|
OBSERVACIONES SOBRE LA BIOLOGIA, ECOLOGIA Y
EXPLOTACIÓN DE MEGAPITARIA AURANTIACA (SOW., 1831), M. SQUALIDA (SOW.,
1835) Y DOSINIA PONDEROSA (GRAY, 1838) (BIVALVIA: VENERIDAE) DE LA BAHÍA
DE ZIHUATANEJO E ISLA IXTAPA, GRO., MÉXICO
Trabajo recibido el 12
de agosto de 1976 y aceptado para su publicación el 15 de octubre de
1976.
E. BAQUEIRO
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología.
J. STUARDO
Proyecto Multinacional en Ciencias del Mar de la OEA
en México. Contribución 65 del Centro de Ciencias del Mar y
Limnología, UNAM.
Se hace un análisis de las
características de longitud, altura y peso total de las poblaciones de
Megapitaria aurantiaca, M. squalida y Dosinia ponderosa
durante el ciclo anual comprendido entre septiembre de 1974 y septiembre de
1975. Estas tres especies de "almejas" de la Bahía de Zihuatanejo e Isla
Ixtapa, Gro., son de elevado valor comercial en las pesquerías del
área.
Las características hidrológicas
generales encontradas son muy semejantes a lo reportado por los escasos autores
que han estudiado el área, disminuyendo la salinidad a un mínimo
en el mes de agosto, al igual que la temperatura en marzo; los valores
máximos para la salinidad se encontraron en diciembre y para la
temperatura en septiembre de 1974 y 1975.
Del análisis de las variaciones de la longitud
total, alto y peso total, así como del estudio del desarrollo
gonádico se concluye que: a) Se observa la presencia por lo menos de
tres clases de edad con un incremento paulatino de las clases de menor
tamaño, mismo que no se observa en las clases mayores, ya que
desaparecen por efectos de la explotación; b) La reproducción es
más o menos constante durante todo el año con dos periodos de
desove máximo en los casos de M. aurantiaca y
M. squalida y tres para D. ponderosa,
periodos que coinciden con la presencia de juveniles en la población dos
meses después; c) Las tres especies viven enterradas en el fondo donde
el movimiento del agua es constante; su distribución está
determinada principalmente por el tipo de substrato.
En la fauna acompañante predominan, otros
bivalvos, entre los que Chione undatella presenta posiblidades de
explotación.
Se dan las características del ciclo
gonádico de las tres especies y se hacen recomendaciones para una
explotación controlada basada en la determinación de tallas
mínimas y vedas alternadas de los bancos en explotación,
planteándose la posibilidad de mantener ésta durante todo el
año con artes de pesca mecanizados, siempre y cuando se amplíen
las áreas de pesca.
An analysis of the weight, height, length and width
in populations of the "clams" Megapitaria aurantiaca, M.
squalida and Dosinia ponderosa from Zihuatanejo
and Isla Ixtapa, Guerrero, Mexico including a description of its reproductive
cycle is given for an annual period going from september 1974 to september
1975.
The general oceanographic characteristics found in
the area at the time of study seem similar to those described by a few other
authors: low salinity values were recorded in august and the lowest
temperatures in march; the highest salinity values were measured in december
and the highest temperatures in september.
The main conclusions drawn from the analysis of the
structure of the population follow: a) there are at least three classes with
more or less constant growth, better observed in the smaller classes but not in
the older ones as they are rapidly withdrawn from the population by fishing; b)
reproduction is constant throughout the year with two maximal spawning periods
for M. aurantiaca and M. squalida and
three for D. ponderosa; these periods were always followed
by the appearance of conspicuous juveniles two months later; c) the three
species live buried in bottoms where there is a constant water movement but
their distribution is regulated by the nature of the substratum.
The associated fauna is mainly represented by bivalves
of which Chione undatella may have commercial fishery
importance.
Recommendations are given for a controlled fishery,
bases on minimal length and alternated exploitation of the banks. It is
suggested that a mechanized fishery could sustain production throughout the
year.
Este trabajo formó parte de los estudios sobre Uso de la Zona Costera de los Estados de Michoacán y Guerrero que el Centro de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México, realizara en contrato para la Comisión del Río Balsas de la Secretaría de Recursos Hidráulicos entre 1974 y 1975. De los resultados obtenidos en el estudio prospectivo de la zona costera de Michoacán y Guerrero (Stuardo et al., 1974), se concluyó que la explotación de moluscos marinos, representa una de las pesquerías que a nivel nacional ha venido aumentando progresivamente y corresponde al tercer rubro más importante dentro de la explotación pesquera nacional. Para Zihuatanejo, Gro., la producción de "almejas" es de suma importancia en la pesquería local ya que en orden de importancia ocupa el segundo lugar, con una producción en el año de 1972 de 87 670 docenas con valor de $ 615,916.00, superada sólo por la del ostión, que el mismo año produjo 159 447 kilos con un valor de $ 645,565.00. Esta importancia es el reflejo de la abundancia y calidad de las especies reunidas bajo ese nombre, que habitan en la Bahía de Zihuatanejo y sus alrededores. El desconocimiento total de los aspectos biológicos y ecológicos de las especies explotadas, así como la disminución indudablemente notoria del tamaño medio de sus poblaciones por sobrexplotación, determinó la necesidad de estudiarlas, lo que corresponde al objeto de la presente contribución. Los trabajos de tipo faunístico, ecológicos generales o monográficos publicados referentes a la costa de Guerrero y/o Zihuatanejo son escasos. Stuardo y Martínez (1974) y en especial Stuardo et al. (1974) en una prospección de estas costas ponen énfasis especialmente en aquellas especies, particularmente invertebrados de la costa rocosa, de reconocida importancia económica y discuten la mayoría de las referencias conocidas; poco antes Tovar y Sánchez (1974) efectuaron un estudio estacional de los grupos planctónicos recalcando la abundancia relativa de las larvas de moluscos bivalvos en la Bahía de Zihuatanejo, incluyendo la descripción general de los sedimentos y su contenido de carbonatos y materia orgánica. Esta información se ha utilizado parcialmente en las discusiones de este trabajo. Aun cuando hasta la fecha no se habían realizado investigaciones sobre la biología, dinámica de poblaciones o ecología de las especies de "almejas" de Zihuatanejo, los estudios realizados sobre la anatomía general, distribución y ciclo reproductor de especies foráneas del mismo género (Norton, 1917; Lozano, 1952) o géneros afines (Ansell, 1961; Trevallion y Edwards, 1970) u otras familias (Trevallion, 1971; Olivier et al., 1971) si no directamente comparables, sobre todo porque provienen de especies de áreas climáticamente muy diferentes, sirvieron inicialmente para precisar la extensión y contenido de los estudios realizados en Zihuatanejo sobre estructura de las poblaciones y sus variaciones en el tiempo, crecimiento, anatomía y reproducción de las "almejas roja, negra y blanca" "Megapitaria aurantiaca, M. squalida y Dosinia ponderosa. No todos los resultados obtenidos se incluyen en el presente trabajo. ÁREA DE ESTUDIOEl Estado de Guerrero se encuentra en la vertiente del Océano Pacífico, donde la planicie costera es estrecha, bordeada por la Sierra Madre Occidental. El clima en esta zona ha sido clasificado por García (1973) como del subtipo AW" ó (W) i, o sea, cálido subhúmedo con dos periodos de lluvias, uno máximo en verano (mayo-agosto) más lluvias invernales ocasionales (diciembre-enero) separados por sequías. Las temperaturas más altas se presentan antes del mes de junio; la temperatura media para el periodo de 1944-1972 fue de 26.2º C y la precipitación pluvial anual de 110.3 mm. Los vientos dominantes de S-SW, están dados por la cercanía de la Sierra. El principal aporte fluvial cercano lo constituye el Río Balsas localizado a 14 km de Isla Ixtapa, el que junto a otros arroyos pequeños vierten agua dulce durante el periodo de lluvias. En la Bahía de Zihuatanejo descargan también algunos arroyos a través de tres pequeños esteros o lagunetas. Respecto a las características hidrológicas costeras, Tovar y Sánchez (1974) reportan que en un ciclo anual de controles estacionales la temperatura máxima del agua del mar registrada fue de 32º C en el verano y la mínima de 20° C en primavera con una media anual de 27.3º C; la salinidad máxima reportada fue de 36%o en verano, otoño e invierno, con una media anual de 35.5 %. 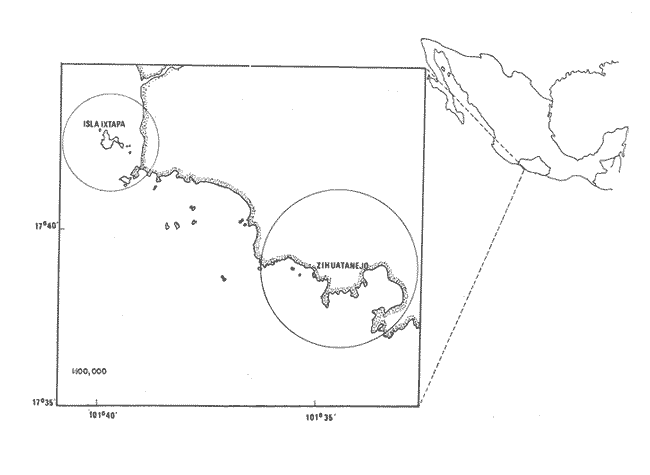 Mapa 1. Localización de las áreas de estudio en la costa del estado de Guerrero, México. El área de estudio que comprendió sectores de la Bahía de Zihuatanejo y de los alrededores de Isla Ixtapa, está localizada en la porción norte de la costa del Estado de Guerrero (Mapa 1), y es clasificada como un litoral rocoso con acantilados (Lankford, 1974). En esta zona se encuentran algunas playas de arena, ubicadas principalmente en bahías y ensenadas. La Bahía de Zihuatanejo (Mapa 2a) , está localizada en los 17° 37' de latitud norte y 101º 33' de longitud oeste; tiene una extensión de 3.52 km2 y presenta varias playas arenosas (separadas por puntas rocosas), de poca pendiente que se continúan hasta una profundidad máxima de 25 m al centro de la boca de la bahía. Los sedimentos están constituidos principalmente por arena o arenalimo; en algunos puntos se encuentra arena gruesa y grava, especialmente en las puntas y boca de la bahía, en donde Tovar y Sánchez (op. cit.) reportan fondos de arena, areno-migajosos, migajón-arenosos y de roca. La Isla Ixtapa o Isla Grande (Mapa 3a) está localizada entre los 17° 40' de latitud norte y 101º 40' de longitud oeste, a 366 mts de la playa vecina apunta Ixtapa (playa quieta), hayándose separada de la costa por un canal de poca profundidad (10 cm como máximo) con fondo plano, arenoso o areno-limoso. La isla es pequeña, de forma irregular, bordeada por farallones rocosos y playas arenosas pequeñas en bahías de fondo arenoso y coralino. MATERIALES Y MÉTODOSEste estudio se inició en septiembre de 1974, habiéndose llevado a cabo 10 muestreos mensuales en las siguientes fechas: 6 de septiembre, 12 de octubre, 2 de noviembre y 6 de diciembre de 1974; 14 de marzo, 4 de mayo, 10 de junio, 7 de julio, 6 de agosto y 1° de septiembre de 1975. No fue posible realizar los muestreos de enero, febrero y abril de 1975, por diversas causas. La posición de las estaciones fue fijada una vez que se ubicaron y exploraron todos los bancos conocidos, tanto en la Bahía de Zihuatanejo, como en los alrededores de Isla Ixtapa, escogiéndose los puntos de muestreo por su accesibilidad y abundancia de ejemplares. Dichos puntos están representados en los mapas 2a y 3a que corresponden a 4 bancos de la Bahía de Zihuatanejo y 4 en Isla Ixtapa. Para obtener las muestras se empleó una lancha de fibra de vidrio con motor fuera de borda; los ejemplares fueron recolectados a mano por medio de buceo autónomo (S. C. U. B. A.), localizándolos con la ayuda de una barra de metal que servía de sondeador, técnica seguida por los pescadores locales. La determinación de la densidad de población se realizó empleando un marco de aluminio de un metro cuadrado que se fijó al fondo. Escarbando en la arena con un rastrillo hasta una profundidad de 30 cm se extraían todos los macrorganismos que ahí se encontraban. Conjuntamente al muestreo de densidad de población, se medía la profundidad con una sondaleza de mano y la temperatura del agua del fondo y del sedimento con un termómetro, con divisiones de 1º C y escala de -15º C a 150º C. Además, se obtenían muestras de agua, con una botella de plástico que era llenada en el fondo, para conocer la salinidad, la cual era determinada en la estación de Isla Ixtapa con un refractómetro de mano "Spekol". En cada muestreo se obtuvieron 50 ejemplares de cada una de las especies estudiadas, provenientes de los bancos 1 y 4 de Zihuatanejo y 2 y 4 de Isla Ixtapa, y muestras adicionales de 1 m2, con un número variable de ejemplares y especies. En el laboratorio se midió el largo, ancho, alto, peso total y peso húmedo sin concha, de cada uno de los ejemplares obtenidos. Las mediciones se hicieron con un calibrador (Vernier) hasta centésimas de milímetro y los pesos se midieron con una báscula de brazo, hasta décimas de gramo. Doce de los ejemplares colectados de cada especie fueron fijados en Bouin para ser procesados con las técnicas histológicas corrientes a fin de determinar el ciclo reproductor y madurez sexual; algunos otros ejemplares de cada especie fueron fijados en formol al 10% para estudios anatómicos posteriores. A partir del mes de marzo de 1975, se fijaron al fondo, en las cercanías de Isla Ixtapa, jaulas de metal con tela plástica en las que se colocaron ejemplares marcados para control de crecimiento de cada una de las especies estudiadas. Las medidas y recuentos obtenidos se han procesado estadísticamente con la ayuda de un programa S. P. S. S. (Statistical Package for the Social Sciences. University of Chicago and Alberta) y representado, en tablas y gráficas que objetivan las características de las poblaciones estudiadas. Los valores de tendencia central dados por la media, moda y mediana, que en una curva normal de población deben presentar el mismo valor, presentan un desplazamiento dado por los valores de Kurtosis y Skewness que indican la presencia y dominancia de una clase de edad dada. En todos los casos el rango de valores determinados es muy amplio, lo que se refleja en los elevados valores para la Varianza y Desviación Standard, mismos que indican lo heterogéneo de las muestras. Las especies encontradas se han determinado según las técnicas taxonómicas usuales, utilizando la literatura especializada respectiva.  ABREVIACIONES EMPLEADAS EN EL ANÁLISIS ESTADÍSTICO RESULTADOS
TAXONOMÍA Y DISTRIBUCIÓNDescripciones generales de estas especies han sido dadas pr Olsson (1961) y Keen (1958, 1971). Las tres especies pertenecen a la familia Veneridae, representando a dos sub-familias diferentes. En la sub-familia Pitarinae se incluye Megapitaria aurantiaca y M. squalida; y en la Dosiniinae a Dosinia ponderosa. Estas especies junto al "ostión de roca" Crassostrea iridescens y la "lapa", Ancistromesus mexicanus, son los moluscos económicamente más importantes en la región. Género MegapitariaMegapitaria aurantiaca (Sowerby, 1831). Lámina 1. a y b. Descripción: Concha gruesa, de forma triangular, anteroposteriormente alargada. Perióstraco grueso. Charnela gruesa con tres dientes cardinales y laterales anteriores muy marcados (heterodonta); a cada diente corresponde un cóndilo en la valva contraria. Lúnula poco marcada. Ligamento externo, colocado sobre una plataforma ligeramente escarbada en la porción posterodorsal de la charnela. Huella paleal bien marcada; seno paleal en forma de lengua, extendiéndose horizontalmente hasta la mitad de la concha. Huella de los aductores circular, un poco mayor la posterior, a la cual se une la huella del retractor posterior; huella del aductor anterior hundida, bien marcada. Perióstraco de color caférojizo. Concha de color rosáceo o encarnado. Interior blanco. Chamela púrpura en su parte posterior. Distribución: Esta especie es reportada desde el golfo de California, México a Salinas, Ecuador (Olsson 1961; Keen 1971) incluyendo la Bahía de Zihuatanejo, costas adyacentes e Isla Ixtapa, Gro., Méx. (Stuardo et al. 1974). Observaciones: La coloración de los juveniles varía presentando ornamentación de color caférojizo en forma de zig-zag, concéntrica, sobre fondo blanco. Keen (1971) reporta ejemplares de 112 mm de largo; en este estudio el ejemplar más grande encontrado fue de 87.6 mm. Megapitaria squalida. (Sowerby, 1835). Lámina 1 c y d. Descripción: Concha gruesa porcelanosa, lisa, de forma triangular, alargada anteroposteriormente. Perióstraco grueso. Ligamento externo, situado sobre una plataforma ligeramente incrustada en la parte posterior de la charnela. Charnela fuerte con tres dientes cardinales y dientes anteriores muy marcados (heterodonta). Línea paleal bien marcada; seno paleal profundo en forma de lengua, llegando hasta la mitad de la concha; ligeramente inclinado. Huellas de los aductores circulares, ligeramente mayor la posterior, junto a la cual se encuentra la huella del retractor posterior; la huella del retractor anteríor ligeramente hundida en la concha. Color de la concha variado, café claro o crema u ornamentado con líneas concéntricas, en zig-zag y líneas radiales de color café obscuro. Perióstraco de color café claro, brillante. El interior de la concha es blanco con un tinte violáceo ligero. Distribución: Ha sido reportada desde laguna Ojo de Liebre (Scammon), Baja California, México a Mancora, Perú (01sson, 1961; Keen 1971) incluyendo la Bahía de Zihuatanejo y castas adyacentes e Isla Ixtapa, Gro., Méx. (Stuardo et al. 1974) Observaciones: Cuando la almeja se encuentra en fondos en que domina el limo toma una coloración negra, la cual desaparece al tenerlas en agua limpia. En los ejemplares jóvenes predomina la coloración en zig-zag o de líneas radiales, la cual se mantiene en la parte dorsal de los adultos. Keen (1971) reporta ejemplares de 120 mm de longitud; en este estudio el ejemplar más grande encontrado fue de 72.4 mm de largo. Género DosiniaDosinia ponderosa (Gray 1838) Lámina 1. e y f. Descripción: Concha de forma circular o lenticular con escultura concéntrica bien marcada. Perióstraco muy delgado y brillante. Ligamento externo situado sobre una plataforma incrustada en la parte posterodorsal de la charnela, quedando parcialmente oculto por la concha. Lúnula cardiforme bien marcada. Charnela con tres dientes cardinales, más cortos y gruesos en la valva derecha; a cada diente corresponde un cóndilo en la valva opuesta; la valva izquierda presenta un diente lateral anterior muy reducido (heterodonta). Línea paleal bien marcada; seno paleal triangular, inclinado diagonalmente, llegando hasta el centro de la concha. Huellas de los músculos aductores ligeramente elípticas, mayor la posterior, junto a la cual se encuentra la huella del retractor posterior. La huella del retractor anterior se observa en la parte interna anterior de la charnela. Concha de color blanco translúcida, con franjas concéntricas de tonos diferentes; interior blanco. Distribución: Se ha reportado entre Laguna Ojo de Liebre (Scammon) Baja California, México a Paita, Perú (Olsson 1961; Keen, 1971) incluyendo la Bahía de Zihuatanejo (Stuardo et al. 1974). Keen (1971) reporta el ejemplar más grande de 145 mm de largo; en este estudio el ejemplar mayor encontrado fue de 93.2 mm. ASPECTOS HIDROLÓGICOSEn la tabla 1 y en la figura 1 se presentan las variaciones de temperatura y salinidad observadas en la Bahía de Zihuatanejo e Isla Ixtapa durante un año completo. Su comparación con lo reportado por Tovar y Sánchez (1974) aparece en la tabla 1 y se ilustra en las figuras 8a. , 9a. , y 10a. En la figura 1 se observa que la temperatura promedio descendió de 29ºC en septiembre de 1974 a 25.5ºC en marzo de 1975, elevándose hasta 28.5ºC en junio y volviendo a 29ºC en el mes de julio; bajó nuevamente en agosto a 28.5ºC aumentando hasta 30°C en el mes de septiembre de 1975. La salinidad promedio se comporta en forma contraria a la temperatura. Entre septiembre y diciembre de 1974 ascendió de 32 % a 33.8 %; mientras que entre diciembre de 1974 y marzo de 1975 descendió junto con la temperatura hasta 32.5 % y luego subió a 33.5 %. en el mes de mayo, manteniéndose así durante el mes de junio, para bajar en julio y agosto durante el periodo de lluvia llegando a un mínimo de 31.3 %; con posterioridad ascendió nuevamente a 33.1 % en septiembre de 1975. El valor máximo registrado para la salinidad fue de 34.35 % para el mes de diciembre en Isla Ixtapa y el mínimo, también para la misma localidad, igual a 3l %, en el mes de agosto. 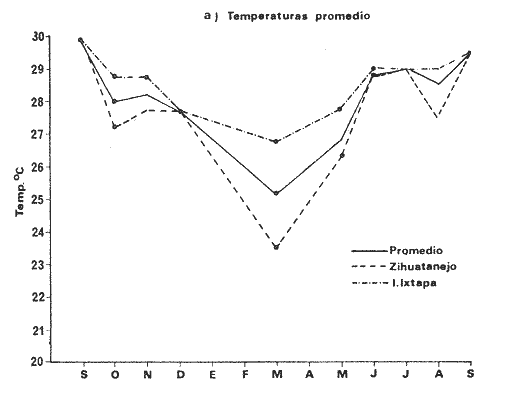 Fig. 1 Variaciones promedio y medias mensuales de temperatura (a) y salinidad (b) en el área de estudio entre septiembre de 1974 y septiembre de 1975 (detalle de valores en Tabla 1). 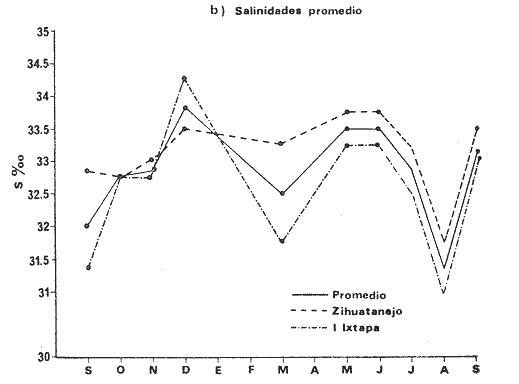 continuación En la misma figura se constata que las variaciones de temperatura entre Zihuatanejo e Ixtapa son similares, pero los rangos de variación son muchos mayores en Zihuatanejo, en donde se observaron rangos entre 23.5ºC y 29.8ºC, no así en Isla Ixtapa, en donde el rango de variación fue sólo de 27.75ºC a 29.8ºC. La salinidad se constata más estable en Zihuatanejo sufriendo una caída extrema en el mes de agosto, con valores que fluctúan entre 31.75 % y 33.75 % rango que para Isla Ixtapa varía entre 31.0 % y 34.35 %. DISTRIBUCIÓN DE LOS BANCOS EN EL ÁREA DE ESTUDIO
Bahía de ZihuatanejoSe localizaron 4 bancos principales en esta bahía (Mapa 2a). Los bancos 1 y 2 ubicados casi al centro de la bahía entre 4 y 12 m de profundidad, están formados por las especies Megapitaria squalida y Dosinia ponderosa. El fondo es de arena con limo. Los bancos 3 y 4 se ubican en la entrada de la bahía y en su parte norte interna a poca distancia de la costa, respectivamente. Ambos están formados por la especie Megapitaria aurantiaca, aunque en el banco 3 ésta se halla asociada con la "almeja roñosa" Ventricolaria iscordia. Este banco se encuentra en la parte más profunda de la bahía, entre 14 y 18 m, en un canal con fondo de arena gruesa con algunos cantos, donde la corriente es fuerte. El banco 4 se encuentra a profundidades que varían entre 5 y 10 m, sobre un fondo de rocas con arena gruesa y limo. Aquí M. aurantiaca no es muy abundante pero se halla asociada con diversas especies de almejas. Isla IxtapaEn los fondos estudiados alrededor de la isla se localizaron también cuatro bancos importantes (Mapa 3a); los bancos 1 y 4 localizados cerca de la isla y los bancos 2 y 3 en el canal que la separa de la costa. Los bancos 1 y 2 están ubicados en fondos de arena fina entre 5 y 12 metros de profundidad, separados por una franja de arena fina y limo. El banco 1 incluye sólo a la especie Megapitaria squalida; el banco 2 está formado principalmente por M. squalida y secundariamente por Dosinia ponderosa, así como por otras especies de almejas menos abundantes, como se menciona en fauna asociada. 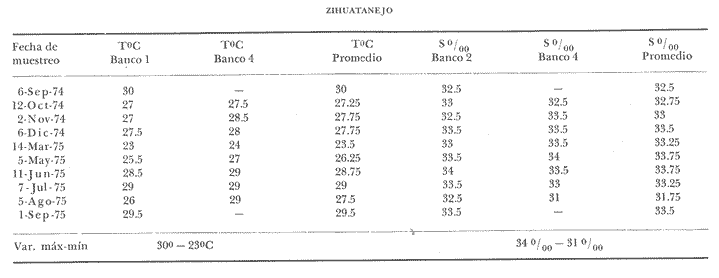 TABLA 1 TEMPERATURA Y SALINIDAD EN LOS BANCOS DEESTUDIO ZIHUATANEJO 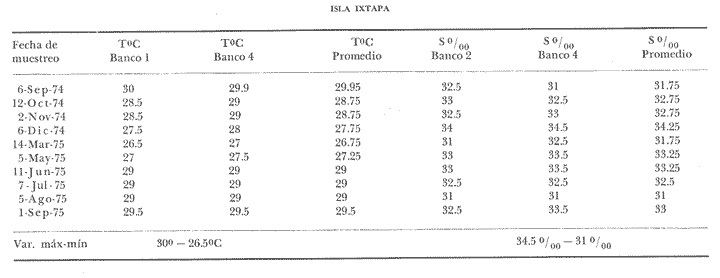 ISLA IXTAPA 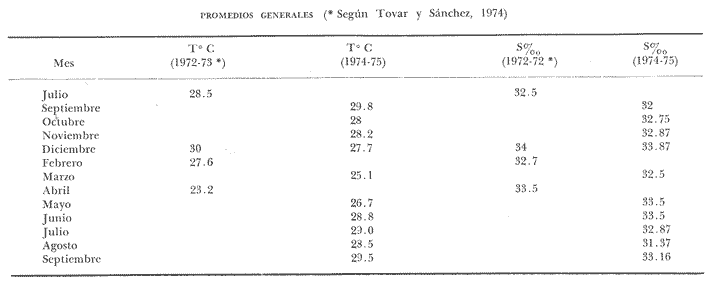 PROMEDIOS GENERALES (* Según Tovar ySánchez, 1974) 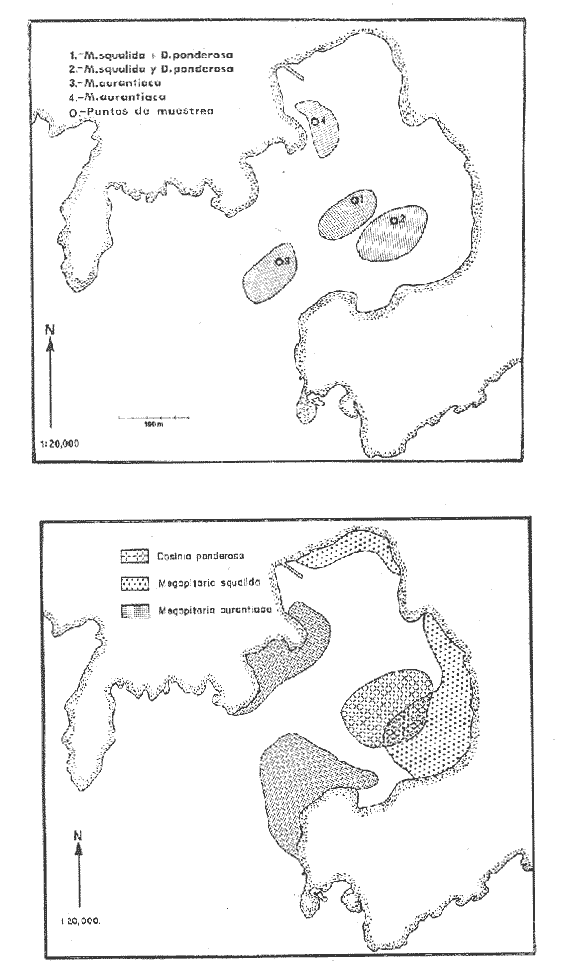 Mapa 2. Bahía de Zihuatanejo. a) Localización de los bancos de "almeja". b) Distribución de M. aurantiaca, M. Squalida y D. ponderosa. El banco 3 se encuentra en una zona de mucha corriente con influencia del oleaje, en fondo de arena gruesa, a profundidad de 6 a 8 metros. Está formado principalmente por Megapitaria aurantiaca. El banco 4 se halla frente a la costa, en profundidades que van de 5 a 8 metros, en fondo de rocas y arena gruesa. Además de M. aurantiaca, especie que predomina, se encuentran algunos ejemplares de M. squalida, y un gran número de otras especies, La localización y delimitación de estos bancos corresponden a los puntos más conocidos por los pescadores y de mayor abundancia de las diversas, almejas", pero su distribución en ambas localidades es más amplia. En lo que respecta a la "almeja roja" (Megapitaria aurantiaca) , es posible agregar que se encuentra en mayor o menor cantidad en todas las zonas con fondo de arena gruesa y donde la acción de las olas no es muy fuerte. Como ya se mencionó, la distribución de la "almeja negra"(Megapitaria squalida) y la "almeja blanca" (Dosinia ponderosa) está dada por el tipo de substrato, siendo éste muy afín para las dos especies, por lo que en la mayoría de los casos, los bancos de ambas se encuentran superpuestos. En los mapas 2a y 3a, se representa la localización de los bancos estudiados, y en los mapas 2b y 3b la distribución general de las tres especies en la zona de estudio. 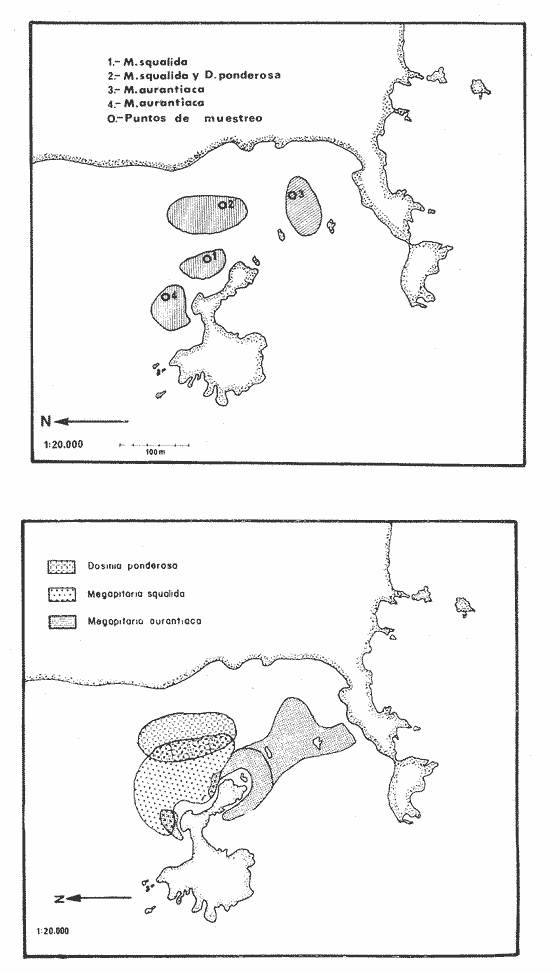 Mapa 3. Isla Ixtapa.a) Localización de los bancos de almeja.b) Distribución de M. aurantica, M. squalidad y D. poderosa ABUNDANCIA Y ESTRUCTURA DE LAS POBLACIONESLa información por especies aquí detallada incluye características de tamaño y peso de cada una de las especies estudiadas en muestras combinadas de las poblaciones provenientes de los bancos 1 y 4 de la Bahía de Zihuatanejo y 2 y 4 de Isla Ixtapa. La abundancia corresponde a muestras obtenidas en 1 m² de fondo en los bancos correspondientes. 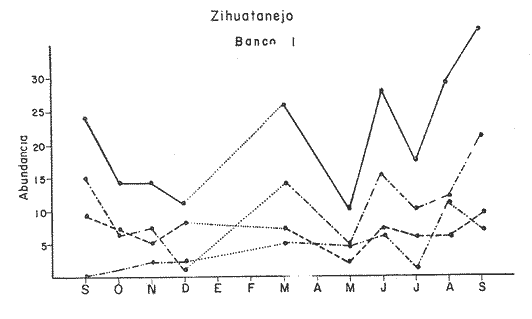 Fig. 2. abundancia comparada de M. aurantica, M. squalidad y D. ponderosapor unidad de área (m2) en los bancos 1 y 4 de Zihuatanejo. 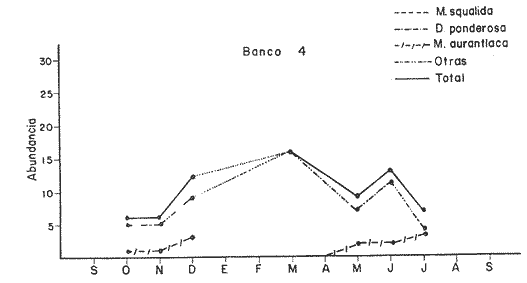 Fig. 2. abundancia comparada de M. aurantica, M. squalidad y D. ponderosapor unidad de área (m2) en los bancos 1 y 4 de Zihuatanejo. 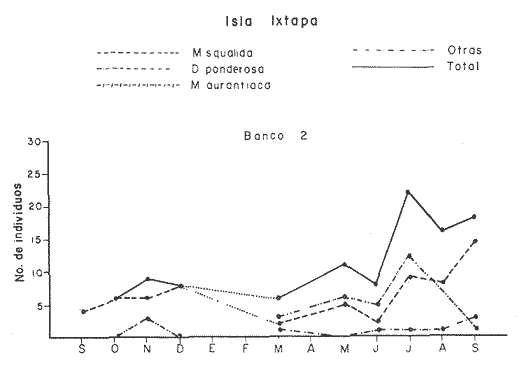 Fig. 3 Abundancia comparada de M. aurantiaca, M. squalida y D. ponderosa por unidad de área (m2) en los bancos 2 y 4 de Isla Ixtapa 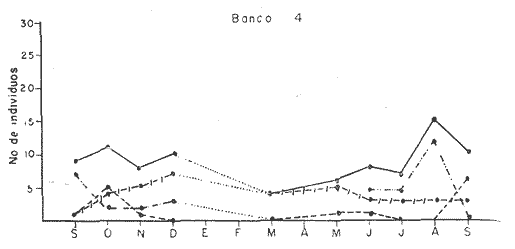 Fig. 3 Abundancia comparada de M. aurantiaca, M. squalida y D. ponderosa por unidad de área (m2) en los bancos 2 y 4 de Isla Ixtapa
Fig. 4 Fig. 5 Fig. 6 Megapitaria aurantiacaLas observaciones realizadas sobre la estructura de la población de esta especie se presentan en las figuras 4a, b, c y en las tablas 3a, b, c, d y e. Su abundancia comparada a la de otras especies de bivalvos se ilustra en las figuras 2 y 3 y tablas 2a y b. a) LargoEn la figura 4a es posible observar que los tamaños máximos alcanzados por los ejemplares de esta población no pasan de la clase de 90 mm en los meses de marzo y mayo. Del análisis gráfico resulta un desplazamiento de la moda hacia la derecha entre los meses de septiembre a noviembre de 1974, lo que indica un crecimiento mensual de aproximadamente 5 mm. En diciembre se desplaza la moda hacia la izquierda por el ingreso de ejemplares juveniles; en marzo de 1975 la moda vuelve a desplazarse considerablemente hacia la derecha. En mayo 1975 se constata una población muy heterogénea en la que se pueden distinguir tres modas dadas por el ingreso a la población de juveniles, productos del intenso desove de los meses anteriores, y otras dos de adultos. 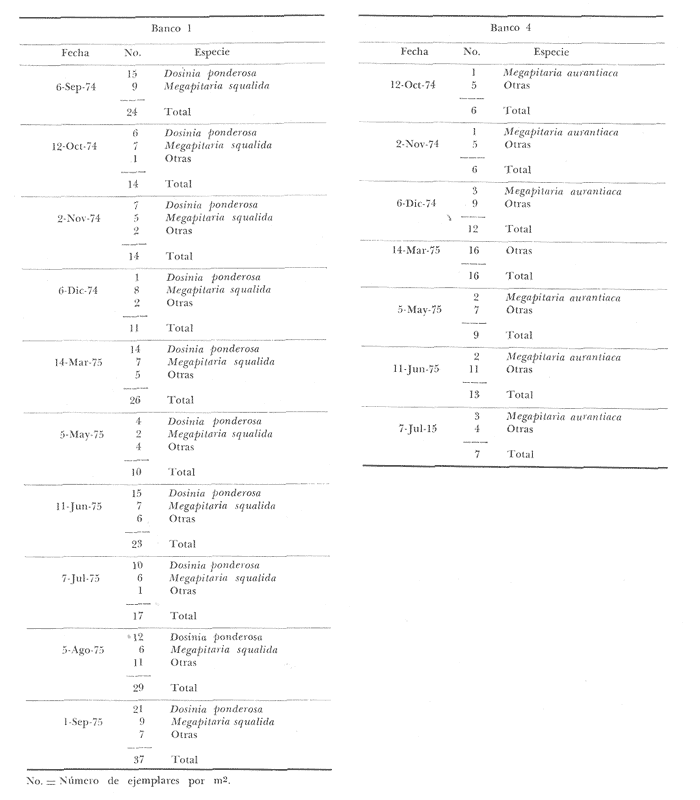 TABLA 2a ABUNDANCI RELATIVA DE LAS ESPECIES ESTUDIADAS ZIHUATANEJO 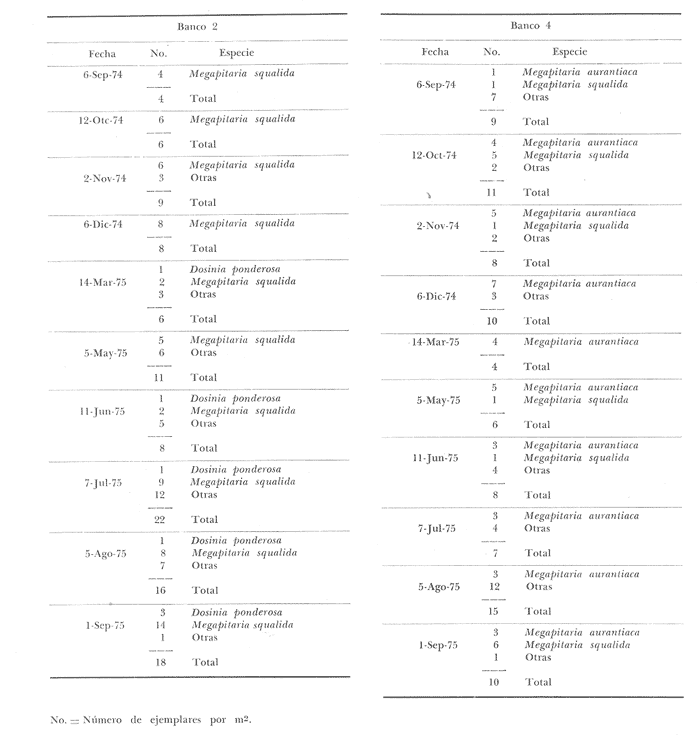 TABLA 2 b ABUNDANCIA RELATIVA DE LAS ESPECIES ESTUDIADAS I S L A I X T A P A En los meses siguientes la población va haciéndose más homogénea en torno a la clase de 40-45 mm que se hace más notoria mes a mes para volver a presentar en noviembre de 1975 una curva de características muy similares a la del año anterior. El tamaño mínimo encontrado fue de 10.6 mm en el mes de mayo y el máximo de 87.6 mm en el mes de marzo. Las clases sombreadas corresponden a grupos de edades que se desplazan notoriamente hacia la derecha (Fig. 4a), indicando el crecimiento alcanzado. b) AltoEn la figura 4b se observa el comportamiento de la población en relación al alto de la concha. En general es paralelo al del largo, encontrándose solamente un poco más marcadas las modas de las diferentes clases de edades probables. Las clases dominantes son las de 30-35 mm y 45-50 mm (de alto), fluctuando las medidas entre 2540 mm; el máximo observado fue de 72.85 mm para el mes de mayo y el mínimo de 8.6 mm para el mismo mes. c) AnchoLa variación observada en este carácter no ha sido representada en la figura 4, pero, los valores de la tabla 3c indican fluctuaciones similares a las del largo y alto. El número de clases resultante es menor como reflejo del crecimiento más lento de este parámetro, razón por la cual, el rango de fluctuación es menor, presentándose el máximo de 46.7 mm en el mes de marzo y el mínimo de 5.8 mm en el mes de mayo. d) Peso totalEl comportamiento de la curva de peso (Fig. 4c) sigue en cierta medida a la de crecimiento, encontrándose un desplazamiento constante de la moda de 10-30 g hacia la derecha desde septiembre hasta el mes de noviembre en que llega a la clase de 20-30 g con una regresión en el mes de diciembre de 1974 a la de 10-20 g. Luego, el peso aumenta nuevamente dominando porcentualmente las clases más elevadas para volver a bajar hasta la de 020 g en agosto, que se hace exageradamente dominante en el mes de septiembre de 1975. Resulta de interés constatar la semejanza de esta curva con la de septiembre de 1974. El peso mínimo registrado fue de 0.5 g en el mes de mayo y el máximo de 156 g para el mes de marzo. e) Peso sin conchaLa variación en el peso de las partes blandas tampoco ha sido incluida en la figura 4, pero se resume en la tabla 3e. Aún cuando los rangos de clases son diferentes, presenta las mismas variaciones que el peso total, sufriendo sólo una reducción en la frecuencia de las clases mayores. El peso máximo, de 41.4 g, fue encontrado en el mes de mayo y el mínimo, de 0.3 g, en los meses de mayo y septiembre de 1975. d) AbundanciaLa abundancia de esta especie medida por metro cuadrado y representada en las figuras 2 y 3, fluctúa grandemente en los bancos de estudio (banco 4 en Zihuatanejo y banco 4 en Isla Ixtapa). Se observa que la densidad aumenta en los dos bancos hacia el mes de diciembre, coincidiendo con el ingreso a la población de juveniles (Fig. 4), densidad que para Isla Ixtapa se mantiene más o menos constante durante los meses de marzo a mayo de 1975, cayendo ligeramente en el mes de junio para mantenerse estable hasta el mes de diciembre. En Isla Ixtapa durante los meses de septiembre a diciembre, el aumento en la densidad de M. aurantiaca corresponde a la disminución de otras especies, no así en los meses de junio a septiembre de 1975 en que su densidad se mantiene constante independientemente del aumento en densidad de otras especies. El banco 4 de Zihuatanejo sólo se reporta hasta el mes de julio de 1975, a partir del cual no se encontró ninguna almeja viva por efecto de un intenso acarreo de sedimentos finos que cubrieron el área a causa del dragado de la boca del estero cercano. Presentó dos aumentos bien marcados desde octubre a diciembre de 1974 y entre abril y julio de 1975. En este último banco, la densidad varió entre 1 y 3 ejemplares por m2, mientras que en el banco 4 de Isla Ixtapa la densidad varió de 1 a 7 ejemplares por M2 con un promedio de 4 ejemplares por M2. Megapitaria squalidaEn las figuras 5a, b y c y en las tablas 4a, b, c, d y e, se representa la estructura de la población de esta especie para la zona de estudio. Su abundancia comparada con otras especies se ilustra en las figuras 2 y 3 y en las tablas 2a y b. a) LargoEn la figura 5a, se constata que el largo máximo de la población no pasa de la clase de 70-75 mm con modas que fluctúan entre 35 y 60 mm. En el periodo comprendido entre septiembre de 1974 y marzo de 1975 se observa un desplazamiento gradual de las clases pequeñas (15-20 mm) hacia las clases de 5060 mm, con un aumento notorio de éstas; en el mes de diciembre de 1974, se observa la aparición de juveniles que se hacen también aparentes en el mes de marzo de 1975, y especialmente en mayo lo que corresponde a un intenso desove que se efectúa a partir de octubre (Fig. 9). En el mes de mayo de 1975 se tiene una clara separación de tres clases de edades con modas de 15-20, 34-40 y 55-60 mm; en junio la moda de 15-20 mm se ha sumado a la de 3540 mm. En julio la moda dominante del mes anterior sufre un desplazamiento hacia la derecha, pero aparece otra moda que agrupa un mayor número de ejemplares, producto del desove total que sufrió la población en los meses pasados (Fig. 9b). En los meses de agosto y septiembre la moda se desplaza nuevamente hacia la derecha, dominando la clase de 40-45 mm. El largo máximo registrado fue de 72.4 mm en el mes de septiembre de 1974 y el mínimo de 11.5 mm en mayo de 1975. Las clases sombreadas comprenden a clases de edades que se desplazan hacia la derecha (Fig. 5a) b) AltoNuevamente en esta especie es notorio el paralelismo existente entre el largo y el alto (Fig. 5b), haciéndose igualmente notoria la separación de grupos o clases de edades, como es posible constatar en los meses de diciembre de 1974, marzo, mayo, junio y julio de 1975, en que se pueden observar clases separadas o la dominancia de dos o más modas. En esta especie no se observa separación entre las clases de mayor tamaño y las más pequeñas como en el caso de M. aurantiaca. La clase dominante es la de 40-45 mm, la cual se desplaza entre septiembre y diciembre de 1974 a la de 45-50 mm; en julio de 1975 la clase que domina es la de 25-30 mm, la cual se va desplazando hasta 3540 mm en el mes de septiembre de 1975. El alto máximo registrado fue de 60.9 mm en el mes de septiembre y el mínimo de 9.5 mm en mayo, correspondiendo a los máximos y mínimos del largo. c) AnchoAl igual que en la especie anterior, la variación del ancho no se ha incluido en la figura 5, pero se presenta en la tabla 4c. Fluctúa en forma paralela al largo y alto, con un número menor de clases por lo que las clases dominantes son más marcadas, acentuándose así las variaciones de la curva. El valor máximo encontrado fue de 39.3 mm en el mes de septiembre de 1974; el mínimo de 5.1 mm en septiembre de 1975; de igual forma el rango de fluctuación de los valores es menor que en el largo y alto. d) Peso totalComo en el caso de M. aurantiaca, la curva de peso (Fig. 5c) sigue un desplazamiento correspondiente al crecimiento. Nótese sin embargo, que el rango de peso para el mes de septiembre de 1974 (0-95 g), es mayor que en los muestreos posteriores, hasta los meses de junio y agosto de 1975, en que nuevamente aparece la clase de 75-80 g, pese a que los anteriores incluyen ejemplares de tamaño máximo similares. Esta diferencia se explica por el mayor porcentaje de ejemplares no desovados en estos meses, lo que es muy claro en los meses de julio, agosto y septiembre (de 1975), en que la curva de peso se desplaza totalmente hacia la izquierda en parte porque correspondió a un periodo de indiferenciación gonádica de la población (Fig.9) y por el ingreso masivo de juveniles. 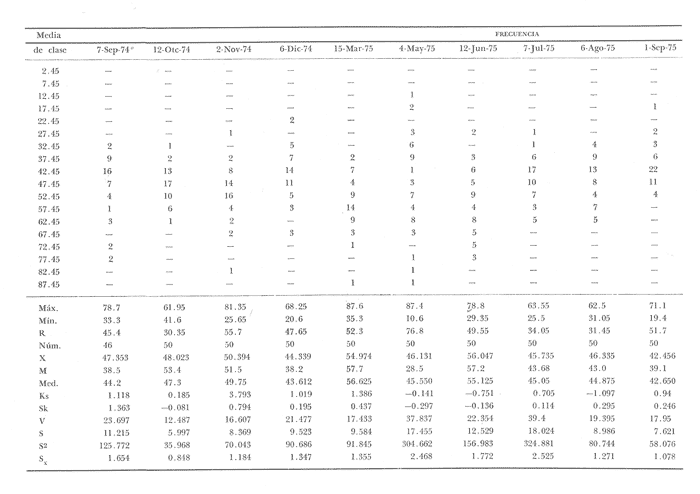 TABLA 3a ESTRUCTURA DE POBLACIONES Mega pitaria aurantiaca (Largo) 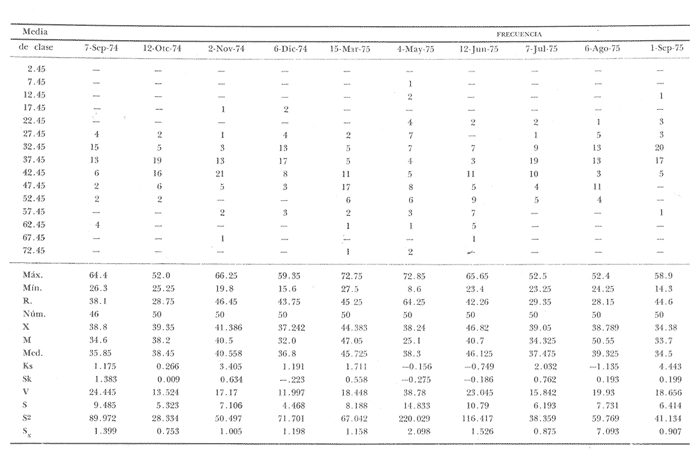 TABLA 3 b ESTRUCTURA DE POBLACIONES Megapiataria aurantiaca (Alto) 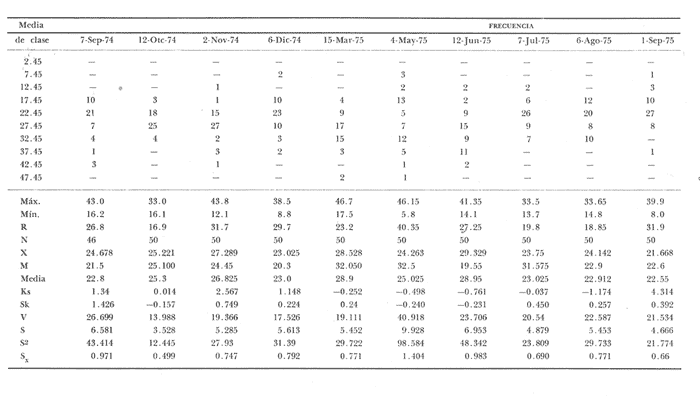 TABLA 3c ESTRUCTUIRA DE POBLACIONES Megapiatara aurantiaca (Ancho) 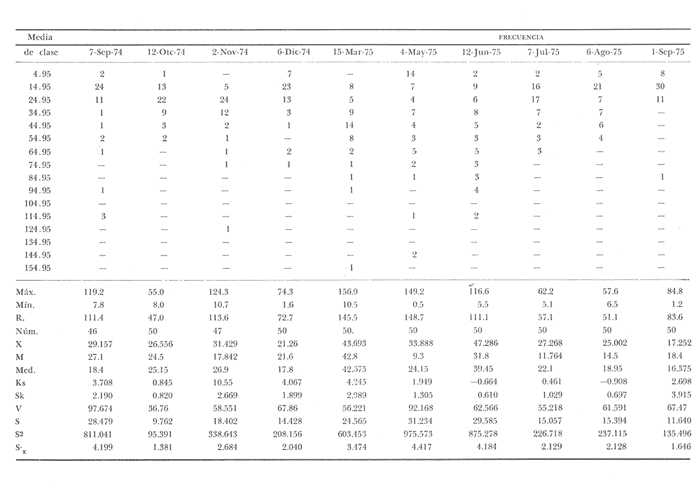 TABLA 3 d ESTRUCTURA DE POBLACIONES Megapiataria aurantica (Peso total) 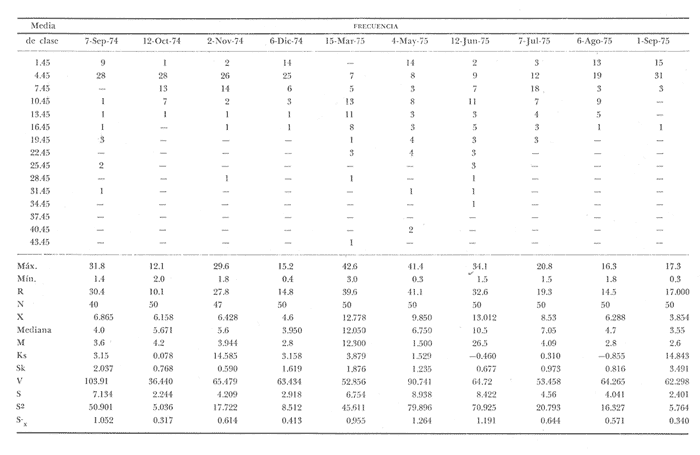 TABLA 3e ESTRUCTURA DE POBLACIONES Megapiataria aurantica ( Peso sin concha) e) Peso sin conchaLos valores agrupados del peso de las partes blandas se presentan en la tabla 4e y no se incluyen en la figura 5. Muestran gran paralelismo con el peso total, reduciéndose la frecuencia de las clases mayores, en la misma forma que en la especie anterior. El peso máximo, de 41.5 g, fue registrado en el mes de junio y el mínimo de 0.2 g en septiembre de 1975. f) AbundanciaLa abundancia de esta especie por metro cuadrado (Figs. 2 y 3) ha variado en los bancos estudiados (banco 1 en Zihuatanejo y 2 y 4 de Isla Ixtapa). En todos los bancos estudiados se observan fluctuaciones similares, teniendo aumentos en densidad en los meses de diciembre de 1974 y septiembre de 1975 que corresponden a la aparición de juveniles. Al comparar estas curvas con la gráfica de madurez gonádica (Fig. 9) se ve que en ambos casos coincide con un desove masivo ocurrido dos meses antes. En el banco 1 de Zihuatanejo la densidad varió entre 2 y 9 ejemplares por metro cuadrado con un promedio de 6 ejemplares; en este banco M. squalida aparece como especie acompañante de la "almeja blanca" (Dosinia ponderosa) que es la especie dominante. En el banco 2 de Isla Ixtapa la densidad varió entre 2 y 9 ejemplares por m2, con promedio de 6 ejemplares, siendo aquí la especie dominante. En el banco 4 de Isla Ixtapa su abundancia varió de 1 a 6 ejemplares por M2 apareciendo como especie acompañante de Megapitaria aurantiaca. Dosinia ponderosaLas observaciones para la 'estructura de la población de esta especie se presentan en las figuras 6a, b y c y en las tablas 5a, b, c, d y e. Su abundancia comparada a la de otras especies de bivalvos, se da en las figuras 2 y 3 y en las tablas 2a y b. a) LargoEn la figura 6a se muestra que el largo de la población no excede de la clase de 90-95 mm, con un mínimo de 18.2 mm en el mes de julio y un máximo de 93.2 mm en mayo. La moda varía entre 55 y 70 mm entre los meses de septiembre de 1974 y marzo de 1975, pero en el mes de mayo es hace muy notoria la moda de 65-70 mm y aparece una clase pequeña de juveniles, la cual se va desplazando hacia la derecha en los meses siguientes, con un claro incremento mensual de 5 mm en pro medio. En general, entre los meses de mayo y septiembre, se nota un gran incremento de juveniles en la población. El incremento de juveniles en el mes de mayo corresponde a un intenso desove en el mes de marzo (Fig. 1 Ob) b) AltoPor la forma de la concha, el alto (Fig. 6b) se incrementa paralelamente con el largo, por lo que las curvas son muy semejantes. Es importante mencionar que en las curvas de altura es más notorio el desplazamiento de la moda de las clases de 50-55 a 60-65 mm entre los meses de marzo a junio de 1975, observándose también la aparición de juveniles en los meses de mayo y julio. Relacionando el estudio de la madurez gonádica (Fig. 10b) con la presencia de juveniles, se constata que corresponden a desoves durante los meses anteriores a mayo y junio. El alto máximo registrado fue de - 38.2 mm en el mes de mayo y el mínimo de 17.3 mm en julio. Las clases sombreadas corresponden a grupos de edad o clases de edad que se van desplazando hacia la derecha, observándose en este caso la presencia al menos de tres clases de edad durante el año. 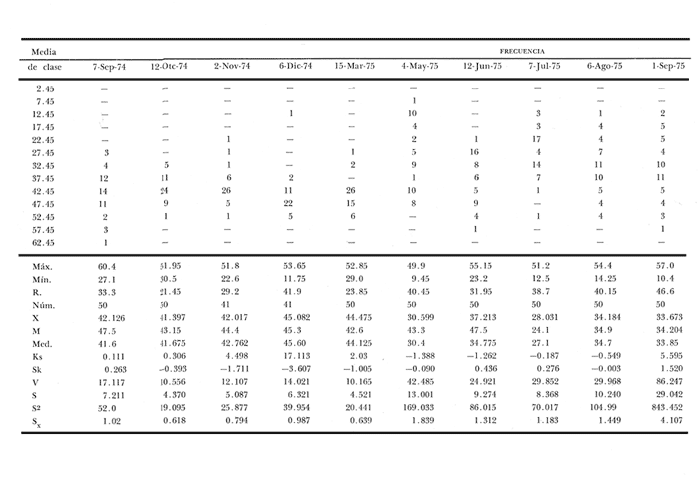 TABLA 4a ESTRUCTURA DE POBLACIONES Megapitaria squalida (Largo) 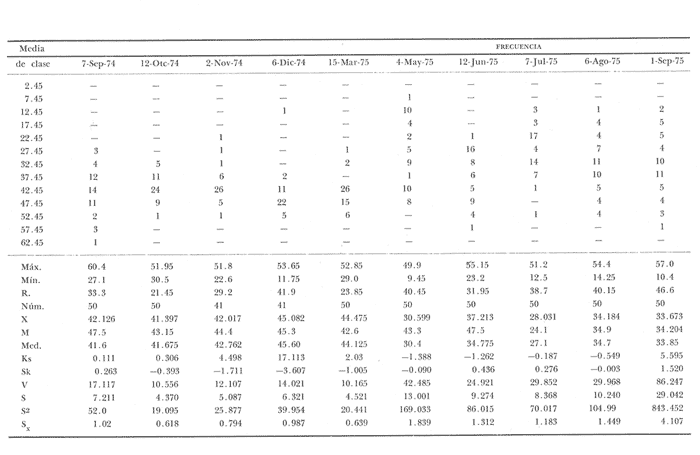 TABLA 4b ESTRUCTURA DEPOBLACIONES Megapitaria squalida (Alto) 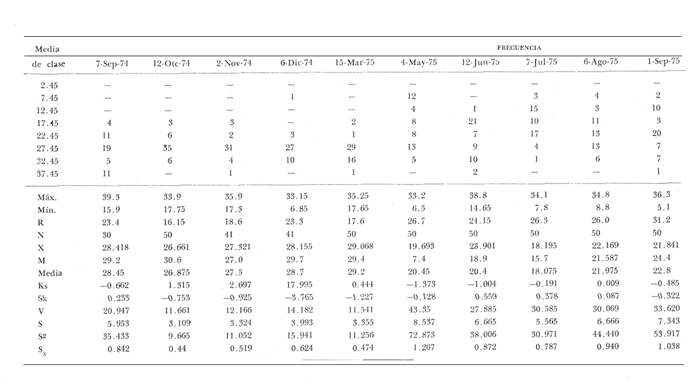 TABLA 4c ESTRUCTURA DE POBLACIONES Megapiataria squalida (Ancho) 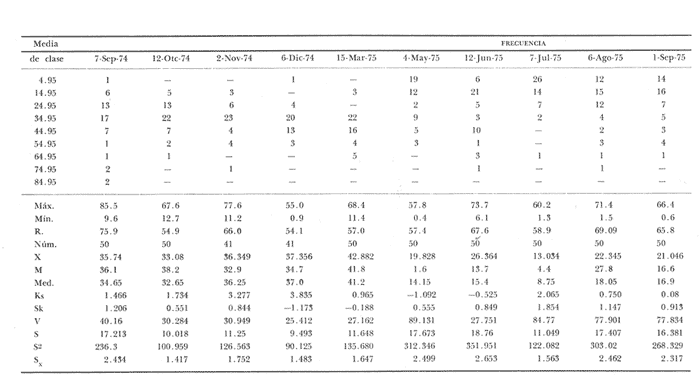 TABLA 4d ESTRUCTURA DE POBLACIONES Megapiataria squalida (Peso total) 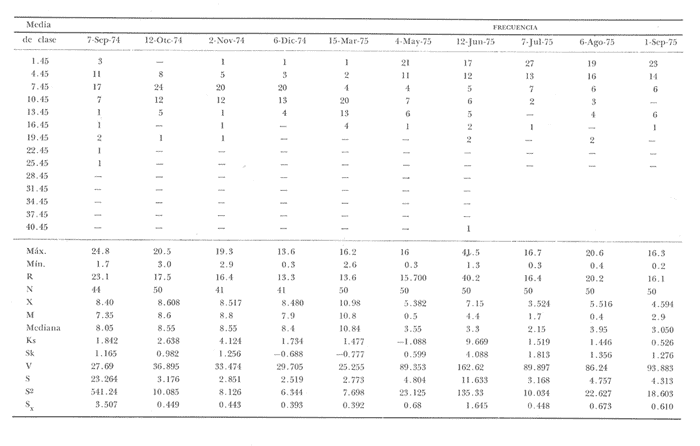 TABLA 4e ESTRUCTURA DE POBLACIONES Megapiataria squalida (Peso sin concha) c) AnchoLos valores respectivos no se presentan en la figura 6 que ilustra la variación de las otras medidas, pero su agrupación se incluye en la tabla 5c. Obviamente, el crecimiento en el ancho es lento lo que determina no sólo un rango de variación estrecho, sino también un número reducido de clases. El valor máximo medido igual a 45.2 mm, se obtuvo en el mes de mayo y el mínimo de 7.7 mm, en julio. d) PesoEn la curva de peso (Fig. 6c) no es posible diferenciar clases dominantes cuyo desplazamiento indique un aumento relacionado con el crecimiento de tamaño de los individuos en la población. En algunos meses se observa un aumento en las clases de mayor peso (70-180 g), pero consecuencia de muestras que incluyen ejemplares de mayor longitud. A partir de la aparición de juveniles en el mes de mayo y junio de 1975 (y aparentemente antes de septiembre de 1974) es proporcionalmente mayor el número de ejemplares agrupados en las clases de más de 80 g que en los otros meses, resultado del desplazamiento de clases menores hacia las clases superiores. El peso máximo registrado fue de 179.4 g para el mes de mayo y el mínimo de 1.1 g para julio, valores que corresponden a los máximos y mínimos de altura y longitud. e) Peso sin conchaComo en el caso de las otras especies las fluctuaciones del peso de las partes blandas son aquí similares a las del peso total, constatándose un paralelismo más marcado, probablemente a causa de una concha más delgada. (Tabla 5e) El valor máximo igual a 57.6 g se registró en el mes de mayo y el mínimo de 0.4 g en julio. f) AbundanciaEsta especie se ha encontrado abundante en el banco 1 de Zihuatanejo, ya que en Isla Ixtapa es una especie acompañante de M. squalida. En el banco 1 de Zihuatanejo varía entre 1 y 21 ejemplares por m2 con un promedio de 10 ejemplares (Fig. 2). Su abundancia fluctúa conjuntamente con las otras especies acompañantes, de las que Megapitaria squalida es la segunda en importancia después de Dosinia ponderosa. ANÁLISIS DE CRECIMIENTOEn la tabla 6a y b y figura 7b, se constata que para M. aurantiaca, el incremento mensual medio aumenta desde las clases menores hacia las tallas medias, registrándose el incremento máximo (5.54 mm mensuales) en los ejemplares de 65 a 70 mm de largo, volviendo a disminuir en los ejemplares de talla mayor con un incremento mensual de .05 mm para los ejemplares de 90 a 95 mm. En M. squalida se observa que el incremento mensual medio va disminuyendo de los ejemplares menores (25-30 mm) con un incremento mensual de 4.5 mm hasta 0.6 mm en los ejemplares mayores (70-75 mm). Para D. ponderosa se observa un comportamiento similar al de M. squalida, teniendo el máximo incremento de 2.3 mm mensuales en los ejemplares de 40 a 45 mm de largo y el mínimo de 0.28 mm mensuales en los ejemplares de 75 a 80 mm. Las alteraciones en las curvas de M. squalida y D. ponderosa, en que se observa un aumento en el incremento mensual promedio de las clases de 50-55 mm y 55-60 mm respectivamente, así como la gran irregularidad observada en los incrementos registrados por M. aurantiaca, pueden deberse a la tolerancia de los organismos de mayor tamaño a la manipulación y presiones a que estuvieron sometidos durante el experimento. La tabla 6b y figura 7a indican el incremento mensual promedio o ritmo de crecimiento del total de los ejemplares marcados de cada una una de las especies estudiadas. Se determina que las tres especies siguen un patrón similar con un incremento máximo en el mes de junio. Correlacionando esta gráfica con el desarrollo gonádico (Figs. 8, 9 y 10) se observa que a este periodo de incremento máximo corresponde un periodo de actividad gonádica mínima; la temperatura para este periodo fue de 29º C y la salinidad de 32.9%. (Fig. l), lo que puede indicar valores cercanos a los óptimos para el desarrollo de estas especies. 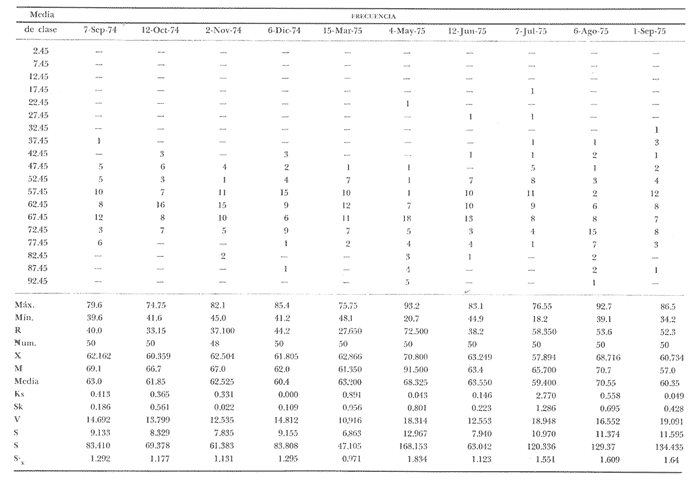 TABLA 5a ESTRUCTURA DE POBLACIONES Dosinia ponderosa (Largo) 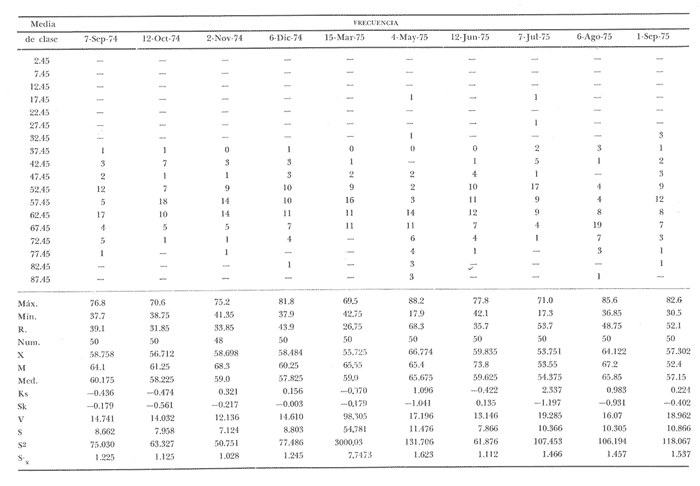 TABLA 5b ESTRUCTURA DE POBLACIONES Dosinia ponderosa (Alto) 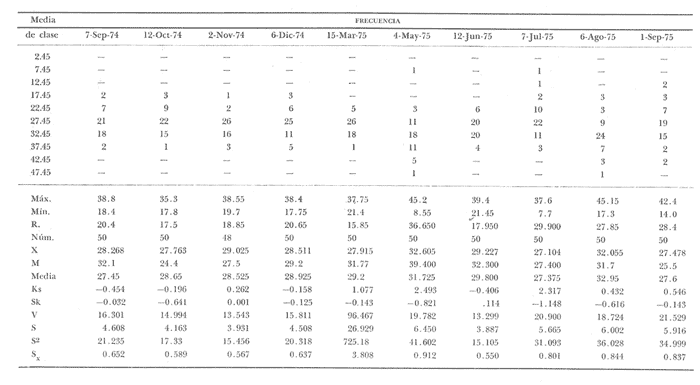 TABLA 5c ESTRUCTURA DE POBLACIONES dosinia ponderosa (Ancho) 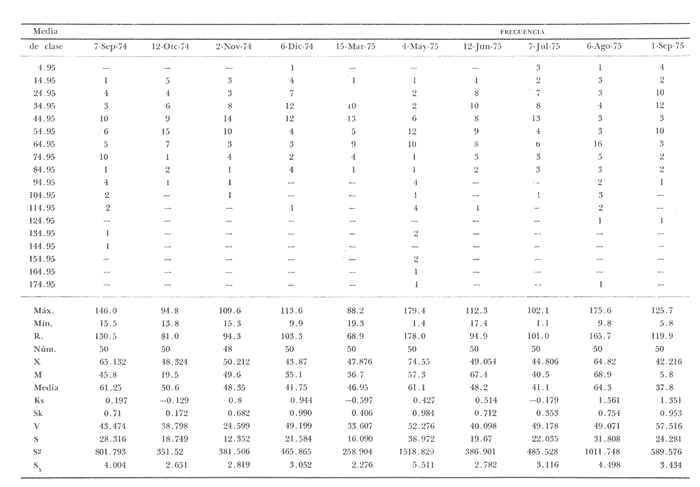 TABLA 5d ESTRUCTURA DE POBLACIONES Dosinia ponderosa (Peso total) 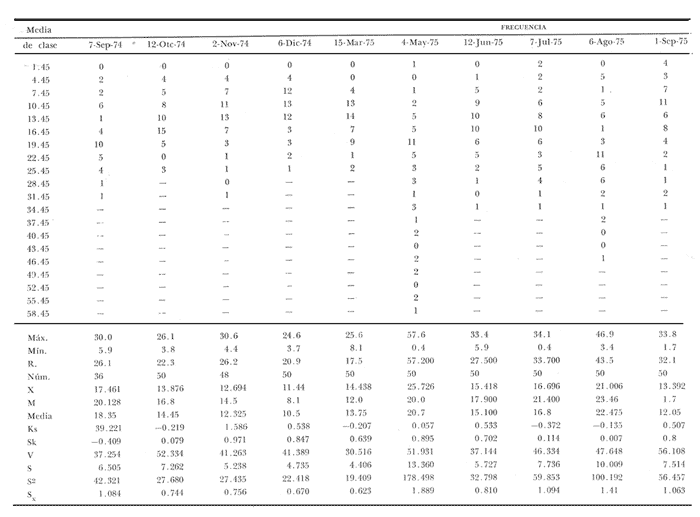 TABLA 5e ESTRUCTURA DE POBLACIONES Dosinia ponderosa (Peso sin concha) Esta estimación de incremento de tamaño es sólo aproximada, ya que en las mediciones se han incluido ejemplares tanto pequeños como grandes en diferentes meses, reconociendo que los ejemplares pequeños crecen más rápidamente que los grandes. 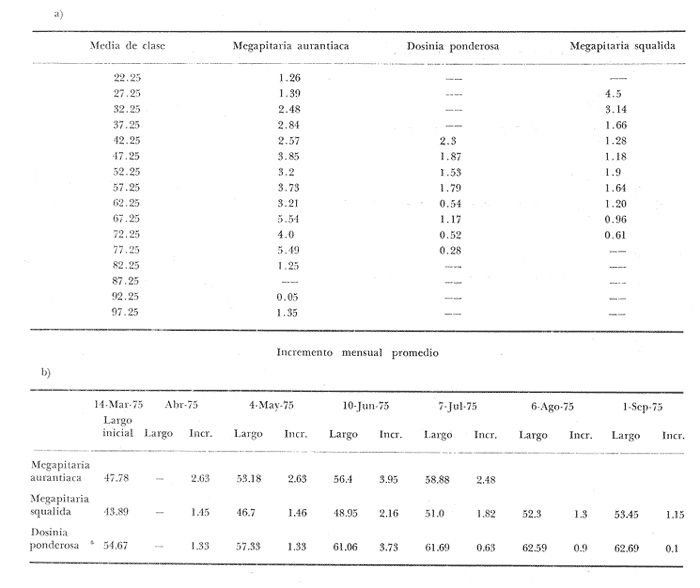 TABLA 6 PROMEDIO ANUAL DE INCREMENTO POR TALLAS DESARROLLO GONÁDICOLas tres especies presentan características semejantes durante el desarrollo gonádico; por lo tanto, la descripción que a continuación se da es aplicable a cualquiera de ellas. El ciclo reproductor se ha dividido en cinco fases que se caracterizan por la estructura de la gónada, cantidad y estado de los gametos, características que se describen a continuación. 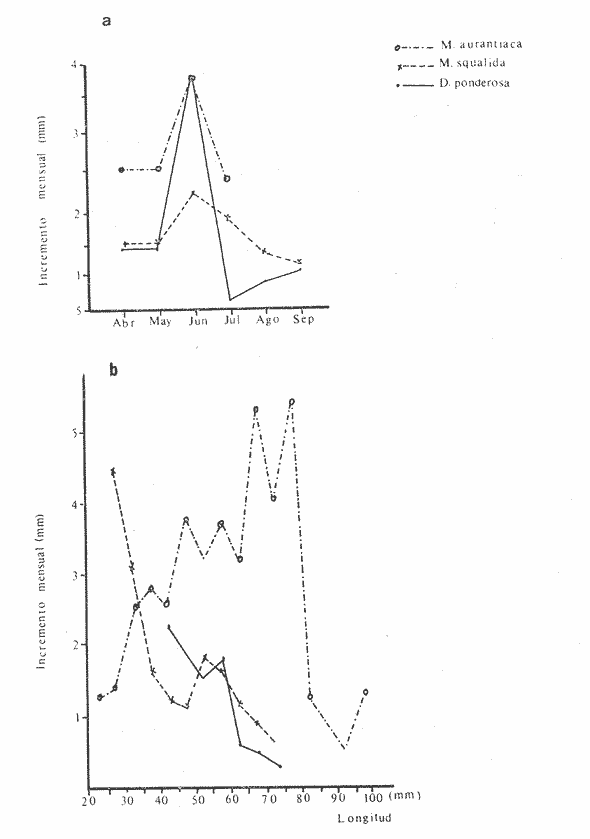 Fig. 7 a) Incremento mensual promedio por especie. b) Promedioanual de incremento mensual por tamaño y especie. FASE I Indiferenciación (Láms. 2a y 3a)También llamada de reposo, se caracteriza por la ausencia total de folículos y gametos. El tejido conectivo ocupa todo el espacio observable entre la glándula digestiva y el tracto digestivo. En sus primeras etapas es posible observar restos de folículos muy destruidos invadidos por fagocitos. FASE II Gametogénesis (Láms. 2b y 3b)Entre el tejido conectivo que llena la cavidad visceral se empiezan a formar los folículos, en cuyo interior es posible observar gran cantidad de células germinales. En el macho los gametos inmaduros llegan a ocupar toda la luz del folículo. En estados avanzados de esta etapa no se observa tejido conectivo o está muy reducido. En las hembras los folículos son más pequeños y están separados por tejido conectivo; los gametos femeninos inmaduros son de forma marcadamente piriforme con la parte angosta unida a la pared del folículo; al centro de éste se van agrupando los huevos maduros. En esta fase las paredes foliculares son más gruesas en los machos que en las hembras. FASE III Madurez (Láms. 2c y 3c)Esta fase corresponde al fin de la gametogénesis e inicio del desove. Por lo general no es posible diferenciarla, ya que el desove se inicia antes de que termine la gametogénesis o ésta se continúa durante el desove. En el macho los folículos se presentan muy aglomerados; no se observa tejido conectivo y sus paredes prácticamente se confunden. En la luz del folículo se aglomeran los haces de espermatozoides. En la hembra esta fase es más marcada, ya que una vez maduros los óvulos, la pared germinal de los folículos se reduce a una capa de células muy delgada. En esta fase los folículos se encuentran llenos de óvulos maduros que, por la aglomeración, presentan forma irregular. FASE IV Desove (Láms. 2d y 3d)Corresponde al periodo de evacuación de los gametos. Las gónadas se observan más o menos vacías dependiendo de lo avanzado del desove; los folículos generalmente se rompen y confunden entre sí. En algunos de ellos la gametogénesis puede continuar mientras que otros están casi completamente desovados. Una vez que la mayoría de los gametos han sido evacuados, el folículo es invadido por fagocitos. Simultáneamente al desove y conforme éste avanza, se va formando el tejido conectivo que inicia su aparición alrededor del tracto, digestivo y la glándula digestiva como un paquete compacto, el cual se va disgregando conforme invade el espacio dejado. por la gónada vacía. FASE V Posdesove (Láms. 2e y 3e)En esta fase la gónada ha evacuado progresivamente la mayoría de los gametos; los folículos muy rotos son invadidos por gran cantidad de fagocitos que destruyen los restos de las paredes de los folículos y los gametos que no hayan sido desalojados durante el desove. El tejido conectivo continúa invadiendo todo el espacio que ha dejado la gónada. ESTUDIO GONÁDICO Y MADUREZ SEXUAL EN LAS ESPECIES ESTUDIADASLos resultados del examen histológico de las gónadas de las tres especies, para el periodo comprendido entre septiembre de 1974 y septiembre de 1975, se presentan en las figuras 8, 9 y 10 y en la tabla 7. Las características histológicas del desarrollo gonádico de las tres especies (M. squalida, M. aurantiaca o D. ponderosa) son muy semejantes, no existiendo diferencia interespecífica alguna entre una fase de madurez, desove o indiferenciación. Para el análisis del comportamiento de la población se hace un análisis por especie. Megapitaria aurantiacaLa figura 8 muestra las características del desarrollo gonádico de esta especie expresadas porcentual e individualmente (Figs. 8b y c) y su relación con la temperatura (Fig. 8a). En la figura 8b los porcentajes están dados por el área cubierta con los símbolos correspondientes. Esta especie presenta un periodo de desove que se prolonga a lo largo de todo el año, con un máximo en los meses de octubre a noviembre de 1974 (y entre febrero y mayo de 1975). Este segundo desove masivo puede ser el más importante, ya que posteriormente aparece un periodo de indiferenciación o reposo gonádico en un 75%, de los ejemplares examinados, quedando algunos ejemplares en gametogénesis activa. La fase de gametogénesis va estrechamente relacionada a los periodos de desove e indiferenciación, ya que con posterioridad a una etapa de indiferenciación sigue la maduración de los gametos y posteriormente el desove, el cual puede ser total seguido por un periodo de posdesove, o puede continuar reiniciando la gametogénesis y el desove. Es posible detectar cierta fluctuación entre los periodos de gametogénesis y desove con las fluctuaciones de la temperatura. En el mes de octubre se observa que cuando la temperatura baja, la gametogénesis disminuye y el desove aumenta; conforme la temperatura sigue descendiendo la gametogénesis aumenta hasta el mes de marzo en que alcanza un 50% de la población y la temperatura su valor mínimo; posteriormente la temperatura aumenta mientras que la gametogénesis disminuye y el desove aumenta, hasta un punto en que toda la población ha desovado y la temperatura se estabiliza. En los primeros meses la relación de sexos es muy variada y no parece guardar relación alguna (Fig. 8c), pero en los meses de diciembre de 1974 y entre mayo y septiembre de 1975 se ve una proporción de 1 a 1. Debe considerarse, sin embargo, que la representatividad de la muestra pudiera ser baja. La talla mínima en que se detectó gametogénesis es de 42.6 mm de largo. Megapitaria squalidaEn la figura 9 se ilustran las características del desarrollo gonádico (Figs. 9b y c) y su relación con la temperatura (Fig. 9a) para el ciclo anual de esta especie, entre septiembre de 1974 y septiembre de 1975. En esta especie, como en la anterior, la época de desove se prolonga durante todo el año con dos periodos de máximo desove: uno en el mes de octubre (87.5%) y otro en mayo (58.3%) coincidiendo ambos con la aparición de juveniles en la clase de 15-20 mm de largo dos meses después (Fig. 5). En esta especie también se observa un periodo de indiferenciación o reposo gonádico en el mes de julio que precede a un prolongado desove. Aun cuando durante este mes un 72% de la población se encuentra en fase indiferenciada, el 28% restante se encuentra en gametogénesis, la cual es constante con un mínimo en octubre en que no se encuentran ejemplares en gametogénesis pura, mas sí combinada con desove (Fig. 9c), alcanzando un máximo (50, 45 y 50%) en los meses de septiembre de 1974, marzo y septiembre de 1975, antes de los periodos de máximo desove. 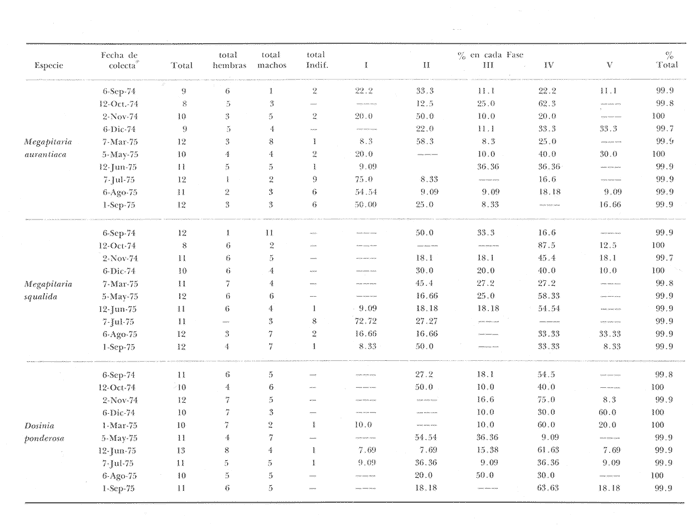 TABLA 7 EJEMPLARES UTILIZADOS EN EL ESTUDIO GONADICO 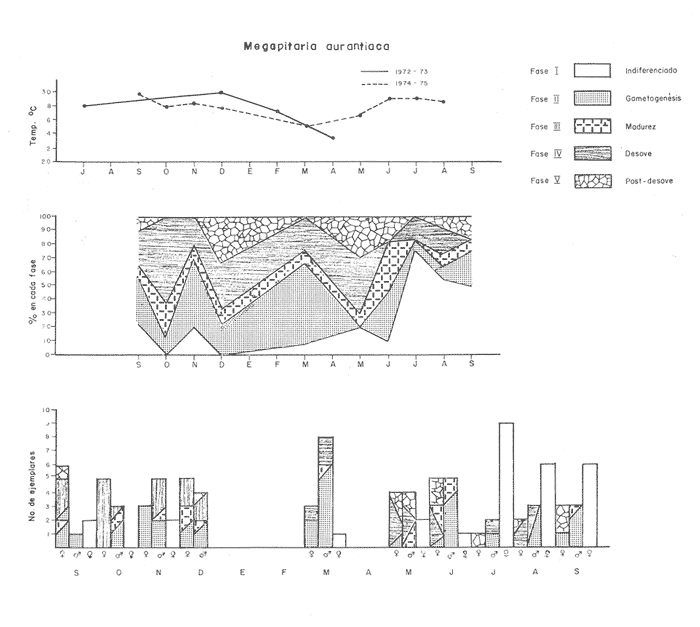 Fig. 8. Análisis cualitativo y porcentual del desarrollo gonádico de las poblaciones combinadas de M. aurantiaca de Zihuatanejo e Isla Ixtapa, durante el ciclo anual comprendido entre septiembre de 1974 y septiembre de 1975 comparado a la variación de temperatura. a) Variación mensual de temperatura; b) Porcentaje mensual de las diferentes fases; c) Análisis individual por sexos. La presencia de ejemplares en posdesove, especialmente en agosto y la ausencia de ejemplares indiferenciados con posterioridad a septiembre, indica que la gónada tiene una gran capacidad de regeneración, manteniendo su actividad todo el año con una sola etapa de reposo, casi general, entre los meses de junio y agosto. No es posible determinar una relación directa entre las variaciones de temperatura y el desarrollo gonádico. La proporción de sexos inferida de la tabla 3 y figura 9c, presenta un número mayor de machos en los meses de septiembre de 1974, julio, agosto y septiembre de 1975, y mayor número de hembras en los meses restantes, a excepción del mes de mayo, en que la relación es de 1 a 1. El promedio total del muestreo de hembras y machos da una relación de 1 a 1, característica que no puede ser demostrada en los muestreos mensuales por lo pequeño de la muestra. La talla mínima en que se detectó gametogénesis es de 41.7 mm de largo. Dosinia ponderosaLa figura 10 muestra las características gonádicas de la población de esta especie para el periodo de septiembre de 1974 a septiembre de 1975 (Figs. 10b y c) y su relación con la temperatura (Fig. 10a). El periodo de desove es aquí mucho más largo que en las anteriores, no pudiéndose determinar un desove preciso, pero sí tres periodos en que la mayoría de la población se encuentra en desove, correspondiente a los meses de noviembre (75%), marzo (60%) y septiembre de 1975 (63.6%). Estos periodos de desove están precedidos por otros de máxima actividad gonádica (gametogénesis) en los meses de octubre (50%), mayo (54%) y julio (36%). La gran cantidad de ejemplares desovados encontrados en el mes de diciembre, indica ya sea que en los meses de enero y febrero pudo haber una mayor cantidad de ejemplares indiferenciados cuyo remanente se presenta como un 10% en marzo, o que esta especie tiene una gran capacidad de regeneración, ya que en marzo sufre un gran desove que no es seguido como en las otras especies por un periodo de indiferenciación. La presencia de gametogénesis en el mes de mayo apoya esta segunda hipótesis. Al igual que en las especies anteriores, no se puede establecer aquí una relación clara entre las variaciones de temperatura y la actividad gonádica, ya que en los meses de septiembre de 1974 y 1975 los máximos de desove corresponden a un máximo de temperatura y el máximo de desove en marzo a un mínimo de temperatura. La relación de sexos para esta especie se desprende de la tabla 10 y figura l0c. Para los meses de julio, agosto y septiembre de 1975 la relación de sexos es de 1 a 1, pero en los otros meses no es posible determinarla, ya que la porporción varía mes a mes. Sin embargo, de la suma total de todas las muestras se obtiene una proporción de 1 a 1. La talla mínima en que se detectó gametogénesis es de 50 mm de largo. ESPECIES ESTUDIADAS Y FAUNA ASOCIADAEntre los aspectos de mayor interés, en relación al tipo de habitat en que viven estas especies, está la determinación de la fauna acompañante. La fauna encontrada es muy raviada estando representada por diversos phyla, tales como Equinodermos (Ofiuroideos y Asteroideos), Anélidos, Sipuncúlidos, etcétera, pero, en todos los casos el grupo más abundante y al que por ello se le prestó mayor consideración fue el de los moluscos, en especial, los bivalvos nombrados en la tabla 8. Algunas de las especies incluidas en esta tabla, tales como Eucrassatella digueti y Chione undatella alcanzan hasta 92 y 60 mm de longitud respectivamente, pero sólo Ch. undatella es abunndante y podría ser explotada en algunos bancos, como en el caso del banco 4 de Zihuatanejo donde era más abundante que M. aurantitaca (Fig. 2b, Banco 4; tabla 7). Las otras especies son todas pequeñas y poco abundantes, a excepción de Pinna rugosa que generalmente alcanza alrededor de 20 cm de largo, y Laevicardium elenense que, aunque no alcanza más de 20 mm, aparece esporádicamente con densidades de hasta 200 ejemplares por metro cuadrado. 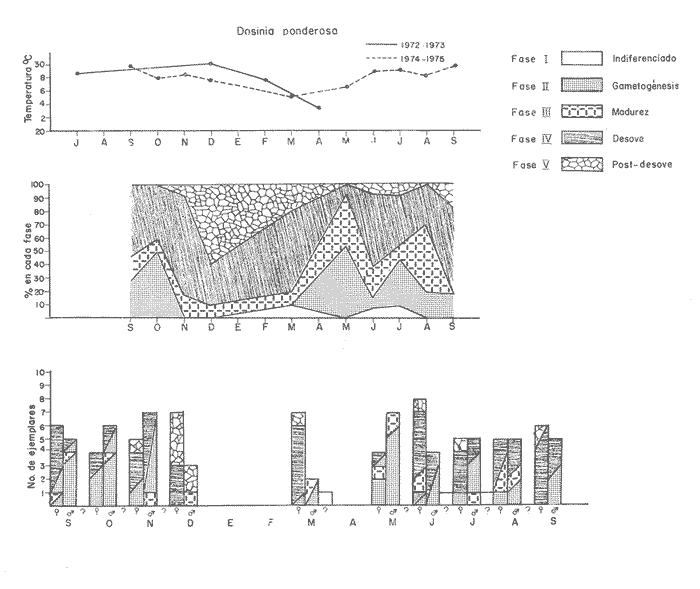 Fig.10. Análisis cualitativo y porcentual del desarrollo gonádico de las poblaciones combinadas de D. ponderosa de Zihuatanejo e IslaIxtapa,durante el ciclo anual comprendido entre septiembre de 1974 y septiembre de 1975 comparado a la variación de temperatura. a) Variación mensual de temperatura; b) Porcentaje mensual de las diferentes fases; c) Análisis individual porsexos. 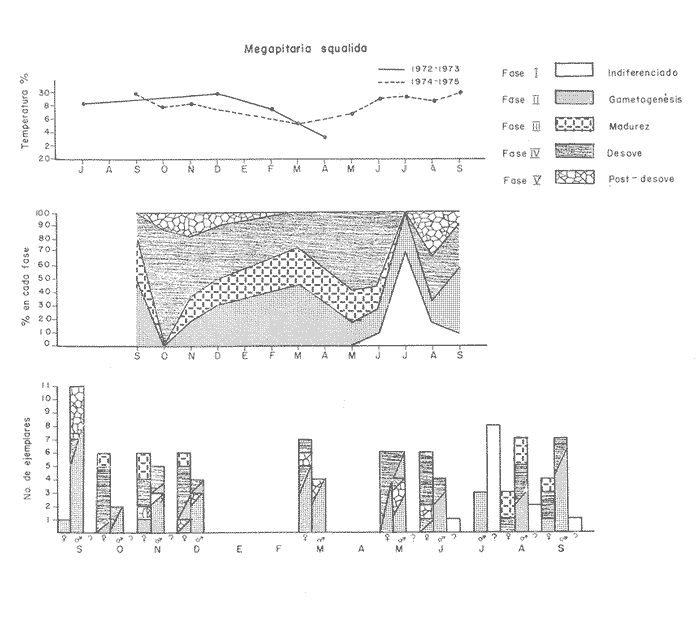 Fig. 9. Análisis cualitativo y porcentual del desarrollo gonádico de las poblaciones combinadas de M. squalida de Zihuatanejo e Isla Ixtapa, durante el ciclo anual comprendido entre septiembre de 1974 y septiembre de 1975 comprado a la variación de temperatura. a) Variación mensual de temperatura; b) Porcentaje mensual de las diferentes fases; c) Análisis individual por sexos.  TABLA 8 ESPECIES DE PELECIPODOS ASOCIADAS 9 MEGAPITARIA AURANTICA MEGAPITARIA SQUALIDAD Y DOSINIA PONDEROSA ZIHUTANEJO Los principales gasterópodos encontrados en los diferentes bancos fueron, en orden de abundancia: Terebra formosa, T. glauca, Turritella gonostomata, T. banksi, Strombus (Triconis) peruvianus, Oliva polpasta, O. incrassata, Conus sp., Cymakra granata, Mitra muricata, Haminoea angelensis y Polinices uber. ANÁLISIS DE LA PRODUCCIÓNEn la tabla 9 figura 11 se da la producción mensual de Megapitaria aurantiaca y Megapitaria squalida + Dosinia ponderosa, para los meses de marzo de 1974 a julio de 1975, según lo registrado por la Cooperativa Vicente Guerrero de Zihuatanejo, Gro. La producción de estas especies está dada por la demanda, la cual viene solamente de cuatro centros de consumo que son: Lázaro Cárdenas, Mich.; Acapulco y Zihuatanejo, Gro., y la Ciudad de México. Esta demanda puede ser cubierta solamente durante los meses de buen tiempo, es decir, entre noviembre y mayo, ya que el resto del año (junio a noviembre) las lluvias, vientos fuertes del sureste o vientos huracanados no permiten la pesca por encontrarse el agua muy turbia. Aun cuando las poblaciones de almeja no son explotadas intensivamente durante todo el año, la explotación se limita a unos cuantos bancos, los cuales son sobreexplotados en los meses de buen tiempo, por lo que, el tamaño medio de la población ha venido disminuyendo gradualmente. La producción durante estos 17 meses fue de 149 803 docenas, equivalente a 63 129.4 kilos (calculado del peso promedio para cada una de las especies) , lo que aportó un total de 2.122,702.00 pesos a los pescadores, valor que aumenta en un 30% al salir de la Cooperativa. 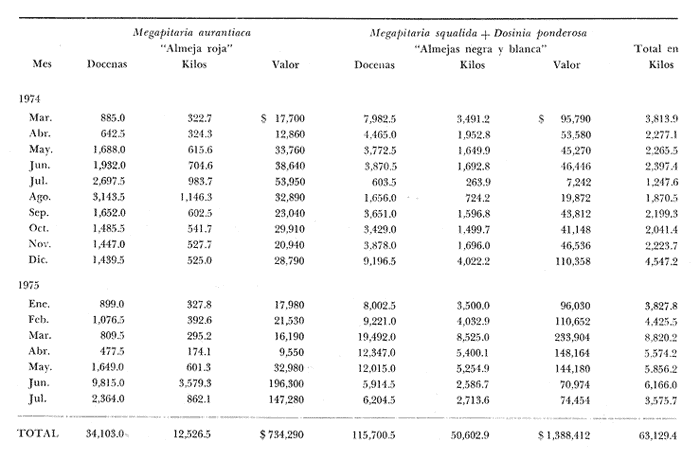 TABLA 9 PRODUCCIÓN MENSUAL DE "ALMEJAS" DE LA COOPERATIVA VICENTE GUERRERO (Perido de marzo, 1974 a julio, 1975) 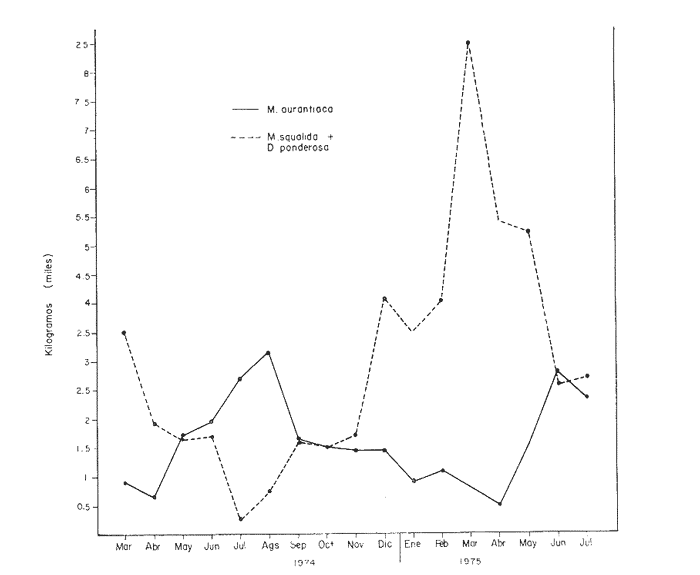 Fig. 11 Análisis de la producción mensual en kilos de M.aurantiaca y M. squalida + D. Ponderosa de la cooperativa Vicente Guerrero de Zihuantanejo entre los meses de marzo de 1974 a julio de 1975. ConclusionesDISCUSIÓN Y CONCLUSIONESLas variaciones extremas de temperatura para el agua de la Bahía de Zihuatanejo, así como su mayor estabilidad en la salinidad, se pueden explicar por ser un cuerpo de agua aparentemente con poca influencia de las corrientes costeras, sufriendo más intensamene los efectos de insolación y enfriamiento. El agua de fondo se mantiene más estable con respecto a la salinidad al no sufrir una renovación constante por efecto de las corrientes, quedando el agua estratificada. Esto no ocurre, sin embargo, en los lugares de poca profundidad, donde por efecto del viento, el agua de superficie es mezclada con la de fondo, como ocurre en el banco 4 de Zihuatanejo. Las variaciones extremas de salinidad, así como las temperaturas más estables en Isla Ixtapa, podrían explicarse por una corriente costera procedente del norte cuya intensidad de flujo disminuye sólo durante los periodos en que el viento del sureste sopla constantemente, mezclando el agua en la totalidad de la columna. De este modo la temperatura parece verse afectada por los cambios estacionales generales, por el aumento en intensidad de las corrientes del norte o por las aguas del sur que empujan los vientos del sureste. La salinidad se ve igualmente influenciada en forma muy especial por los ríos de la zona litoral que, además de disminuir la salinidad en los periodos de lluvia, aportan una gran cantidad de materia en suspensión que enturbia grandemente el agua. La disminución tan marcada de la salinidad que se registra en Isla Ixtapa durante el mes de marzo, puede explicarse si se considera que en esta época del año los vientos dominantes son del noroeste, que aunados a la corriente costera del norte, acarrean aguas de menor salinidad provenientes de la desembocadura del Río Balsas. La distribución de M. aurantiaca, M. squalida y D. ponderosa está dada preferencialmente por el tipo de sustrato y la profundidad, influyendo grandemente la acción del oleaje y corrientes que, como es de suponerse, tiene mayor efecto sobre la dispersión de las fases larvarias. Las tres especies habitan por debajo del límite inferior de las mareas, habiéndose encontrado hasta los 18 m de profundidad. Es posible que se encuentren a mayores profundidades, pero las actividades de buceo no se han realizado más allá. Keen (1971) reporta registros de 10, 160 y 60 m para M. aurantiaca, M. squalida y D. ponderosa respectivamente como profundidades máximas a las que se han encontrado. Todas viven enterradas en el fondo pero a profundidades diferentes: M. aurantiaca y D. ponderosa se encuentran preferentemente a 15 y 20 cm y M. squalida entre 5 y 10 cm de profundidad. Respecto de la naturaleza del fondo, M. aurantiaca ha sido encontrada en fondos de grava y arena en comunidad con una gran variedad de especies, principalmente bivalvos, compartiendo el mismo sustrato con M. squalida cuando el fondo es más arenoso. M. squalida y D. ponderosa son especies comunes en fondos de arena fina asociadas a otras especies de bivalvos relativamente escasos. La gran variación de abundancia de las dos especies en un mismo banco (Tabla 2a y b), es debido a que se trata de la sobreposición de dos especies con afinidad parcial de sustrato, ya que M. squalida se encuentra en fondos preferentemente arenosos y D. ponderosa, en fondos areno-limosos. Comparando la distribución determinada para cada una de las especies (Mapas 2b y 3b) con los datos dados por Tovar y Sánchez (1974) para la distribución del tipo de sedimentos de la bahía, resultaría que, de acuerdo a la clasificación usada por ellos, la "almeja roja" (M. aurantiaca) se distribuiría en fondo rocoso (!) la "almeja negra" (M. squalida) en fondos arenosos y la "almeja blanca" (D. ponderosa) en fondos areno-migajosos. Decididamente tanto la distribución de los tipos de sedimentos que ellas encontraron y la nomenclatura utilizada son conflictivas. El incremento mensual promedio de la talla observado en el análisis de las características de las poblaciones es, como ya se mencionó, de aproximadametne 5 mm, sin embargo, esto no ha sido posible compararlo con las mediciones hechas directamente sobre ejemplares marcados y aislados, habiéndose observado una disminución en el incremento promedio conforme aumenta el tamaño de los organismos. Sólo en algunos casos se registró crecimiento individual de más de 5 mm, lo cual puede ser el resultado de las condiciones adversas a que fueron sometidos, retardando su crecimiento en mayor o menor grado, según la tolerancia de cada ejemplar. Estos factores se introdujeron al mantenerlos en jaulas forradas de tela plástica, la cual se cubría rápidamente de epizoos reduciendo el flujo y calidad del agua que pasaba sobre las almejas. Otra alteración muy importante fue al sacar los ejemplares periódicamente para su medición, lo que detuvo su crecimiento por periodos más o menos prolongados, siendo más sensibles los ejemplares pequeños. Este receso en el crecimiento se demostró con la aparición de líneas de crecimiento muy marcadas después de cada medición, las que aparecen en forma natural en especies que habitan en climas fríos, apareciendo las líneas durante el invierno y durante la época de reproducción en que detienen su crecimiento (Sevilla, 1960; Olivier et al., 1971; Purchon, 1968). Debido a estas alteraciones se descartó la posibilidad de efectuar un cálculo de crecimiento por métodos estadísticos que permitieran determinar un ritmo natural de crecimiento con características isomtricas o alométricas de las variables consideradas. Respecto de la estuctura por tamaños y peso, todas las medidas de tendencia central, variabilidad y dispersión calculadas, por ejemplo, los valores elevados de variabilidad (coeficiente de dispersión) , indican que las muestras son heterogéneas incluyendo ejemplares de diversa edad y peso. Sin embargo, es importante hacer notar que el análisis de las características de la población indica que durante todo el año existe un reclutamiento de juveniles con algunos periodos en que es más notorio (dos en el caso de M. aurantiaca y M. squalida y tres para D. ponderosa), lo que está íntimamente ligado a las épocas de desove. La actividad gonádica se lleva a cabo durante todo el año independientemente de las variaciones de temperatura, realizando las tres especies un ciclo anual de desarrollo gonádico con un periodo de indiferenciación posterior a un periodo de desove máximo, ya sea prolongado a lo largo de varios meses o más o menos definido. La gametogénesis se mantiene durante todo el año pasando generalmente de una etapa de post-desove a una nueva etapa de gametogénesis. El análisis de gónadas efectuado sugiere cierta alternancia en los periodos de gametogénesis y desove de D. ponderosa y M. squalida. Esta última presenta desove máximo en los meses de octubre, abril, mayo y junio, en los que se presenta también un mayor porcentaje de gametogénesis de D. ponderosa. Esta especie, al contrario de la anterior, presenta en los meses de septiembre, marzo y junio un mayor porcentaje de ejemplares en desove, mientras que M. squalida presenta gametogénesis en septiembre y marzo e indiferenciación en junio y julio. Esta alternancia en los periodos de desove para estas dos especies que habitan en sustratos muy afines, de ser efectiva podría ser un mecanismo de adaptación para atenuar la competencia por espacio. La presencia de otras especies comestibles presenta un recurso adicional que puede ser aprovechado. Tanto Chione undatella, cuyo número es mucho mayor que el de M. aurantiaca en algunos bancos, como Laevicardium elenense aunque es una especie pequeña, harían posible una explotación temporal por su gran abundancia en ciertas épocas del año. Por último, la explotación de otras especies en los mismos bancos favorece a las especies de mayor interés comercial al disminuir la competencia. RECOMENDACIONESEl método de pesca empleado para la captura de las almejas puede ser considerado artesanal, sistema con el cual no se capturan todos los ejemplares disponibles a la pesquería, situación que mantiene un número de ejemplares adultos que aseguran la repoblación de los bancos. Sin embargo, en algunos lugares, en donde la captura se hace más intensa en los periodos en que no es posible pescar en toda el área, se llegan a obtener ejemplares que no han adquirido aún la madurez como consecuencia de la sobreexplotación. El uso de sistemas mecanizados para la pesca aumentaría la eficiencia de captura-hombre, ayudaría a mantener la producción durante todo el año independientemente de las condiciones del agua (que en la actualidad impiden el buceo durante algunos meses del año) y haría posible la explotación de áreas que, por su localización o profundidad, no son explotadas. La explotación mecanizada, racional y bajo control puede aumentar la pesquería y mantenerla indefinidamente, pero de llevarse a cabo sin control de tallas ni observando vedas, puede traer la destrucción del recurso en poco tiempo. Para la introducción de sistemas mecánicos de explotación, se recomienda determinar y fijar: a) Tallas mínimas de captura. De acuerdo a lo determinado en el ciclo gonádico, las tallas que se recomiendan para cada especie son: Megapitaria aurantiaca 50 mm Megapitaria squalida 50 mm Dosinia ponderosa 60 mm b) Veda temporal de los bancos de captura. Cuando el porcentaje de ejemplares mayores a la talla mínima sea menos de: 40% Megapitaria aurantiaca 50% Megapitaria squalida 40% Dosinia Ponderosa c) Determinar nuevas áreas de captura. Las áreas explotadas actualmente, en especial las comprendidas dentro de la Bahía de Zihuatanejo, son las más intensamente explotadas y no soportarían una pesca intensiva durante todo el año
AgradecimientosLos autores agradecen a las autoridades del Centro de Ciencias del Mar y Limnología de la UNAM, las facilidades otorgadas para la realización de este trabajo; y al Comité Editorial de los Anales de este Centro la gentileza de aceptar esta publicación. LITERATURAANSELL, A. D. Reproduction, growth and niortality of Venus striatula (Da Costa) in Karnes Bay, Millport, Scotland J. mar. biol. Ass. U. K. 1961 191-215 41 GARCÍA, E. Modificaciones al sistema de clasificación climática de Koppen (para adaptarlos a las condiciones de la República Mexicana). Instituto de Geografía, Univ. Nal. Autón. México 1973 246 p. KEEN, A. M. Sea Shells of Tropical West America. Marine Mollusks from Baja California, México to Peru. Standford University Press, U. S. A. Second Ed. 1971 1064 p., 22 láms. figs. (1a. edic. 1958). LANKFORD, R. Descripción general de la zona costera Guerrero y Michoacán. Subprograma de geología. In: Informe Final Ira. Eetapa Programa Uso de la Zona Costera de Michoacán y Guerrero. Convenio Comisión del Río Balsas, S. R. H. y Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México, Contrato de Estudios No. OCE-03-73 1974 42 p., 12 figs. LOZANO, F. Notas anatómicas sobre especies españolas de moluscos de interés pesquero. Bol. R. Soc. Esp. Hist. Nat., Sec. Biol. 1952 255-278, láms. 1015. 50 (2) NORTON, O. A. Some ecological observations on Dosinia discus Reeve at Beaufort, N. C. Ecology 1947 199-204 28 (2) OLIVIER, R. S., D. A. A. CAPEZZANI, J. 1. CARRETO, H. E. CHRISTIANSEN, V. J. MORENO, J. E. AIZPUN y P. E. PENCHASZADEH Estructura de la comunidad, dinámica de la población y biología de la "almeja amarilla" (Mesodesma mactroides Desh. 1854) en el mar Azul (Pdo. de Gral. Madariaga Bs. As., Argentina). Inst. Biol. Mar. Mar del Plata, Ser. Contribuciones 1971 3-90, 17 tablas, 35 figs. 122 OLSSON, A. A. Mollusks of the Tropical Eastern Pacific, Particulary from the Southern Half of the Panamic-Pacific Faunal Province (Panama to Peru). PanamicPacific Pelecypoda. Paleontological Research Institution, lthaca N. Y. 1961 574 p., 86 láms. PURCHON, R. D. The Biology of the Mollusca. Pergamon Press Oxford 1968 560 p., 185 figs. SEVILLA, M. L. Contribución al conocimiento de la madreperla Pínctada mazatlanica (Hanley) 1845. Revta. Soc. mex. Hist. Nat. 1969 223-261 30 STUARDO, J. y A. MARTíNEZ Resultados generales de una prospección de los recursos biológicos y pesqueros del sistema laguna costero de Guerrero, México. Acta Politécnica Mexicana 1975 99 (72) 2a. época, 16 STUARDO, J., A. MARTÍNEZ, A. YÁÑEZ, J. A. WEINBORN Y J. RUIZ Informe Final Ira. Etapa Programa Uso de la Zona Costera de Michoacán y Guerrero. Subprograma de Biología. Prospección de los recursos biológicos y pesqueros del sistema lagunar de Guerrero y en parte, del litoral rocosa de Michoacán. Convenio Comisión del Río Balsas, S. R. H. y Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México, Contrato de Estudios No. OC-E-03-73 1974 515 p., 20 mapas, 42 tablas, 19 figs., 34 láms. TOVAR, M. M. E. y M. A. SÁNCHEZ Tésis Profesional. Descripción de las variaciones estacionales de algunos grupos planctónicos en relación con elementos ambientales en la Bahía de Zihuatanejo, Gro. Fac. Ciencias Univ. Nal. Autón. México 1974 34 p., 51 figs. TREVALLION, A. Studies on Tellina tenuis Da Costa III. Aspects of general biology and energy flow. J. exp. Mar. Biol. Ecol. 1971 95-122 7 TREVALLION, A. J. H. STEELE y R. C. EDWARDS Dinamics of a benthic bivalve. J. H. Steele Marine Food Chains. Oliver and Boyd, Edinburgh 1970 285-295
|