|
GENERALIDADES ICTIOLÓGICAS DEL SISTEMA LAGUNAR
COSTERO DE HUIZACHE-CAIMANERO, SINALOA, MÉXICO
Trabajo recibido el
15 de enero de 1976 y aceptado para su publicación el 15 de octubre de
1976.
FELIPE AMEZCUA-LINARES
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología, Laboratorio de
Ictiología y Ecología Estuarina. Contribución 59 del
Centro de Ciencias del Mar y Limnología, UNAM.
Las comunidades ictiofaunísticas del sistema
lagunar costero de Huizache-Caimanero (costa noroccidental del Pacífico
de México) , tienen una alta diversidad compuesta por 27 familias, 46
géneros y 60 especies, distribuidas en un 8% de peces
dulceacuícolas, 8% de peces típicamente estuarinos, 31% de peces
marinos que visitan el estuario como adultos y para alimentarse, 33% de peces
marinos que utilizan el estuario como áreas naturales de crianza y 20%
de peces marinos que son visitantes ocasionales. Las familias mejor
representadas en diversidad son: Carangidae (7 especies) , Gerridae (5
especies) , Sciaenidae (5 especies) , Gobiidae (5 especies) ; y con 3 especies,
Mugilidae, Centropomidae, Pomadasyidae Engraulidae y Clupeidae.
Las comunidades varían en su composición
y abundancia relativa de especies de acuerdo a: 1) las condiciones
hidrológicas del sistema, consecuencia de 2) la estación del
año y 3) la localidad dentro del estuario y sus gradientes de salinidad.
No obstante, las especies más representativas en número de
individuos y biomasa durante todo el año son: Mugil curema,
Galeichthys caerulescens, Diapterus peruvianus y Pomadasys
macracanthus.
Se discuten aspectos biológicos generales sobre
alimentación, crecimiento, madurez y migraciones para
Galeichthys caerulescens y Mugil curema,
las 2 especies más importantes desde el punto de vista biológico,
ecológico y pesquero del sistema lagunar. Se recomiendan futuras
investigaciones conducentes a evaluar, administrar y explotar racionalmente los
recursos pesqueros de Huizache-Caimanero y sus perspectivas de óptimo
aprovechamiento.
The ichthyofaunistic communities of the
Huizache-Caimanero coastal lagoon system (northwestern Pacific coast of Mexico)
have a high diversity composed of 27 families, 46 genuses, and 60 species,
distributed as follows: 8 percent freshwater fishes, 8 percent typically
estuarine fishes, 31 percent marine fishes which visit the estuary as adults
looking for food, 33 percent marine fishes which use the estuary as a natural
breeding area, and 20 percent marine fishes which are occasional visitors. The
families of greatest diversity are: Carangidae (7 species), Gerridae (5
species), Sciaenidae (5 species), and Gobiidae (5 species). Those with 3
species are: Mugilidae, Centropomidae, Pomadasyidae Engraulidae and
Clupeidae.
The communities vary in their compositíon and
relative abundance of species in relation to: 1) the hydrologic conditions of
the system, which are a consequence of 2) the season of the year, and 3) the
zone within the estuary and its degrees of salinity. Nevertheless, the species
which are most representative in terms of number of individuals and biomass
during the year are: Mugil curema, Ga1eichthys caerulescens, Diapterus
peruvianus and Pomadasys macracanthus.
General biological aspects of feeding, growth,
maturity and migrations are discussed for Galeichthys caerulescens and Mugil
curema, the two most important species from the point of view of the
biological, ecological and fishing aspects of the lagoon system. Further
research is recommended to evaluate, administer and rationally exploit the
fishing resources of Huizache-Caimanero.
Las lagunas costeras del Sur del Estado de Sinaloa y en general del noroeste de México, se han estudiado desde hace algunos años por diversas instituciones debido a la importancia que tienen dentro de las pesquerías nacionales. Entre ellas, la Universidad Nacional Autónoma de México inició en 1969 diversos proyectos encaminados a efectuar estudios relacionados con la biología del camarón y otros aspectos ecológicos de las lagunas costeras situadas en el Pacífico noroccidental por la importancia que estos crustáceos revisten en la economía pesquera mexicana. Los aspectos ictiológicos más interesantes de estas investigaciones se encuentran condensados en los informes de Carranza (1969a, 1969b, 1969c, 1970), Carranza y Amezcua-Linares (197la, 1971b), Amezcua-Linares (1972), González-Villaseñor (1972) . Durante el desarrollo de los mencionados programas, se fundó la Estación de Investigaciones Marinas Mazatlán, dependiente del Centro de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México (UNAM), en la que se llevó a efecto este trabajo como parte de un convenio actualmente en desarrollo, celebrado entre la propia UNAM y el Consejo Británico, con la participación de personal de ambas nacionalidades. Inicialmente el mencionado proyecto se enfocó al estudio de los principales depredadores y competidores del camarón (braquiuros y peces) , y parte de esos resultados se presentan en este trabajo con información obtenida y procesada en Mazatlán e interpretada e integrada en el Laboratorio de Ictiología del Centro de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México. ANTECEDENTESLa importancia de las lagunas costeras y estuarios de México como áreas potencialmente productivas, ha sido destacada desde hace bastante tiempo y en diferentes trabajos, los cuales han sido por lo general aislados, poco frecuentes y normalmente de objetivos dispares. Sin embargo, recientemente y dentro de la línea de investigación de Biología y Ecología de Peces de lagunas costeras y estuarios que se desarrolla en el Laboratorio de Ictiología del Centro de Ciencias del Mar y Limnología de la UNAM, se ha señalado la necesidad de enfatizar en los estudios ictiológicos desde un punto de vista sistemático, multidisciplinario e integral del ecosistema, con el objeto de hacer una buena evaluación de los recursos biológicos y pesqueros de esas áreas altamente productivas, y de ese modo recomendar una correcta administración de los recursos ictiológicos en México; Stuardo et al. (1974), Yáñez-Arancibia (1975a, 1975b, 1975c), Yáñez-Arancibia y Nugent, 1975 y 1976, Yáñez-Arancibia et al. (1976). Anterior a estos trabajos se han hecho estudios que deben de ser considerados como básicos en la continuidad de la línea de investigación ya mencionada y cuyos resultados son necesariamente valiosos (Ramírez, 1952; Darnell, 1962; Álvarez, 1970; Carranza, 1970; Carranza y Amezcua-Linares, 1971b; Toral, 1971; Amezcua-Linares, 1972; GonzálezVillaseñor, 1972; Castro, 1974; Castro et al., 1974; Reséndez, 1970 y 1973) . La complejidad de estas áreas dificulta en gran medida la interpretación biológica y ecológica de las comunidades ictiofaunísticas que allí habitan. Yáñez-Arancibia (1975a, 1975b y 1975c) y Yáñez-Arancibia et al. (1976) , discuten metodologías conducentes a la prospección, evaluación y explotación racional de los recursos ícticos lagunares. Dentro de esta línea de investigación se ha desarrollado el presente estudio. ÁREA DE ESTUDIOEl área ha sido estudiada desde el punto de vista geológico por Ayala-Castañares et al. (1970), desde el punto de vista hidrológico por Arenas-Fuentes (1970) y biológicamente, en forma parcial, por García-Cubas (1970), Gómez-Aguirre et al. (1970 y 1974), Ortega (1970) y Cabrera (1970). Es característico encontrar abundantes manglares entre la vegetación macrófita de las orillas. GEOGRAFÍALas lagunas de Caimanero y Huizache con superficie aproximada de 175 Km2 (Chapa y Soto, 1969), se encuentran situadas en la planicie costera del Sur del Estado de Sinaloa, México, entre los 22º 50' y 23º 05' latitud norte y los 105° 55' y 106° 20' longitud oeste. Poseen dos aportes fluviales, uno localizado al noroeste, el río Presidio y otro al sureste, el río Baluarte. Son lagunas costeras de aguas someras y de nivel muy variable de acuerdo a las estaciones del año; su nivel máximo ocurre en la temporada de lluvias (junio a septiembre) cuando en las regiones centrales existe una profundidad máxima de 2 m, mientras que en la temporada de secas el nivel es muy bajo y la extensión muy reducida quedando la mayor parte del embalse totalmente descubierto. Esto se debe a que las lagunas se están azolvando rápidamente con sedimentos acarreados por los ríos, provenientes de zonas altas como resultado de un proceso geológico natural. En las figuras 1 y 2 se señala el área de estudio con las características topográficas y las estaciones de colecta. 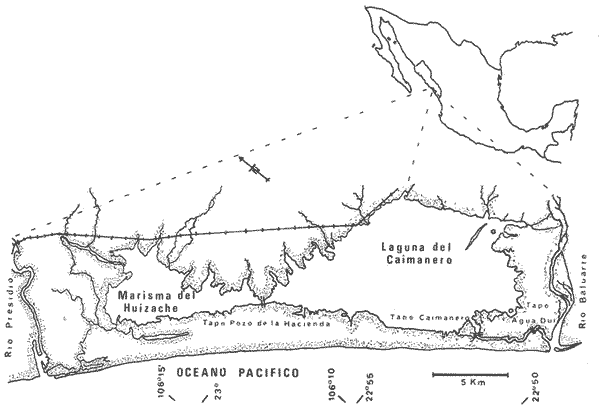 Fig. 1. Sistema lagunar costero de Huizache-Caimanero, Sinaloa, México. Se señala la superficie aproximada y datos topográficos 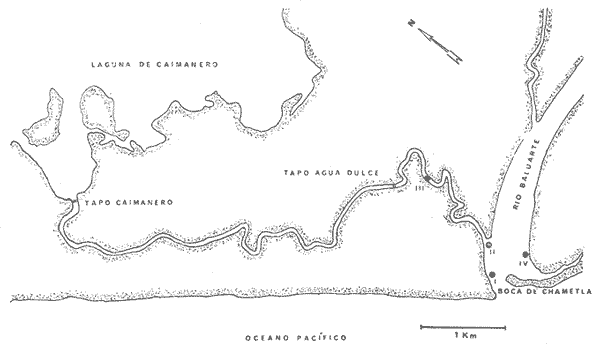 Fig. 2. Detalle del Canal de Agua Dulce, donde se ubican las estaciones de muestreo HIDROLOGÍALas salinidades medias son de 19 %, existiendo un rango de 0 % a 36% de acuerdo a la época del año. En la época de lluvias la salinidad se abate por completo con los grandes aportes fluviales. Durante la sequía (febrero a mayo) el agua marina predomina dentro del sistema. Es necesario mencionar que esto sucede en el estuario y esteros, ya que en las lagunas durante la época de sequía se registran salinidades hasta de 60%, llegando incluso la laguna del Huizache a una total desecación. La temperatura media es de 24º C, presentándose un rango de 15° a 32º C dependiendo de la época del año, ya que las temperaturas más bajas se registran en enero y las más altas en agosto. La temperatura se eleva considerablemente en la parte central de las lagunas por la continua insolación a que están expuestas, llegando a los 36º C. La cantidad de oxígeno disuelto en el agua ha sido estudiada con detalle en la zona de trabajo por Arenas-Fuentes (1970), quien encontró valores siempre superiores a los mínimos requeridos por los peces para su sobrevivencia (3.0 ml/I). En las figuras 3 a 6 se señalan los valores medios de las salinidades y temperaturas para las 4 estaciones de colecta. Las salinidades más bajas se registraron durante la temporada de lluvias, junio a octubre. Sin embargo en la figura 3 se observa un valor alto debido a que es la zona de la boca y existía en ese momento marea viva. Se señala en las citadas figuras que las temperaturas más bajas se registraron de diciembre a febrero, existiendo aparentemente un rango de 12º C entre la época fria y las épocas más cálidas que son entre junio y septiembre. CLIMADe acuerdo con la modificación de García (1973), a partir del sistema de Köepen, el área ,presenta un clima cálido, subhúmedo y con precipitaciones en verano y en invierno, aunque en esta última estación las lluvias son mínimas ya que están representadas por menos del 5% del total anual, el que supera los 750 mm. 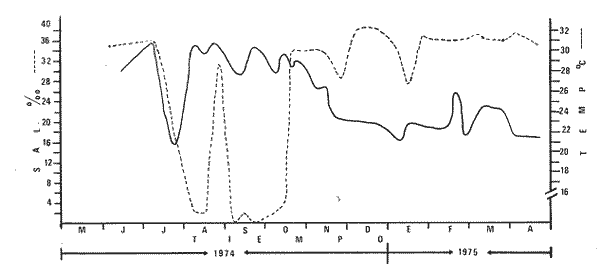 Fig. 3. Salinidad y temperatura registrada durante el periodo de mayo de 1974 a abril de 1975 en la estación 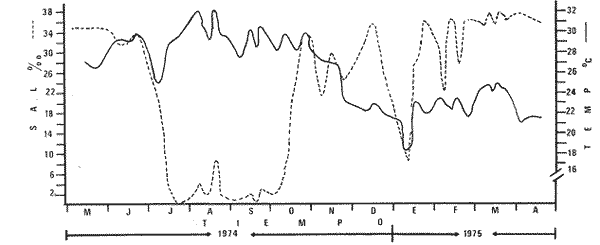 Fig. 4. Salinidad y temperatura registrada durante el periodo de mayo de 1974 a abril de 1975 en la estación II, Boca del Canal de Agua Dulce. 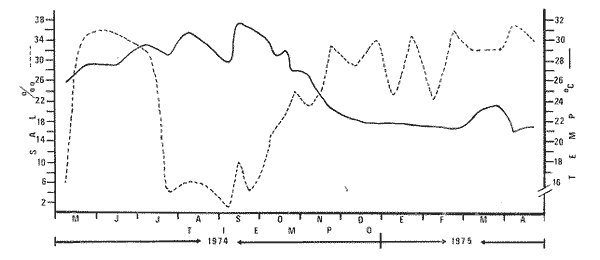 Fig. 5. Salinidad y temperatura registrada durante el periodo de mayo de 1974 a abril de 1975 en la estación III, Canal de Agua Dulce. 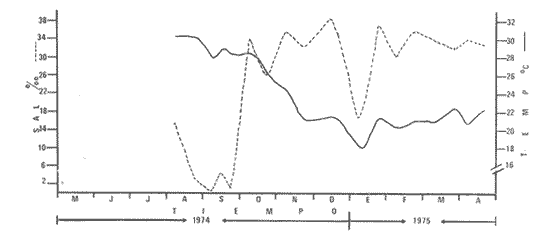 Fig. 6. Salinidad y temperatura registrada durante el periodo de mayo de 1974 a abril de 1975 en la estación IV, Estuario de Chametla. Existe un régimen térmico invernal uniforme, no bien definido, con poca variación en la temperatura media mensual a través del año, la que siempre es superior a los 18º3 C. La diferencia entre el mes más cálido y el más frío oscila entre los 7º C y 14' C (Tamayo 1949). MATERIALES Y MÉTODOS
MATERIAL DE ESTUDIOSe examinaron para las curvas de crecimiento, relación peso-longitud y desarrollo sexual, 292 ejemplares de Galeichthys caerulescens con un rango de tallas entre 133436 mm de longitud y de 28-1419 g de peso. Así mismo, para la lisa Mugil curema fueron examinados 535. ejemplares con tallas entre 14-350 mm y entre 1-580 g de peso. En ambas especies los ejemplares proceden de las 4 estaciones de colecta y de prácticamente todas las fechas de muestreo. Para el análisis de estómagos, se tomaron 23 ejemplares de Galeichthys caerulescens de las 4 localidades, el resto se encuentra en estudio para complementar estas observaciones. La lista de 27 familias, 46 géneros y 59 especies fue obtenida y determinada del análisis taxonómico de 2,500 especímenes colectados en las 4 estaciones durante el ciclo anual. ACTIVIDADES DE CAMPOPara las colectas se utilizó una lancha plana de fibra de vidrio tipo trimarán de 5.0 m de eslora, con un motor fuera de borda de 35 HP, desde la cual se operaron las siguientes artes de pesca: a) Chinchorro playero, de 54.0 m de largo por 2.55 m de alto, con malla de hilo nylon y luz variable; alas de 1.7 m con luz de malla de 3.3 cm; partes medias de 3.57 m con luz de malla de 1.8 cm y la bolsa de 12.92 m con luz de malla de 1.2 cm. b) Red de arrastre de prueba camaronera, con tablas de 45.5 por 23.5 cm, 2.13 m de abertura en la boca y luz de malla de 1.2 cm c) Atarraya de hilo nylon con 950 mallas, luz de 3.2 cm y 8.0 m de diámetro. Ocasionalmente se empleó una atarraya de hilo nylon NI? 1 con luz de malla de 1.2 cm y 4.18 m de diámetro. Para medir la temperatura se usó un termómetro marca Taylor con escala de 1 a 500 C, y para mediciones de salinidad se usaron refractómetros American Optical de lectura directa, con compensador automático de temperatura lectura mínima de 1%. Se efectuaron 25 muestreos en 4 diferentes estaciones seleccionadas y relacionadas con las colectas de larvas de camarón en las fechas inmediatas correspondientes a las fases lunares (Tabla 1). En cada una de ellas se muestreó con los diferentes tipos de artes de pesca tratando de cubrir de la mejor manera posible los habitats aumentando de esta manera la eficiencia de captura. Los 4 lugares de colecta para los muestreos de peces, se denominaron en números romanos, como se indica en la figura 2 y cuya descripción es la siguiente:  TABLA 1 FECHA DE LOS MUESTREOS DE PECES YFASES LUNARES EN HUIZACHE-CAIMANERO, SINALOA, MEXICO 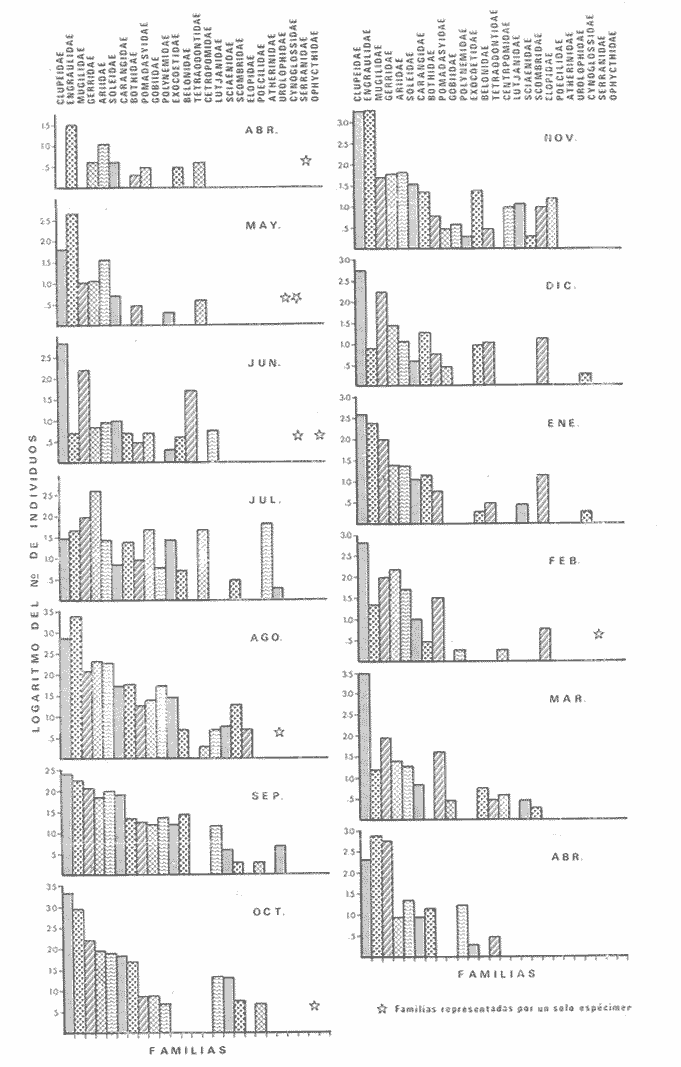 Fig. 7 Variación estacional anual de las principales familias registradas entre el Canal de agua Dulce y la Boca de Chametla. ESTACIÓN I. Corresponde a la zona de la boca de Chametla; vía de acceso al mar localizada al sur del sistema Huizache-Caimanero. ESTACIÓN II. Se localiza en la boca del canal de Agua Dulce, en la intersección de éste con el río Baluarte, prácticamente dentro del estuario. ESTACIÓN III. Está situada en el canal del estero de Agua Dulce, en regiones cercanas al tapo del mismo nombre. ESTACIÓN IV. Localizada en el margen sur y enfrente a la Estación II, donde el embalse recibe directamente- la descarga de aguas fluviales. A partir del mes de agosto se incluyó esta colecta consistente únicamente en lances de chinchorro playero, ya que representaba un interés excepcional por la gran cantidad de individuos y especies que ahí se encontraban, sobre todo durante la época de precipitaciones. En la Estación III, sólo se usó con regularidad la atarraya, debido a que el chinchorro playero y la red de arrastre eran difíciles de operar efectivamente por las zonas de manglar. En cada una de las estaciones se efectuaron un lance con el chinchorro, uno con la red de arrastre y cinco con la atarraya, a excepción de algunos casos en que debido a circunstancias especiales se efectuó un lance extra. Prácticamente la totalidad de los muestreos fueron hechos en marea alta o flujo en las horas próximas y posteriores al crepúsculo y el alba. Esto fue programado originalmente con el fin de relacionar los resultados con los muestreos de larvas de Penaeus spp. que se efectuaron en las mismas condiciones. Los ejemplares fueron fijados en el campo con formalina al 10%, haciendo una insición en el vientre para que el fijador llegara inmediatamente al estómago y con esto detener el proceso digestivo, ya que al estar sometidos a altas temperaturas se acelera este proceso. Posteriormente fueron introducidos en bolsas de polietileno con su etiqueta respectiva y éstas a su vez en cubetas de plástico de 18 litros con tapa hermética, para ser trasladadas al laboratorio. ACTIVIDADES DE LABORATORIOEn el laboratorio se usaron diversos instrumentos para estudiar los ejemplares, tales como un ictiómetro convencional de 50 cm de largo y graduado en mm; una balanza granataria marca OHAUS con capacidad máxima de 2,610 g y con precisión de 0. 1 g; un microscopio estereoscópico marca Karl-ZEISS con diferentes aumentos. Para efectuar las mediciones se usó la longitud furcal, entendida ésta como la distancia comprendida entre la parte anterior del hocico y extremo posterior de los radios medios de la aleta caudal, que corresponden precisamente a la furca de la aleta. Para determinar el sexo y las fases del desarrollo gonádico, se siguió el criterio de NikoIsky (1963), ya que se estima el más adecuado para el tipo de peces en estudio. Generalmente la determinación de sexo y estado sexual fue hecho con los ejemplares ya fijados en formol al 10%, aproximadamente unos días después de su captura. Este procedimiento puede tener el inconveniente de que los órganos sexuales sufran decoloración por acción del fijador, sin embargo dada la cantidad de peces, resultó inoperable efectuar determinaciones en fresco y se optó por examinar ejemplares fijados. Un solo método de análisis de estómago no resulta completo en la mayoría de los casos debido a la variedad de hábitos alimenticios que presentan los peces, por ello se utilizaron 3 tipos de métodos diferentes para una mejor evaluación (i.e., volumétrico, numérico y de frecuencia) ampliamente discutidos en trabajos recientes de Yáñez-Arancibia (1 975a y 1975b) y Yáñez-Arancibia et al. (1976). Todo el material biológico empleado para los análisis de estómagos proviene exclusivamente de lances con chinchorro playero y atarraya, ya que muchos de los peces al sentirse atrapados regurgitan el alimento, resultando inadecuadas otras artes para este tipo de estudios. Por ello, sólo es posible obtener buenas muestras cuando los peces son capturados y fijados en un lapso breve (Ricker, 1968). Todo el material quedó depositado en el laboratorio de Mazatlán preservado en alcohol isopropílico al 40%, y algunas submuestras fueron trasladadas al Laboratorio de Ictiología y Eco logía Estuarina -del Centro de Ciencias del Mar y Limnología de la UNAM. RESULTADOS Y DISCUSIÓN
1) COMPOSICIÓN DE LA ICTIOFAUNAEl catálogo de las especies reconocidas durante el ciclo anual de colectas en la zona de Chametla-Agua Dulce, se ha hecho siguiendo el criterio de Greenwood et al. (1966) para las categorías taxonómicas supragenéricas. Clase Chondrichthyes Orden Batoidei Suborden Myliobatoidea I Familia Urolophidae 1) Urotrygon asterias (Jordan y Gilbert) Clase Osteichthyes Subclase Teleostei División 1 Superorden Elopomorpha Orden Elopiformes Suborden Elopoidei II Familia Elopidae 2) Elops affinis Regan Orden Anguilliformes Suborden Anguilloidei III Familia Muraenidae 3) Gymnothorax dovii (Günther) IV Familia Ophychthidae 4) Ophichthus zophochir (Jordan y Gilbert) Superorden Clupeomorpha Orden Clupeoidei V Familia Clupeidae 5) Lile stolifera (Jordan y Gilbert) 6) Opisthonema libertate (Günther) 7) Opisthopterus lutipinnis (Jordan y Gilbert) VI Familia Engraulidae 8) Anchovia macrolepidota (Kner y Stein dachner) 9) Anchoa panamensis (Steindachner) 10) Anchoa mundeoloides (Breder) División III Superorden Ostariophysí Orden Siluriformes VII Familia Ariidae 11) Galeichthys caerulescens (Günther) 12) Arius liropus (Bristol) Superorden Atherinomorpha Orden Atheriniformes Suborden Exocoetoidei VIII Familia Exocoetidae (incluye Hemirhamphidae) 13) Hyporhamphus unifasciatus (Jordan y Gilbert) IX Familia Belonidae 14) Strongylura stolzmanni (Steindachner) Suborden Cyprinodontoidei X Familia Poecilidae 15) Poecilopsis latidens (Garman) Suborden Atherinoidei XI Familia Atherinidae 16) Thyrinops crystallina (Jordan y Culver) Superorden Acanthopterygi Orden Perciformes Suborden Percoidei XII Familia Centropornidae 17) Centropomus nigrescens (Gúnther) 18) Centropomus robalito (Jordan y Gilbert) 19) Centropomus pectinatus (Poey) XIII Familia Serranidae 20) Epinephelus labriformis (Venys) XIV Familia Carangidae 21) Nematistius pectoralis (Gill) 22) Caranx hippos (Linnaeus) 23) Trachinotus rhodopus (Gill) 24) Trachinotus paloma Jordan y, Starks 25) Oligoplites mundus Jordan y Starks 26) Oligoplites saurus (Bloch y Schneider) 27) Selene brevoortii. (Gill) XV Familia Lutjanidae 28) Lutjanus novemfasciatus (Gill) 29) Lutjanus argentiventris (Peters) XVI Familia Gerridae 30) Gerres cinereus Walbaum, 31) Eugerres axillaris (Günther) 32) Diapterus peruvianus (Cuvier y Valen ciennes) 33) Eucinostomus californiensis (Gill) 34) Eucinostomus sp. XVII Familia Pornadasyidae 35) Pomadasys leticiscus (Günther) 36) Pomadasys macracanthus (Günther) 37) Pomadasys branickii (Steindachner) XVIII Familia Scianidae 38) Cynoscion xanthulus Jordan y Gilbert 39) Larimus acclivis Jordan y Bristol 40) Bairdiella icistia (Jordan y Gilbert) 41) Micropogon ectenes Jordan y Gilbert 42) Menticirrhus elongatus (Günther) Suborden Mugiloidei XIX Familia Mugilidae 43) Mugil cephalus Linnaeus 44) Mugil curema Valenciennes 45) Mugil hospes Jordán y Culver Suborden Polynemoidei XX Familia Polynemidae 46) Polynemus approximans Lay y Bennet Suborden Gobioidei XXI Familia Gobiidae (incluye Eleotridae) 47) Gobiomorus maculatus (Günther) 48) Dormitator latifrons (Richardson) 49) Eleotris pictus (Kner y Steindachner) 50) Gobionellus sagittula (Günther) 51) Gobionellus microdon (Gilbert) Suborden Scombroidei XXII Familia Scombridae 52) Scomberomorus maculatus (Mitchill) Orden Pleuronectiformes Suborden Pleuronectoidei XXIII Familia Bothidae 53) Cytharichthys gilberti Jenkins y Ever man Descrita como Eucinostomus currani por Zauranech (1967), cuyo trabajo aún no ha sido publicado. Miller y Lee (1972), hacen la misma observación. La frecuencia de esta especie y otra similar, Eucinostomus entomelas Zauranech (1967), en la costa central del Pacífico mexicano hacen necesaria la descripción definitiva de ellas en un futuro próximo. Las mismas especies han sido encontradas en las lagunas costeras de Guerrero (A. Yáñez-Arancibia comunicación personal y Amezcua-Linares, 1972) . Suborden Soleoidei XXIV Familia Soleídae 54) Achirus mazatlanus (Steindachner) 55) Achirus panamensis (Steindachner) XXV Familia Cynoglossidae 56) Symphurus atricaudus (Jordan y Gilbert) Orden Tetraodontiformes Suborden Tetraodontoidei XXVI Familia Tetraodontidae 57) Sphoeroides annulatus (Jenys) Además de las especies mencionadas, existen 2 que no se registraron en este estudio, pero que sin embargo son bien conocidas en el sistema de Huizache-Caimanero. Ellas son: 58) Rypticus saponaceus bicolor Valenciennes, de la familia Serranidae. Es una especie que entra muy poco en aguas estuarinas, ya que pertenece a una familia típicamente marina y sólo lo hace cuando las condiciones ambientales son propicias, esto es durante las marcas vivas. Se le ha localizado en las inmediaciones de la Boca de Chametla. 59) Chanos chanos (Förskal) , pertenece al Orden Cetomimiformes, Suborden Chanoidei, Familia Chanidae. Esta especie conocida comúnmente como "sabalote", e introducida a la República probablemente de Hawaii (Ramírez G., 1963; Carranza, 1970), se ha adaptado perfectamente a las aguas del Pacífico mexicano desde el Golfo de California a Oaxaca. Es una especie comestible que llega a medir hasta 1.5 m como máximo, por lo que tiene importancia desde el punto de vista pesquero; penetra con regularidad a las lagunas costeras del litoral y corresponde a la especie más importante en cultivos de aguas salobres en la Región Indo Pacífica (Pillay, 1972). Kevin Warburton de la Universidad de Liverpool (comunicación personal), ha capturado ejemplares de 261 mm en la zona del tapo Botaderos en el área de estudio. 2) CARACTERIZACIÓN ICTIOLÓGICA GENERALDentro de los sistemas estuarinos de las costas del Pacífico norte, es posible reconocer una diversidad relativamente grande de especies de peces. Algunas especies pasan la mayor parte de su ciclo vital en los estuarios, otras desarrollan parte de él en éstos, y por último las hay que sólo invaden circunstancialmente este medio. Yáñez-Arancibia (1975c) , señala que las comunidades ictiológicas lagunares o estuarinas pueden estar constituidas por 6 tipos de peces, a saber: dulceacuícolas, anádromos y catádromos, estuarinos, marinos visitantes en el estado adulto para alimentarse, marinos que utilizan el estuario como áreas de crianza, y visitantes marinos ocasionales. En la figura 7 se han enumerado las familias identificadas y su proporción del número de individuos para cada uno de los meses, ordenadas de izquierda a derecha en sentido progresivo descendente, con el fin de observar la incidencia cuantitativa de las diferentes familias. Se tomó en cuenta esta categoría taxonómica (familias) debido a que siempre existe una especie más representativa cuantitativamente y en otros casos hay sólo una especie en cada tina de ellas, de manera que a este nivel taxonómico resulta más práctico hacer comparaciones numéricas de la abundancia relativa. En la figura 7 podemos observar que existe un grupo de familias que aparece prácticamente en forma invariable y en número considerable de individuos; éstos constituyen las especies típicamente estuarinas, mientras que las otras aparecen en ciertas épocas del año, como por ejemplo la familia Gobiidae -durante las lluvias. Finalmente, los visitantes ocasionales, tanto de aguas marinas, Ophychthidae, Serranidae, como de aguas dulces, Atherinidae. En la figura 8 se muestra la variación anual numérica de las principales familias, partiendo de la más abundante. Podemos observar cómo el volumen y la continuidad se reducen progresivamente en cada familia, hasta aparecer sólamente como pequeñas cimas interrumpidas en ciertas épocas del año de acuerdo a las variaciones ambientales, como las marcas y la aportación de agua fluvial. De acuerdo al criterio de Günter (1956), McHugh (1967, Yáñez-Arancibia (1975c), se puede caracterizar a la ictiofauna de HuizacheCaímanero de la siguiente manera: PECES DULCEACUÍCOLASPoecilopsis latidens Thyrinops cristallina Eleotris pictus Gobiomorus maculatus Dormitator latifrons PECES ANÁDROMOS
PECES CATÁDROMOS
PECES ESTUARINOSLile stolifera Galeichthys caerulescens Arius liropus Gobionellus sagittula Gobionellus microdon PECES MARINOS QUE UTILIZAN EL ESTUARIO COMO ÁREAS DE CRIANZACentropomus nigrescens Centropomus robalito Centropomus pectinatus Anchoa panamensis Anchoa mundeoloides Caranx hippos Oligoplites mundus Oligoplites saurus Gerres cinereus Eugerres axillaris Diapterus peruvianus Eucinostomus californiensis Eucinostomus sp. Mugil cephalus Mugil curema Polynemus approximans Cytharichthys gilberti Achirus mazatlanus Achirus panamensis Sphoeroides annulatus PECES MARINOS QUE UTILIZAN EL ESTUARIO COMO ADULTOS PARA ALIMENTARSEElops affinis Opisthonema libertate Opisthopterus lutipinnis Anchovia macrolepidota Anchoa mundeoloides Anchoa panamensis Hyporhamphus unifasciatus Strongylura stolzmanni Centropomus nigrescens Centropomus robalito Selene brevoorti Lutjanus novemfasciatus Lutjanus argentiventris Pomadasys macracanthus Pomadasys leuciscus Cynoscion xanthulus Bairdiella icistia Micropogon ectenes Scomberomorus maculatus PECES MARINOS QUE SON VISITANTES OCASIONALESUrotrygon asterias Gymnothorax dovii Ophichthus zophochir Epinephelus labriformis Nematistius pectoralis Trachinotus rodophus Trachinotus paloma Pomadasys branickii Larimus acclivis Menticirrhus elongatus Mugil hospes Symphurus atricaudus 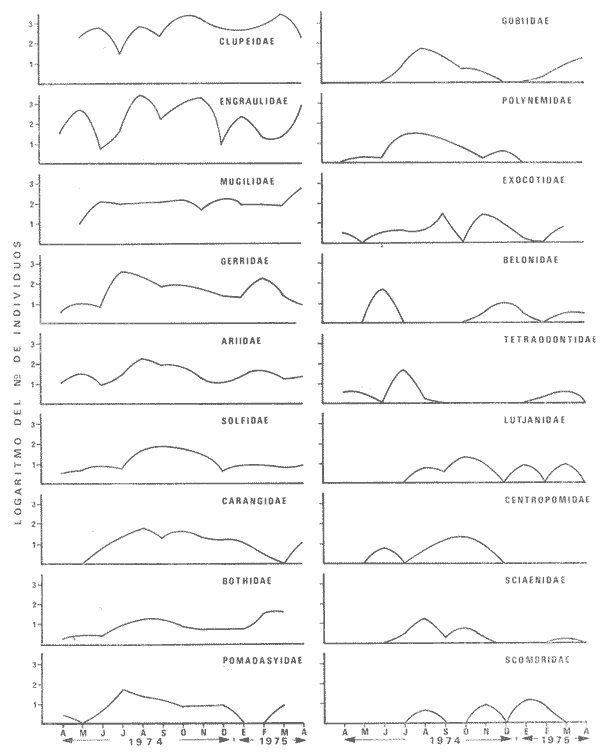 Fig. 8 Variaciónnual yabundanci relativade cada una delas familiaspresentesentre la Boca de Chametla yel Canaldel agua Dulce, Sinaloa, México En la evaluación estimativa de las poblaciones se usó un método de evaluación que fuese fácil de precisar, así se consideraron áreas y no volúmenes para la cuantificación, tal y como aparece en las figuras 9, 10 y 11, donde se muestran los diferentes resultados obtenidos con las 3 artes de pesca utilizadas para las capturas. Nótese que sólo en las capturas con chinchorro los promedios de individuos señalan números enteros por M2, mientras que con atarraya y red de arrastre los valores son de décimas en una y de centécimas en la otra. Esto indica que el arte de pesca más efectivo fue el chinchorro playero. Nótese que las 3 gráficas citadas coinciden en un pico hacia el mes de noviembre y que abarca de septiembre a diciembre, en donde se registran los máximos valores de densidad de población. Es posible que esto se deba al incremento de individuos y especies, que tiene lugar durante el abatimiento de salinidad resultante del aporte de agua dulce que proviene de los ríos durante la temporada de lluvias. Éstas se inician en los meses de julio y agosto, y como consecuencia la localidad es invadida por especies de la familia Gobiidae, entre otras, e incluso especies típicamente dulceacuícolas cuando las condiciones ambientales son favorables (Figs. 7 y 8). La caracterización de la fauna ictiológica de HuizacheCaimanero permite precisar que corresponde a una comunidad ictiofaunística con la mayor parte de los componentes comunitarios señalados por Günter (1956), Mc Hugh (1963) y Yáñez-Arancibia (1975c). 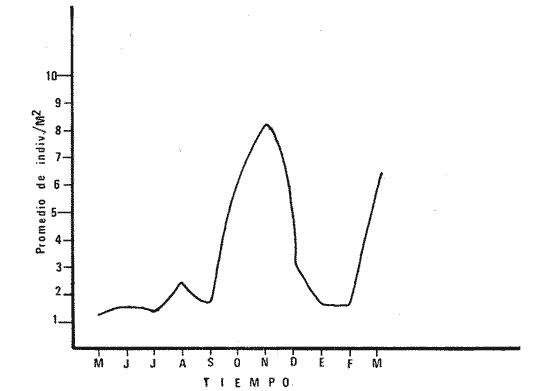 Fig. 9. Densidad relativa de peces por m2, en el área de la Boca de Chametla y Canal de Agua Dulce, recolectados con chinchorro playero, durante mayo de 1974 a marzo de 1975. 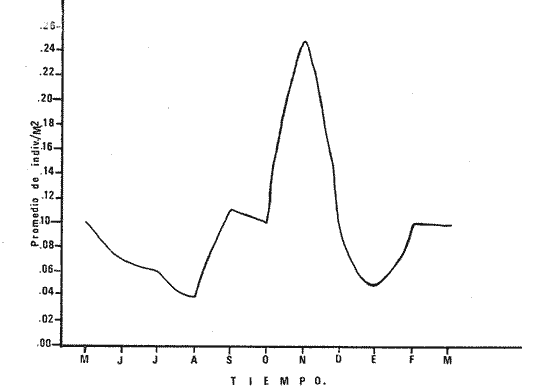 Fig. 10. Densidad relativa a peces por m2, en el área de la Boca de Chametla y Canal de Agua Dulce, recolectados con atarraya, durante mayo de 1974 a marzo de 1975. 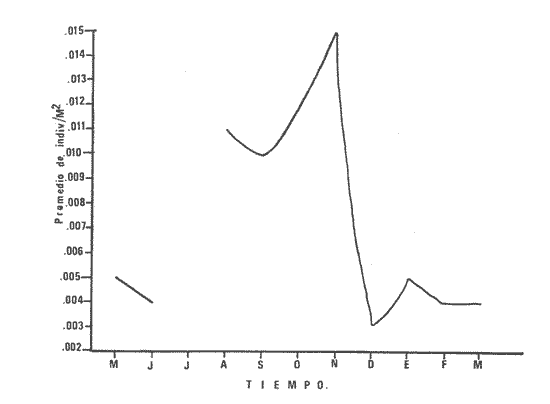 Fig. 11. Densidad relativa de peces por m2, en el área de la Boca de Chametla y Canal de Agua Dulce recolectados con red de prueba camaronera, durante mayo de 1974 a marzo de 1975. La combinación de las 3 artes de pesca ya mencionadas, permitió las colectas de aproximadamente 60 especies, de las cuales el 8.3% (5 especies) son dulceacuícolas, el 8.3% (5 especies) peces típicamente estuarinos, el 33.3% (20 especies) corresponde a peces marinos que utilizan el estuario como áreas naturales de crianza, el 30.1% (19 especies) corresponde a peces marinos que visitan el estuario generalmente como adultos y para alimentarse, y finalmente las comunidades también están constituidas por peces marinos que son visitantes ocasionales o circunstanciales a los que les corresponde el 20% (12 especies), lo cual no puede pasar desapercibido aun cuando son poco significativos en número y biomasa. El predominio de uno u otro de los componentes ictiofaunísticos de la comunidad descrita depende de: a) la localidad , b) la estación de] año, c) las condiciones físico-químicas del agua y d) la disponibilidad de alimento. 3) OBSERVACIONES SOBRE GALEICHTHYS CAERULESCENS (GÜNTHER)Esta especie se conoce comúnmente en Huizache-Caimanero con el nombre de "chihuil", pero en las costas de Guerrero se le denomina cuatete" (Yáñez-Arancibia et al., 1976) . El chihuil" es una especie regularmente abundante en el sistema, sobre todo en la laguna de Caimanero y las inmediaciones del tapo del mismo nombre donde se les localiza en grandes cardúrnenes de cientos de individuos, que generalmente se reúnen atraídos por el alimento que ahí se encuentra cuando los tapos están en operación. Por lo general estos peces no son consumidos y son rechazados por los pescadores cuando llegan a aparecer en sus redes, excepto durante la cuaresma en que comerciantes del centro del país los adquieren, o bien cuando la cantidad es suficiente para hacer fertilizantes. Algunas veces también se les utiliza para preparar harina, aunque tienen el inconveniente de ser muy grasosos. Por el contrario, en la costa de Guerrero constituye una de las especies de importancia comercial más significativas (Yáñez-Arancibia et al., 1976). En la zona estudiada en este trabajo, los chihuiles se encuentran presentes todos los meses del año aunque no en la misma cantidad, como lo muestra la figura 12 donde se pueden observar 3 cantidades principales, la más alta en febrero, la otra en septiembre y la última en mayo. Con la información disponible es difícil explicar este hecho, sin embargo la abundancia registrada en septiembre y octubre puede deberse a que en el primero de estos meses se lleva a cabo el desove. Las otras dos cantidades aparentemente no tienen una explicación razonable sin embargo la mayor abundancia podría estar determinada por la presencia de alimento, ya que son peces muy voraces. El resto del año hay una población más o menos uniforme. Es posible que la relativa abundancia que generalmente muestra esta especie esté asociada directamente con el mecanismo de defensa con que cuenta. Este consiste, aparte de sus hábitos bentónicos, en la presencia de fuertes espinas ponzoñosas con una sierra inversa en su borde posterior en las aletas pectorales y dorsal, las cuales levanta a voluntad cuando se siente en peligro y cuya picadura es muy dolorosa para el hombre. De este modo constituye un alimento poco apetecible para cualquier depredador, y al menos en el sistema lagunar no se ha podido reconocer alguno de importancia, a excepción del hombre. Yáñez-Arancibia et al. (1976) ha señalado anteriormente que Galeichthys caerulescens no sufre presiones de predación por parte de otros peces, pero sí es un "exportador de energía" del ecosistema al ser predado por el hombre y ciertas aves. 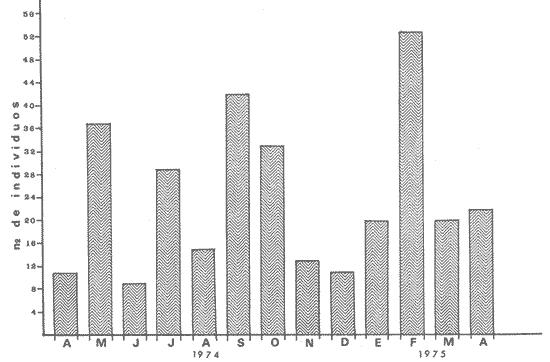 Fig. 12. Abundancia relativa anual de Galeichthys caerulescens en el sistema lagunar costero de Huizache-Caimanero, Sinaloa, México. En las figuras 13 y 14 se muestra la relación peso-longitud obtenida con las mediciones de todas las colectas y después de haber recibido conversión logarítmica con el fin de obtener figuras lineales y calcular dicha relación. En la figura 13 la escala logarítmica agrupa a la mayoría de los puntos referentes al peso fresco, mas no sucede así en los que representan el peso seco, cuyos puntos se encuentran ligeramente más dispersos. También se puede observar que la mayor parte de los puntos se encuentran localizados entre los 150 y 500 g para peso fresco y entre los 35 y 180 g para peso seco, y de 220 a 380 mm de talla en ambos pesos. La apariencia de una sola línea parece indicar que el patrón de incremento en peso es el mismo para los individuos mayores de 200 mm; no sucede lo mismo con los ejemplares pequeños según lo muestra la figura 14 donde aparecen 2 líneas diferentes que representan un incremento más rápido en peso en fases jóvenes y otras más lenta que se mantiene prácticamente estable hasta el estado adulto en los individuos más grandes. Por el contrario, González-Villaseñor (1972) y Yáñez-Arancibia et al. (1976) han indicado que el incremento en peso es mucho mayor después de los 200 mm en relación a la talla que no crece con tanta rapidez. Al estado actual de estos estudios no tiene explicación este hecho, pero parece razonable suponer que el incremento en peso después de los 200 mm se debe al desarrollo gonádico. Se efectuaron curvas a partir de muestras de las poblaciones que se encontraron en la zona de trabajo. En la figura 15 aparecen los resultados considerando el porcentaje del número total de individuos debido a la amplia variación registrada en cada mes; en ella podemos observar que no existe desplazamiento en las modas y siempre se encuentra mayoría de individuos de una talla determinada entre los 226 mm a 325 mm en cada uno de los ciclos mensuales. Esto de hecho no indica que las poblaciones no crezcan, sino que hay una constante sucesión de las mismas en el área, resultado de las migraciones que indudablemente efectúan estos peces dentro del sistema. En la figura 16 aparece la suma anual de todos los individuos y se observa la existencia de una sola moda localizada en las tallas más frecuentes a lo largo del año, siendo comparativamente muy escasas el resto de las tallas, lo que viene a corroborar la anterior afirmación en el sentido de que en las inmediaciones del estuario de Chametla y en el canal de Agua Dulce, hasta el tapo, existen por lo regular poblaciones sucesivas de una talla determinada, emigrando posteriormente por causas aún no bien conocidas, aparentemente con dirección a la laguna. Yáñez-Arancibia et al. (1976) señala que G. caerulescens busca salinidades bajas para desovar, y de acuerdo a eso es posible que la especie migre al interior de la laguna durante la época de reproducción. 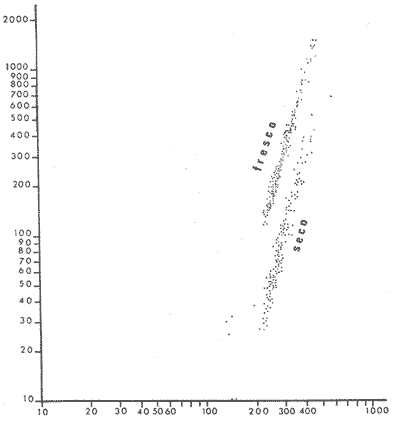 Fig. 13. Relación peso-longitud de Galeichthys caerulescens. 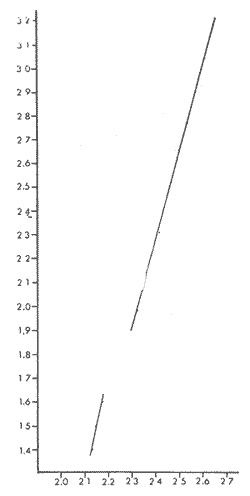 Fig. 14. Promedios logarítmicos de la relación peso-longitud en Galeichthys caerulescens. Como se muestra en la figura 17 el desove de esta especie en el área tiene lugar en los meses de agosto y septiembre, en los cuales es mayor la incidencia de hembras maduras, e inmediatamente después, de machos desovados. Es posible que el desove se efectúe con más intensidad en otras partes del sistema, dada la poca cantidad de ejemplares localizados en el área de Chametla y Agua Dulce. El desove también puede ser detectado por la presencia, en temporadas inmediatas, de gran número de juveniles (Fig. 15). 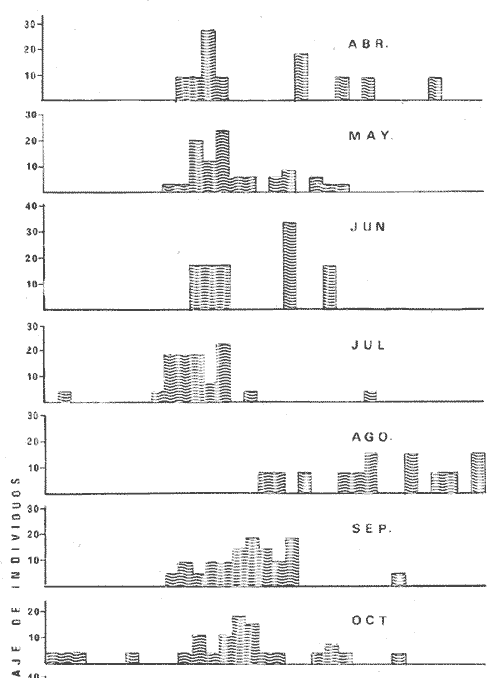 Fig. 15. Frecuencia de tablas en Galeichthys careulescens durante el ciclo anual. Se observa la continua incidencia de tallas determinadas. 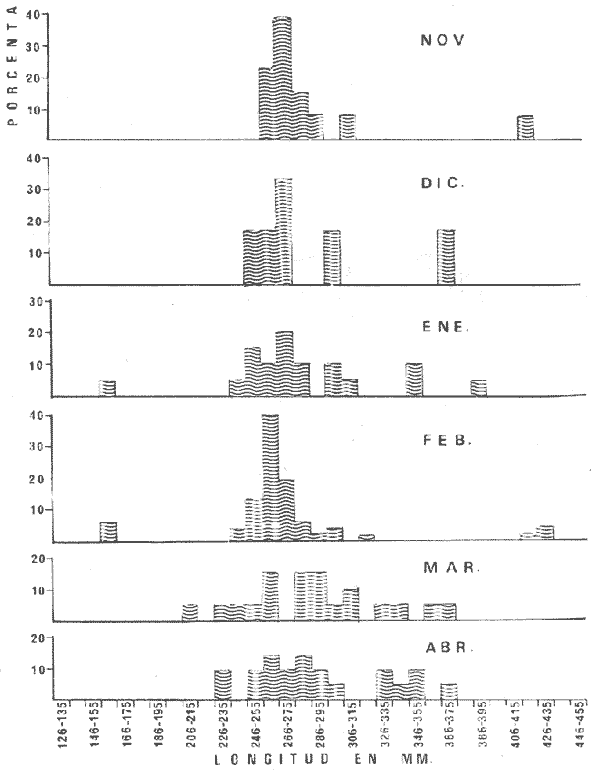 Fig. 15. Frecuencia de tablas en Galeichthys careulescens durante el ciclo anual. Se observa la continua incidencia de tallas determinadas. 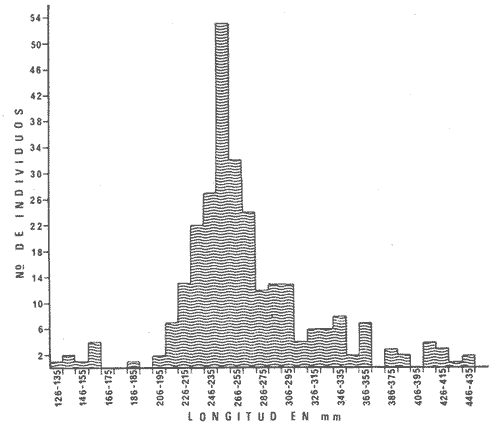 Fig. 16. Tallas más frecuentes registradas durante todo el ciclo anual de colectas donde se observa que la mayoría de los individuos de Galeichthys caerulescens se encuentran entre los 216 y 315 mm de longitud. En el resto del ciclo anual sólo se detectaron las 3 primeras fases de desarrollo, sobresaliendo por su incidencia la fase 111. Aun cuando de esto se deriva que, para La especie sólo existe un desove -durante su ciclo anual de vida en la zona de muestreo, en otras localidades del sur como las lagunas costeras de Guerrero, Yáñez-Arancibia et al. (1976) encontraron 2 épocas de desove para esta misma especie, una en mayo y otra en octubre. Las gónadas femeninas maduras de ejemplares de 425 mm y 950 g alcanzan por lo regular un peso entre 150 y .900 a con una longitud de 10 a 12 cm ocupando la mayor parte de la cavidad celómica. Cada una de ellas contienen de 50 a 60 huevecillos de aproximadamente 1.5 cm de diámetro y de color amarillo yema, lo que demuestra la gran cantidad de vitelo que contienen. Esto corrobora las observaciones de Yáñez-Arancibia et al. (1976) en que no es una especie fecunda en sentido estricto, pero su adaptación para sobrevivir consiste en los especiales hábitos reproductivos que posee. Sin embrago el tamaño de los huevos que menciona Yáñez-Arancibia et al. (1976) son mucho menores. 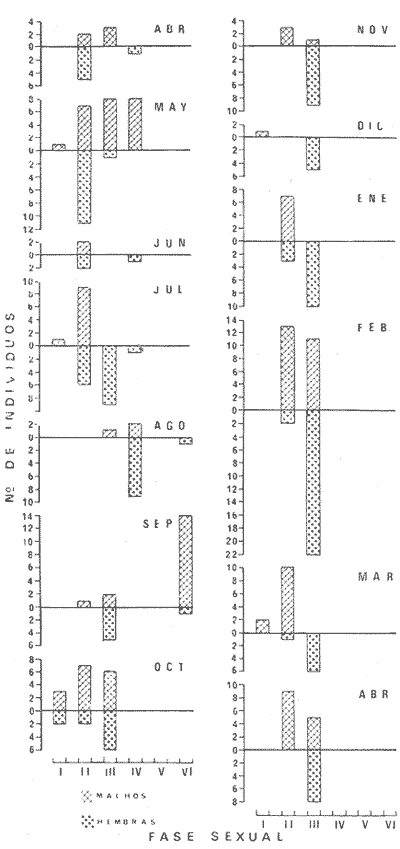 Fig. 17. Desarrollo sexual y proporción de sexos mensual de Galeichthys caerulescens, durante el ciclo anual de colectas, donde se aprecia que el desove tiene lugar durante agosto y septiembre El desarrollo de las gónadas generalmente está asociado a la aparición de caracteres secundarios, siendo el más común la diferencia en el tamaño, como sucede en esta especie, donde la hembra madura es mayor por algunos centímetros que el macho; pero también las hembras presentan otro carácter que es la presencia de una formación carnosa en la base superior de las aletas pélvicas que va agrandándose conforme transcurre la madurez, acompañada de un obscurecimiento paulatino en dicha zona. Yáñez-Arancibia et al. (1976) discuten en extenso la reproducción de Galeichthys caerulescens en lo referente a dimorfismo sexual, forma de reproducción, tipo y época de reproducción, fecundidad y desarrollo embrionario. La talla promedio de los juveniles se encuentra entre los 4 y 5 cm, igual al señalado por Yáñez-Arancibia et al. (1976). La proporción entre machos y hembras es de 1 : 1 durante toda la colecta anual. Los chihuiles son peces muy voraces con cierta tendencia a incluir animales en su dieta. Se ha observado en algunas ocasiones que llegan a ingerir carroña y materia orgánica de desecho. Por lo tanto se parte del hecho de que no presentan una selección o preferencia por determinada clase o tipo de alimento y que frecuentemente consumen lo que se encuentra disponible en el medio. Esto se puede corroborar observando la tabla 2, donde se muestra una variedad notable de grupos zoológicos y algunos vegetales pero con predominancia de los artrópodos, tanto en número de ejemplares ingeridos como en frecuencia y en volumen; le siguen los anélidos y dentro de ellos principalmente los poliquetos. Los camarones representan una cifra alta en el número de individuos porque están representados en su mayoría por postlarvas y juveniles, lo que se refleja en el bajo volumen obtenido. Yáñez-Arancibia et al. (1976), han señalado que esta especie es un carnívoro consumidor de segundo y tercer orden. Además hace un análisis detallado de la alimentación, hábitos alimenticios y relaciones tróficas. En el sistema Huizache-Caimanero ingiere grandes cantidades de camarón, pero su preferencia alimenticia depende de la disponibilidad de alimento, estación del año y la localidad. En el trayecto comprendido entre la Boca de Chametla y el tapo de Agua Dulce, existe una migración intensa de los individuos de esta especie durante el año y presuntamente lo hacen a lo largo de todo el sistema. Existen fundamentos para considerar que dichas migraciones son por alimento y se llevan a cabo de las partes bajas del estero hacia la laguna donde se desarrolla el camarón. Por lo que se refiere a las migraciones por desove, no se tienen datos precisos acerca de que las efectúen como sucede en otras especies en el mismo sistema. Sin embargo, como se ha mencionado, las migraciones existen dentro del sistema, principalmente desde el estero hacia la laguna. Tales migraciones se ven aparentemente interrumpidas, en una magnitud desconocida, por los obstáculos físicos que representan los tapos. 4) OBSERVACIONES SOBRE MUGIL CUREMA VALENCIENNESEsta especie, conjuntamente con Mugil cephalus, constituye uno de los principales recursos pesqueros que se desarrollan en aguas estuarinas, de ahí la importancia de los estudios para conocer el ciclo biológico de estas especies (Stuardo et al., 1974; Yáñez-Arancibia 1975a, 1975b) . Las lisas representan una de las especies más abundantes y mejor distribuidas dentro del sistema Huizache-Caimanero, siendo posible encontrarlas en mayor o menor número durante todos los meses del año. Como se observa en las gráficas de abundancia (Figs. 7 y 8), la familia de los mugílidos ocupa el tercer lugar numérico. Sin embargo volumétricamente le corresponde. el primero, ya que las familias que la superan en el primer sentido, Clupéidos y Engraúlidos, están integrados por especies de pequeño tamaño y peso, no comparables con las que alcanza la especie aquí discutida. La figura 18 muestra la abundancia relativa de la especie donde se distinguen una serie de altas y bajas en las columnas que son difíciles de interpretar. Sin embargo se ha visto que la mayor captura comercial de la lisa, de ambas especies, se lleva a efecto al finalizar la temporada de lluvias cuando el embalse está a su máxima capacidad, temporada en la cual la pesca del camarón se encuentra intensificada. -Por lo que, en apariencia, existe una relación directa entre peces y crustáceos que bien pudiera ser una competencia entre unos y otros como lo ha demostrado Yáñez-Arancibia (1975b). 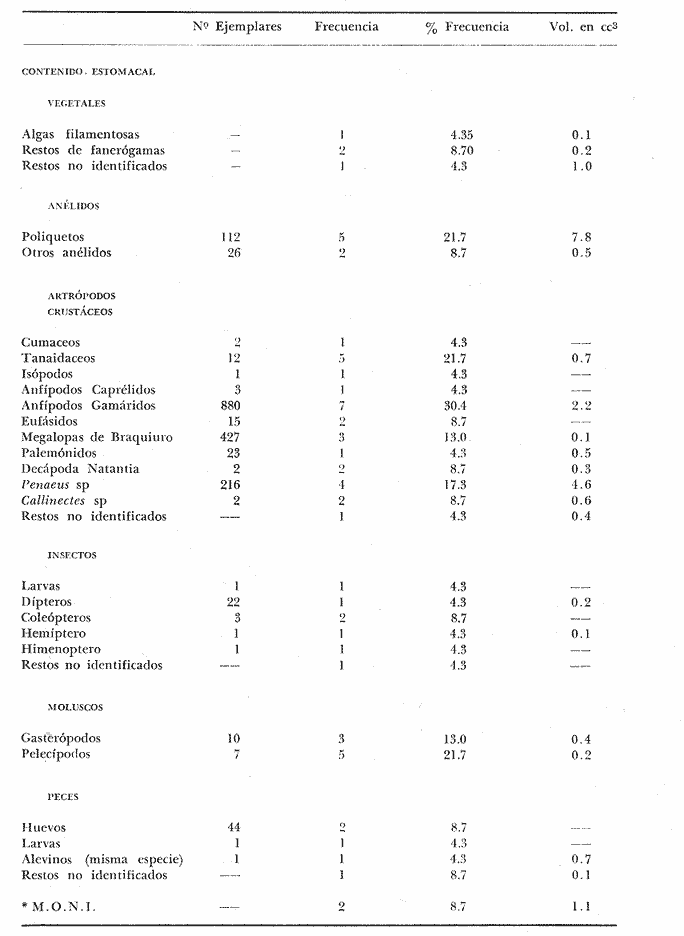 TABLA 2 RESULTADO DE 23 ESTÓMAGOS ANALIZADOS DE GALEICHTHYS CAERULECENS EN HUIZACHE CAIMANERO, SINALOA, MÉXICO 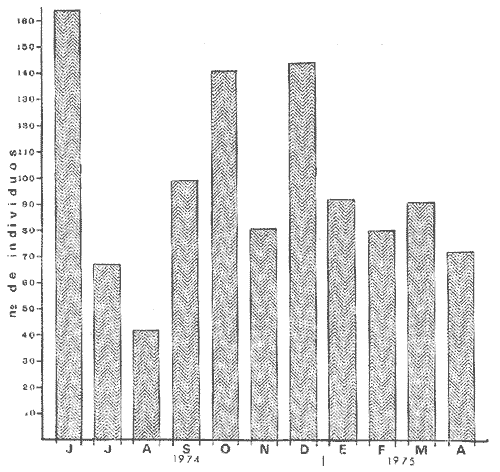 Fig. 18 Abundancia relativa anual de Mugil curema en el sistema lagunar costero de Huízache- Caimanero, Sinaloa, México. En la figura 19 se muestra la relación peso-longitud que se presenta en esta especie a través de la escala logarítmica, en ella se observa como la mayoría de los puntos inciden sobre una línea recta teórica en el peso fresco y se encuentran un poco más dispersos en el peso seco. Tanto en esta especie como en la anterior, se obtuvieron resultados más uniformes' con peso fresco que con peso seco. En la figura 20 de logaritmos promedios se visualiza mejor la recta en ejemplares que pasan los 6 g y los 75 mm; en individuos menores parece existir una relación diferente, pero no hay suficientes puntos para mostrarlo con claridad. En la figura 21 se observa el movimiento de las tallas modales a través del tiempo; este desplazamiento de las modas no sigue una secuencia regular, sino que va sufriendo una, serie de cambios; algunas veces el desplazamiento es rápido y otras lento, lo que puede explicarse dado que el crecimiento no es uniforme, es más rápido en los juveniles y decrece paulatinamente hasta llegar a estabilizarse. En la figura 22 se muestran las 6 diferentes modas registradas durante el ciclo de colectas; en ella se ve que la forma de las curvas está de acuerdo con lo anterior; sin embargo, 3 de las líneas modales están interrumpidas y se nota en ellas una tendencia a converger en un punto imaginario extrapolando las líneas indefinidamente, lo cual significa que hay una tendencia hacia la estabilización del crecimiento.  Fig. 19 Relación peso-longitud de Mugil curema. El desove tiene lugar hacia la segunda mitad de la primavera, ya que en abril es cuando se detecta una mayor proporción de ejemplares maduros, lo mismo en mayo, pero se tienen datos insuficientes en este mes. Es de notarse la completa ausencia de fases V y VI correspondientes a peces en desove y desovados, pero esto se debe a que lo efectúan fuera del sistema para lo que tienen que realizar migraciones hacia el mar, por lo que prácticamente la totalidad de las lisas maduras se localiza en las inmediaciones de la boca con tránsito hacia el mar. Es conocido por otros trabajos sobre diferentes especies de lisas en otros países, que el desove tiene lugar en mar abierto sobre la plataforma continental (Anderson, 1957; Caldwell y Anderson, 1959; Thompson, 1954; Jacot, 1920; Yáñez-Arancibia 1975a, 1975b). La época de desove se puede corroborar con la presencia de numerosos juveniles en el mes de abril (Fig. 21) . El resto del año se mantiene una población más o menos regular en la que predomina la fase 1 inmediatamente después del desove, que parece ser el único durante todo el ciclo anual. 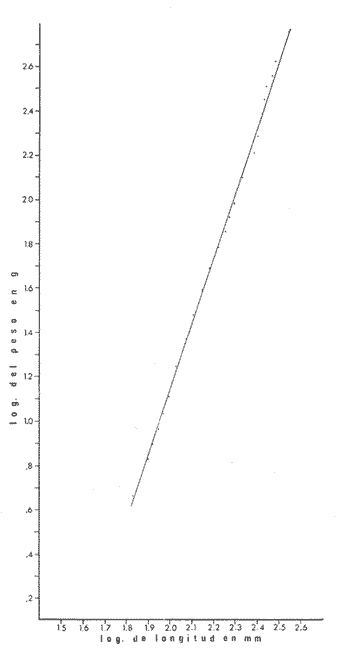 Fig. 20. Promedios logarítmicos de la relación peso-longitud de Mugil curema. 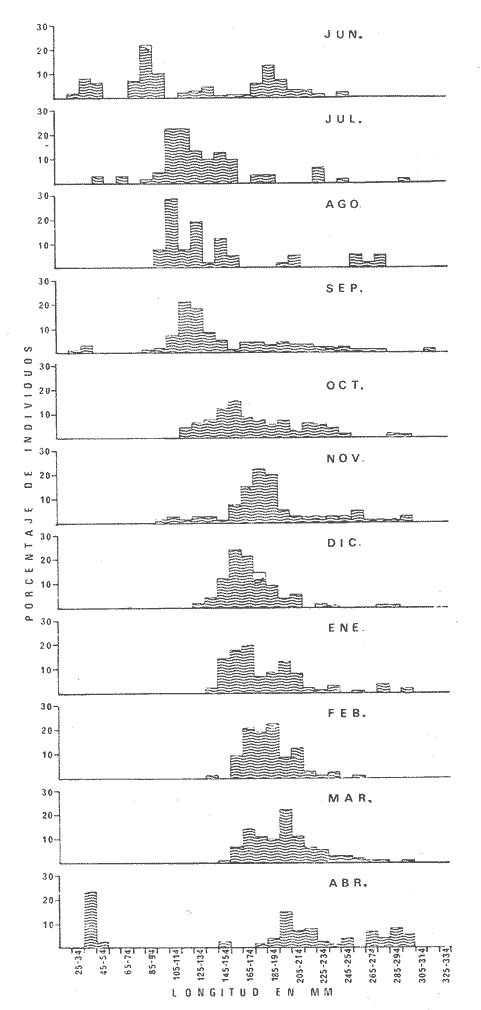 Fig. 21. Frecuencia de tallas en Mugil curema durante el ciclo anual. Se observa el movimiento de las modas. 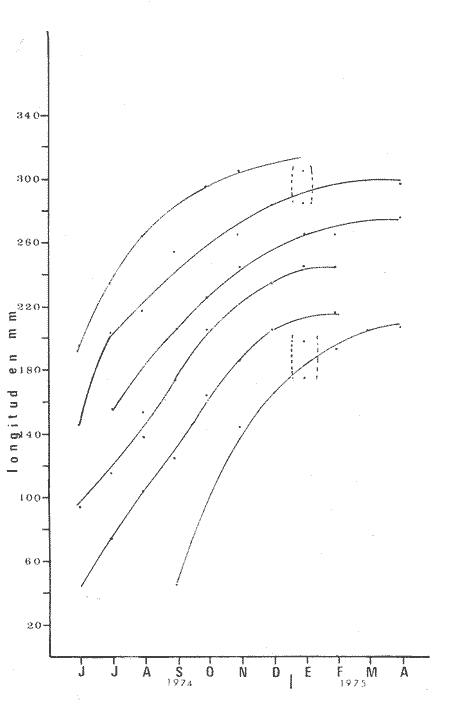 Fig. 22. Diferentes modas registradas a través del ciclo anual en Mugil curema Supuestamente tanto la fecundación como el completo desarrollo embrionario se lleva a cabo en aguas marinas, así para cuando entran a la Boca de Chametla hacia el mes de junio principalmente, son ya juveniles de 14 mm de longitud y 1 g de peso. La proporción de sexos anual entre machos y hembras es de 1.4:1 (Fig. 23). La lisa por lo regular consume la materia del fondo que se encuentra en el fango de los estuarios y esteros, del cual toma detritus orgánicos, algunas algas y pequeños. organismos conjuntamente con los sedimentos. Por esta causa se ha considerado que vive en estrecha relación con los camarones que viven en el fondo posiblemente comiendo lo mismo. De aquí se deriva la idea de que puedan ser competidores del crustáceo por alimento y espacio. Este punto ha sido discutido en otras áreas por Carranza (1970) y Yáñez-Arancibia (1975b). 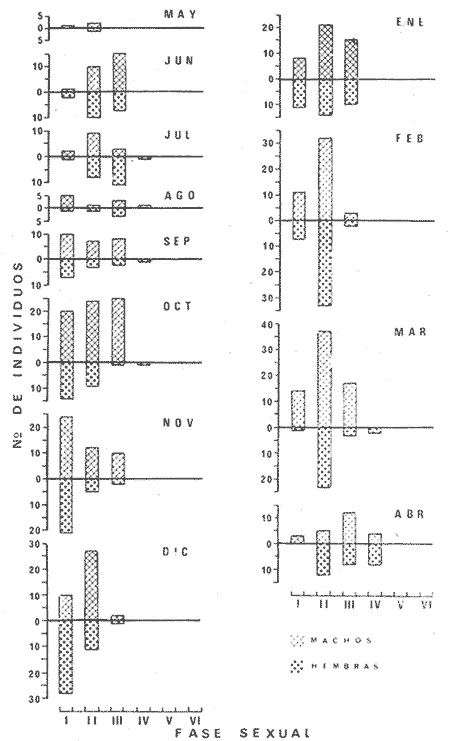 Fig. 23. Desarrollo sexual y proporción de sexos de Mugil curema durante el ciclo anual de colectas. Se nota la completa auscencia de fases V y Vi Las lisas presentan dos clases de migraciones perfectamente definidas: por alimento y por desove. Las migraciones por alimento las efectúan una vez que eclosiona el huevo en el mar abierto y es capaz de desplazarse buscando las bocas de los ríos y penetrando en estuarios, esteros y lagunas costeras, donde pasa la mayor parte del ciclo anual alimentándose y creciendo hasta alcanzar la madurez sexual cuando busca nuevamente las aguas marinas, abandonando definitivamente los estuarios para regresar a las áreas de desove. Actualmente se desconoce por completo la localización de tales áreas y el trayecto que siguen las larvas para alcanzar las aguas estuarinas en cuyo lapso se transforman en juveniles. Esto hace necesario en el futuro un estudio de la plataforma continental frente a las bocas de los sistemas estuarinos. La razón por la que esta especie busca las aguas estuarinas para su desarrollo, se puede deber a que éstas tienen una alta productividad como consecuencia de los aportes de aguas continentales que acarrean nutrientes inorgánicos, además de la alta disponibilidad de alimentos (detritus) y protección que tienen contra los depredadores (Yáñez-Arancibia 1975b). Los tapos no son un obstáculo para las migraciones por desove y alimentación que realizan estos peces, porque en la temporada que las efectúan, dichos obstáculos no se encuentran en operación, esto es, entre los meses de abril y julio que es principalmente cuando hacen sus movimientos migratorios. Conclusiones1. La fauna ictiológica del sistema lagunar de Huizache-Caimanero está compuesta por 27 familias, 46 géneros y 60 especies comunes. 2. Estas comunidades son características de los sistemas lagunares y/o estuarinos de la costa del Pacífico mexicano presentando el 8% de peces dulceacuícolas, el 8% de peces típicamente estuarinos, 30% de peces marinos que visitan el estuario generalmente como adultos y para alimentarse, el 33% de peces marinos que utilizan el estuario como áreas naturales de crianza y el 20% de peces marinos que son visitantes ocasionales. No se encontraron peces anádromos ni catádromos. 3. Las diferentes especies en particular y las comunidades en general sufren variaciones en su composición y abundancia relativa de acuerdo a: 1) las condiciones hidrológicas del sistema, consecuencia de 2) la estación del año, y 3) la localidad dentro del sistema y 1 sus gradientes de salinidad. 4. Las familias mejor representadas en diversidad son: Carangidae (7 especies), Gerridae (5 especies) , Sciaenidae (5 especies) , Gobiidae (5 especies) ; y con 3 especies las familias: Mugilidae, Centropromidae, Pornadasyidae, Engraulidae y Clupeidae. 5. Las especies mejor representadas en NÚMERO durante la mayor parte del año son: Lile stolifera, Anchoa panamensis, Mugil curema, Galeichthys caerulescens, Achirus mazatlanus, Diapterus peruvianus, Eucinostomus spp., Pomadasys macracanthus, Cytharichthys gilberti y Caranx hippos. 6. Las especies mejor representadas en BIOMASA durante la mayor parte del ciclo anual son: Mugil curema, Mugil cephalus, Galeichthys caerulescens, Pomadasys macracantus, Diapterus peruvianus, Cynoscion xanthulus y Achirus mazatlanus. 7. Galeichthys caerulescens es una especie típicamente estuarina que completa todo su ciclo de vida en el interior del sistema lagunar, de hábitos bentónicos y predominantemente carnívoros, incluyendo camarones del género Penaeus spp. otros crustáceos y peces en su dicta, con volumen y frecuencia significativos. Realiza migraciones buscando alimento. y durante la época de reproducción. 8. Mugil curema es una especie marina que utiliza el estuario como área natural de crianza para protegerse, alimentarse (de detritus) y crecer. Grandes cardúmenes de juveniles (15 a 18 mm) penetran desde el mar en los meses de junio y julio y abandonan el estuario al inicio de la primavera del año subsiguiente para volver al mar definitivamente. La época de desove en el mar se efectúa entre los meses de marzo y mayo. 9. Al estado actual de las investigaciones se hacen necesarios estudios: 1) biológico-pesqueros y 2) de relaciones tróficas de las especies de importancia comercial con el objeto de: a) recomendar una correcta administración de los recursos pesqueros y b) determinación del rol ecológico de los peces y la tendencia del flujo energético en las comunidades del sistema lagunar de HuizacheCaimanero y sus perspectivas de óptimo aprovechamiento. AgradecimientosEl autor agradece a las siguientes personas: Alfredo Laguarda Fígueras, Director del Centro de Ciencias del Mar y Limnología de la UNAM, por el apoyo y las facilidades otorgadas durante el desarrollo del presente trabajo. Alejandro Yáñez-Arancibia por la gran ayuda y asesoría en la integración del trabajo y la revisión crítica, del manuscrito. Alberto Ramírez Flores jefe de la Estación Mazatlán del Centro de Ciencías del Mar y Limnología de la UNAM, por su apoyo y acertada revisión del manuscrito. Gerardo Green y Richard Nugent por la revisión del manuscrito y sus valiosos consejos y sugerencias. Efraín Chavarín por la ayuda en el trabajo de laboratorio y campo. Este estudio fue financiado por el Programa de Investigación Científica de Colaboración celebrado entre la Universidad Nacional Autónoma de México, a través del Centro de Ciencias del Mar y Limnología, y el Consejo Británico a través de la Universidad de Liverpool, Inglaterra. LITERATURAÁLVAREZ, V. J. Peces mexicanos (Claves). Com. Nal.. Cosul. Pes. I.N.I.B.P. 1970 166 p. AMEZCUA-LINARES, F. Tesis Profesional. Aportación al conocimiento de los peces del sistema de Agua Brava, Nayarit. Fac. Ciencias. Univ. Nal. Autón. México. 1972 209 p. ANDERSON, W. W. Early development spawning, growth, and occurr ence of the silves mullet (Mugil cephalus), along the South Atlantic coast of the Uníted States. Fish Bull. U. S. Fish Wildl. Serv. 1957 501-519 58 (144) ARENAS-FUENTES, V. 29 Informe del Contrato de Estudios No. EI-69-93 Informe final de las investigaciones correspondientes a hidrología y productividad en los planes piloto Escuinapa y Yavaros. Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1970 191-233 AYALA-CASTAÑARES, A., M. GUTIÉRREZ y V. M. MALPICA Informe final de los estudios de Geología Marina en las regiones de Yavaros, Son., Huizache y Caimanero, Sin. y Agiabampo, Sin., durante la primera etapa. 29 Informe del Contrato de Estudios Nº EI-69-93 Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1970 3-190 CABRERA, J. Informe sobre el programa de biología del camarón en los planes piloto Escuinapa-Yavaros. 29 Informe del Contrato de Estudios Nº EI-69-93 Univ. Nal. Autón. México,-Inst. Biol. Depto. Cienc. del Mar Limnol. y S. R. H. 1970 384-409 CALDWELL, D. K. y W. W. ANDERSON Off shore occurrence of larval silver mullet Mugil curema in the Western Gulf of Mexico Copeia 1959 252-253 3 CARRANZA, J. Reconocimiento preliminar de la fauna ictiológica en las áreas de los planes piloto Escuinapa y Yavaros y programa de las investigaciones. ler. Informe del Contrato de Estudios EI-69-51 Univ. Nal. Autón México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1969a 31 p. ---, 29 Informe de Contrato de Estudios Nº EI-69-51 Primer informe sobre el estudio taxonómico de la fauna de peces de los planes piloto Escuinapa y Yavaros. Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol, y S. R. H. 1969b 39 p. ---, 3er. Informe de Contrato de Esudios-EI-69-51 Informe preliminar sobre la alimentación y hábitos alimenticios de las principales especies de peces de las zonas de los planes piloto Yavaros y Escuinapa. Univ. Nal. Autón. México Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1969c 50 ---, Informe final Informe final sobre la primera etapa del estudio de la fauna ictiológica y depredaciones del camarón en las lagunas y esteros de los planes piloto Escuinapa, Sin. y Yavaros, Son. Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1970 28 p. CARRANZA, J. y F. AMEZCUA-LINARES, . , : . 2a. Parte del 29 Informe de Contrato de Estudios Nº NAY-EST-7 Plan Nayarit, S. R. H. Informe de avance de trabajo, incluyendo información ecológica básica datos de distribución de postlarvas e informe preliminar sobre la fauna ictiológica. Univ. Nal. Autón. México Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1971a 58-80 CARRANZA, J. y F. AMEZCUA-LINARES 2a. Parte del Informe Final de Contrato de Estudios Nº NAY-EST-7 Plan Nayarit, S. R. H. Resultados finales de hidrología, plancton y fauna ictiológica en el sistema Teacapán-Agua Brava (octubre 1970-junio 1971) Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1971b 88-115 CASTRO, J. L. Aspectos zoogeográficos de los peces marinos que penetran a las aguas continentales de México. Mem. VI Congr. Latinoam. Zool. México (en prensa) . 1974 CASTRO, J-., L., MA. DE J. PARRA y F. DE LA CHICA Los peces de las lagunas oriental y occidental, Oaxaca, y sus relaciones con la temperatura y salinidad. Mem. V Congr. Nac- Oceanogr., México (en prensa). 1974 CHAPA-SALDARA, 14. y R. SOTO-LÓPEZ Resultados preliminares del estudio ecológico y, pesquero de las lagunas litorales del sur de Sinaloa, México. Ayala-Castañares, A. y F. B. Phleger Lagunas Costeras, un Simposio. Mem. Simp. Intern. Lagunas Costeras, UNAM-UNESCO Nov. 28-30, 1967 1969 653-670 DARNELL, R. M. Fishes of Rio Tamesí and Related Coast al Lagoons in East-Central México. Inst. Mar. Sci. Univ. Tex. 1962 299-365 8 GARCÍA-CUBAS, A. 29 Informe del Contrato de Estudios Nº EI69-93 Informe sobre el avance de trabajo en el programa de estudio de los moluscos en los planes piloto Yavaros v Escuinapa. Univ. Nal. Autón México. Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1970 347-371 GARCÍA, E. Modificaciones al sistema de clasificación climática de Koppen. Instituto de Geografía, Univ. Nal. Autón. México. 1973 246 p. GÓMEZ-AGUIRRE, S., 11 C. FLORES-COTO, S. LICEADURÁN, H. SANTOYO y A. MARTÍNEZ 20 Informe del Contrato de Estudios Nº EI69-93 Informe final de la 2a. Etapa de los estudios del plancton en los planes piloto Escuinapa-Yavaros. Univ. Nal. Autón. México, Inst. Biol. Depto. Cienc. del Mar y Limnol. y, S. R. H. 1970 234-286 GÓMEZ-AGUIRRE, S., S. LICEA-DURÁN y C. FLORESCOTO Ciclo anual del plancton en el sistema Huizache-Caimanero, México (1969-1970). An. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México 1974 83-98 1 (1) GONZÁLEZ-VILLASEÑOR, L. I. Tesis profesional Aspectos biológicos y distribución de algunas especies de peces de la familia Arilidae de las lagunas litorales del noroeste de Mexico. Fac. Ciencias, Univ. Nal. Autón. México 1972 88 p. GREENWOOD, P. H., D. E. ROSEN, S. H-. WEITZMAN Y G. S. MYERS Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bull. Am. Mus. Nat. Hist. 1966 341-455 131 (4) GUNTER, G. A revised list of eurihalin fishes of north and middle America. Amer. Midl. Nat. 1956 345-354 56 (2) JACOT, A. P. Age, growth and scale characters of the mullet Mugil cephalus and M. curema. Trans. Amer. micr. Soc. 1920 199-229 39 McHUGH, J. L. Estuarine nektom G. H. Lauff Estuaries Publ. Amer. Ass. Adv. Sci. 1967 581-619 83 MILLER, J. D. y R. N. LEA Guide to the coastal marine fishes of California. Fish. Bull. Calif. Dept. Fish and Game, Fish. 1972 1-235 157 NIKOLSKY, G. V. The Ecology of Fishes. Academic Press, London New York 1963 352 p. ORTEGA, G. M. 29 Informe del Contrato de Estudios Nº EI-69-93 Informe de avance del estudio de la vegetación sumergida en los planes piloto Yavaros y Escuinapa. Univ. Autón. México Inst. Biol. Depto. Cienc. del Mar y Limnol. y S. R. H. 1970 287-346 PILLAY, T. V. R. Coastal Aquaculture in the IndoPacific Region. PILLAY, T. V. R. Fishing News (Books) Ltd., England, (FAO) 1972 497 p. RAMIREZ, R. Estudio ecológico preliminar de las lagunas costeras cercanas a Acapulco. Revta. Soc. mex. Hist. Nat. 1952 199-218 13 ---, Nociones sobre Hidrología Aplicada a la Pesca. S. I. C., Com. Nal. Cosul. Pes. I. N. I. B. P. 1963 361 p. RESÉNDEZ, A. Estudio de los peces de la laguna de Tamiahua, Veracruz, México. An. Inst. Biol. Univ. Nal. Autón. México, Ser. Cienc. del Mar y Limnol. 1970 79-146 41 (1) ---, Estudio de los peces de la laguna de Alvarado, Veracruz, México. Revta. Soc. mex. Hist. Nal. 1973 183-281 31 RICKER, W. E. Methods for assesment of fish production in fresh wather. Blackwell Scientific Pub. 1968 225 p. STUARDO, J., A. MARTINEZ, L. A. YÁÑEZ, J. A. WEINBORN y J. RUIZ Prospección de los recursos biológicos y pesqueros del sistema lagunar de Guerrero, y, en parte, del litoral rocoso de Michoacán. An. Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México (en prensa). 1974 TAMAYO, J. Geografía general de México. Talleres gráficos de la Nación. 1949 1: 1-628, 2: 1-580 THOMPSON, J. M. The mugilidae of Australia and adjacent seas. Aust. J. Mar. Freshw. Res. 1954 70-131 5 (1) TORAL, S. Tesis profesional Estudios de los Cichlidae (Pisces: Perciformes) de la laguna de Términos y sus afluentes. Fac. Ciencias, Univ. Nal. Autón. México 1971 32 p YÁÑEZ-ARANCIBIA, L. A. Informe Final de la 2da. Etapa del Programa de Uso de la Zona Costera de Michoacán y Guerrero. Relaciones tróficas de la fauna ictiológica del sistema lagunar costero de Guerrero y aspectos parciales de la dinámica de las poblaciones de los peces de importancia comercial. 230 p. 104 tablas, 71 figs. Convenio Comisión del Río Balsas, S. R. H. y Centro de Ciencias del Mar y Limnología, U. N. A. M., México. Contrato N9 OC-E-03-74. 1975a. 750 p. ---, Observaciones sobre Mugil curema Valencíennes en áreas naturales de crianza, México. Alimentación, crecimiento, madurez y relaciones ecoIógicas. An. Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México 1975b 92-124 3 (1) ---, Sobre los estudios de peces en las lagunas costeras: Nota científica. An. Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México 1975c 53-60 2 (1) YÁÑEZ-ARANCIBIA, L. A. y R. S. NUGENT Síntesis de antecedentes bióticos y abióticos de los ecosistemas estuarinos de Nayarit, México. Publ. Ciencias Biológicas. Univ. Autón. Nuevo León, México. Vol. in memoriam Dr. E. Caballero (en prensa). 1975 YÁÑEZ-ARANCIBIA, L. A., J. CURIEL-GÓMEZ y V. LEYTON DE YÁÑEZ Prospección biológica y ecológica de bagre marino Galeichthys caerulescens (Gunther) en el sistema lagunar costero de Guerrero, México. (Pisces: Ariidae). An Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México 1976 125-180 3 (1) ZAURANECH, J. B. Tesis Master of Science in Oceanography. The gerrid fishes of Eucinostomus in the Eastern Pacif. Univ. California, La Jolla. 1967 74 p.
|