|
ASPECTOS ECOLÓGICOS Y DISTRIBUCIÓN DE LOS MOLUSCOS EN LAS LAGUNAS COSTERAS DE GUERRERO, MÉXICO
JOSÉ STUARDO
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología y Proyecto
Multinacional en Ciencias del Mar, OEA.
MARÍA VILLARROEL
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y
Limnología.
Se discuten las peculiaridades de la
distribución de los moluscos que habitan el sistema lagunar del
estado
de Guerrero y su posible dependencia de los siguientes factores
ecológicos: salinidad, oxígeno, profundidad, turbidez,
sustrato y
sedimentos, y contenido de carbono orgánico (materia
orgánica) y
carbonatos en los últimos.
La fauna de moluscos vivos es escasa, representado por
sólo 8 especies: 3 "caracoles", las especies
Amnicola
sp., Melongena patula y Neritina
cassiculum de las cuales sólo la segunda es muy grande,
y 5
bivalvos, entre los cuales se encuentra la "ostra" u "ostión"
Crassostrea iridescens, que penetra
esporádicamente
sólo a la laguna de Potosí, el "mejillón de laguna"
Mytella strigata, la "navaja" Tagelus
longisinuatus muy abundante en los fondos blandos-arenosos y
las
"almejas" Iphigenia altior y Felaniella
sericata. Todos los bivalvos son de importancia
económica,
pero no necesariamente conviven juntos en una misma laguna.
Entre los factores estudiados, el más
importante y que regula la distribución de estos moluscos y otros
organismos lagunares es la salinidad.
El contenido de materia orgánica de los
sedimentos es variable con respecto al tipo de sedimento estudiado,
corroborándose su gran abundancia en los más finos. Sin
embargo,
no es totalmente aprovechado ya que aparentemente limita el desarrollo de
la
infauna debido a procesos redox ya conocidos.
The peculiarities of the distribution of the molluscs
inhabiting the coastal lagoons of the state of Guerrero and their
apparent
dependence on some ecological factors are discussed. The studied factors
are:
salinity, oxygen, depth, turbidity, substrate and sediments, and in the
latter,
organic carbon and carbonates.
The number of living molluscs is very low, being
represented by only 8 species: the 3 snai1s Amnicola
sp.
Melongena patula and Neritina
cassiculum
of which only the second one is large sized, and 5 bivalves, namely, the
oyster
("ostion") , Crassostrea iridescens, which
sporadically
enters the Potosi lagoon; the mussel, Mytella strigata
; the
razor clam, Tagelus longisinuatus, very abundant in
the
sandy bottoms; and the clams, Iphigenia altior and
Felaniella sericata. All the bivalves have economic
importance but do not necessarily live together in a same
lagoon.
Among the various environmental factors studied, the
most important in regulating the distribution of these molluscs and other
lagunar organisms is salinity.
The content of organic matter in the bottoms studied
varies according to sediment particle size, the higher organic content
being
found in the finer sized sediments. Moreover, as has been shown
elsewhere, the
redox processes in the latter appear to limit their utilization by the
infauna.
Este trabajo forma parte de un estudio prospectivo más amplio de los recursos biológicos y pesqueros de la zona costera de los estados de Michoacán y Guerrero, recientemente reportados por Stuardo et al. (1974) y complementa la información sobre moluscos de las lagunas costeras del estado de Guerrero allí presentada. La información general publicada con anterioridad sobre el área de estudio es mínima y corresponde a un estudio ecológico preliminar de las lagunas de Coyuca y Tres Palos casi dulceacuícolas, vecinas a Acapulco, realizado por Ramírez-Granados (1952) en el que no se mencionan moluscos. Los moluscos corresponden tradicionalmente a uno de los grupos mejor estudiados taxonómicamente y por lo menos la fauna de moluscos marinos del Pacífico mexicano ha sido incluida recientemente en una compilación de los moluscos tropicales con concha de la costa occidental de América realizada por Keen (1971) Existen, sin embargo, numerosas especies y aun taxa superiores incluidos en la fauna mexicana que son mal conocidos. Éste es el caso de los moluscos dulceacuícolas y de aguas salobres. Se debe a estas deficiencias que las identificaciones en muchos casos, no pueden pasar más allá de géneros. Los estudios sobre ecología y biología de los moluscos del Pacífico mexicano están comparativamente muy atrasados y fuera de investigaciones realizadas sobre algunos moluscos de importancia económica, sobre todo ostréidos (Sevilla, 1959; Stuardo y Martínez, 1976) y de alguna información inédita, no hay trabajo serios realizados al respecto. Frente a los objetivos de un estudio de este tipo es necesario referirse a dos aspectos conceptuales: uno es la definición de laguna costera; el otro es el origen y la distribución de la fauna denominada de estuarios o estuarina. El concepto de laguna costera se incluye generalmente en el de estuario aunque, aparentemente, hay la tendencia por diversos autores (por ejemplo Emery y Stevenson, 1957; Emery, 1967) de aceptar una separación aún imprecisa, pero importante entre los dos, como lo demuestra también el hecho de que se haya efectuado un simposio reciente sobre lagunas costeras en México (Ayala-Castañares y Phleger, editores, 1969). Pritchard (1967) ha propuesto y discutido una definición de estuario que incluye a las lagunas costeras: "Un estuario es un cuerpo de agua costero, semi-encerrado que tiene una conexión libre con el mar abierto y dentro del cual el agua de mar es mensurablemente diluida con agua dulce derivada de drenaje terrestre- (traducción, op. cit.). Esta definición es satisfactoria desde el punto de vista físico, ya que no hay separación abrupta entre los gradientes (de salinidad) de estuarios y lagunas, pero no dice nada de los aspectos biológicos. Caspers (1967) discutiendo las definiciones de estuario ha hecho notar las dificultades de llegar a una definición precisa sobre todo al tratar de caracterizar a un estuario desde el punto de vista faunístico. Sugiere que sería posible distinguir entre lagunas y estuarios analizando otro carácter hidrológico: la inestabilidad de la salinidad. Asi, cuando el aporte de agua dulce en un vaso separado desarrolla un cuerpo de agua salobre estable, podría hablarse de una laguna. Si la mezcla de agua dulce y marina no es estable sino que muestra cambios periódicos, el vaso puede ser considerando un estuario. Indudablemente esta separación no se aplica a los cuerpos de agua de llanuras costeras como los existentes en las costas de Guerrero, identificados corrientemente y fisiográficamente con el nombre de lagunas costeras. Mientras no existan definiciones más satisfactoria, si es que llegaran a haberlas, aceptamos el criterio de Emery y Stevenson (1957) de separar estuarios y lagunas desde un punto de vista fisíográfico aplicable a las lagunas de Guerrero. De acuerdo con estos autores: "Estuarios son cuerpos de agua bordeados y parcialmente separados del mar por masas terrestres -formadas originalmente por agentes no marinos. Generalmente son perpendiculares a la línea de costa y la mayoría de ellos ocupan las bocas anegadas de valles de ríos y, en consecuencia, son considerados corrientemente como evidencia de sumergencia" (traducción, op. cit.). "Lagunas son cuerpos de agua separados en la mayoría de los casos del océano por barras costeras o islas de origen marino y son generalmente paralelos a la línea de costa. Se consideran corrientemente resultado de emergencia, pero pueden formarse también como resultado de sumergencia de una planicie costera" (traducción, op. cit.). Respecto del origen y la distribución de la fauna de estuarios se acepta que se deriva parcialmente de la fauna dulceacuícola y parcialmente de la fauna marina (Hedgpeth, 1957). Day (1951, 1967) reconoce al respecto 5 componentes, que creemos conveniente seguir por razones de claridad: 1) Un componente dulceacuícola que comprende unas pocas especies derivadas de ríos que son tolerantes de sal. 2) Un componente marino estenohalino restringido a aguas de alta salinidad cerca de la boca del estuario. 3) Un componente marino eurihalino que se extiende desde el mar a través de todo el estuario. 4) Un componente estuarino que ha evolucionado de formas marinas, pero que se halla ahora restringido a estuarios y no es nunca encontrado en el mar. 5) Un componente migratorio que incluye una variedad de formas activas tales como aves, peces, camarones, langostinos y cefalópodos que entran y salen del estuario de acuerdo a migraciones de alimentación y desove. A estos componentes que enfatizan la importancia de la salinidad, este autor agregó posteriormente (1967) un nuevo componente al concluir que muchas de las especies denominadas estuarinas son en realidad especies de aguas tranquilas y que la habilidad para sobrevivir en aguas protegidas es tan importante en la formación de una población estuarina como su tolerancia a una salinidad reducida. Como lo ha hecho notar Phleger (1969) la necesidad de entender la naturaleza, dinámica e historia de las lagunas costeras modernas tiene muchas implicaciones científicas y económicas, en especial para México en donde por lo menos la tercera parte de su línea de costa corresponde a lagunas costeras con un potencial económico-pesquero considerable. La información discutida en este trabajo intenta contribuir al conocimiento faunístico y ecológico en especial de algunas lagunas de Guerrero. ÁREA DE ESTUDIOEl conjunto de lagunas costeras estudiadas cubre gran parte de los 490 km de costa del estado de Guerrero, en el Pacífico mexicano (Mapa 1) . Una descripción detallada de las lagunas es dada por Stuardo et al. (1974) y se resume en otra contribución reciente (Stuardo y Martínez, 1975). Los aspectos fisiográficos, geológicos y climatológicos generales del área de estudio han sido discutidos también recientemente por Lankford (1974). Existen alrededor de 11 lagunas de las cuales por su superficie, profundidad, características biológicas y pesqueras son importantes sólo 9 que enumeradas de sur a norte son las siguientes (Mapa 1): 1. Salinas de Apozahualco 2. Chautengo o Nexpa 3. San Marcos o Tecomate 4. Tres Palos 5. Coyuca 6. Mitla 7. Nuxco 8. Salinas del Cuajo 9. Potosí. Del total enumerado, la laguna de Tres Palos, una de las más grandes no fue estudiada originalmente. Por otra parte las lagunas Salinas de Apozahualco, Chautengo y Nuxco han continuado siendo estudiadas en mayor detalle. Las características hidrológicas de estas lagunas junto a información de otro tipo se incluyen en las tablas 2, 3, 4 y 5. 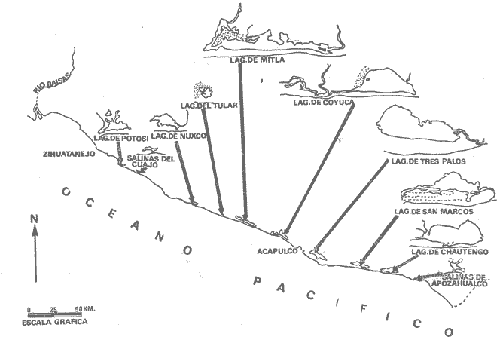 Mapa 1. Localización de las lagunas estudiadas en la costa del estado de Guerrero, México. MATERIALES Y MÉTODOSLas especies de moluscos provienen casi en su mayoría de un total de 50 muestras cuantitativas bentónicas obtenidas entre el 12 de septiembre de 1973 y el 11 de enero de 1974 en el área de estudio. En muchos casos las colectas se completaron con material colectado a mano o por buceo. Los datos hidrológícos provienen de muestras obtenidas hasta agosto de 1974 y los de sedimentos corresponden a muestras obtenidas conjuntamente con las anteriores en los meses de mayo y julio-agosto de 1974, respectivamente. Las fechas precisas se indican en las tablas respectivas. El material de moluscos obtenido en estas dos últimas oportunidades se halla aún en proceso de separación, pero no modifica sustancialmente los resultados aquí discutidos. Las muestras bentónicas fueron obtenidas con una draga Van Veen de 3.75 litros de capacidad y las de sedimentos con una draga del mismo tipo de 10 litros de capacidad. La separación de los organismos se efectuó mediante un set de tamices de 0.5, 2 y 10 mm respectivamente. Las muestras hidrológicas fueron colectadas con una botella Van Dorn de 3 litros de capacidad y procesada en el laboratorio de Oceanografía del Centro de Ciencias del Mar y Limnología. Para el estudio de las relaciones entre sedimentos y moluscos se eligieron las lagunas Salinas de Apozahualco, Chautengo y Nuxco, que se muestrearon en dos oportunidades: desde el 16 al 22 de mayo, en el periodo de secas y desde el 29 de julio al 3 de agosto en el periodo de lluvias. Se tomó un total de 51 muestras, distribuidas de la siguiente manera: 10 en Salinas de Apozahualco, 23 en Chautengo y 18 en Nuxco, las que correspondieron a 5, 12 y 10 estaciones respectivamente en el primer muestreo y 5, 8 y 11 respectivamente, en el segundo muestreo. De las muestras de 1 litro de sedimento obtenidas se hicieron dos tipos de análisis: granulométrico y químico. En el primer caso se siguieron las técnicas de análisis granulométrico descritas por Folk (1969) de acuerdo a los procesos descritos originalmente por Emery (1938) y Krumbein (1932). El análisis se realizó de la siguiente manera: 1. Se tomó una submuestra de 10 a 20 gramos (2 a 3 cucharaditas) y se lavó a través de un tamiz U. S. Standard 230, con abertura de malla de 0.0625 mm, con una solución peptizadora de carbonato y oxalato de sodio. 2. La fracción arenosa en las muestras, con 5% o más de arena retenida en este tamiz, fue analizada en el tubo de sedimentación, según el proceso descrito por Emery (1938). 3. La fracción limo-arcillosa, que pasó a través del mismo, fue analizada por el método de pipeteo descrito por Krumbein (1932). Con los resultados obtenidos se calcularon los porcentajes de arena-limo-arcilla, elaborándose mapas de distribución para cada laguna con el empleo del triángulo de Shepard. Basándonos en la distribución de las muestras en el triángulo de Shepard se definieron tres grupos de sedimentos que caracterizan a las lagunas estudiadas, a saber: Grupo I. Arenas; con un porcentaje de arena mayor a un 60%. Grupo II. Mezcla de arena-limo-arcilla; con un porcentaje de arena inferior a 60% y superior a 20%, y Grupo III. Limo-arcilla, arcilla-limo; con un porcentaje de arena inferior a un 20%. En el análisis químico de los sedimentos se determinó el contenido de carbonatos y carbono orgánico. Los porcentajes de carbonatos se obtuvieron mediante el método acidimétrico, que se resume a continuación. 1. A 0.5 gramos de muestra se agregan 10 ml de ácido sulfúrico 0.01 - 0.02 N. 2. Se filtra con bomba de vacío y filtro de fibra de vidrio. 3. Se calienta hasta el inicio de la ebullición; el exceso de ácido se titula con solución de hidróxido de sodio, usando como indicador fenolftaleina. El porcentaje de carbonato está dado por la siguiente fórmula:
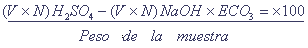 V= Volumen, N=Normalidad, E= Equivalentes. El porcentaje de carbono orgánico se determinó mediante el método de la vía húmeda de Schollenberger (1927) modificado posteriormente por Walkley y Black (1934). El análisis consistió básicamente en los siguientes pasos: 1. 0.5 gramos de muestra se colocan en un matraz Erlenmeyer de 250 ml. 2. Se agregan 2 m de solución de sulfato de plata (Ag2 SO4) - 3. Se añaden 5 m de una solución normal de K2 Cr2 O7 Y 10 mI de H2S04 concentrado. 4. Se agita durante 1 minuto y se deja reposar 30 minutos sin agitar. 5. Se afora a 100 ml con agua destilada y se agregan 5 ml de H3 PO4 y como indicador 5 a 6 gotas de sulfonato de bario difenilamina. 6. Se titula el exceso de dicromato (K2 Cr2 O7) con una solución 0.5 N de sulfato ferroso hasta que varíe de púrpura o azul a verde. El porcentaje de carbono orgánico está dado por la siguiente fórmula: 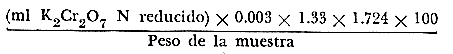 Tanto para la determinación de carbono orgánico como para la de carbonatos, las muestras fueron preparadas secando una alícuota a una temperatura menor de 80ºC; luego se separó el material grueso mayor de 1 mm y se pulverizó posteriormente en un mortero de porcelana. RESULTADOS Y DISCUSIÓN
CARACTERIZACIÓN ECOLÓGICA DE LAS LAGUNAS COSTERAS ESTUDIADASStuardo et al. (1974) han sugerido que los rasgos generales de las lagunas estudiadas permiten definir un sistema lagunar que puede caracterizarse de la manera siguiente: a) Todas las lagunas corresponden a cuerpos de agua someros, con volúmenes muy diferentes unos de otros. b) La superficie y el volumen individuales son variables a lo largo del año, siendo máximo en el periodo de lluvias y mínimo en el de secas, pudiendo reducirse las más someras en más de un 50%. c) La mayoría de las lagunas están separadas del mar por una barra, cuya abertura está supeditada a su altura, al aumento del volumen anual con la formación de una cabeza hidrostática que rompe a la barra por rebasamiento, o al fuerte impacto del oleaje y las corrientes marinas durante huracanes y otras catástrofes naturales. d) La entrada de agua de mar, como resultado de la abertura de las bocas, puede estar en el primer caso, restringida a un ciclo anual, o ser esporádica en el segundo, variable en el tiempo, pudiendo una laguna permanecer cerrada durante uno o más años. e) La duración de la abertura de las bocas es variable en el tiempo. f) Los aportes de agua dulce para cada laguna son diversos en su origen y variables en el tiempo. g) La salinidad interlagunar es prácticamente diferente en cada laguna y la intralagunar es variable a lo largo del año. Sin embargo todas las lagunas son salobres. h) Los fondos dominantes son lodosos, negruzcos, reductores. i) Existen escasos animales bentónicos de la macroinfauna y la macroepifauna vivos. j) La vegetación acuática es escasa aparte de los mangles y ocasionalmente de la vegetación semiacuática. k) La diversidad y productividad bentónica general es variable a lo largo del año. l) La diversidad y productividad planctónica general es también variable a lo largo del año. m) La producción pesquera actual y el potencial pesquero de cada laguna- son también variables. FAUNA DE MOLUSCOS LAGUNARESSe ha diferenciado un total de 23 especies de moluscos cuantitativamente importantes en los fondos lagunares, algunos de ellos identificables aún sólo a nivel de género debido a la falta de revisiones taxonómicas, sobre todo en grupos de micromoluscos y fauna dulceacuícola. No se han considerado conchas y restos de especies que penetran a las bocas arrastradas por las corrientes de mareas. La tabla 1, incluye el total de especies estudiadas. Puede constatarse que de las 23 especies sólo las 8 siguientes han sido encontradas vivas: Amnicola sp., Melongena patula, Neritina cassiculum, Crassostrea iridescens, Felaniella (Zemysia) sericata, Iphigenia altior, Tagelus (Tagelus) longisinuatus y Mytella strigata. (láminas 1 y 2) . De ellas, la última es la única que forma en algunas lagunas poblaciones epifaunísticas cuantitativamente importantes. T. (T.) longisinuatus, I. altior y F. (Z.) sericata todas integrantes de la infauna muestran poblaciones localizadas, de gran abundancia esporádica. De las especies restantes, N. cassiculum parece estar asociada principalmente al "mejillón de laguna" M. strigata y aparentemente abunda sólo donde hay poblaciones también vivas de la última. Una dependencia similar parece existir entre el "caracolillo" Amnicola sp. y el alga Cladophora sp., asociación que quizás incluya también algunas fanerógamas acuáticas. La mayoría, con excepción de Crassostrea iridescens, parecen ser especies adaptadas a condiciones salobres y/o aguas tranquilas. 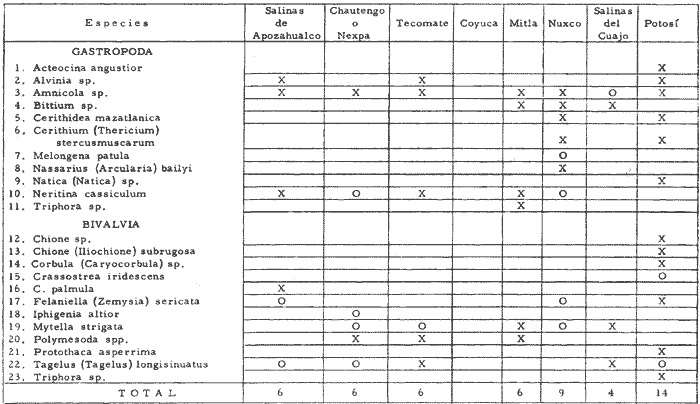 TABLA 1 DISTRIBUCION DE LOS MOLUSCOS EN LAS LAGUNAS ESTUDIADAS (O= vivos, X= muertos) Las 15 especies restantes están representadas sólo por valvas y restos de conchas. De ellas son importantes por su abundancia conchas del género Polymesoda, las que muestran una variación tal que es difícil identificarlas con algunas de las especies conocidas. No se han encontrado ejemplares vivos, pero a juzgar por la abundancia de conchas es factible que existan poblaciones en sectores de algunas de las lagunas que no hayan sido aún muestreados, o lo que es igualmente probable que las poblaciones ahora muertas se hayan desarrollado mientras existían condiciones de habitat que han desaparecido en la actualidad, o al menos en el momento del estudio. Éste es indudablemente el caso de los restos de especies marinas encontradas en gran abundancia en la laguna de Potosí: Acteocina angustior, Natica sp., Chione sp., Corbula sp., etcétera. Todas estas especies habitaron esta laguna mientras estuvo abierta al mar por 2 o 3 años consecutivos después del huracán Beulah. Lo mismo ocurre en la laguna de Mitla, donde los restos de conchas del "mejillón de laguna" son sumamente abundantes, pero en la que esta especie no puede vivir por las condiciones casi dulceacuícolas (oligohalínas) existentes en la actualidad. Es más, es indudable que el ciclo de variación anual existente en algunas lagunas, especialmente de la salinidad suele permitir el establecimiento temporal de poblaciones de origen marino, luego de abrirse las barras, mientras duran las condiciones marinas, o dulceacuícolas y salobres cuando la salinidad disminuye por efecto de las lluvias. El caso extremo se presenta hacia fines del periodo de secas (marzo-mayo) cuando grandes sectores de algunas lagunas quedan totalmente en seco o bajo una capa delgadísima de agua, a menudo con salinidades y temperaturas elevadas, determinando mortalidades masivas de organismos. Para entender mejor la distribución de los moluscos en las lagunas estudiadas es conveniente analizar algunos de los factores abióticos que la determinan. DEPENDENCIA DE LOS MOLUSCOS LAGUNARES DE LOS PARÁMETROS AMBIENTALESEl análisis de las características que presentan las distintas lagunas y los tipos de habítat predominantes permiten sugerir la importancia de algunos parámetros basándonos en la presencia o ausencia y en la distribución de los moluscos encontrados en las lagunas. Se considera que la salinidad y el sustrato son determinantes y en menor grado el oxígeno, la profundidad y la turbidez. SalinidadÉste es uno de los parámetros ambientales más y mejor estudiados en el mar y zonas estuarinas, y referencia a/o revisiones generales recientes sobre su significado ecológico e importancia en la distribución de los organismos estuarinos pueden encontrarse en los trabajos de Emery y Stevenson (1957), Pearse y Gunther (1957), Gunther (1961), Carriker (1967) y Kinne (1967 y 1971). Respecto de las lagunas de Guerrero, los valores de salinidad encontrados durante cuatro muestreos que corresponden aproximadamente a un ciclo anual, se resumen en la tabla 2. Los rangos máximos de salinidad conocidos para cada laguna se representan en la figura 8 y su variación en cada una de las estaciones muestreadás en las lagunas Salinas de Apozahualco, Chautengo y Nuxco, en las figuras 1, 2 y 3. Puede concluirse que en relación a la salinidad no existen dos lagunas que sean iguales. Se ha mencionado que la clasificación de los organismos estuarinos y lagunares sigue a su origen dulceacuícola o marino y desde este punto de vista, las lagunas de Guerrero pueden agruparse de acuerdo a dos características principales: a) Aquellas que mantienen rangos de salinidad bajísimos durante todo el año como son: Coyuca y Mitla, con características casi dulceacuícolas y con un aporte mínimo o sin ningún aporte de agua de mar (00. 11-04.98 % S), y b) Aquellas esencialmente salobres con valores de salinidad cercanos a los del agua de mar o superiores, y entrada de agua de mar anual, cíclica u ocasional. Aquí se incluyen todas las lagunas restantes (>05.00 % S). Los valores pueden disminuir considerablemente durante el periodo de lluvias. En las dos primeras lagunas no se han encontrado moluscos vivos en el fondo y aparentemente estas lagunas pueden caracterizarse mejor por la presencia de una vegetación sumergida o semiacuática, insectos e isópodos dulceacuícolas. El segundo grupo se caracteriza por un número de especies de moluscos de origen dulceacuícola o marino, que, sin embargo, no se identifican con ninguna laguna en particular sino con las condiciones existentes en el tiempo, en cada una de ellas. De este modo su presencia puede ser desde estacional a más o menos permanente, pero en general la variación de los parámetros hidrológicos determina la existencia de organismos sólo dentro de límites más o menos determinados, conclusión similar a la que ha llevado a postular efectos concretos de la salinidad en la distribución de los invertebrados en habitats con gradientes de salinidad pronunciados tales como estuarios, esteros y lagunas y en aguas costeras, lo que ha resultado en las numerosas clasificaciones propuestas al respecto. 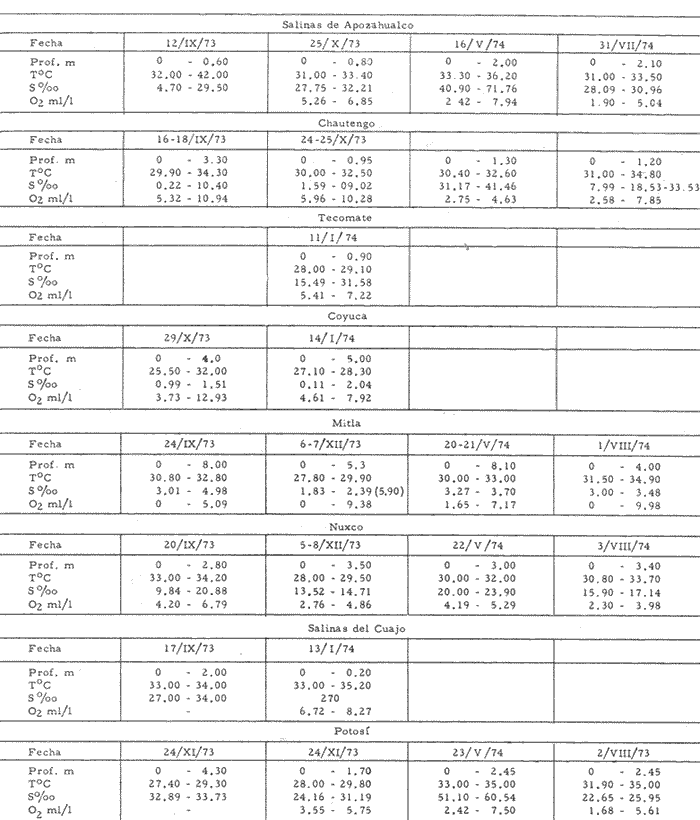 TABLA 2 MARIACIONES MÁXIMAS Y MINIMAS EM LAS LAGUNAS DE GUERRERO La figura 8 basada en una clasificación de "zonas estuarinas" propuesta recientemente por Kinne (1971), representa la distribución de los moluscos vivos encontrados en las distintas lagunas y los límites aproximados de salinidad a que se los encuentra. Esta clasificación se ha ampliado para incluir otras dos zonas salinas que caracterizan también a las lagunas: hipersalina y ultrahalina. Por razones de espacio se han incluido sólo los nombres genéricos de las especies. De las especies enumeradas, las estuarinas que forman parte de la epifauna, Mytella, Neritina y Amnicola son las únicas cuya distribución parece estar determinada por límites de salinidad más o menos precisos, aunque en el caso de Amnicola se ha observado que su presencia depende de algunas algas, de las que probablemente se alimenta. La distribución de las especies restantes sigue una combinación de condiciones más salinas y la presencia de arena. En general, estos resultados y los siguientes confirman la aserción que ecológicamente estuarios y lagunas representan zonas de competencia reducidas en las que los factores físicos más que los biológicos son los que determinan la dinámica de las poblaciones. OxígenoÉste es otro de los factores mejor estudiados en el mar y en general se piensa que no es un factor limitante aunque se han reportado casos de mortalidades masivas causadas por deficiencias de oxígeno (Brengersma-Sanders, 1957; Tulkki, 1965). Los límites más bajos de oxígeno disuelto adecuados para la, respiración de la mayoría de los organismos marinos parecen ser del orden de 1.0 a 2.0 ml/l, pero en general los límites de supervivencia para diversas especies y habitats son aún pobremente conocidas (Vernberg, 1972). La tabla 2 resume los valores máximos y mínimos de oxígeno encontrados en todas las lagunas estudiadas y las figuras 1 a 3 detallan estos valores por estación para las lagunas Salinas de Apozahualco, Chautengo y Nuxco. Se observa que en general los valores de oxígeno son relativamente elevados y corresponden a los valores máximos de saturación del agua de mar y de agua dulce a temperaturas similares (alrededor de 30ºC) y en muchos casos son aún superiores. Aparentemente hay una diferencia entre los valores obtenidos durante el periodo de secas y el de lluvias ya que durante los meses de mayo y junio se nota una tendencia a bajar (tabla 2, Figs. 1, 2 y 3). A pesar de la presencia de fondos reductores malolientes, en gran parte de los fondos lagunares, se han encontrado condiciones anaeróbicas sólo en algunas estaciones de la laguna de Mitla de características casi dulceacuícolas, en lugares donde las profundidades son mayores a 4.0 m. La variación de los valores de un punto a otro en esta y otras lagunas, es bastante grande, pero en general dentro de límites (2.30 a 12.93 ml/l) que indudablemente no alteran el metabolismo de los organismos que las habitan, ni afecta su tolerancia, reproducción o distribución. SustratoLa distribución de los organismos bentónicos sésiles o parcialmente sésiles está determinada básicamente por características de dureza del sustrato, lo que llevó a Petersen (1913, 1918) a proponer la separación de aquellos que viven dentro del sustrato o sobre él introduciendo los términos de epifauna e infauna. Fuera de esta separación global la caracterización de la distribución de organismos bentónicos en sustratos blandos o semiduros ha seguido dos tendencias interpretativas: la que apoya la dependencia primordial de distintos grupos de organismos al sustrato, y los que la niegan, estableciendo una dependencia sólo de tipo secundario. Como ocurre a menudo con disyuntivas de este tipo es indudable que uno u otro de estos criterios es válido para planteamientos y situaciones diversas. Por ejemplo, si una clasificación de los sedimentos distingue grupos tan amplios como lodo, lodo-arena, arena fina y grava de conchas (Craig y Jones, 1966) la distribución de diversos organismos presentará claramente una dependencia a estos tipos de sustrato. Por el contrario si la clasificación distingue entre distintos tipos de lodo, como ocurre en el estudio granulométrico de los sedimentos, la dependencia de organismos da sedimentos muy finos como limo y arcilla es oscura y es necesario considerar otras variables tales como sus características químicas (Nelson, 1962), la presencia de materia orgánica (Bader, 1954) o adaptaciones morfológicas funcionales de los organismos estudiados. 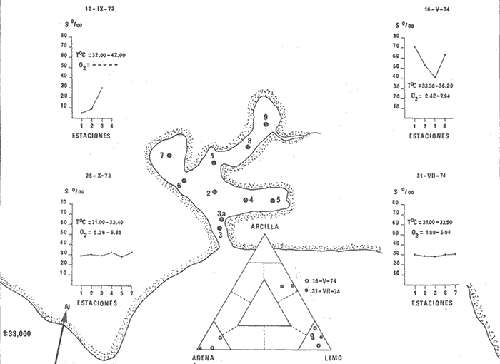 Fig. 1. Ubicacion de las estaciones (centro), distribución de las muestras de sedimentos en el triágulo de Shepard (abajo) y variación de los parámetros hidrológicos en laguna Salinas de Apozahualco. Establecer relaciones es aún más difícil cuando el número de organismos es reducido y las variaciones del medio físico son grandes. Carriker (1967) ha discutido estos problemas in extenso, deduciendo que algunas de las conclusiones más estimulantes obtenidas en estudios en aguas costeras, como la de la dependencia de organismos bentónicos de la infauna a determinado tamaño de partícula, deben ser detalladamente estudiadas. Éste es el caso, por ejemplo, de la dependencia sugerida por Bader (1954) y Sanders (1956, 1958) de los organismos detritívoros a fondo arcillosos por la capacidad de estas partículas de "fijar" materia orgánica. De las 8 especies de moluscos vivos encontrados en las distintas lagunas sólo los gasterópodos Amnicola sp. y Neritina cassiculum y el bivalvo Crassostrea corteziensis no presentan relaciones directas con los fondos lagunares. Los dos gasterópodos son aparentemente herbívoros, pero mientras Amnicola sp. depende en su distribución como ya se ha mencionado- de algunas algas, Neritina cassiculum se presenta casi siempre asociada a poblaciones del "mejillón de laguna" Mytella strigata. La "ostra" C. corteziensis se fija a raíces de mangle y fondos duros pero, únicamente, mientras existan condiciones marinas. 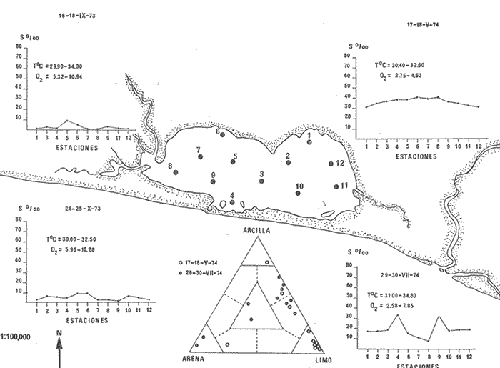 Fig. 2. Ubicació n de las estaciones (centro), distribución de las muestras de sedimentos en el triángulo de Shepard (abajo) y variación de los párametros hidrológicos en la laguna de Chautengo. Los moluscos restantes en mayor o menor grado siguen una distribución acorde con distintos tipos de sedimentos, relación que se ha estudiado en algún detalle en sólo tres lagunas de Guerrero: Salinas de Apozahualco, Chautengo y Nuxco. En las tablas 3, 4 y 5 se da una lista de las muestras (estaciones) estudiadas en cada una de estas lagunas, indicando los porcentajes de arena, limo y arcilla encontrados, junto a otros parámetros medidos. Los porcentajes obtenidos se han representado en cada caso por un triángulo de Shepard insertado en las figuras 1 a 3. Su distribución de acuerdo a los tres tipos definidos en Materiales y Métodos y denominados Grupo I (arena), Grupo II (arena-limo-arcilla) y Grupo III (limoarcilla; arcilla-limo), se incluyen en las figuras 5, 6 y 7. La clasificación en sólo tres tipos se ha hecho considerando que los únicos moluscos que en su distribución siguen a algunos de estos tipos son tres especies de la infauna y una de la epifauna. Las tres especies de la infauna son las "almejas" Felaniella sericata, Iphigenia altior y Tagelus longisinuatus. La especie que forma parte de la epifauna es el "mejillón de laguna" Mytella strigata. Las dos primeras son filtradoras de partículas en suspensión. Tagelus longisinuatus y las especies del género Tagelus, como las de Tellinacea, en general, eran tradicionalmente consideradas detritívoras (se alimentan de depósitos), pero Pohlo (1973) y Villarroel y Stuardo (en prensa) han demostrado que especies del género Tagelus, incluyendo a la aquí mencionada, se alimentan de partículas en suspensión. En consecuencia, su dependencia del sustrato, si la hay, es de otro tipo, aunque no se puede dejar de considerar que la gran cantidad de sedimentos en suspensión que existe en el agua de estas lagunas pudiera indirectamente favorecer su alimentación. 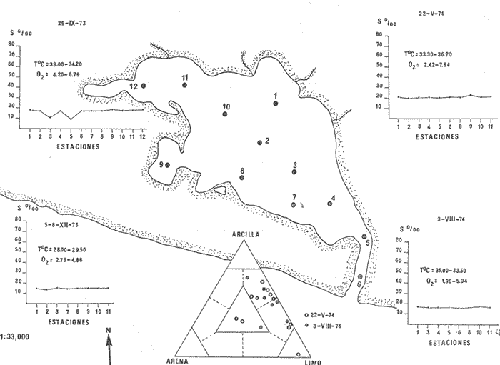 Fig. 3. Ubicación de las estaciones (centro), distribución de las muestras de sedimentos en el triángulo de Shepard (abajo) y variación de los parámetros hidrológicos en la laguna de Nuxco. Las figuras 5, 6 y 7 muestran la distribución de estas especies y la de los tres tipos de sedimentos. Se observa que tanto Felaniella sericata como Iphigenia altior se encuentran viviendo sólo en arena y en las cercanías de las bocas. Poblaciones de la primera especie variando entre 60 y 120 ejemplares por m2 (Stuardo et al., 1974) se han encontrado sólo en Salinas de Apozahualco y en Nuxco, cerca de las barras, y en la parte interior de ellas. Poblaciones de Iphigenia altior existen sólo en fondos de arena gruesa del interior de la barra de Chautengo en el sector noroeste. En otras lagunas no se han encontrado poblaciones vivas de esta especie, ni conchas (valvas) o restos de ellas. 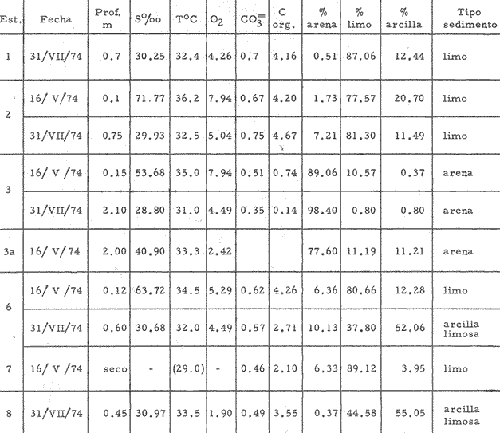 TABLA 3 PORCENTJES DE ARENA, LIMO, ARCILLA Y CONTENIDO DE CARBONATOS Y CORBONO ORGANICO EN LOS SEDIMIENTOS DE LA LAGUNA DE LAS SALINAS DE APOZAHUALCO, PARA LOS PERIODOS DIFERENTES (MAYO, 1974 - JULIO, 1974) Tagelus longisinuatus también forma poblaciones importantes (hasta de aproximadamente 120 ejemplares por m2) especialmente en las proximidades de las barras y de preferencia en fondo arenosos, pero como se demuestra en la figura 5, en la laguna Salinas de Apozahualco, invade también fondos con sedimentos del grupo III. No se encontraron ejemplares vivos de esta especie en sedimentos finos de este grupo en ninguna otra laguna, pero la abundancia de conchas muertas en todos los fondos lagunares estudiados sugiere presencia -si no abundancia-, por lo menos esporádica de esta especie en fondos blandos de sedimentos arcillo-limosos o limo-arcillosos. 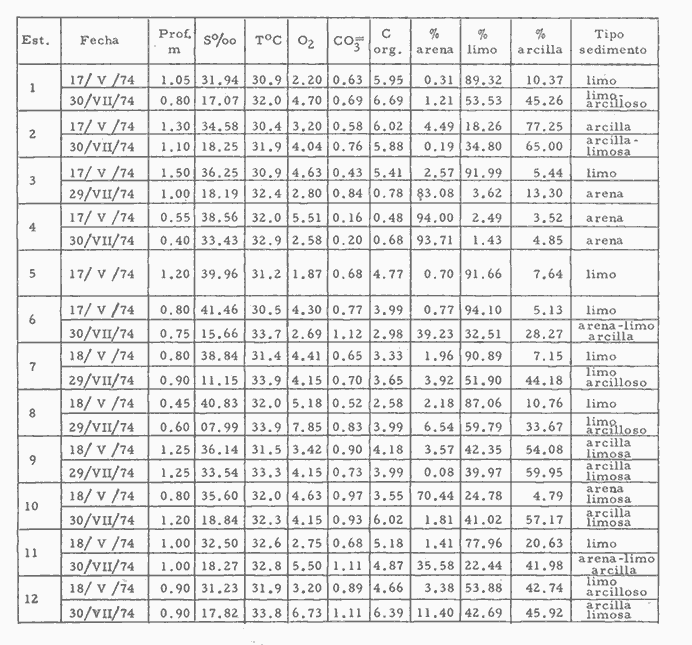 TABLA 4 PORCENTAJES DE ARENA, ARCILLA, Y CONTENIDO DE CARBONATOS Y CORBONO ORGANICOEN LOS SEDIMIENTOS DE LA LAGUNA DE CHAUTENGO PARA LOS PERIODOS DIFERENTES (MAYO, 1974 - JULIO, 1974) La epifauna de fondos blandos está representada solamente por poblaciones del "mejillón de laguna" Mytella strigata que habita sólo en las lagunas de Chautengo y Nuxco, pero no en Salinas de Apozahualco. En la distribución de esta especie filtradora de partículas en suspensión interviene como factor limitante principal la salinidad (Fig. 8) y su adaptación a los fondos blandos ha determinado la formación de una comunidad importante que incluye como integrantes secundarios a Neritina cassiculum, Balanus amphitrite, juveniles y especies pequeñas de "jaibas" (Callinectes, Hexapanopeus) y poliquetos indeterminados. Naturalmente este mejillón crece también sobre fondos duros tales como rocas, palos y piedras, pero no en arena. 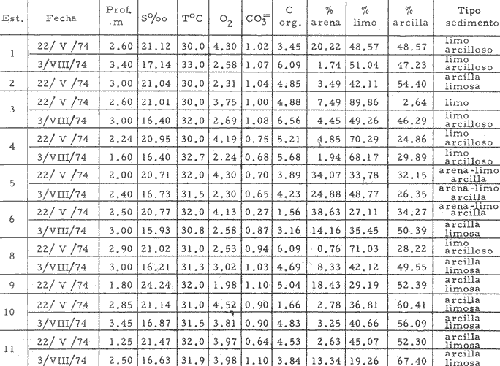 TABLA 5 PORCENTAJES DE ARENA, LIMO Y CONTENIDO DE CARBONATOS L CARBONO ORGÁNICO EN LOS SEDIMENTOS DE LA LAGUNA DE NUXCO PARA DOS PERIODOS DIFERENTES (MAYO, 1974 - AGOSTO,1974) El "caracol" Melongena patula es la única otra especie encontrada sólo en los fondos arenosos próximos a la barra de la laguna de Nuxco. TurbidezLa turbidez como una propiedad óptica de partículas en suspensión en el agua es notoria en aguas costeras y, particularmente, en estuarios y lagunas costeras en donde varía ampliamente a, lo largo del año alcanzando un máximo durante las inundaciones de la estación lluviosa (Emery y Stevenson, 1957). Sin embargo, la información de sus efectos sobre peces y organismos bentónicos es aún muy controvertible (Kinne, 1971). La mayor parte de los estudios se han basado en especies importantes para las pesqueríasde aguas dulces como son los salmónidos y de organismos sésiles como la "ostra". Para cualquier observador es indudable que la abundancia de algunos organismos faunísticos en estuarios y lagunas costeras, aun en aquellos donde la turbidez es muy notoria, no parece influenciar grandemente ni su abundancia ni su distribución. Por el contrario su efecto sobre la reducción de la zona fotosintética es conocido. 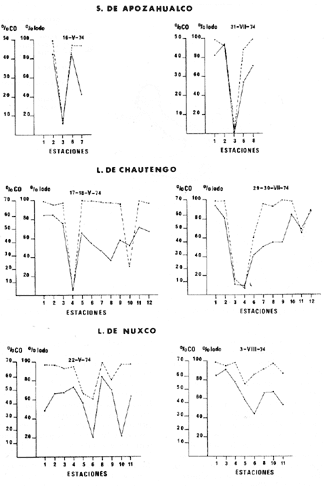 Fig. 4. Relación de % de carbono orgánico y % de lodo para dos períodos diferentes en las lagunas Salinas de Apozahualco, Chautengo y Nuxco. 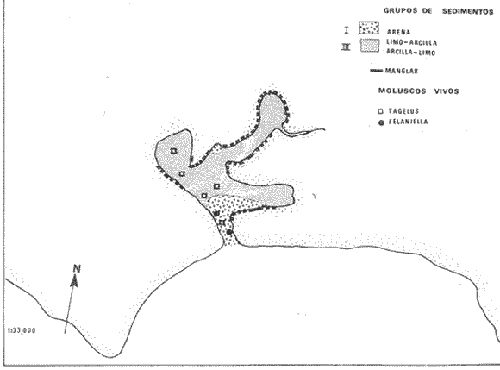 Fig. 5. Relación entre tipos de sedimentos y la distribución observada de los moluscos bentónicos en la laguna Salinas de Apozahualco. En las tres lagunas estudiadas no se ha medido la turbidez sino a través de observaciones indirectas mediante el empleo del disco de Secchi, cuya visibilidad a menudo es ya nula aproximadamente a los 15 cm de profundidad. En el caso de los moluscos filtradores, aparte de algunas adaptaciones en la amplitud de la cavidad paleal y el tamaño de las branquias y palpos que facilitan la filtración de partículas no se conocen efectos desfavorables, salvo quizás, por enterramiento en el caso de velocidades extremas de sedimentación. En el hecho se ha sugerido que la materia orgánica en suspensión puede favorecer no sólo la actividad de alimentación de los adultos sino también de las larvas (Loosanoff y Davies, 1963). En consecuencia, el papel que juega la turbidez en la distribución de los moluscos lagunares no es determinante aunque será necesario estudiar con mayor precisión sus efectos sobre todo en relación a la vegetación sumergida. En términos generales la turbidez de la laguna de Nuxco es mínima comparada a la de las otras dos lagunas, especialmente Chautengo, relación que está demostrada en la primera de estas dos lagunas por la presencia de extensas poblaciones de la fanerógama marina Ruppia maritima. En Chautengo la densidad y abundancia del "mejillón de laguna" Mytella strigata en gran parte de los fondos es prácticamente la misma por unidad de superficie, independientemente del grado de turbidez. 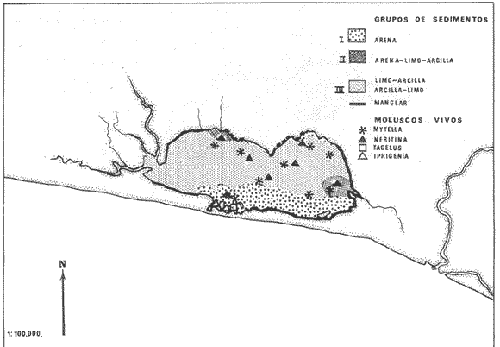 Fig. 6. Relación entre tipos de sedimentos y la distribución observada de los moluscos bentónicos en la laguna de Chautengo. ProfundidadTratándose de cuerpos de agua relativamente someros, la profundidad no juega ningún papel de importancia en la distribución de los moluscos estudiados (no así en el caso de la vegetación sumergida; ver Turbidez) . Probablemente su importancia es indirecta en algunas lagunas como en la de Mitla, donde la presencia de fondos anóxicos podría limitar la distribución de algunas especies en profundidad, si las hubiere. En la actualidad esta laguna no presenta poblaciones vivas de moluscos bentónicos, ni aun en la epifauna, debido a su baja salinidad y quizás a la presencia de fondos muy reductores. Carbono orgánico (materia orgánica)La importancia de la materia orgánica en la distribución de los organismos bentónicos, especialmente moluscos en aguas costeras ha sido enfatizada por diversos autores, especialmente por Bader (1954, 1962). La materia orgánica sirve como fuente de energía tanto a bacterias heterotróficas, como a organismos bentónicos de mayor tamaño. En fondos ricos en detrito orgánico, la microfauna, macrofauna, depredadores y omnívoros abundan a sus expensas. En condiciones estuarinas y lagunares el contenido orgánico de los sedimentos es elevado, no así la diversidad de los organismos que los habitan y en consecuencia, su importancia comparada a la variación de algunas características del agua, tales como salinidad y temperatura aparece secundaria. 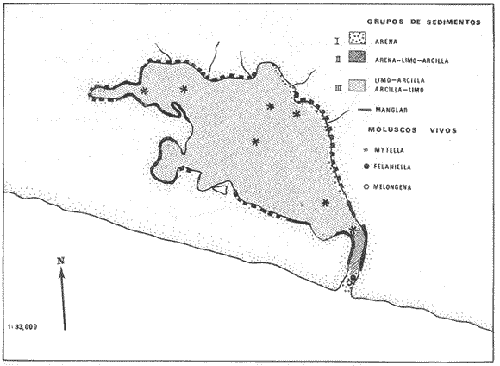 Fig. 7. Relación entre tipos de sedimentos y la distribución observada de los moluscos bentónicos en la laguna de Nuxco. La comparación de los valores obtenidos con los publicados en diversos trabajos es difícil, ya que no existe uniformidad de criterios para medirla y diversos autores utilizan coeficientes diferentes de conversión de carbono orgánico a materia orgánica. Los porcentajes de carbono orgánico indicados para estuarios, lagunas y aguas costeras varían entre 0.6 a 1 y 7 a 8% (Trask, 1939; Bader, 1954; Oppenheimer y Ward, 1963). En este trabajo se analizó el contenido de materia orgánica en las lagunas Salinas de Apozahualco, Chautengo y Nuxco para estimar una abundancia sugerida por el color negruzco de gran parte de los sedimentos de ellas. Los resultados que se presentan en las tablas 3, 4 y 5, en las tres lagunas variaron entre 0. 14 y 6.69%. Salinas de Apozahualco, al igual que en el caso de los porcentajes de carbonatos, fue la que presentó los valores más bajos, fluctuando entre 0.14 y 4.67%. Esto puede explicarse por sus grandes diferencias ecológicas respecto de las otras lagunas. Su vaso es muy pequeño, los aportes de agua dulce por escurrimiento son mucho menores y sus ciclos de variación son extremos, yendo del anegamiento a un desecamiento casi total. En las otras lagunas con un volumen de agua relativamente constante los valores variaron entre 0.48 y 6.69 %. 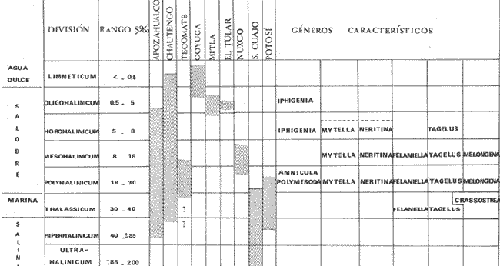 Fig.8. Relaciones entre moluscos vivos y rangos de salinidad en las lagunas de Guerrero. Dentro de cada laguna, los porcentajes se distribuyeron en relación al tipo de sedimento. Los valores más bajos se midieron en sedimentos gruesos en las cercanías de las bocas; en cambio, como era de esperar los valores más altos se obtuvieron en aquellos fondos con sedimentos finos. Esta distribución está de acuerdo con la comúnmente descrita para otros ambientes lagunares (Krumbein, 1939). No existe relación entre la distribución de los moluscos y los porcentajes de materia orgánica obtenidos, lo que se debe a aquellas especies encontradas son filtradoras de partículas pero esto a su vez parece ser consecuencia de una causa mucho más importante. Bader (1954) ha encontrado una relación entre depósitos de grano extremadamente fino y una fauna de diversidad muy baja y baja diversidad taxonómica. Esta relación puede ser atribuida principalmente a una circulación intersticial pobre que resulta en la acumulación de sedimento fino y dentro de él de productos de descomposición tóxica y/o deplesión del oxígeno disponible (Purdy, 1964). Tal combinación limita la distribución de los pelecípodos ínfaunísticos- y probablemente de otros invertebrados y de la fauna intersticial- en sedimentos que contienen más de un 3% de materia orgánica. Según Bader (op cit.) el rango óptimo de materia orgánica estaría entre 1.18 y 2.80%. En consecuencia, valores más altos o muy altos pueden indicar indirectamente pobreza o ausencia de organismos bentónicos aeróbicos, como es el caso de las lagunas estudiadas. La relación directa entre sedimentos de grano fino y contenidos más altos de carbono orgánico y viceversa, pobreza de carbono orgánico en sedimentos gruesos es una generalización conocida ya desde hace años (Trask, 1939; Stewart, 1958; Purdy, 1964, etcétera). Esta relación es claramente observable en las tres lagunas estudiadas (Fig. 4). CarbonatosLos porcentajes de carbonatos medidos tanto en la época de lluvias como en la de secas fueron, en la mayoría de las muestras, bajos (tablas 3, 4 y 5) y se encuentran dentro del rango de 0 a 2 % dado por Trask (1932: 99), valores que corresponden al contenido medio en sedimentos de ambientes salobres. Se constató que los porcentajes fueron ligeramente mayores en la época de lluvias. En general, los porcentajes obtenidos variaron entre 0.20 y 1.12 % en las tres lagunas estudiadas. La variación de los valores individuales por laguna fue la siguiente: 0.35 a 0.75 en Salinas de Apozahualco 0. 16 a 1. 12 en Chautengo, y 0.27 a 1.10 en Nuxco. Los porcentajes más bajos en Salinas de Apozahualco podrían explicarse por el mayor aporte de material calcáreo hecho en las lagunas de Chautengo y Nuxco por las densas poblaciones del "mejillón" Mytella strigata. Sin embargo, al nivel de estos estudios no es posible precisar las diversas causas que pueden determinar estas variaciones. Por otra parte la información sobre la distribución de carbonatos en sedimentos parece a veces conflictiva. Por ejemplo Warme (1969: 147) al estudiar la laguna Mugu en la costa sur de California, encontró porcentajes altos de carbonato en los sedimentos arenosos. Sin embargo, atribuye también estos valores a la presencia de fragmentos de conchas. No encontró carbonatos en los lodos. De acuerdo a nuestros resultados los porcentajes menores, en cualquiera de las tres lagunas, se encuentran junto a las barras, donde los sedimentos son de arena gruesa y grava sin conchas; en cambio los porcentajes mayores corresponden a las áreas donde los sedimentos son más finos, salvo en las estaciones 6, 11 y 12 de la laguna de Chautengo, en que son altos, aunque presentan sedimentos del tipo arena-limoarcilla. Valores altos en sedimentos gruesos de origen fluvial han sido descritos también por Gutiérrez-Estrada (1971) en dos estaciones de su estudio sedimentológico del delta del Río Balsas. Estos valores no han sido aún explicados. CARACTERIZACIÓN DE LA FAUNA DE MOLUSCOS DE LAS LAGUNAS ESTUDIADASSe ha concluido que la fauna de moluscos existentes en las lagunas de Guerrero no hace excepción a las generalizaciones obtenidas en otras áreas estuarinas y que las especies pueden agruparse siguiendo algunas de las clasificaciones de zonas propuestas por rangos de salinidad (Fig. 8; adaptado de Kinne, 1971). Del mismo modo, dependiendo de sus relaciones con los valores de salinidad encontrados pueden tratar de agruparse según su origen siguiendo la clasificación de Day (1951, 1967) ya mencionada. De acuerdo con ella, no hay especies de moluscos que caractericen a un componente dulceacuícola a no ser que la especie de Amnicola encontrada resulte tener una distribución amplia en ríos y aguas dulces vecinas. Es sugestivo, sin embargo, que hasta ahora se hayan encontrado ejemplares vivos de esta especie sólo en la laguna Salinas de Apozahualco en salinidades relativamente elevadas (27 a 34 %), o muy bajas como en la de Mitla. Puede reconocerse un componente marino estenohalino representado por las especies Crassostrea corteziesis y C. palmula las que penetran al menos a dos lagunas (Salinas de Apozahualco y Potosí) en estado larvario cuando se abren las barras. La permanencia de las poblaciones que así se forman dependerá de la duración de condiciones marinas, aunque en el caso de la laguna de Potosí se han encontrado ejemplares juveniles vivos hasta en salinidades de 60.54 %. Es difícil precisar la posible existencia de un componente marino eurihalino. Las especies Felaniella sericata e Iphigenia altior son descritas de aguas costeras y, aparentemente como ya se ha discutido, su distribución más que estar sujeta a rangos de salinidad lagunares parece depender de la naturaleza del sustrato. La única especie verdaderamente marino-eurihalina podría ser Tagelus longisinuatus, si efectivamente habita también en aguas costeras como se registra en la literatura malacológica. Recientemente, Villarroel y Stuardo (1976) han concluido que parece tratarse de una especie que habita estuarios, esteros y lagunas costeras, por lo que sería necesario incluirla en el componente estuarino o salobre. En el hecho el único componente definido en el área de estudio es el componente estuarino representado por las especies Mytella strigata, Neritina cassiculum y Polymesodasp. Todas ellas son incapaces de vivir en aguas costeras marinas, aunque es posible que como lo sugiere Day (1967) la presencia de aguas tranquilas sea también un factor importante en su distribución. Melongena patula puede considerarse también como un organismo típicamente estuarino. No se conocen especies que formen un componente migratorio entre los moluscos estudiados. Si bien la caracterización de estos componentes lagunares-estuarinos es difícil de precisar, si se considera sólo a los moluscos, no lo es al incluir a otros organismos encontrados en la prospección de las lagunas. Relaciones faunísticas generales de este tipo, sobre todo de los invertebrados son discutidas en otro trabajo (Stuardo y Martínez, 1975). Un aspecto más, de gran importancia en estudios de la fauna de estuarios y algunas costeras, se refiere a los aspectos taxonómicos de ella. Sobre todo en trabajos de tipo ecológico en áreas pobremente conocidas, el investigador debe ser extremadamente cuidadoso ya que como Muus (1963, 1967) lo ha hecho notar, la presunción de que organismos abundantes deben ser especies bien definidas, no es siempre verdadera. Por una combinación de estudios morfilógicos y ecológicos este autor ha demostrado que aún en regiones supuestamente muy bien conocidas como es la nordeuropea, nombres de especies familiares encierran a veces a más de una especie, o a grupos subespecíficos producto de la variación como resultado natural de procesos de adaptación y aislamiento. AgradecimientosLos autores expresan sus agradecimientos a Robert Lankford y a Raymundo Lecuanda por las facilidades otorgadas en el laboratorio de Geología Marina del Centro de Ciencias del Mar y Limnología, UNAM, para el procesamiento granulométrico de las muestras y muy valiosas sugerencias; a Lisandro Castellanos y a Rubén Rodríguez por su consejo en la elección de los métodos empleados en las determinaciones de materia orgánica y carbonatos y por poner a nuestra disposición los datos de salinidad y oxígeno obtenidos en las lagunas estudiadas; a Daniel Méndez y Luis González por su ayuda en la obtención de las muestras y procesamiento parcial del material colectado y a Susana Arellano por su eficiente ayuda mecanográfica. Agradecemos también a Alfredo Laguarda Figueras, Director del Centro de Ciencias del Mar y Limnología y a las autoridades de la Comisión del Río Balsas, Secretaría de Recursos Hidráulicos quienes otorgaron los medios para realizar este trabajo. LITERATURALagunas Costeras. Un Simposio. Mem. Simp. Intern. Lagunas Costeras. AYALA-CASTAÑARES, A. y F. B. PHLEGER UNAM-UNESCO 1969 686 p. Nov. 28-30, 1967. BADER, R. G. The role of organic matter in determining the distribution of pelecypods in marine sediments. J. mar. Res. 1954 32-47 13 ---, Some experimental studies with organic compounds and minerals. In Symposium on the Envionmental Chemistry of Marine Sediments. Marshall, N. Narragensett Marina Lab. Occ. Publ. 1962 42-57 1 BRONGERSMA-SANDERS, M. Mass mortality in the sea. Treatise on Marine Ecology and Paleoecology. Hedgpeth, J. W. Geol. Soc. America, Mem. 1957 941-1010 67, 1 CARRIKER, M. R. Ecology of estuarine benthic invertebrates: a perspective. Estuaries. Publ. Amer. Ass. Adv. Sci. Lauff, G. H. 1967 442-487 83 CASPERS, H. Estuaries: Analysis of definitions and biological considerations. Estuaries. Publ. Amer. Ass. Adv. Sci. Lauff, G. H. 1967 442-487 83 CRAIG, G. Y. y N. S. JONES Marine benthos, substrate and paleoecology. Palaeontology 1966 30-38 9 (1) DAY, J. H. The ecology of South African estuaries. Part 1. General considerations. Trans. R. Soc. S. Afr 1951 53-91 33 ---, The biology of Knysna Estuary, South Africa. Estuaries. Publ. Amer. Ass. Adv. Sci. Lauff, G. H. 1967 397-407 83 EMERY, K. O. Rapid method of mechanical analysis of sands J. sedim. Petrol. 1938 105-111 8 (3) ---, Estuaries and Lagoons, in Relation to Continental Shelves. Estuaries. Publ. Amer. Ass. Adv. Sci. Lauff. G. H. 1967 911 83 EMERY, K. O. y E. STEVENSON Estuaries and Lagoons. I Physical and chemical characteristics. III Sedimentation in estuaries, tidal flats and marshes Treatise on Marine Ecology and Paleoecology. Hedgpeth, J. W. Geol. Soc. America, Mem. 1957 673-693 y 729-749 67, 1 FOLK, R. L. Traducción de Carmen SchIaepfer y Rebeca M. de Schmitter. Petrología de las rocas sedimentarias. Inst. Geol. UNAM 1969 405 p. GUNTER, G. Some relations of estuarine organisms to salinity. Limnol. Oceanogr. 1961 182-190 6 (2) GUTIÉRREZ-ESTRADA, M. Fisiografía y sedimentología del delta del Río Balsas. Michoacán, México. Boln. Inst. Geol. Méx 1971 98 p. 93 HEDGPETH, J. W. Estuaries and Lagoons, II Biological Aspects. Treatise on Marine Ecology and Paleoecology. Hedgpeth, J. W. Geol. Soc. America, Mem. 1957 673-729 67, 1 KEEN, M. Sea shells of tropical West America. Stanford Univ. Press. 1971 1064 p. KINNE, O. Physiology of estuarine organisms with special reference to salinity and temperature: General aspects. Lauff, G. H. Estuaries. Publ. Amer. Ass. Adv. Sci. 1967 525-540 83 ---, Marine Ecology Salinity. 4.3 Animals. 4.31 Invertebrates. Kinne, O. 1971 821-995 1 (2) KRUMBEIN, W. C. The mechanical analysis of finegrained sediments. J. sedim. Petrol. 1932 140-149 2 (3) ---, Recent Maríne Sediments, a Symposium. Tidal Lagoon Sediments on the Mississippi Delta. Trask, F. D. American Assoc. Petrol. Geol. Tulsa, Oklahoma 1939 178-194 LANKFORD, R., . , Informe del estudio de reconocimiento de la región costera de los estados de Michoacán y Guerrero. Subprograma de Geología. Descripción general de la zona costera de Guerrero y Míchoacán. Informe Centro de Ciencias del Mar y Limnología. UNAM 1974 42 p. LOOSANOFF, V. L. y H. C. DAVIS Rearing of bivalve mollusks. Adv. mar. Biol. 1963 1-136 1 Muus, B. J. Some Danish Hydrobiidae with the description of a new species, Hydrobia neglecta. Proc. malac. Soc. Lond. 1963 131-138 35 ---, Some problems facing the ecologist concerning races and subspecies of brackish-water animals. Lauff, G. H. Estuaries. Publ. Amer. Ass. Adv. Sci. 1967 558-563 83 NELSON, B. W. Symposium on the Environmental Chemistry of Marine Sediments. Important aspects of estuarine sediment chemistry for benthic ecology. Marshall, N. Narragansett Marine Lab., Occ. Pub. 1962 27-41 1 OPPENHEIMER, D. H. y R. A. WARD Symposium, on Marine Microbiology Release and capillary movement of phosphorus in exposed tidal sediments. Oppenheimer, C. H. 1963 664-673 PEARSE, A. S. y G. GUNTER Treatise on Marine Ecology and Paleoecology. Salinity Hedgpeth, J. W. Geol. Soc. America, Mem. 1957 129-158 67, 1 PETERSEN, C. G. J. Rep. Dan. biol. Stn. Valuation of the sea. II. The animal communities of the sea bottom and their importance for marine zoogeography. 1913 1-42 21 ---, Rep. Dan. biol. Stn. The sea bottom and its production of fish foods; a survey of the work done in conexion with valuation of Danish waters from 1883-1917. 1918 1-62 25 PHLEGER, F. R., . , .: . Lagunas Costeras. Un Simposio. Mem. Simp. Intern. Lagunas Costeras. Some general features of coastal lagoons. Ayala-Castañares A. y F. B. Phleger UNAM-UNESCO Nov. 2830, 1967, 1969 5-25 POHLO, R. H. Feeding and associated functional morphology in Tagelus californianus and Florimetis obesa (Biv. Tellinacea). Malacologia 1973 1-11 12 (1) PRITCHARD, D. W. What is an Estuary: Physical viewpoint. Lauff, G. H. Estuaries. Publ. Amer. Ass. Advanc. Sci. 1967 3-5 83 PURDY, E. G. Approaches to Paleoecology. Sediments as substrates. Imbrie. J. y N. D.. Newell John Wiley and Sons New York 1964 238-271 RAMÍREZ-GRÁNADOS, R. Estudio ecológico preliminar de las lagunas costeras cercanas a Acapulco, Gro. Revta. Soc. mex. Hist. Nat. 1952 199-218 13 SANDERS, H. L. Oceanography of Long Island Sound, 195254. Part X; The biology of marine bottom communities. Bull. Bingham oceanogr. Coll. 1956 345-414 15 ---, Benthic studies in Buzzards Bay. I. Animalsediment relationships. Limnol. Oceanogr. 1958 245-258 3 SCHOLLENBERGER, C. J. A rapid approximate method for determining soil organic matter. Soil. Sci. 1928 65-68 24 SEVILLA, M. L. Datos biológicos para el cultivo del ostión en Guaymas. Secretaría de Industria y Comercio. Dirección General de Pesca. 1959 87 p. STEWART, H. B. Sedimentary reflections of depositional environment in San Miguel lagoon, Baja California, México. Bull Am. Ass. Petrol. Geol. 1958 2567-2698 42 (11) STUARDO, J. y A. MARTÍNEZ Acta Politecnica Mexicana Resultados generales de una prospección de los recursos biológicos y pesqueros del sistema lagunar costero de Guerrero, México. 1975 99-115 16 (72) STUARDO, J. y A. MARTÍNEZ Relaciones entre algunos factores ecológicos y la biología de poblaciones de Crassostrea corteziensis Hertlein, 1951, de San Blas, Nayarit, México. An. Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México 1976 84-130 2 (1) STUARDO, J., A. MARTÍNEZ, A. YÁÑEZ, J. A. WEINBORN Y J. RUIZ. Prospección de los recursos biológicos y pesqueros del sistema lagunar de Guerrero y, en parte del litoral rocoso de Michoacán. An. Centro Cienc. del Mar y Limnol., Univ. Nal. Autón. México. (en prensa). 1974 TULKKI, P. Disappearance of the benthic fauna from the Basin of Bornholm (Southern Baltic) due to oxygen deficiency. Cah. Biol. mar. 1965 455-463 6 TRASK, P. D., . , Origin and environment of source sediments of petroleum. Gulf Publishing Company Houston, Texas. 1932 XV + 323 p., 38 figs. ---, Recent Marine Sediments, a Symposium. Organic content of recent marine sediments Trask, F. D. Amer. Assoc. Petrol. Geol. Tulsa Ok1ahoma 1939 428-453 VERNBERG, F. J. Dissolved gases. 9.3 Animal. O. Kinne Marine Ecology 1972 1491-1526 1 (3) VILLARROEL, M. y J. STUARDO Observaciones sobre la morfología general, musculatura y aparato digestivo en Tagelus (Tagelus) dombeii y T. (T.) longisinuatus (Tellinacea Solecurtidae). Malacologia, 16 (en prensa). 1976 WALKLEY, A. y I. A. BLACK An examination of the Degtjareff Method for determining soil organic matter and a proposed modification of the Chromic Acid Titration Method. Soil Sci. 1934 29-38 37 WARME, J. E. Lagunas costeras. Un Simposio. Mugu Lagoon, coastal southern California; origin, sediments and productivity. Ayala- Castañ ares, A. y F. B. Phleger Mem. Simp. Intern. Lagunas Costeras. UNAM-UNESCO. Nov. 28-30, 1967 1969 137-154 LAMINAS
 Lám. 1. Fig. 1. Melongena patula (Broderip y Sowerby, 1829) Alto: 165 mm. Interior de la barra de la Laguna de Nuxco, Gro. Fig. 2. Neritina cassiculum (Sowerby, 1836) Alto: 19 mm. Laguna de Tecomate, Gro. Fig. 3. Crassostrea iridescens Hanley, 1854) Largo 200 mm. La Unión. Gro. Fig. 4. Amnicola sp. Alto: 2.2 mm. Laguna de Apozahualco, Gro. Fig. 5. Mytella strigata (Hanley, 1843) Largo: 90 mm. Laguna de Nexpa o Chautengo, Gro. 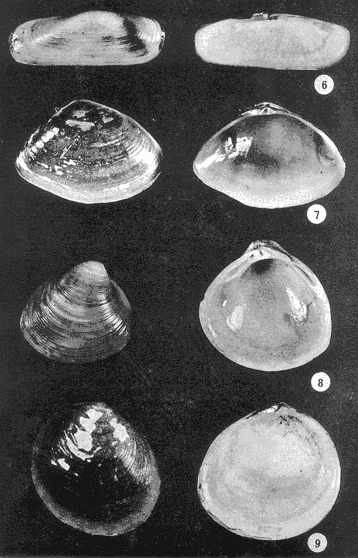 Lám. 2. Fig. 6. Tagelus (Tagelus) longisinuatus (Pilsbry y Lowe, 1932) Largo: 55 mm. Laguna de Nexpa o Chautengo, Gro. Fig. 7. Iphygenia altior (Sowerby, 1833) Largo: 35 mm. Laguna de Nexpa o Chautengo, Gro. Fig. 8. Polymesoda sp. Largo: 22 mm. Laguna de Tecomate, Gro. Fig 9. Felaniella (Zemysia) sericata (Reeve, 1850) Largo: 21 mm. Salinas de Apozahualco, Gro.
|