|
MÉTODO BASADO EN LA TEORÍA DE LA
INFORMACIÓN PARA CALCULAR EL TAMAÑO DE MUESTRA DE ANIMALES
MARINOS
Trabajo recibido en marzo de 1976 y aceptado para su
publicación el 15 de octubre de 1976.
RENÉ J. BUESA
Centro de Investigaciones Pesqueras, La Habana,
Cuba.
Se propone un método para calcular el
tamaño de la muestra de animales marinos que consiste en graficar contra
muestras sucesivamente crecientes el valor correspondiente del índice de
diversidad de Brillouin, en bitios por individuo. El
tamaño adecuado será aquél al cual deje de obtenerse
información adicional sobre la población, el cual corresponde al
punto de la gráfica donde comienza a disminuir el valor de los bitios
por individuo. Se considera que el tamaño de muestra calculado con este
método posee una confianza superior a P
0.99
It is proposed a method based on the information
theory in order to calculate sample sizes for marine animals. The method
consists in plotting increasing sample sizes against Brillouin's diversity
index specific values (in bits) . Adequate sample size will
be the one where no aditional information is obtained, expressed as a decrease
in specific bitial value. Sample size calculated with this method possess a
confidence higher than P 0.99.
Los índices de diversidad derivados de la teoría de la información han servido para caracterizar los tipos más variados de comunidades naturales. Así tenemos que se han empleado para estudiar la producción primaria y el fitoplancton (Patten, 1963) ; el macrofitobentos marino (Buesa, 1974) ; el fitoplancton (Margalef, 1967), y varios tipos de comunidades, tales como las de corales (Loya, 1972) ; hormigas (Culver, 1972) ; insectos en general (Cameron, 1972) ; reptiles y anfibios (Lloyd et al., 1968) ; aves (Tramer, 1969) ; troglobiontes (Poulson y Culver, 1969) y peces estuarinos (Mc Erlean et al., 1973), así como la comunidad epifítica de Zostera (Marsh, 1973), la pelágica en general (Gueredrat et al., 1972) y hasta las plantas en una cronosecuencia (Reiners et al., 1971), entre otras, y todos ellos con resultados adecuados a los propósitos de los trabajos. La diversidad, como es sabido, es directamente proporcional a la información total, a la incertidumbre de predicción y a la entropía, todas referidas al conjunto en el cual se mide. Consecuentemente, el concepto de diversidad derivado de la teoría de la información, es inversamente proporcional al de la información inherente a cada subconjunto, al de la certeza de predicción sobre el conjunto y al de la uniformidad del conjunto. El contenido de información de una comunidad, o conjunto, o universo a muestrear, se puede expresar como la probabilidad con la que se puede predecir correctamente la muestra siguiente que se tome al azar (Cameron, 1972) . Mientras más subconjuntos haya en un conjunto y mientras más equitativamente se encuentren distribuidos aquéllos, habrá menor certeza en la predicción, es decir, habrá una mayor incertidumbre con respecto a la muestra siguiente, con un índice de diversidad concomitantemente mayor. Si partimos del hecho de que el objetivo de tomar una muestra es el de obtener información válida sobre el universo que se está estudiando, es lícito considerar como teóricamente correcto el empleo de las fórmulas de la teoría de la información para calcular el tamaño de muestra adecuado. La secuencia del razonamiento es como sigue: 1. Se sabe que el valor en bitios del índice de diversidad está en correspondencia con la entropía, y que ésta varía en función de la distribución de los elementos y del total de subconjuntos propios; y 2. Podemos decir que dentro de un cierto ecosistema, la población de una especie dada está constituida por el conjunto de sus ejemplares, que al mismo tiempo es el universo a muestrear. 3. Cualquier muestra que se tome de ese conjunto es un subconjunto propio, formado a su vez por clases de edad, largo, peso u otra categoría, que son transitivamente subconjuntos propios de la muestra, cada uno con una frecuencia conocida. 4. Cada tamaño, de muestra poseerá un valor de diversidad, de información sobre el universo del cual procede, que variará en función de las características del propio universo, y según el tamaño de la muestra. 5. El universo o conjunto a muestrear posee un valor de diversidad y las muestras o subconjuntos que se extraigan de él poseerán otro valor de diversidad, tan próximo al del universo mientras más representativas de él sean. 6. Mientras que el valor del índice de diversidad de muestras de frecuencia sucesivamente mayores aumente, querrá ello decir que estamos obteniendo más información, o información adicional sobre el conjunto, pero cuando el valor en bitios se mantenga es table o incluso comience a disminuir, querrá ello decir que no se obtiene información adicional sobre la población aunque aumentemos el tamaño de la muestra, o lo que es igual, que la muestra, por ser representativa del conjunto, se hace estable en cuanto a su nivel de información sobre éste. 7. En consecuencia, el tamaño de muestra adecuado será aquél cuya diversidad sea máxima, y no otro. MATERIALES Y MÉTODOSSe utilizaron datos de dos crustáceos: el camarón acanalado (Penaeus duorarum) y la langosta espinosa (Panulirus argus) ; de tres peces costeros demersales: la lisa o lebrancho (Mugil liza). la biajaiba (Lutjanus synagris) y el bajonado o mojarra rayada (Calamus penna), y de dos especies de peces pelágicos: el bonito o barrilete (Katsuwonus pelamis) y la albacora (Thunnus atlanticus), es decir, se seleccionaron especies de importancia comercial, pero que desde el punto de vista sistemático y ecológico se encuentran bien "alejadas" entre sí. Para evitar cualquier error como posible consecuencia de que algunas de las muestras poseyera algún tipo de sesgo, así como porque es adecuado para muestras pequeñas, se utilizó el índice de Brillouin (1960) para calcular la diversidad individual de las muestras de los organismos marinos, según la expresión: B = 1/N log2 N!/πni! donde: B = valor de la diversidad individual en bitios N = total de elementos en el conjunto (muestra) n = total de elementos en el subconjunto i-ésimo (clase dentro de la muestra) En todos los casos se seleccionaron muestras consideradas como aleatorias y crecientes en número (cantidad de elementos), hasta valores superiores a quinientos ejemplares (elementos) en casi todas las especies estudiadas. En algunos casos las frecuencias muestreales por clases de largo se fueron acumulando, y siempre se determinó la desviación típica para calcular el tamaño de la muestra por métodos estadísticos convencionales, como el que aparece a continuación, donde: N= (sZ/T)2 N = tamaño de muestra s = desviación típica muestral Z = desviación relativa correspondiente al nivel de confiabilidad deseado T = valor de la precisión deseada El valor del índice de diversidad en bitios por individuo se graficó contra el tamaño de la muestra utilizada para calcularlo y los puntos se ajustaron a "mano alzada" para conocer aproximadamente el punto de inflexión o asintotismo de las curvas. RESULTADOSEl primer análisis se efectuó con relación al efecto que pudiera causar en el índice de diversidad el hecho de trabajar con muestras aisladas o acumuladas, y se pudo comprobar (tabla 1) que el tamaño de muestra al cual la información sobre el conjunto comienza a disminuir es menor cuando se calcula en las acumuladas. Esto se debe a la permanencia en las muestras más numerosas de las clases sucesivamente precedentes, lo cual tiende a aumentar la uniformidad. 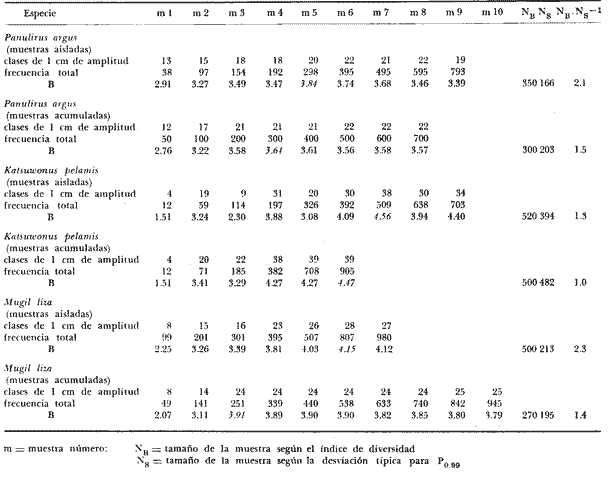 TABLA 1 VARIACIÓN DEL ÍNDICE DE DIVERSIDAD DEBRILLOUIN (B) PARA MUESTRAS AISLADAS Y ACUMULADAS La amplitud de las clases, al comparar las de uno y de cinco centímetros (tabla 2), si bien no hizo variar mucho el tamaño de las muestras calculado según los bitios individuales de información (i. e., la variación del valor de B) , sí determinó un incremento del tamaño calculado según la desviación típica muestral, así como una disminución de los valores absolutos de "B" por muestra, debido a la reducción del número de subconjuntos propios o clases de largo. 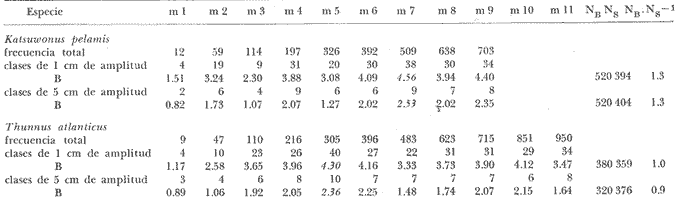 TABLA 2 VARIACIÓN DEL ÍNDICE DE DIVERSIDAD DEBRILLOUIN (B) PARA MUESTRAS AISLADASCON AMPLITUDESDE INTERVALOS DECLASES DIFERENTES 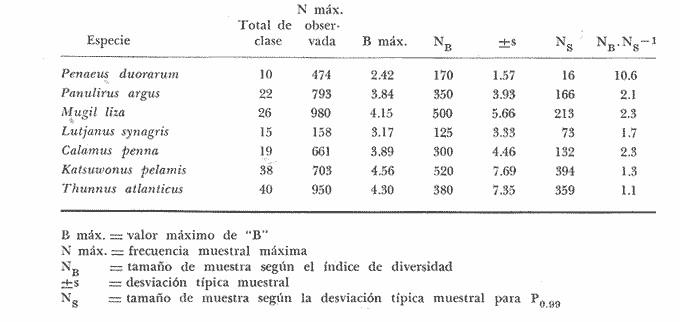 TABLA 3 RESUMEN DE LOS RESULTADOS OBTENIDOS CON MUESTRAS AISLADAS (INDIVIDUALIZADAS) DISCRIMINADAS EN CLASES DE UN CENTIMETRO DE AMPLITUD PARA EL LARGO DE LOS EJEMPLARES Para las especies consideradas el tamaño de la muestra calculado con el índice de diversidad de Brillouin (tabla 3 y figura 1) fue entre 1.1 y 10.6 veces mayor que el tamaño de muestra calculado a partir de la desviación típica muestral para una P 0.99 y en todos los casos fue menor que el tamaño calculado para una P 0.999 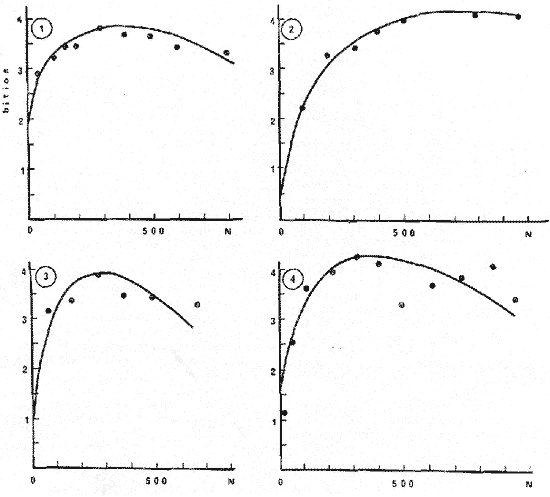 Fig. 1. Tamaño de la muestra calculado según la gráfica de la variación del índice de diversidad de Brillouin (B) contra el tamaño de la muestra (N), para cuatro especies de animales Thunnus atlanticus. DISCUSIÓNSe ha descrito que en las poblaciones fitoplanctónicas después de identificar y contar trescientas o setecientas células, la diversidad en bitios por célula no suele variar más allá de la segunda cifra decimal, y que, empíricamente, se ha encontrado que la diversidad individual tiende a valores próximos a los 5 bitios (Margalef, 1967). En los casos presentados ningún valor del índice de diversidad individual alcanzó el límite empírico señalado, y el mayor fue de 4.56 bitios (para el K. pelamis). Por ello el comportamiento señalado para el fitoplancton se puede extender al caso de las especies que incluimos, a pesar de su "lejanía" ecológica y sistemática. Aunque el límite de 5 puede pensarse que constituye algo en contra del método propuesto, el índice de diversidad individual de las especies consideradas presentó un comportamiento semejante en todas, aumentando hasta un cierto valor para después disminuir o mantenerse más o menos estable a partir de él, independientemente de que en algunos casos el máximo no llegó a alcanzar ni siquiera los 3 bitios por individuo (tabla 3). Por todo lo anterior se puede considerar que el comportamiento del índice de diversidad con respecto al tamaño de las muestras es independiente de su valor absoluto. Los resultados parecen indicar que realmente al aumentar el tamaño de la muestra el valor de la información individual expresado por los bitios aumenta hasta un cierto valor máximo para un tamaño de muestra dado, a partir del cual comienza a disminuir o se hace más o menos estable, o asintótico, por lo que no es necesario obtener muestras mayores a las que poseen dicha información máxima sobre el universo muestreado, ni es correcto obtenerlas menores. Los ejemplos presentados, basados en clases de largo, realmente dentro del conjunto constituyen un continuo y su separación puede ser considerada como arbitraria. El empleo del método basado en las clases de edad puede que sea más justificado, aunque quizá no se puede efectuar en todos los casos. En esencia el método viene a sustituir a una variancia, aunque puede detectar mejor los efectos de factores naturales predecibles, tales como la natalidad, crecimiento y mortalidad sobre los subconjuntos del continuo, sobre todo si se tiene en cuenta que para alcanzar una cierta talla la especie, los ejemplares, se han visto sometidos a todos los factores señalados. El método puede servir también para definir la suficiencia de las muestras con las que se deben estudiar las fracciones precomerciales de las poblaciones, con pocas clases de edad, y talla, y con diversidad baja, tanto como las fracciones adultas, más viejas y con estructuras poblacionales más diversas. El tamaño de muestra calculado con el desvío para una P 0.99 es menor que el correspondiente a la gráfica de la variación de "B" contra la frecuencia total de la, muestra (N), por lo que se infiere que el resultado que se puede esperar con este método es superior al de la probabilidad señalada. La gráfica de B contra N parece que puede ser ajustada matemáticamente mediante un polinomio. Ese ajuste, de resultar significativo según métodos convencionales como el de GaussDoolittle, permitirá calcular el tamaño exacto de la muestra a partir de la evaluación de la primera derivada de la ecuación correspondiente a cada especie. Este tratamiento matemático obviará la imperfección y el subjetivismo al que están expuestos todos los métodos gráficos, como el propuesto. ConclusionesEl tamaño de muestra conveniente para animales marinos puede ser calculado a partir de la diversidad calculada para una serie de muestras aleatorias de cada especie, ya que es un indicador de la información total que se obtiene sobre la población. El método propuesto se basa en la obtención de muestras crecientes en número (e. g.: 50, 100, 200, 300, 500 y 700 individuos) de una especie dada. A cada una, considerada individualmente y discriminada en clases de largo de intervalos pequeños, se le calculará el ínclice de diversidad de Brillouin. Los resultados en bitios por ejemplar se graficarán contra el tamaño de la muestra correspondiente y se escogerá como tamaño conveniente para los trabajos subsiguientes aquél al cual el índice de información individual decaiga o se mantenga estable con respecto al valor máximo obtenido. Se considerara que el ajuste de los resultados mediante un polinomio, y la evaluación posterior de la primera derivada de la ecuación estadísticamente, significativa, puede obviar en gran medida el subjetivismo que implica el método gráfico propuesto, sobre todo en los casos en los cuales la evaluación del índice de diversidad contra el tamaño de la muestra no es muy evidente o fácil de ajustar a "mano alzada". AgradecimientosEl autor desea expresar su agradecimiento a Ramón Margalef por la lectura y comentarios sobre el manuscrito. Se agradecen también los comentarios de G. González sobre este artículo. L. S. Álvarez-Lajonchere facilitó sus datos de lisa, y los colegas Ma. E. Ruiz de Quevedo y C. A. Carles los de camarones y peces pelágicos, respectivamente. LITERATURABRILLOUIN, L. Science and Information Theory. Academic Nueva York 1960 BUESA, R. J. Biomasa del macrofitobentos de la plataforma noroccidental de Cuba. INP/CIP, Cuba; Res. Invest. 1974 51-54 1 CAMERON, G. N. Analysis of insect trophic diversity in two salt marsh communities. Ecology 1972 58-73 53 (1) CULVER, D. C. A niche analysis of Colorado ants. Ecology 1972 126-131 53 (1) GUERFDRAT, J. A., R. GRANDPERRIN y C. ROGER Diversité spécifique dans le Pacifique Equatorial: évolution de l'ecosystéme. Cah. OTROM, sér. Océanogr. 1972 57-69 10 (1) LOYA, Y. Community structure and species diversity of hermatypic corals at Eilat, Red Sea. Marine Biology 1972 100-123 13 (2) LLOYD, M., R. F. INGER y F. W. KING On the diversity of reptile and amphibian species in a Bornean rain forest. Amer. Natur 1968 497-515 102 MARGALEF, R. El ecosistema. H. Ginés y R. Margalef Ecología Marina. Fundación La Salle de Ciencias Naturales, Monogr. 1967 377-453 14 MARSH, G. A. The Zostera epifaunal community in the York River, Virginia. Chesap. Sci. 1973 87-97 14 (2) McERLEAN, A. J., S. G. O'CONNOR, J. A. MIHURSKY Y C. I. GIBSON Abundance, diversity and seasonal patterns of estuarine fish populations. Estuar. Coast. mar. Sci. 1973 19-36 1 (1) PATEEN, B. C. The information concept in ecology: some aspects of information-gathering behavior in plankton. W. Fields y W. Abbott Information Storage and Neural Control, Cap. 7, Thomas, Illinois. 1963 POULSON, T. L. y D. C. CULVER Diversity in terrestrial cave communities. Ecology 1969 153-158 50 (1) REINERS, W. A., I. A. WORLEY y O. B. LAWRENCE Plant diversity in a chronosequence at Glacier Bay, Alaska. Ecology 1971 55-69 52 (1) TRAMER, E. J. Bird species diversity: components of Shannon's formula. Ecology 1969 927-929 50 (5)
|