuantitativa que exista en el fitoplancton no sólo sirve para identificar y describir a la o las comunidades, sino también para apreciar el efecto que pueden tener los factores ambientales sobre las especies existentes en un mismo ambiente, además de la mejor comprensión de las relaciones poblacionales y los cambios biológicos asociados con fenómenos de sucesión en esta taxocenosis. Considerando que el fitoplancton tiene, de manera general, una tasa de renovaciones elevada, la evolución de las poblaciones es rápida, por consiguiente se deriva el interés de analizar la similaridad o afinidad en periodos cortos. Los índices de afinidad o similaridad han sido aplicados de manera preponderante al estudio de comunidades terrestes (Renkonen, 1938; Kontkanen, 1950; Elton y Miller, 1954; Davis, 1963; Southwood, 1966) . Son pocos los trabajos que se conocen donde se aplique este método para el estudio de comunidades acuáticas; entre éstos se podría citar el de Whittaker y Fairbanks (1958) donde se aborda el estudio de las comunidades zooplanctónicas de diversos cuerpos de agua; Sanders (1960) describe y delimita comunidades bénticas; Fager (1963) estudia las comunidades marinas de manera general. Con relación al fitoplancton, a pesar de que éste forma parte de un ecosistema abierto, se han contemplado aspectos de similaridad tanto de especies como de sus densidades (Fager, 1963) ; sin embargo, no se conocen trabajos específicos donde se aplique el uso de índices de afinidad en el estudio de este grupo. 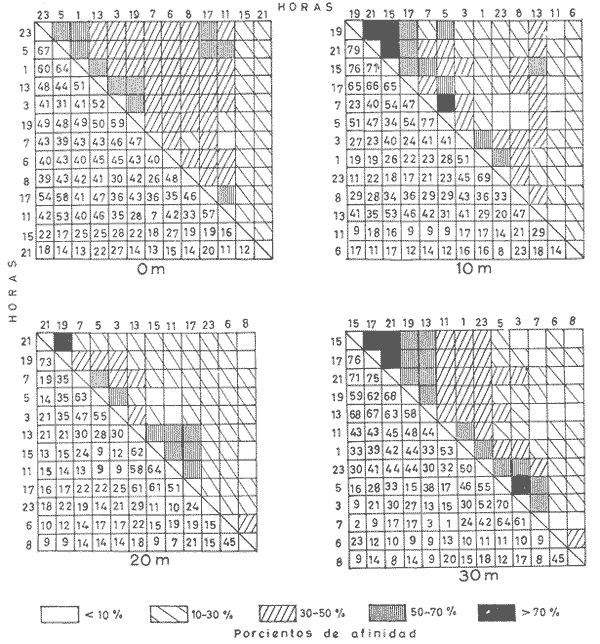 Fig. 4. Variación de la afinidad en cada nivel durante el ciclo nictemeral. En el presente estudio los valores de afinidad se calcularon aplicando la fórmula de Renkonen (1938) con la cual se obtienen porcientos de afinidad entre dos muestras; el valor resultante corresponde a la suma de los porcientos menores de las especies comunes en ambas muestras de acuerdo a la siguiente fórmula: A = Sumatoria min (a,b,...,x) donde A = porciento de afinidad Sumatoria min = suma de los porcientos menores de las especies a, b, ..., x, comunes de dos muestras Para el análisis de la afinidad en el ciclo nictemeral se hicieron los cálculos para cada nivel entre las diversas horas (Fig. 4) y para cada hora entre los diversos niveles (Fig. 5). Los valores obtenidos presentaron marcadas fluctuaciones durante el ciclo nictemeral, siendo el valor mínimo de 1 porciento, encontrado en el nivel de 30 metros, entre las 0700 y las 1100 horas y el máximo de 82.9 porciento, encontrado a las 1900 horas entre los niveles de 10 y 20 metros. Considerando las afinidades calculadas para cada nivel entre las diversas horas, se registraron núcleos muy aparentes de gran afinidad. En el nivel superficial se presentaron valores mayores del 50 porciento, dados principalmente por una marcada abundancia de Thalassiothrix frauenfeldii y dinoflageladas desnudas; por otro lado, entre las 0700 y las 1100 horas la afinidad fue mínima con un valor de 7 porciento; hay que considerar que a las 0700 horas se registró una abundancia considerable de Trichodesmium Thiebautii mientras que a las 1100 horas abundaron otras especies tales como Nitzschia acicularis y Thalassiothrix frauenfeldii. De una manera general la afinidad en este nivel fue relativamente regular (Fig. 4). En el nivel de 10 metros los valores fluctuaron más ampliamente, presentándose núcleos aparentes de gran afinidad (mayor del 70 porciento) en las primeras horas de oscuridad y en la madrugada (Fig. 4), caracterizados por la abundancia de Thalassionema nitzschioides, Thalassiothrix frauenfeldii y dinoflageladas desnudas y entre las 1900 y 2100 horas por fitoflageladas cuya proporción fue mayor del 60 porciento en ambas horas. Los mínimos de afinidad se presentaron en varias ocasiones a las 1100 horas, siendo que la composición fitoplanctónica a esta hora fue particular debido a una proporción conspicua de Nitzschia acicularis (tanto en el nivel superficial como en el de 10 metros) y de clorofitas, fítoplanctontes que no aparecen en otras horas o de ser así en baja proporción. En el nivel de 20 metros la mayoría de los valores se encuentran entre 10 y 30 porciento pero se identifica un valor de afinidad elevado entre las 1900 y las 2100 horas, con una marcada abundancia de fitoflageladas. Se registraron igualmente valores del 50 al 70 porciento en las horas del medio día con Nitzschia longissima como especie abundante. Los valores de afinidad bajos se presentaron entre horas discontinuas. 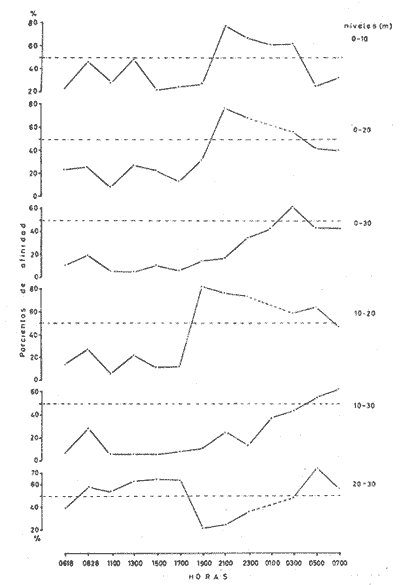 Fig. 5. Variación de la afinidad entre niveles durante el ciclo nictemeral. En el nivel de los 30 metros se registró una mayor afinidad al través del tiempo presentándose los valores más altos al atardecer y en la madrugada, con la presencia de Nitzschía longissima y Pleurosigma normanii como especies más abundantes en el primer caso y con Thalassionema nitzschioides, Thalassiothrix frauenfeldii y dinoflageladas desnudas en el segundo. Se registró igualmente un núcleo de afinidad inferior al 10 porciento en las horas de la mañana y otro entre las 0700 y diversas horas. Considerando las afinidades calculadas entre los diversos niveles, se obtuvieron igualmente fluctuaciones considerables (Fig. 5). En la capa de 0 a 20 metros la afinidad presentó fluctuaciones de las primeras horas al atardecer, a partir del cual hubo un incremento notable en los valores (de menos del 10 porciento a más del 70 porciento). La afinidad se mantuvo alta durante la noche decreciendo paulatinamente hasta la madrugada donde los valores permanecieron cercanos al 50 porciento. En el nivel de los 30 metros se observó un comportamiento inverso de la afinidad, siendo que los valores más altos se registraron durante el día y al amanecer, mientras que al atardecer se presentó un decremento brusco de la afinidad la cual se fue incrementando nuevamente hacia el crepúsculo matutino. Se observa pues, la presencia de dos capas con características opuestas; con respecto a esto hay que considerar que en la capa profunda se presentó una abundancia notable de diatomeas de hábitos bénticos que pasan a formar parte del ticoplancton o plancton accidental, otorgándole una estructura diferente a la de las capas suprayacentes. Por otro lado, la afinidad calculada entre niveles discontinuos presentó igualmente fluctuaciones; sin embargo, entre la superficie y los 20 metros se registró un patrón de variación semejante al de los valores obtenidos entre las capas continuas; entre los niveles superficie-30 metros y 10-30 metros la afinidad fue notablemente baja, sin embargo, en el crepúsculo matutino los valores fueron superiores al 50 porciento o cercanos a este valor. ConclusionesDISCUSIÓN Y CONCLUSIONESLa expresión probabilística de Shannon y Weaver da una medida de la organización o grado de complejidad de la estructura de una comunidad; considerando lo anterior, la taxocenosis fitoplanctónica tuvo modificaciones estructurales al través del tiempo. En el ciclo nictemeral se registraron por un lado estados estructurales pobres y por otro estados con mayor complejidad, significando que el potencial de organización no se mantiene en la comunidad, como resultado de las fluctuaciones poblacionales así como de las condiciones ambientales. La estructura se vio afectada principalmente por el comportamiento de las fitoflageladas y dinoflageladas, grupos que presentan migraciones verticales durante el ciclo nictemeral en las que la iluminación parece intervenir de manera directa (Santoyo y Signoret, 1975). Existen otros factores que intervienen igualmente en las alteraciones de la estructura de la comunidad tales como: la acumulación de metabolitos en aguas estratificadas que tienen un efecto deletéreo sobre, algunas especies significando una ventaja para otras (Margalef, 1958), la reproducción diferencial de las diversas especies, la segregación específica, la competencia, la depredación por zooplanctontes fitófagos o filtradores, etcétera. Finalmente hay que considerar que la densidad de una especie no sólo es función de las otras sino también de la suya propia, tanto en áreas vecinas como en un solo punto en tiempos diferentes y que los cambios cualitativos y cuantitativos en una comunidad son resultado de la respuesta de las especies dándole al conjunto características que no son dadas por ellas en particular. Las marcadas fluctuaciones registradas señalan un gran dinamismo de la comunidad, dando como resultado un cambio continuo en su estructura y por ende en su organización. Santoyo y Signoret (1975) ya habían planteado el hecho de que la comunidad fitoplanctónica en la Bahía de Campeche se caracterizaba por su heterogeneidad. El acelerado dinamismo y la heterogeneidad de esta taxocenosis hacen que la correspondéncia entre las condiciones ambientales y las poblaciones de diferentes especies sea imprecisa, por lo que es recomendable buscar las relaciones no con factores aislados, sino con características globales del medio ambiente tales como la estabilidad, turbulencia, etcétera (Margalef, 1970) . Las reducidas fluctuaciones que presentaron la temperatura y la salinidad (de 24.5ºC a 26.7ºC y de 36.60 a 36.91 O/oo) en el área de estudio sugieren la presencia de masas de agua estables y estratificadas (Santoyo y Signoret, 1975). Esta estratificación en las masas de agua se proyecta en la comunidad fitoplanctónica por los valores de diversidad y de afinidad. De acuerdo a estos parámetros, se identificaron dos capas diferentes en estructura, la capa superficial, con valores inconstantes de diversidad y afinidad baja, con fluctuaciones al través del tiempo y la capa del fondo con valores altos tanto de diversidad como de afinidad y con una composición diferente, caracterizada por la presencia de diatomeas de hábitos bénticos. Entre ambas capas se presenta una zona de transición con valores de diversidad en general bajos y con afinidad variable. El análisis de los patrones de variación de la diversidad y de la afinidad durante el ciclo nictemeral conduce a hacer varias consideraciones: durante el periodo de 24 horas hubo poblaciones persistentes al través del tiempo, variando únicamente su densidad numérica (Thalassiothrix frauenfeldii, Thalassionema nitzschioides, dinoflageladas desnudas y tecadas) además de otras especies que persisten pero en un determinado nivel. Por otro lado, durante el ciclo nictemeral la relativa estabilidad ambiental observada, propicia en determinados momentos una distribución casi equifrecuente de individuos en especies y por lo tanto con un valor de J' elevado, implicando posiblemente una superposición de nichos o a lo que se le ha llamado la "paradoja de Hutchinson" (Hutchinson, 1961; Margalef, 1969). Margalef (1970, 1974) plantea que la diversidad no es una buena medida de la organización de un sistema si no se considera lo que él llama "persistencia" de la estructura de la comunidad, por lo cual, durante el ciclo nictemeral se pretendió observar este parámetro pero sin llegar a evaluarlo de manera directa; según este autor, la persistencia es una expresión cuantitativa de las medias armónicas de la vida media de las especies presentes. Tal vida media también involucra las pérdidas por difusión, hundimiento, etcétera, sin embargo, la utilización de los índices de afinidad coadyuvaron a interpretar los cambios estructurales de la comunidad. La ubicación de una estación fija de muestreo no representa un estado estacionario de las condiciones bióticas y abióticas y permite hacer inferencia del comportamiento y cambios de estructura de una taxocenosis lábil como lo es el fitoplancton en periodos cortos. AgradecimientosSe hace patente el reconocimiento a Guillermo Villarreal por su inapreciable ayuda en los cálculos de los índices de diversidad y afinidad de este estudio. LITERATURADAVIS, B. N. K. A study of micro-arthropod communities in mineral soils near Corby, Northants. J. Anim. Ecol. 1963 49-71 32 ELTON, C. S. y R. S. MILLER The ecological survey of animal communities: with a practical system of classifying habitats by structural characters. J. Ecol. 1954 460-489 42 FAGER, E. W. The composition of sea-water comparative and descriptive oceanography. Communities of organism.s. In: M. N. Hill (Ed.), Thea Sea. Ideas and Observations on Progress in the Study of the Seas. II. Interscience Publ., Londres 1963 415-437 FAGER, P. E. y A. D. HASLER Species diversity in lacustrine phytoplankton. I. The components of the index of diversity from Shannon's formula. Amer. Natur. 1969 51-59 103 (929) HUTCHINSON, G. E. The paradox of the plankton. Amer. Natur. 1961 137-145 95 KONTKANEN, P. Quantitative and seasonal studies on the leafhopper fauna of the field stratum on open areas in north Karelia Ann. Zool. Soc. Vanamo 1950 191 13 (8) LLOYD, M. y R. S. GHELARDI A table for calculation the "equitatibility" component of species diversity J. Anim. Ecol. 1964 217-225 33 LLOYD, M., J. H. ZAR y J. R. KARR On the calculation of information-theoretical measures of diversity. Amer. Natur. 1968 257-272 79 (2) MARGALEF, R. La teoría de la información en ecología. Mem. Real. Acad. Cienc. Artes Barcelona 1957 373-445 32 (3) Temporal succession and spatial heterogeneity in phytoplankton. A. A. Buzzati-Traverso Perspectives in marine biology. Calif. Univ. Press Berkeley 323-348 1958 Estudios sobre la distribución a pequeña escala del fitoplancton marino. Mem. Real Acad. Cienc. Artes Barcelona 1969 223-243 40 (1) Organization and distribution of phytoplankton communities. Final Tech. Rep. Office Naval Res., (mimeogr.) 1970 16 p. Ecología. Omega Barcelona 1974 951 p. PIELOU, E. C. The measurement of diversity in different types of biological collections. J. Theoret. Biol. 1966a 131-144 13 Shannon's formula as a measurement of specific diversity: its use and misuse. Amer. Natur. 1966b 463-465 100 (914) Species diversity and pattern-diversity in the study of ecological succession. J. Theoret. Biol. 1966c 370-383 10 RENKONEN, O. Statistisch-ökologische untersuchungen über die terrestrische käferwelt der finnischen bruchmoore. Ann. Zool. Soc. Vanamo 1938 1-227 6 (1) SANDERS, H. L. Benthic studies in Buzzards Bay. III. The structure of the soft-bottom community. Limnol. Oceanogr. 1960 138-153 5 SANTOYO, H. y M. SIGNORET Variación nictemeral del fitoplancton en la Bahía de Campeche, México. Rev. lat-amer. Microbiol. 1975 161-157 17 (3) SHANNON, C. E. y W. WEAVER The Mathematical Theory of Communication. Univ. Illinois Press, Urbana. 1963 SOUTHWOOD, T. R. E. Ecological Methods with Particular Reference to the Study of Insect Populations. Methuen Londres 1966 391 p. WHITTAKER, R. H. y C. W. FAIRBANKS A study of -plankton copepod communities in the Columbia Basin, southeastern Washington. Ecology 1958 46-65 39 (1)
|