|
RELACIONES ENTRE ALGUNOS FACTORES ECOLÓGICOS Y LA
BIOLOGÍA, DE POBLACIONES DE CRASSOSTREA CORTEZIENSIS HERTLEIN, 1951, DE
SAN BLAS, NAYARIT, MÉXICO
Trabajo presentado al V Congreso Nacional
de Oceanografía, México, octubre 22 de 1974.
JOSÉ STUARDO
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología y Proyecto
Multinacional en Ciencias del Mar. O. E. A.
ALFREDO MARTíNEZ
Universidad Nacional Autónoma de
México, Centro de Ciencias del Mar y Limnología
Se analizan los resultados parciales de un estudio de
la biología de Crassostrea corteúensis
Hertlein, 1951 realizado entre octubre de 1971 y septiembre de 1973 en los
esteros de San Blas, Nayarit dando énfasis especial a las variaciones
hidrológicas y su efecto sobre la reproducción de esta
especie.
Se concluye, en primer lugar, que existe una
variación cíclica anual de temperatura caracterizada por una
disminución entre noviembre y marzo con un mínimo a los 23ºC
y un aumento posterior hasta un máximo de 32ºC. La salinidad es
relativamente uniforme, cercana a la del agua de mar, excepto en años
muy lluviosos en que puede llegar a 0.00%o en las partes alejadas de las bocas
de los esteros. La madurez gonádica, y en particular el desove con la
presencia consiguiente de larvas velígeras en el plancton, parecen
seguir a la variación cíclica de la temperatura.
El crecimiento de C. corteziensis
en esta área es rápido, alcanzando 70 y 75 mm de
longitud en los primeros 7 meses de vida, pero el ritmo es particularmente
rápido durante los dos primeros meses, llegando a ser de aproximadamente
40 mm de crecimiento máximo. Con posterioridad hasta los 100 o 110 mm el
ritmo puede ser igual a 10 mm de crecimiento máximo por mes.
La depredación sobre las poblaciones estudiadas
parece ser en ciertos tamaños críticos considerable, siendo
efectuada principalmente por el pez Sphaeroides annulatus,
denominado vulgarmente "botete", y en segundo término por
jaibas del género Callinectes.
Partial results of a study of the biology of
Crassostrea corteziensis Hertlein, 1951, carried out from
October, 1971 to September, 1973 in the estuaries at San Blas, Nayarit are
presented. Special emphasis is given to the hydrological changes and their
effects on the reproduction of this species.
It is concluded that there is a cyclic annual
variation of the temperature characterized by a decrease to a minimum of
23ºC, between November and March and a subsequent increase to a maximum of
about 33ºC between June and September. Salinity is fairly constant, close
to that of the sea, except in rainy years in which it can decrease at all
depths to 0.000/00 in places located not far from the inlets. Gonadic maturity
and particulary spawning, and the ensuing appeareance of veligers in the
plancton seem to follow the annual temperature fluctuation.
Growth of C. corteziensis in this
area is fast reaching 70 to 75 mm in length in the first 7 months, but this
rate is particularly high during the first 2 months, constituting about 40 mm
of maximum growth. Later, the growth rate can be up to 10 mm per month till the
oysters reach 100 or 110 mm in length.
Predation on the studied populations seems to be
critical for certain sizes. The main predator is the fish Sphaeroides
annulatus, locally called "botete", followed by crabs of the
genus Callinectes.
Este trabajo incluye parte de los resultados obtenidos durante la fase final del estudio de la posible influencia de los cambios hidrológicos producidos por el Río Santiago sobre la biología del "ostión de placer", Crassostera corteziensis en San Blas, Nayarit, México. Los resultados generales han sido discutidos recientemente por Stuardo, Martínez y Escobar (1973). Los únicos estudios publicados con anterioridad sobre la biología y ecología de esta especie son los de Sevilla (1959) y Ramírez y Sevilla (1965) efectuados en poblaciones de Guaymas, Sonora. Se trata de una de las especies de mayor importancia económica entre los ostréidos del Pacífico mexicano, y la constatación de diferencias apreciables en su comportamiento biológico y ecológico a lo largo de su distribución, al menos en la costa mexicana, hace necesario el análisis y profundización de estos estudios, lo que constituye el objetivo principal de esta nueva contribución. ÁREA DE ESTUDIOLa región de estudio se encuentra en la llanura costera del Estado de Nayarit, más o menos en el cruce del paralelo 21º35´N, con el meridiano 105º20´W (Mapa l). En forma general el área corresponde a una llanura de depositación marina, compuesta de sedimentos del Cuaternario. En esta zona los cambios de nivel del mar han provocado un aumento en el rango de depositación a lo largo de la línea de costa aumentando, por lo tanto, la amplitud de la planicie aluvial y formando un conjunto de bermas semiparalelas por acarreo a lo largo de las playas. Cortando esta planicie se encuentran los esteros (fide Lankford, R. R. y R. Saenger in Stuardo et al., 1973). La llanura costera nayarita presenta, además, lagunas dispersas entre planicies inundadas por ríos (Curray et al., 1969). Comprende la mayoría del Estado de Nayarit y parte sur de Sinaloa en la costa del Pacífico de México. El río más grande es el río Santiago que nace en el lago de Chapala cerca de Guadalajara, el cual es alimentado por el río Lerma que nace cerca de la ciudad de México. Este río tiene un efecto profundo sobre la naturaleza de la región, ya que aporta la mayor parte de los sedimentos incluidos en la llanura costera. El clima de la región es subtropical-tropical con temperatura media anual de 25ºC, rangos anuales de lluvia cercanos a los 1660 mm. y vientos dominantes del noroeste en los meses de invierno y del oeste a suroeste en el verano. La época de lluvias se remonta hasta fines del verano y principios, de otoño, frecuentemente acompañada por tormentas tropicales o chubascos provenientes del Sur. Las marcas son mixtas, diurnas y semidiurnas con un rango medio anual de 0.70 m. El área de estudio original abarcó una parte de la planicie costera mencionada, aproximadamente entre el río Zauta y río Santiago (Mapa 2) quedando incluidos en ella los esteros del Pozo y Rey objetos de este estudio. MATERIALES Y MÉTODOS
a) HidrologíaLos resultados hidrológicos discutidos en este trabajo corresponden a observaciones sobre temperatura, salinidad y oxígeno obtenidas semanal o quincenalmente entre octubre de 1971 y septiembre de 1973, en un total de 10 estaciones de los esteros del Pozo y del Rey, de las cuales se seleccionaron 3 en consideración a que representan las variaciones observadas en toda el área de estudio. Estas estaciones son La Garita, Talpita y Tiburón (Mapa 2).  Mapa 1. Sitación geográfica del área de estudio 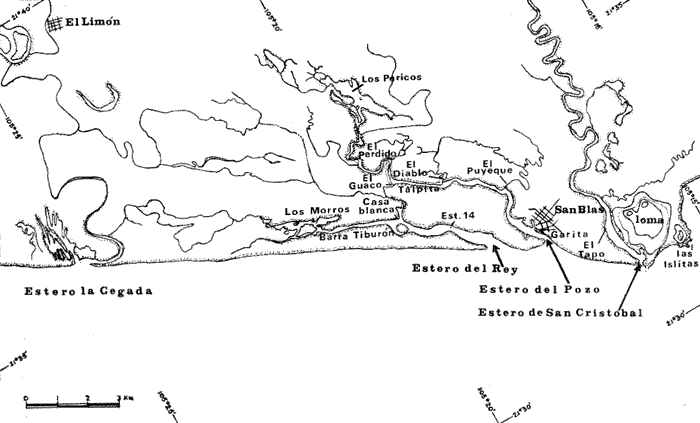 Mapa 2. Toponimia de los lugares estudiados. San Blas, Nay. Los valores de temperatura se leyeron con un termómetro de máxima-mínima con escala de -2 a 51ºC. La salinidad se determinó con un refractómetro Goldberg (American Optical Co., Modelo 10402) y el oxígeno mediante análisis por el método Winkler. El total de las observaciones discutidas corresponde a 267 muestras de salinidad, 198 de oxígeno y 267 lecturas de temperatura. b) Técnicas de cultivoLas técnicas empleadas son, en general, conocidas internacionalmente en las prácticas del cultivo ostrícola y se modificaron en mayor o menor grado para ajustarlas a las condiciones de los esteros estudiados. La falta de aguas someras a lo largo de los esteros motivó el empleo de colectores suspendidos, ya sea de un armazón de troncos y alambre cubriendo una superficie de 2 m de ancho y 2 m de largo, con una altura variable, dependiendo de la profundidad o colgando de ramas de los mangles que bordean las orillas. De esta manera, se evitó en parte la acción depredadora de diversos organismos bentónicos. Los tipos de colectores y técnicas empleadas fueron las siguientes: a) Collar de conchas. Consiste en una serie de conchas de ostión cubiertas, por una mezcla de cal-arena-cemento blanco, con proporción 3-1-1, perforadas cerca del umbo, colocadas en pares de 12 y 15 y atravesadas por un alambre galvanizado de aproximadamente un metro de largo, con tramos de manguera de plástico de 10 cm de largo entre cada par de conchas. b) Tejas encaladas (Lám. 4, b) . Colector en que se utilizan tejas de barro de las comúnmente empleadas en los techos de las casas de la localidad, de 35 cm de largo por 15 cm de ancho, en un número de 8 a 12 tejas por colector. Dichas tejas son perforadas en sus extremos y unidas en pares con alambre galvanizado formando un castillo, el cual es cubierto por una mezcla de cal-arena-cemento blanco en la proporción 3-1-1. En los últimos controles se emplearon series de 4 tejas separadas verticalmente por una distancia de 10 cm una de otra. c) Bolsas de conchas de ostión. Hechas de retazos de malla de chinchorro de 2 pulgadas de abertura, en forma de bolsa, con un volumen aproximado, de 50 1 de capacidad que permite introducir entre 50 y 200 conchas de ostión encaladas, según su tamaño. d) Rejilla o empalizada de mangle. Este colector fue empleado con resultados variables en distintas oportunidades; además de presentar una superficie de fijación considerable, es el más económico. En su construcción se emplearon 100 trozos de ramas de mangle de 50 cm de longitud por 2.5 cm de grueso aproximadamente, repartidas en 10 empalizadas, formando un enrejado cúbico de 50 cm por lado. e) Cajas de crecimiento y engorda. (Lám. 5, b) Formadas por un armazón cuadrangular de alambrón (0.5 pulgadas de diámetro), con una longitud de 1 m, una altura de 20 cm y un ancho de 40 cm, revestida con malla de alambre de 8 mm de abertura para protección contra depredadores. La parte superior de la caja corresponde a la tapa y se suspende de sus extremos mediante 2 asas. Se construyeron 4 de estas cajas ostrícolas para albergar ejemplares provenientes tanto de colectores como de los manglares, y fueron instaladas en 3 áreas, repartidas de la siguiente forma: 2 en La Garita, 1 en Casa Blanca y 1 en la Vena del Diablo. f) Corral de protección. (Lám. 5, a.) Construido sobre el fondo ocupando un área de 12 x 6 m, en aguas del Estero del Pozo (frente a La Garita), estaba formado por empalizadas de 200 cm de altura de malla de alambre de 5 cm de abertura. Planificado para formar un banco de ostión artificial en un sector de corrientes de cierta magnitud, se cubrió el fondo con una cama de conchas. Posteriormente, se fueron depositando en él, colectores con fijaciones y "semilla" procedente de mangle y algunos ostiones adultos, a fin de enriquecer el banco experimental. Este sistema no dio resultado para el área en cuestión, pues se constató un elevado índice de predación y un azolvamiento de las conchas del fondo, las que finalmente fueron totalmente cubiertas por sedimentos. La tabla 6 resume la fecha de instalación, tipo, localidad y permanencia en el agua de los diversos grupos de colectores instalados entre noviembre de 1971 y noviembre de 1972. La información obtenida de colectores instalados con posterioridad, aún en procesamiento, se presenta en las tablas 2 , 3 y 5, y es comentada aquí en mayor o menor amplitud. C) OtrosEn el estudio histológico de las gónadas se siguió la técnica de tinción por hematoxilina-eosina. El número de ejemplares estudiados mensualmente se indica en la tabla 1. La recolección de larvas se efectuó en una columna horizontal de agua de 50 m con una red de plancton de tela de molino No. 25, con abertura de malla de 63 micras, de forma cónica con una boca de entrada de 30 cm de diámetro, una longitud de 1 m y una boca terminal colectora de 5 cm de diámetro. La medición de juveniles y adultos se efectuó con un vernier o escalímetro de precisión. RESULTADOS
HIDROLOGÍALa información hidrológica obtenida a través de los muestreos continuos de temperatura, salinidad y oxígeno ya descritos tuvieron por objeto el proporcionar un panorama general de la variación de estos parámetros, en los esteros estudiados. Es importante mencionar que la variación de los parámetros de temperatura, salinidad y oxígeno no presentó diferencias considerables entre cinco localidades estudiadas, entre 1971 y 1973 (La Garita, Barra de Talpita, Barra de Tiburón, Los Morros y Laguna Los Pericos) (Mapa 2) y que de manera general sigue a las variaciones estacionales de la región. En consecuencia, para la discusión de los resultados se han seleccionanado 3 estaciones de muestreo (Garita, Talpita y Tiburón), que cubren y representan a grandes rasgos el área en cuestión (Figs. 1, 2 y 3). 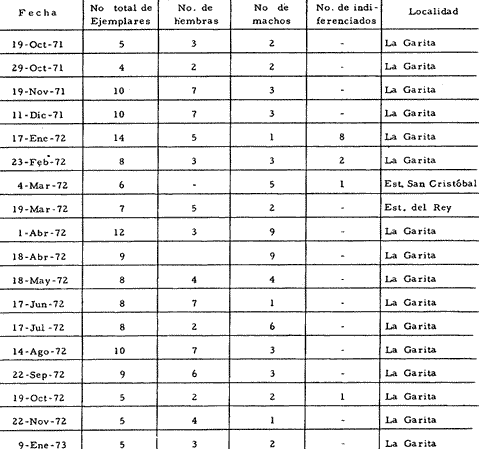 TABLA 1 PROCEDENCIA, FECHA Y SEXO DE LOS EJEMPLARES DE C. CORTEZIENSIS DE SAN BLAS, NAY. UTILIZADAS EN LOS ESTUDIOS HISTOLÓGICOS DE CICLO GONÁDICO ENTRE OCTUBRE DE 1971 Y DICIEMBRE DE 1972 Destaca en primer lugar la presencia de 2 años lluviosos y uno seco, situación que se refleja en las curvas, de salinidad constatándose que a finales de 1971 y 1973 la salinidad se abate hasta menos de 5 %0 mientras que para la misma época en 1972 no alcanza sino hasta 22%0. Es importante hacer ver que comúnmente las diferencias entre los valores de superficie y fondo nunca fueron muy marcadas, a pesar de encontrarse profundidades hasta de 7.0 m. Este hecho nos permite abreviar la información obtenida en las determinaciones de fondo, considerando además que las poblaciones del "ostión de placer" (Crassostrea corteziensis) objeto de esta publicación se encuentran sólo en los primeros 45 a 50 cms. de profundidad (zona medio litoral), sobre raíces de mangle. Con respecto al oxígeno las mismas figuras muestran que los valores obtenidos no representan un factor limitante para el progreso de la vida en estos esteros ya que comúnmente encontramos valores localizados entre 2 y 5 ml/l. . La temperatura presentó una variación anual de aproximadamente 8ºC con los valores mínimos detectados entre diciembre y abril, y los máximos entre mayo y noviembre (Figs. 1, 2, 3 y 6). La discusión de los resultados se hará bajo el siguiente orden: Garita, Talpita y Tiburón. La Garita (Fig. 1)Esta estación muestra claramente lo que sucede en la entrada (boca) de los esteros circunvecinos a San Blas, Nayarit, pues presenta condiciones marinas con cambios no muy pronunciados lo que determina cierta estabilidad ambiental en el tiempo. Esta situación se refleja en el establecimiento y supervivencia de poblaciones del "ostión de roca" (Crassostrea iridescens) en esta localidad, una especie esencialmente marina. La temperatura sigue completamente las variaciones estacionales, encontrándose los máximos térmicos hacia los meses de junio a octubre con lecturas de 30 a 32°C y los mínimos de noviembre a abril (del año siguiente) con valores de 23 a 26ºC. La diferencia observada entre años más o menos lluviosos, se refleja también en este parámetro ya que durante 1972, un año seco, la temperatura no alcanzó a descender más allá de 24ºC. Las diferencias de temperatura del agua entre superficie y fondo casi nunca fueron mayores de 1ºC. En esta estación la salinidad varió muy poco a lo largo del tiempo, como resultado de su proximidad al mar. Esto condicionó una influencia francamente marina, alterada solamente por la época de lluvias, con valores localizados entre 31 y 35 %0 la mayor parte del tiempo (noviembre 1971 a junio 1973) excepto en noviembre de 1972 en que descendió a 27 %0 Durante la temporada de lluvias observamos abatimientos considerables (de este parámetro) hasta 0 %0 como ocurrió a fines de 1971 y en agosto y septiembre de 1973. En relación a la cantidad de oxígeno se manifestaron los valores más elevados para el área en cuestión, ubicados predominantemente entre 3 y 5 ml / 1 con algunos descensos que no excedieron a los 2.5 ml / 1, lo que implica la presencia de aguas bien oxigenadas. Barra de Talpita (Fig. 2)La elección de esta estación de muestreo, así como la de Barra de Tiburón, se basó en el hecho de que por su posición, representan lo que acontece en el interior de los esteros del Pozo y Rey. Asimismo son importantes porque constituyen las áreas de fijación óptimas de la larva del "ostión de placer" (Crassostrea corteziensis). La variación de los parámetros hidrológicos seleccionados sigue casi completamente a la descrita para La Garita, acentuándose algunos cambios en temperatura, salinidad y oxígeno, por la distancia que existe entre el mar y esta estación, así como por hallarse más influenciada en la época de lluvias, por el aporte de agua dulce proveniente del Río Santiago. La temperatura osciló entre 22 y 34ºC presentándose lógicamente los valores mínimos en la época invernal y los máximos en verano. Es evidente la influencia de las estaciones del año en este parámetro y su trascendencia en el ciclo de vida del ostión es definitiva como se discute más adelante. La salinidad varió un poco con respecto a La Garita ya que la mayoría de los valores estuvieron localizados entre 25 y 36 %0 a excepción de aquellos encontrados durante la temporada de lluvias, que descendieron hasta 2 %0 en octubre de 1971 y a 0%0 en agosto y septiembre de 1973. Se considera que la variación de este parámetro en la localidad estudiada, es trascendental para la supervivencia del "ostión de placer", ya que cuando se prolonga por periodos considerables como fue el caso de julio a septiembre de 1973 se aprecian grandes mortalidades del mismo. De manera general, los niveles de oxígeno fueron sólo ligeramente más bajos que en La Garita (entre 2 y 5 ml / 1) . Barra de Tiburón (Fig. 3)En esta localidad el agua de mar tiene un acceso más directo que en la anterior, por lo que comparte características comunes con La Garita, de influencia marítima muy marcada. Aun en años lluviosos esta condición se refleja en cierta estabilidad salina con cambios no muy pronunciados y predominancia de valores entre 28 y 37 %0. 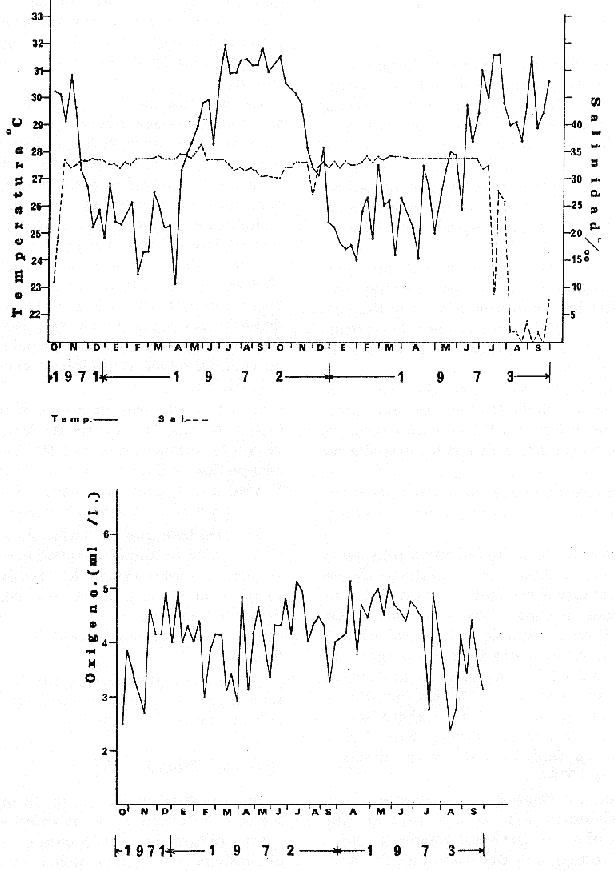 Fig. 1. Variación de salinidad, temperatura y oxígeno entre octubre de 1971 y septiembre de 1973 en La Garita, Estero del Pozo, San Blas, Nay. 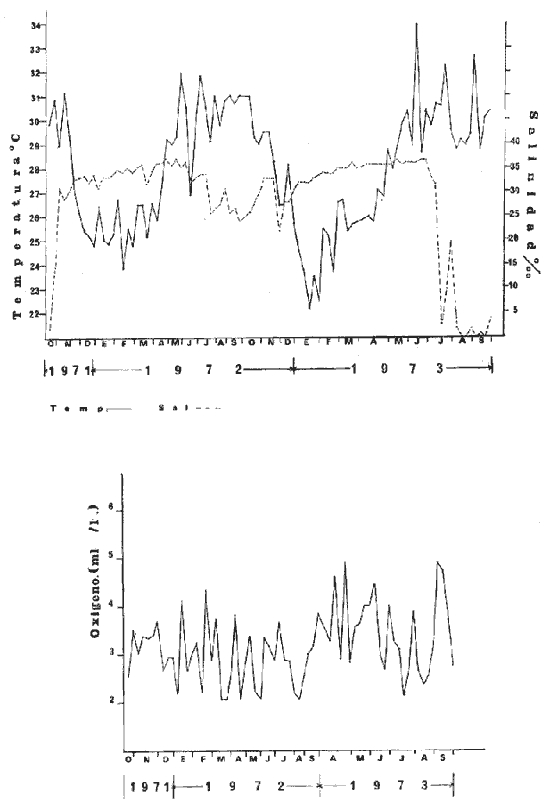 Fig. 2. Variación de salinidad, temperatura y oxígeno entre octubre de 1971 y septiembre de 1973 en Barra de Talpita, Estero del Pozo, San blas, Nay. 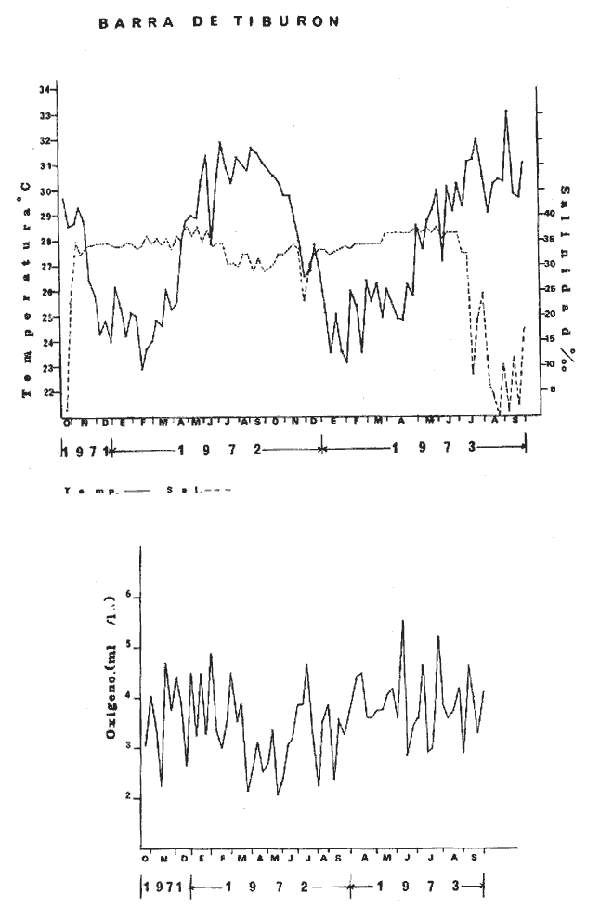 Fig. 3. Variación de salinidad, temperatura y oxígeno enre octubre de 1971 y septiembre de 1973 en Barra de Tiburón, Estero del Rey, San Blas, Nay. Las variaciones de temperatura y oxígeno no representan una excepción al panorama general descrito para las otras dos localidades, lo que en última instancia constituye uno de los factores primordiales que determinan el desarrollo de poblaciones importantes de la especie en cuestión. Las corrientes de marea que determinan en estas mismas áreas una gran concentración de larvas y, en consecuencia, producen los índices de fijación más altos obtenidos, representan junto al sustrato los otros dos factores importantes. POSICIÓN TAXONÓMICA DE LOS OSTIONES DE SAN BLAS, NAYARITLa posición taxonómica de las ostras del Pacífico mexicano y de otras especies de la familia Ostreidae en general es conflictiva, a pesar de que existen antecedentes de tipo anatómico, que ya han contribuido a aclararla considerablemente. Desgraciadamente de tiempo en tiempo aparecen publicaciones importantes que los ignoran basándose exclusivamente en observaciones de tipo conchológico. Éste es el caso de algunas publicaciones de gran divulgación como las, de Olsson (1961) y Keen (1971) que, utilizando solamente características de la concha, contribuyen a mantener la anarquía. De acuerdo, con estos autores habrían 9 o 10 especies de "ostiones", descritos para la costa mexicana del Pacífico: 1)Ostrea angelica Rochebrune, 1895. 2)Ostrea columbiensis Hanley, 1846, incluída en el subgénero Crassostrea por Olsson (1961). 3)Ostrea conchaphila Carpenter, 1857. 4)Ostrea corteziensis Hertlein, 1951, incluída en el subgénero Crassostrea por Olsson (1961). (Lám. 1). 5)Ostrea fisheri Dall, 1914, impropiamente incluída en el subgénero Alectryonia por Olsson (1961). 6)Ostrea iridescens Hanley, 1854. (Lám. 2). 7)Ostrea (Lopha) megodon Hanley, 1846, incluída en el subgénero Alectryonia por Olsson (1961). 8)Ostrea palmula Carpenter, 1857, impropiamente incluída en el subgénero Alectryonia por Olsson (1961). (Lám.3) 9)Ostrea tubulifera Dall, 1914. 10) Ostrea serra Dall, 1914. 11) Ostrea lurida Carpenter, 1864. Entre los autores mexicanos Ramírez y Sevilla (1965), muy acertadamente han insistido en la necesidad de considerar caracteres anatómicos, y sobre todo, la presencia o, ausencia de cámara promial, carácter ya sancionado por los especialistas en el grupo, para diferenciar a especies de los géneros Ostrea y Crassostrea. Desgraciadamente, los estudios de estos autores no han incluido a todas las especies descritas para el Pacífico Mexicano y, en consecuencia, no han podido evaluar críticamente la validez de algunas de ellas. En el género Crassostrea incluyeron a las especies: C. corteziensis C. iridescens y C. palmula. En el género Ostrea a: O. angelica, O. megodon, O. fisheri y O. veatchii (una especie fósil de dudosa presencia en el Reciente). Estudios anatómicos en curso. corroboran la asignación de las especies corteziensis, palmula e iridescens al género Crassostrea y de angelica y lurida al género Ostrea. La posición taxonómica de la especie fisheri no podrá precisarse mientras no se estudien las partes blandas y la especie megodon, sin cámara promial, debe separse en un taxon aparte, que puede corresponder al génera Lopha Röding; 1798 (=Alectryonia Fischer de Waldheim) en el que hasta a.hora se han incluido las especies Ostrea folium Linné, 1758 del Atlántico tropical y subtropical, y O. cristagalli Linné, 1758 del Indopacífico. Thomson (1954) ha sugerido que estas dos especies serían sólo "ecomorfos" de una misma especie, punto de vista aceptado también por Stenzel (1971) pero deberán efectuarse estudios morfológicos completos, para fundamentar esta sugerencia. Las características de la especie megodon también concuerdan bastante con el género Agerostrea Vyalov, 1936 limitado, sin embargo, al Cretácico, Superior. Su asignación precisa a alguno de estos géneros requiere el completar estudios ahora en curso. Respecto de las otras especies, la validez de O. conchaphila, O. tubulifera y O. columbiensis es dudosa y probablemente corresponden a alguna de las otras especies descritas. O. serra no ha vuelto a citarse después de la descripción original. La mayoría de estas especies tienen una distribución geográfica que se extiende desde Baja California o el Golfo de California al norte del Perú, siguiendo los límites de la fauna marina litoral tropical y subtropical del Pacífico Oriental. Ostrea angelica parece estar limitada en su distribución al Golfo de California y parte sur de la Península de Baja California. Ostrea lurida, una especie de aguas frío-temperadas de la costa de Estados Unidos y Canadá, no llega probablemente más al sur del paralelo 26ºN en la costa oeste de Baja California, aunque ha sido citada hasta el Cabo San Lucas hacia el sur. La distribución de estas especies no es continua ni homogénea, sino que depende de sus hábitos de vida. Pueden agruparse así en tres grandes grupos: a) Especies de manglar y aguas salobres b) Especies de mar abierto c) Especies de habitats intermedios a) Especies de manglar y aguas salobres1º Crassostrea corteziensis (Hertlein, 1951) "ostión de placer". En apariencia la distribución de esta especie sigue estrictamente las zonas de manglar, pero no se encuentra en las costas expuestas al mar abierto. Su distribución parece depender esencialmente de condiciones estuarinas más o menos estables y aguas tranquilas. 2º Crassostrea palmula (Carpenter, 1857) ocurre en zonas de manglares en una p:roporción numéricamente muy inferior a C. corteziensis, pero se encuentra de preferencia en costas rocosas expuestas al mar abierto. Ramírez y Sevilla (1965) mencionan que Ostrea angelica Rochebrune, 1895 sería una especie generalmente asociada con C. corteziensis y C. palmula, pero esto no se observa en la costa de Nayarit, probablemente debido a límites de distribución. b) Especies de mar abierto1º Crassostrea iridescens (Hanley, 1854) "ostión de roca". La distribución de esta especie sigue las costas rocosas de mar abierto, muy expuestas al oleaje por debajo de la zona de mareas. En esteros penetra áreas rocosas en las entradas donde prevalecen condiciones esencialmente marinas. 2º Ostrea fisheri (Dall, 1914), además de ser muy escasa parece tener una distribución muy irregular bajo la zona de mareas. 3º Crassostrea palmula (Carpenter, 1857) "goygoqui". Esta especie como, C. iridescens se distribuye a lo largo de las costas rocosas pero en pozas litorales de la zona de mareas. Ya se ha mencionado que penetra en los esteros con mangle y resiste aguas salobres. 4º Ostrea angelica (Rochebrune, 1895). Esta especie ha sido citada por algunos autores con una distribución que comprende desde el Golfo de California a Ecuador. En realidad, su distribución parece ser mucho más restringida limitándose al Golfo de California en donde Rochebrune la describió originalmente formando bancos de considerable extensión (?) . Las especies O. conchaphila Carpenter, O. tubulifera Dall y O. serra han sido citadas de mar abierto. Aparte de O. serra de valor para la discusión de problemas taxonómicos y biogeográficos, las otras dos especies carecen de importancia. c) Especies de habitats intermedios1º Lopha (?) megodon (Hanley, 1946). La distribución de esta especie en México es enigmática. Ha sido encontrada con cierta abundancia en Bahía Magdalena, B. C., donde ha sido dragada entre 3 y 10 brazas. Otros reportes de ejemplares aislados la indican dragada en profundidades de 16 hasta 100 brazas. En la costa central mexicana, los autores han encontrado un solo ejemplar en las cercanías de la boca de la laguna de Chautengo, en Guerrero, fija a raíces de mangle. Especies de importancia económicaDe todas las especies mencionadas sólo Crassostrea corteziensis, C. iridescens y Ostrea lurida.,tienen importancia económica por su abundancia. Las dos primeras por su amplia distribución son las más importantes para el desarrollo ostrícola en México. Ostrea lurida, muy abundante en la costa noroccidental de Baja California, puede contribuir al mercado local o parcialmente al abastecimiento del mercado de los grandes centros de consumo. Lopha (?) megodon puede ser importante en la explotación ostrícola de algunos estados (Baja California) y aparentemente ya se ha ensayado el enlatado de ejemplares de poblaciones provenientes de Bahía Magdalena y alrededor de Bahía Sebastián Víszcaino, B. C., pero el desconocimiento total de su biología y la uniformación muy general sobre su distribución hace necesario estudiarla para concretar las perspectivas de su explotación. En consecuencia, las especies que deben considerarse en un programa de desarrollo ostrícola inmediato en la costa Pacífica del país, son las tres primeras y en especial C. corteziensis y C. iridescens. Estas dos son también las especies más importantes en el área de San Blas, Nayarit, y el desarrollo ostrícola local debe concretarse allí para establecer la potencialidad del cultivo intensivo de ambas especies. Las tres especies presentes en el área de San Blas, Nayarit pueden identificarse con ayuda de la siguiente clave: Clave para diferenciar a las especies de Crassostrea presentes en el área de San Blas, Nayarit 1 . Valvas con dentículos internos laterales a la charnela. De tamaño mediano y muy grande . . . . . . . . . 2 1´. Valvas con el margen interno liso. Sin dentículos internos laterales a la charnela. De tamaño mediano a grande.............................C. corteziensis (Lám. 1)
2. Con fuertes costillas externas en la valva
inferior, fija en forma de copa y margen interno dentado. De 60 - 75 mm de
longitud máxima.........................C. palmula
C.
palmula puede confundirse con O. angelica, pero la
presencia de cámara promial en la primera y no en la segunda permite
diferenciarlas fácilmente.
2´. Sin costillas externas. Márgenes lisos. De hasta 175 mm de longitud máxima............................C. iridescens (Lám. 2) CICLO GONÁDICO (O MADUREZ) DE CRASSOSTREA CORTEZIENSISEl estudio del ciclo gonádico de las especies cultivables es muy importante respecto, de su explotación, y, aunque como se demuestra más adelante, es un aspecto de la biología de las poblaciones que no puede aplicarse aisladamente para interpretar con exactitud periodos de desove, duración de vida larvaria y periodos de fijación es un complemento indispensable al control de la presencia de larvas en el plancton y su fijación; combinados, dan la pauta para establecer el o los periodos de explotación recomendables para una especie. Los resultados que se discuten a continuación comprenden la serie total de observaciones llevadas a cabo entre los meses de octubre de 1971 y diciembre de 1972. Información obtenida con posterioridad confirma estos resultados y será motivo de una publicación especial sobre desarrollo gonádico en C. corteziensis, C. palmula y C. iridescens. Para precisar el grado de madurez en que se encuentra la gónada objeto del estudio histológico se siguió inicialmente un patrón de 4 etapas diferenciables. Aunque el ciclo gonádico de C. corteziensis (y el de C. balmula) permiten reconocer al menos 5, se mantuvo la clasificación para alcanzar uniformidad en las conclusiones. Las 4 fases reconocidas y sus características son las siguientes: I. Fase de reposo gonádico o indiferenciada II. Fase de maduración o prereproductiva III. Fase reproductiva o de desove IV. Fase postreproductiva o de postdesove. La diferenciación de algunas de estas fases se basa esencialmente en la gónada femenina ya que el volumen de la gónada masculina y la cantidad de células sexuales observables sobre todo en la fase II y III hace difícil diferenciar las distintas etapas. I. Fase de reposo gonádico o indiferenciadaLa caracterización de esta etapa en distintas especies es variable. En algunas (C. iridescens) se caracteriza básicamente por la ausencia de células sexuales quedando solamente restos mínimos de tejido gonádico indiferenciado por lo que no se puede determinar el sexo. En otras especies (C. corteziensis, C. palmula) la gónada parece poder llegar ocasionalmente a un estado de regresión similar; pero en la mayoría de los ejemplares observados aparecen restos de gónada en islotes con óvulos o espermatozoides que parecen no ser reabsorbidos totalmente, permaneciendo en este estado hasta el inicio de la etapa de maduración siguiente. En todos los casos la gran abundancia de tejido conjuntivo que ocupa los espacios dejados por el tejido gonádico es característica. Es dentro de esta masa de tejido conjuntivo que se observan los islotes de epitelio gonádico en reposo. II. Fase de maduración o prereproductivaEsta fase implica prácticamente dos etapas diferentes que aquí no se han diferenciado. En la primera, la gónada se caracteriza por presentar gran actividad gametogénica. El epitelio germinal se hace activo formando abundantes células sexuales que se disponen uniformemente en la luz de los conductos. El tejido conjuntivo que se halla en su máximo desarrollo a comienzos de esta fase, empieza a disminuir siendo reemplazado por tejido gonádico y los folículos y gonoductos crecen hasta tocarse. En la etapa siguiente, el tejido reproductor ha alcanzado su máximo desarrollo y la gónada aparece como una masa compacta con folículos totalmente distendidos, uniforme, que ha desplazado totalmente al tejido conjuntivo. En esta etapa hay ejemplares en los que se nota todavía acentuada actividad gametogénica que se continúa aun en la fase siguiente. III. Fase reproductiva o de desoveSe inicia con la expulsión de óvulos y espermios y su progresión se aprecia por la reducción en la cantidad de gametos y la aparición de espacios vacíos ocupando un volumen variable en distintos ejemplares. Es conocido que todos los ejemplares de una población no desovan simultáneamente, ni que todos presentan el mismo comportamiento, lo que se demuestra en la histología de sus gónadas. Así, se observan ejemplares en los que la gametogénesis prosigue acentuadamente durante esta fase sugiriendo que un mismo ejemplar puede desovar durante un periodo largo o más de una vez. Las causas de este comportamiento son aún imprecisas. Aquí también es factible diferenciar sobre todo en las hembras una etapa de desove parcial y otra de desove avanzado. IV. Fase postreproductiva o postdesoveCaracterizada por una combinación de cambios importantes: grandes espacios, con restos de células sexuales de apariencia variable (desde tamaño normal a rotas) , invasión de fagocitos y absorción o degeneración del tejido gonádico. El grado máximo de degeneración se presenta cuando los conductos se observan vacíos, muy contraídos o cuando las paredes se han destruido formando grandes lagunas, o bien cuando sus paredes se ponen en contacto tomando forma alargada. COMPORTAMIENTO MENSUAL DE LAS POBLACIONESLos resultados que se dan a continuación corresponden a una descripción mensual de la evolución gonádica de Crassostrea corteziensis basada en la caracterización de las fases anteriores. La Tabla 1 indica la procedencia, localidad y fecha de las muestras estudiadas y el número de ejemplares incluidos en cada una; la figura 4 representa los resultados mensuales observados, los que pueden resumirse de la manera siguiente: OCTUBRE (1971)La mayoría de los ejemplares del mes de octubre se encontraban en una etapa de desove avanzado, caracterizada por la presencia de folículos fusionados en donde se localizan gran cantidad de óvulos remanentes en su luz que no presentaban fenómenos de citólisis (deformación del citoplasma; pérdida de las membranas celulares). En algunos ejemplares se observaron folículos con elementos sexuales adheridos a la pared folicular o sea que había una leve actividad gametogénica. Fig. 4. Ciclo gonádico de C. corteziensis y parcial de C. palmula y C. iridescens en poblaciones de San Blas, Nay. 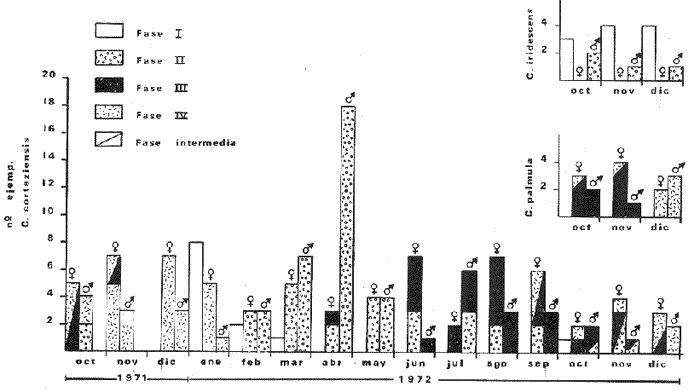 Fig. 4. Ciclo gonádico de C. corteziensis y parcial de C. palmula y C. iridescens en poblaciones de San Blas, Nay. En otros especímenes el tejido gonádico presentó aspecto desorganizado, caracterizado por: presencia de folículos contraídos, pared folicular destruida y grandes espacios en su luz, con óvulos en su mayoría mostrando fenómenos de citólisis; las células fagocíticas son escasas. Estos fenómenos son característicos de la etapa inmediata al desove avanzado, o sea que se estaba iniciando la fase IV o postreproductiva. NOVIEMBRE (1971)Continuaba la fase IV o postreproductiva iniciada en la segunda quincena del mes de octubre. Los fenómenos que caracterizan a esta etapa son: presencia de fagocitos que invaden la gónada para penetrar a los conductos y fagocitar los gametos no desovados, folículos contraídos cuya pared se destruye y fusiona con la de otros folículos. FEBRERO (1972)En la mayoría de los especímenes del mes de diciembre se identificaban células en vías de degeneración, con aparente deformación de citoplasma y pérdida de la membrana nuclear; las reacciones de citólisis eran más aparentes en la porción central de los conductos. Se observaba, además, la presencia de numerosas células fagocíticas que se hacen aparentes en el tejido intersticial. Los fenómenos de citólisis y reabsorción avanzada de la gónada (se observó sólo en algunos ejemplares) se caracterizaban por la presencia de conductos casi vacíos que en los especímenes machos adquieren forma alargada, sobre todo hacia la periferia de la gónada; en las hembras, dichos conductos o presentaban su pared destruida o habían sufrido una contracción, localizándose en su interior de uno a cinco óvulos. En la mayoría de las preparaciones histológicas de este mes, tanto la citólisis como la reabsorción de las gónadas no podían considerarse completas. ENERO (1972)Lo más sobresaliente de las observaciones realizadas en este mes es que, de los catorce especímenes observados, en 8 no se determinó sexo por no localizarse tejido reproductor, siendo el tejido conectivo vesicular muy abundante. Probablemente esta fase indiferenciada o de inactividad de la gónada se inició a fines del mes de diciembre. En los ejemplares hembras se determinó el sexo sólo por la presencia de restos de óvulos. FEBRERO (1972)El epitelio germinal iniciaba su aparición en el tejido conectivo el cual se observó abundante y bien organizado. En las hembras habían ovogonias adosadas a la pared del folículo cuya luz era pequeña. En los machos se observó epitelio germinal en actividad; los folículos no eran de gran diámetro y se disponían en una banda hacia la periferia de la gónada. No se determinó sexo en 2 ejemplares. MARZO (1972)La actividad gametogénica era evidente en ejemplares machos y hembras. En los machos los folículos empezaban a proliferar y aumentar su diámetro. Se observaron todas las fases de la gametogénesis, predominando la de espermatocitos de primero y segundo orden. El tejido interfolicular era aún abundante. La proliferación folicular y el fenómeno de gametogénesis era más avanzado en los machos que en las hembras. ABRIL (1972)Continuaba la fase de gametogénesis iniciada en el mes anterior. En los ejemplares hembras había actividad gametogénica y formación de folículos. Los ovocitos presentaban un nucléolo y cromatina reticular. En los especímenes machos, el tejido reproductor se ramificaba con abundantes espermatogonias pero predominaban los espermatocitos. MAYO (1972)El tejido conectivo disminuía progresivamente, localizándose sobre todo hacia la periferia. Los gónadas en ambos sexos presentaban aspecto maduro. En general, puede considerarse que la gónada se encontraba en una etapa de madurez previa al desove. En ejemplares hembras se observaron ovogonias adheridas a la pared folicular y numerosos óvulos en su luz; las paredes foliculares aparecían fusionadas en su mayoría. En ejemplares machos la fase de la gametogénesis predominante era la de espermatidias y espermatozoides; las paredes foliculares se observaban en estrecho contacto. No había signos de desove. JUNIO (1972)La mayoría de los ejemplares observados presentaron signos de desove parcial; en una minoría se observaba actividad gametogénica con signos de desove. Durante este mes parece haber ocurrido un desove parcial probablemente iniciado, en la primera quincena del mes de junio. JULIO (1972)Continuó la expulsión de elementos sexuales aparentemente iniciada en el mes anterior. Se constató actividad gametogénica en ambos sexos caracterizada en las hembras por la presencia de ovocitos adheridos a la membrana basal y en los machos, por presentar una capa de espermatogonias alrededor del folículo. AGOSTO (1972)Los signos de desove son más acentuados en los ejemplares hembras que en los machos. La mayoría de las hembras examinadas iniciaron un desove total que es evidente por la presencia de folículos contraídos y pared folicular destruida presentando de 1 a 4 óvulos degenerados. En los ejemplares machos se observaron folículos contraídos que son signos de desove. SEPTIEMBRE (1972)A medida que el desove avanzaba se observó la constricción de los conductos y la pared folicular aparecía destruida quedando pocos gametos que no habían sido descargados y que entraban en una etapa de degeneración. No obstante, una minoría de los ejemplares presentó ligera actividad gametogénica. OCTUBRE (1972)El desove continuó, observándose en algunos ejemplares espermatogénesis parcial; otros ejemplares presentaban un postdesove marcado. En un solo ejemplar se constató una fase de indiferenciación gonádica muy marcada. C. palmula parece tener un comportamiento muy similar al de C. corteziensis, al menos en el lugar estudiado. Tanto machos como hembras se encontraban en este mes en pleno desove o desove avanzado. NOVIEMBRE (1972)El comportamiento de la gónada era similar al del mes anterior. Tanto en C. corteziensis como en C. palmula es característico un desove gonadal avanzado. DICIEMBRE (1972)En este mes la diferencia constatada en los meses anteriores era notable. Tanto C. corteziensis como C. palmula presentaban la gónada en estado de postdesove, más acentuado en la segunda especie que en la primera. En relación a lo anterior hay dos errores posibles que podrían alterar en alguna medida los resultados discutidos. El primero corresponde al número reducido de ejemplares que se utilizó para el estudio, histológico de la gónada y que pudiera no ser representativo del ciclo gonádico de la población. Un error de ese tipo es imposible de eliminar directamente, porque el elegir una muestra más representativa (mayor número de ejemplares) significaría una cantidad de trabajo de laboratorio, de procesamiento y de interpretación imposible de realizar. Aun contando con una muestra representativa, tampoco es seguro precisar en qué momento se inicia o termina el desove, lo que es fundamental para establecer las técnicas de cultivo. De ahí que sea necesario correlacionar estas observaciones con otras de presencia de larvas en el plancton y de fijación de "semilla". Estas observaciones, discutidas más adelante, nos permiten interpretar con bastante exactitud el ciclo gonádico observado. Por otra parte, el ciclo gonádico de una población es lo suficientemente general como para poder interpretarla a grandes rasgos estudiando, unos pocos individuos. El segundo error es sólo parcial y se refiere a la posible confusión en algunos casos durante los primeros seis meses, de las especies C. corteziensis y C. palmula. El estudio taxonómico realizado permitió precisar que hay sólo tres especies presentes en el área de San Blas, Nay.: C. corteziensis, C. palmula y C. iridescens. La similitud comprobada en estudios en curso, entre el comportamiento gonádico de las primeras dos especies mencionadas sugiere que el error, de existir, es intrascendente. En consecuencia, las siguientes conclusiones son válidas al menos para condiciones que reflejan aquéllas objeto de estudio. 1. El periodo de "reproducción" o desove en C. corteziensis cubre gran parte del año presentando, como consecuencia, diferencias considerables, respecto de la duración de éste y otros periodos del ciclo gonádico de C. corteziensis en Guaymas, Son., descritos por Sevilla (1959). Según ella la duración del ciclo es como sigue (Fig. 5) a) Fase de maduración - febrero a junio b) Fase de reposo - noviembre a fines de enero c) Fase de postdesove - septiembre a octubre d) Fase de desove - julio a agosto. Las observaciones llevadas a cabo en San Blas, Nay., (Fig. 5) demuestran que la duración de estos periodos es allí mucho mayor y que, en general, como ocurre siempre en la naturaleza, los límites de estas fases se sobreponen a veces considerablemente. La fase de desove es larga, extendiéndose a casi todo el año; sin embargo, durante 1972 la mayor abundancia de larvas se constató durante los meses de septiembre, octubre y noviembre con el máximo en octubre (Fig. 6) , y durante 1973, en los meses de julio y agosto y otra vez en octubre. Esta abundancia se determinó por colectas de plancton y controles de fijación en colectores. 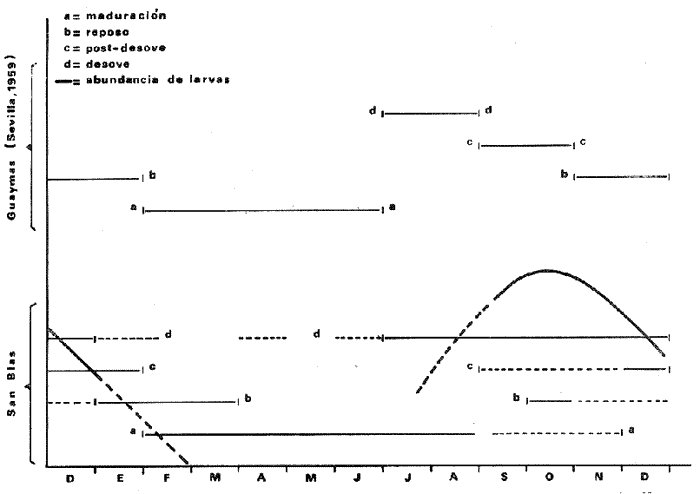 Fig. 5. Duración de las fases del ciclo gonádico de C. corteziensis en Guaymas, Son. (Sevilla, 1959) y San Blas, Nay. Igualmente larga es la fase de gametogénesis y maduración que se extiende entre enero y noviembre, aunque en septiembre y noviembre la proporción de ejemplares en esta fase es menor que la de aquellos en desove. Por otra parte, en numerosos ejemplares en desove se constata en estos mismos meses gametogénesis en diversas partes de la gónada, sugiriendo que estos mismos ejemplares pueden tener un desove muy largo o desoves parciales. La fase de reposo o indiferenciada se presenta en algunos ejemplares (probablemente aquellos que han llegado a tener un desove total) en los meses de enero a marzo, aunque es factible que, esporádicamente, aparezcan individuos aislados en esta fase durante el periodo de desove como ocurrió en el mes de octubre. Ya se ha hecho notar que la indiferenciación en el ciclo gonádico de esta especie y C. palmula no llega a tener la preponderancia que en C. iridescens. La fase de postdesove es apreciable entre septiembre y enero con una predominancia, en los ejemplares estudiados, en los meses de diciembre y enero. Es posible que las diferencias observadas con respecto a Guaymas, correspondan a efectos climáticos distintos. 2. Las poblaciones de C. palmula, por lo menos dentro de los esteros estudiados, parecen presentar un ciclo gonádico muy similar al de C. corteziensis (Fig. 4 ). 3. C. iridescens tiene un comportamiento gonádico muy diferente al de las dos especies anteriores e indudablemente tiene relación con su adaptación a la vida en mar abierto. LARVAS DE OSTIÓNLa literatura sobre larvas de ostréidos de algunas especies muy conocidas tales como Ostrea edulis de los mares europeos y Crassostrea virginica del Atlántico norteamericano y Golfo de México es abundante. Su morfología, desarrollo, ecología y cultivo han sido bien estudiadas; Korringa (1952, 1957), Walne (1964), Galtsoff (1964) y Loosanoff (1965) resumen la mayor parte de la información disponible. 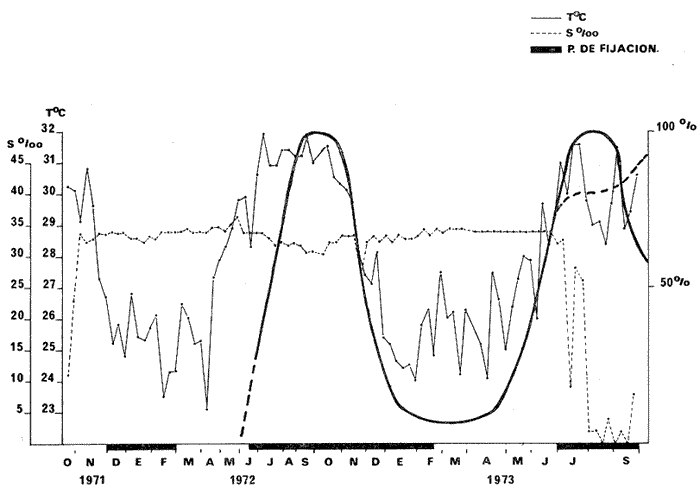 Fig. 6. Relación entre variación de temperatura y salinidad, fijación y aundancia de larvas de C. corteziensis en los eteros del Pozo y El Rey en San Blas, Nay. Valores hidrológicos correspondientes a la localidad La Garita. Por el contrario, de las especies que habitan el Pacífico mexicano sólo se conoce en algún detalle el desarrollo de Ostrea lurida por los trabajos de Stafford (1912, 1913) y Loosanoff et al. (1966). El desarrollo y los estados larvarios de las otras especies es desconocido. El valor de tales estudios desde el punto de vista práctico se refiere esencialmente al reconocimiento de la morfología y ecología de los últimos estados larvarios, denominados de "prodisoconcha I y II" o, de una manera más amplia "pediveliger", para designar aquellos estados en que estas larvas pueden nadar y reptar al mismo tiempo. También es importante conocer la duración de la vida pelágica de las larvas. En relación a la especie objeto de este trabajo el estudio de las larvas pudo iniciarse recién en octubre de 1972, en combinación con el control mensual de fijación en colectores. Sin embargo, su presencia ya se había constatado, en septiembre. En general, es un estudio que requiere de experiencia y observaciones cuidadosas debido a la gran cantidad de bivalvos que presentan estados larvarios planctónicos similares. Por ejemplo, la diferenciación del estado de veliger (larva-D) de un ostréido; cualquiera, del de otros bivalvos con larvas planctónicas es prácticamente imposible. Sólo la aparición de la charnela larvaria a medida que la "prodisoconcha" crece permitirá reconocer a una larva como perteneciente a la Superfamilia Ostreacea, pero la identificación específica y aún genérica, es imposible sin recurrir al desarrollo larvario en laboratorio y a su caracterización por mediciones cuidadosas y descripciones precisas. Aun así resolver el problema específico será difícil existiendo varias especies en la misma área. Afortunadamente, el estado de "prodisoconcha II" y "pediveliger" en muchas especies de Ostreidae, es fácilmente identificable por un desarrollo exagerado y torción de los umbos que le da a la larva en ese estado un aspecto inconfundible que ha llevado a denominarlas también como larva-umbo (lám.4, a) . justamente es necesario conocer la identificación de este estado. en el plancton para establecer las zonas de mayor abundancia de larvas y planificar la implantación de colectores. En la actualidad la identificación de la larva pediveliger de C. corteziensis nos ofrece muy pocas dudas. En primer lugar siendo C. corteziensis la especie dominante en los esteros estudiados hay un elevado margen de seguridad al identificar la larva en cuestión como perteneciente a esta especie, aún en aquellas larvas encontradas en mar abierto, frente a San Blas, Nayarit y alrededores. (No es necesario recurrir a la fecundación artificial y desarrollo en laboratorio para corroborarlo). C. palmula es escasa en los esteros y C. iridescens no se encuentra en desove en los meses estudiados, además de no encontrarse en los esteros y zonas de manglares. No siendo la caracterización morfológica de la larva uno de los objetivos de este estudio, pasaremos a discutir algunos aspectos de su ecología. Sin embargo, es interesante hacer ver la similitud de tamaño y forma constatada entre la pediveliger de C. corteziensis con larvas del mismo tipo de C. virginica descritas por Galtsoff (1964) y otros autores. Relación entre presencia de larvas, época del año y algunos parámetros abióticosEn la figura 5 y en las tablas 2 y 3 se representa la abundancia relativa de larvas pediveliger en muestras de plancton tomadas en una columna horizontal de agua de 50 m en las seis localidades siguientes: Barra Tiburón, Vena del Diablo, La Garita, Estación 14, Boca del Rey e Islitas (Mapa 2). Para verificar la presencia de larvas en los lugares mencionados y su abundancia relativa se tomó, además, una muestra de Plancton superficial durante 5 minutos. 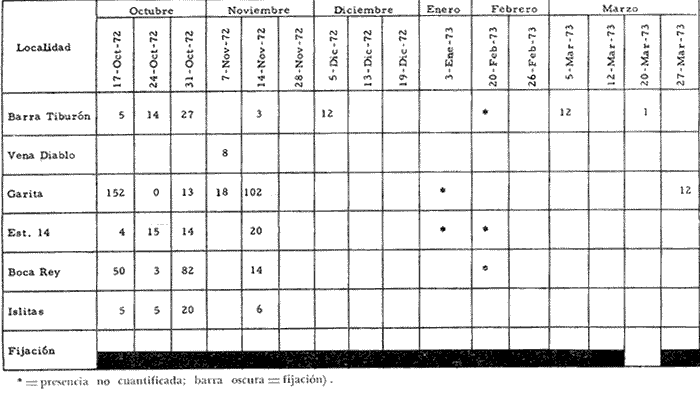 TABLA 2 TOTAL DE LARVAS EN MUESTRAS DE 125 MIL OBTENIDAS EN UNA COLUMNA HORIZONTAL DE AGUA DE 50 M EN LOS ESTEROS DEL POZO Y DEL REY, SAN BLAS, NAY. ENTRE OCTUBRE DE 1972 Y MARZO DE 1973 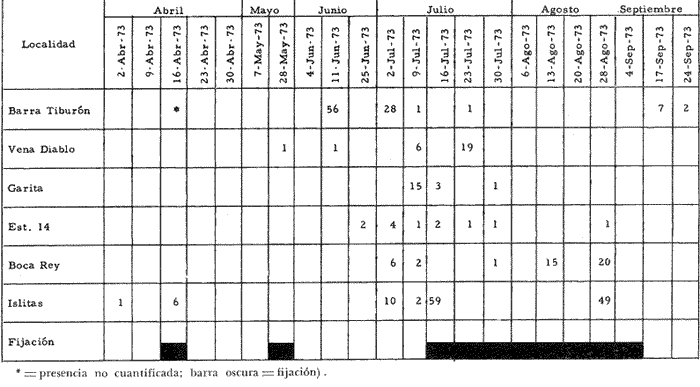 TABLA 3 TOTAL DE LARVAS EN MUESTRA DE 125 MIL OBTENIDAS EN UNA COLUMNA HORIZONTAL DE AGUA 50 M, EN LOS ESTEROS DEL POZO Y DEL REY, SAN BLAS, NAY. ENTRE ABRIL Y SEPTIEMBRE DE 1973 La curva de la figura 5 demuestra que la mayor abundancia de larvas constatada en los meses de septiembre, octubre y noviembre de 1972, con el máximo en octubre, coincide con las observaciones del ciclo gonádico y, como se discute más adelante, con los óptimos de fijación de "semilla", es decir, de larvas metamorfoseadas recién fijadas. De gran interés ha resultado el constatar que en las localidades de La Garita e Islitas se obtiene en ciertos momentos una concentración de larvas mayor que en las otras estaciones de muestreo. En ambos casos predominan larvas de concha muy oscura, es decir "viejas", sugiriendo su transporte y concentracion por efecto de corrientes de marea. La abundancia de larvas frente a la costa (Islitas) demuestra además, el "desperdicio" de larvas que al ser arrastradas por corrientes de mareas y al no encontrar condiciones adecuadas para su fijación se pierden. Indudablemente, la instalación de colectores en los esteros al captar muchas de estas larvas impediría una pérdida exagerada de este tipo para beneficio, de las pesquerías. La figura 6 muestra la abundancia relativa de las larvas, en relación a los periodos de fijación y a la variación de los valores de temperatura y salinidad encontrados en la localidad denominada Garita que, como ya se ha discutido, refleja en general las variaciones encontradas en los dos esteros estudiados. Se considera que tanto la salinidad como la temperatura ocupan un lugar muy importante en la biología de los ostréidos pues existen rangos determinados sobre o bajo los cuales las poblaciones ostrícolas de adultos y larvas de diversas especies no pueden subsistir o son afectadas en mayor o menor grado en su biología. Sólo algunas especies de ostras pueden considerarse como eurihalinas, es decir, como organismos capaces de vivir en agua de mar con un amplio rango de salinidad. Entre las eurihalinas el efecto de las variaciones de salinidad difiere según las especies y dentro de la distribución de una especie, según la latitud. Es de importancia trascendental tratar de precisar la influencia de este factor en la biología de C. corteziensis y de las otras especies que habitan el área de San Blas, Nay., pero sólo una combinación de observación y experimentación permitirá lograrlo. La escasa variación observada durante el año 1972 con valores que fluctuaron entre 29 y 36 %, en total contraste con el fin de año lluvioso de 1971, es probablemente la causa de un año tan productivo desde el punto de vista de la abundancia de larvas y fijación tanto en colectores como en el manglar. Por efecto del azar, el desarrollo del programa ha permitido constatar al menos parcialmente, los efectos de un año relativamente poco lluvioso (1972), y de dos muy lluviosos (1971 y 1973), sobre las variaciones de salinidad. Se sabe que poblaciones de ciertas especies (por ejemplo C. virginica) sufren mortalidad de larvas y adultos ya con valores de 10 %0 de salinidad, si las condiciones se mantienen por más de 10 o 15 días, y se comprobó en San Blas, Nayarit, que a fines de 1971 y a fines de 1973, mientras los valores de salinidad fueron bajos en los esteros, la mortalidad fue elevada y no se observaron fijaciones en raíces de mangles. Es posible que el desove haya sido postergado al término de las lluvias cuando la salinidad regresó a valores superiores a 15 o 20 %0, pero ésta es una hipótesis que necesita ser confirmada. Como se discute más adelante pudiera existir una relación con la temperatura, determinando un efecto combinado sobre la fisiología del "ostión" y, especialmente, sobre su ciclo reproductivo. Durante el año 1972 la variación de la salinidad fue mínima y se constató un largo periodo de desove. Las implicaciones que estas variaciones pueden significar en la explotación ostrícola se discuten más adelante. La importancia de la temperatura en la productividad de los bancos ostrícolas es conocida particularmente en relación a su capacidad reproductiva, crecimiento y engorda. Su importancia en la biología del "ostión de placer" es imprecisa, pero resulta interesante constatar que la inversión entre las curvas de temperatura y salinidad a lo largo de los años 1972 y 973 coincide con el término de la maduración y con el periodo de desove. ¿En qué medida estas variaciones han condicionado el ciclo gonádico y de reproducción en C. corteziensis? La hipótesis planteada anteriormente parece fundamentarse con la información presentada en la figura 6. En consecuencia, podemos adelantar la sugerencia de que el desove se inicia cuando el agua en los esteros después de alcanzar sus valores mínimos de temperatura en invierno y primavera (entre 23ºC y 24ºC) alcanza límites de temperatura superiores a 27ºC o 28ºC y de salinidad inferiores a 34 %0, o al menos que la supervivencia y crecimiento de las larvas pudieran verse favorecidos dentro de esos límites. Es obvio que la curva de desove sigue desfasadamente a la de temperatura. Los controles de fijación indicados en la misma figura corroboran los periodos de aparición de larvas en el plancton. Otros factores abióticos que pueden afectar a las poblaciones tales como sedimentación, corrientes y mareas no parecen tener efecto apreciable sobre las poblaciones ostrícolas, por lo menos en las de las raíces de mangle. La variación de los parámetros restantes (oxígeno, pH, carbonatos, trasparencia) tampoco es determinante. De lo anterior puede concluirse que es indudable que las condiciones hidrológicas generales se mantienen relativamente constantes y favorables al desarrollo de las poblaciones ostrícolas en los esteros del Rey y del Pozo, a pesar de que en años muy lluviosos la disminución extrema de salinidad y su duración por periodos más o menos largos (más de una semana) puede producir mortalidades masivas considerables, por lo menos en los estratos superficiales ocupados por las poblaciones de manglar (que habitan las raíces de mangle). Al respecto, la última visita a las poblaciones de los Esteros del Pozo y del Rey el 7 de noviembre de 1973, demostró que "las avenidas de agua" resultantes de los meses anteriores muy lluviosos, no exterminaron a las poblaciones de "ostión" (ostras), mientras que la mortalidad de esos ejemplares que se hallaban en la zona de marea fue aparentemente total. En esa fecha se constató una enorme cantidad de juveniles de alrededor de 1 a 2 cm de tamaño, que, indudablemente, hicieron su entrada a la población no más de un mes antes es decir, a comienzos o mediados de octubre. Cualquiera que sea su origen eran el resultado de un gran desove producido con posterioridad a las grandes avenidas de agua dulce ocurridas entre julio y septiembre. En consecuencia, es obvio que la conclusión de que hayan sido factores hidrológicos los que han determinado la gran deplesión de esta pesquería en San Blas no tiene fundamento. Indudablemente puede influir en la reducción de las poblaciones de un año a otro, pero las observaciones realizadas demuestran que la capacidad de recuperación de las mismas es excelente, a juzgar por la densidad de fijación en los diversos tipos de colectores utilizados y su capacidad de crecimiento. FIJACIÓNPara estudiar las relaciones entre madurez sexual y fijación de ejemplares y su crecimiento se estableció un programa de instalación de colectores de algunos de los tipos ya descritos (tabla 6). Un primer grupo fue instalado entre los días 30 de noviembre y 9 de diciembre de 1971 y consistió en colectores de teja, collares y bolsas. Fueron controlados prácticamente cada mes a partir del 17 de diciembre, comprendiendo la última visita el 18 de julio de 1972. Se constató fijación de larvas en los meses de noviembre y diciembre, y a juzgar por el tamaño, mínimo de los controles efectuados en los meses siguientes se presume que hubo fijaciones también en enero y febrero, lo que se constató en controles llevados a cabo con posterioridad (Fig. 7). La cantidad de "semillas" obtenida en algunos colectores, sobre todo, los de tejas, fue notable pero en este momento es aún difícil precisar las causas que motivaron el éxito de algunos colectores, sobre todo tratándose de un año (1971) particularmente lluvioso. El segundo grupo de colectores se instaló los días 2 y 3 de junio de 1972 en una fecha que se suponía podría coincidir con los primeros desoves de ese año. Es necesario hacer notar aquí las dificultades que implica el observar la aparición de las primeras fijaciones. Se ha mencionado que el tamaño de las larvas "viejas" de C. corteziensis, equivalente al de otras especies, es de aproximadamente 300 a 320 micras, un tamaño imposible de apreciar en observaciones macroscópicas (al ojo desnudo). De modo que, en el momento de su fijación y en los primeros 10 a 15 días después es muy difícil precisar su ubicación, mucho menos su abundancia. Sólo al alcanzar aproximadamente 3 a 5 mm es posible contarlas con precisión. Medirlas, continúa siendo difícil a no ser que se haga con una lente de aumento y vernier. 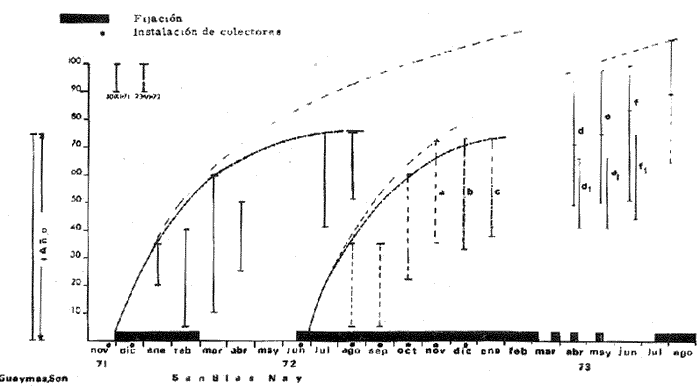 Fig. 7. Perido de fijación y crecimiento de C. corteziensis en colectores experimentales instalados en San Blas, Nay. Este grupo de colectores fue revisado los días 16 y 17 de agosto de 1972 por primera vez. Se constató, en general, una fijación escasa la que a juzgar por el tamaño máximo de algunos ejemplares (35 mm, Fig. 7) debió haberse efectuado ya a fines de junio o comienzos de julio. El día 16 de agosto se instaló, en diversas localidades, un nuevo grupo de colectores (tabla 6) constituido, únicamente por bolsas de conchas con y sin cal, con la doble finalidad de constatar fijación en agosto y establecer posibles preferencias a superficies encaladas. La revisión de estos colectores se efectuó conjuntamente con los de junio entre los días 20 y 21 de septiembre de 1972. Los recuentos y mediciones efectuados en ambos grupos y su correlación permitió establecer fechas aproximadas de fijación. En el mes de octubre se constató la mayor cantidad de fijación por colector de la temporada con valores que fluctuaron entre 1800 y 4000 ejemplares (fijaciones) por colector (4 tejas cada uno). En el mes de noviembre la magnitud de la fijación disminuyó considerablemente variando entre 80 y 288 por colector. Estos valores corresponden a colector de tejas; los de collar (30 conchas cada uno) resultan comparablemente muy inferiores. A fines de noviembre se introdujo un nuevo tipo de colector, hecho de ramas de mangle, en las mismas localidades estudiadas, dada la gran abundancia de "semilla" constatada en las raíces de los árboles de mangle que bordean el estero. Un control efectuado el 3 de enero en los colectores de este tipo, instalados en Barra de Tiburón y Barra Talpita, dio un promedio de 40 fijaciones por trozo para la primera y 20 para la segunda, lo que aproximadamente significa 4000 y 2000 "semillas" por colector, respectivamente, una cifra que para lo avanzado de la temporada de desove indica la gran cantidad de larvas aún presentes en el plancton y las perspectivas de instalar colectores que garanticen una nueva cosecha aun durante el mes de diciembre. El éxito de la instalación depende indudablemente de la elección de lugares en donde se constate mayor abundancia y/o concentración de larvas. La información que se discute a continuación tiende a aclarar estas afirmaciones. Los tamaños encontrados tanto en los colectores de junio como en los de agosto, demuestran que hubo fijación en estos meses, pero que fue mayor en agosto en coincidencia con los periodos de desove. Si hay o no fijación en junio, es decir, si hay o no larvas en ese mes es todavía incierto y deberá precisarse en muestras de plancton tomadas, ex profeso. Sin embargo, una extrapolación de la curva de crecimiento de los ejemplares encontrados los días 16 y 17 de agosto, sugiere que hubo fijación en junio (Fig. 7). La comparación de la efectividad de superficies encaladas versus no encaladas en colectores de bolsas, demostró escasas diferencias que son conflictivas y pueden deberse a diversos factores tales como ubicación del colector, profundidad, número de conchas utilizadas, etcétera. En general, se constata una preferencia de las larvas por colectores encalados. Los tres grupos de colectores instalados el 24 de septiembre, 25 de octubre y 25 de noviembre respectivamente tenían por objeto constatar fijación en cada mes si la hubiera y crecimiento por mes. Para facilitar su control se utilizaron aquí sólo colectores de tejas y sartas de conchas, que se ubicaron en cinco localidades que se considera representan zonas de óptima fijación: Barra de Tiburón, Vena del Perdido, Barra Talpita, Vena del Diablo, Vena del Puyeque y La Garita (Mapa 2). Aparte de su reemplazo mensual por una nueva serie se controló en ellos semanalmente la abundancia. Resultados parciales de algunos de ellos, representados en las figuras 8 y 9 se discuten a continuación. Los resultados obtenidos con posterioridad en colectores de tejas se presentan en la tabla 4. 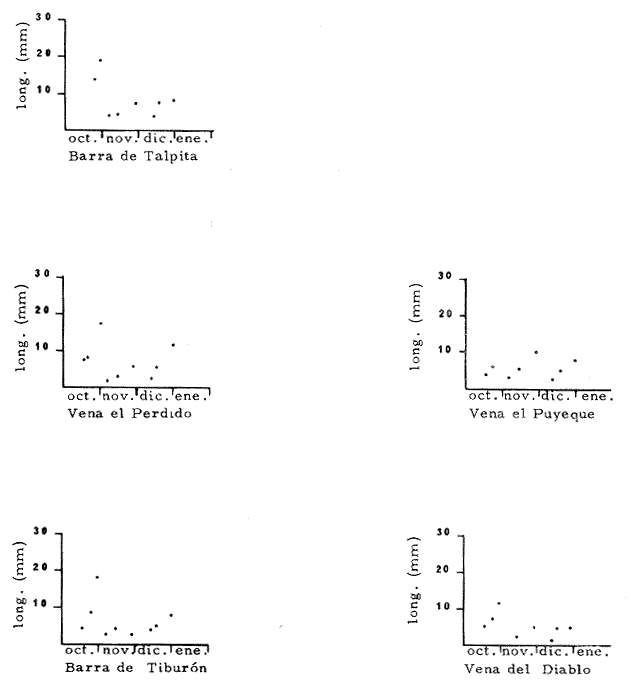 Fig. 8. Ritmo semanal-mensual de crecimiento máximo en juveniles de C. corteziensis provenientes de coectores instalados en los esteros del Pozo y del Rey, San Blas Nay. CRECIMIENTOUno de los aspectos más importantes de la biología de una especie comercial es la determinación de su velocidad de crecimiento en relación a su potencial reproductivo. Es conocido que en las ostras de zonas tropicales y subtropicales la madurez sexual es alcanzada ya con sólo algunos meses de, edad, y aparentemente C. corteziensis no hace excepción a esta generalización, ya que el estudio histológico demostró madurez en ejemplares de 45 mm, apenas con 3 o 4 meses de edad como se discute más adelante. Se ha mencionado en un capítulo anterior que la instalación de colectores tuvo como uno de los objetivos fundamentales el controlar épocas de fijación y crecimiento a partir de ejemplares obtenidos en ellos y medidos periódicamente por un número variable de meses. Para facilitar la discusión conviene agrupar las experiencias de acuerdo a tres tipos principales que se sucedieron en el tiempo. a) Medición periódica de tamaño en ejemplares de colectores que permanecieron sumergidos desde 4 hasta 9 meses. b) Medición mensual de tamaño en ejemplares provenientes de colectores y de raíz de mangle. c) Medición semanal de tamaño en ejemplares de colectores renovados mensualmente. Fig. 8 Ritmo semanl-mensual de cresimiento máximo en juveniles de C. corteziensis provenientes de colores instalados en los estereos del Pozo y del Rey, San Blas Nay. 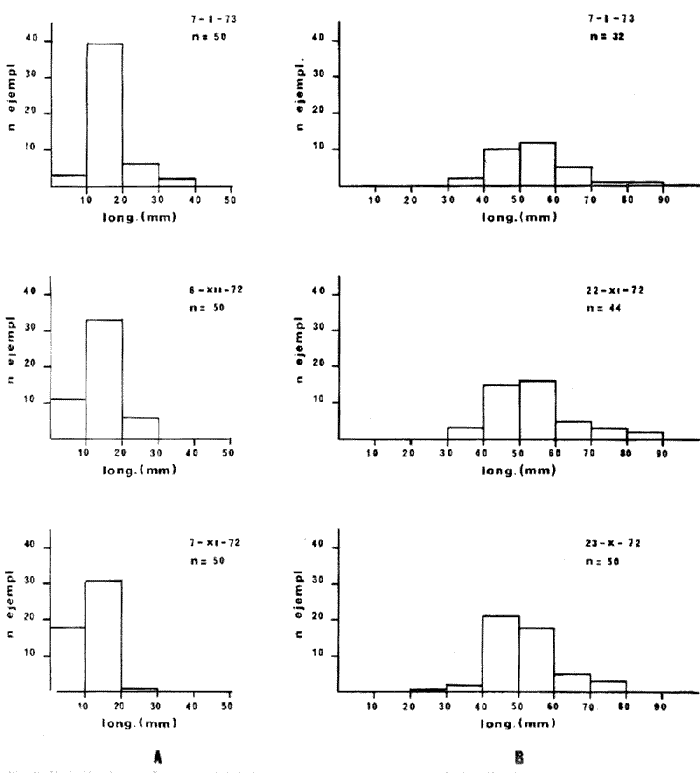 Fig. 9. Variación de tamaño y mortalidad de C. corteziensis en dos de muestras de San Blas, Nay., mantenidas como poblaciones artificiales. a) Medición periódica de tamaño en el ejemplares de colectores sumergidos 4 o más mesesEn el primer grupo de colectores instalados entre el 30 de noviembre de 1971 y el 9 de diciembre de 1971, se efectuaron mediciones de talla máxima y mínima de los ejemplares observados que se representan en la figura 7. Siendo el crecimiento de C. corteziensis relativamente similar en las distintas localidades en que se ubican los colectores (tabla 6) , se han combinado los valores máximos y mínimos representándose en una recta vertical por mes. Por dificultades ajenas al programa no pudieron efectuarse los controles de mayo y junio, sin embargo, la unión de los puntos máximos obtenidos en los meses restantes demuestra una velocidad de crecimiento que puede representarse por una curva cuya forma, al menos inicialmente, aparece corroborada por controles complementarios. 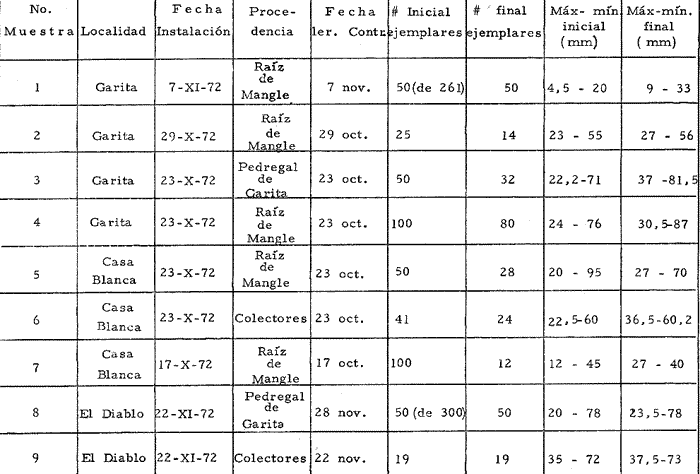 TABLA 4 CONTROL DE CRECIMIENTO MÁXIMO Y MORTALIDAD EN MUESTRAS PROVENIENTES DE MANGLAR, ROCAS O COLECTORES E INSTALADAS EN CAJAS OSTRÍCOLAS EN TRES LOCALIDADES DIFERENTES DE SAN BLAS, NAY.: GARITA, CASA BLANCA Y EL DIABLO De acuerdo a estos controles el tamaño máximo alcanzado después de un mes llegaría a 20 mm (2 cm), al término de dos meses sería de 35 - 40 mm, y al término de tres meses de aproximadamente 50 mm; a los cuatro meses no sobrepasa de 60 - 65 mm, es decir, disminuye y luego la velocidad de crecimiento parecería reducirse considerablemente para alcanzar 70- 75 mm al sexto y séptimo mes. La figura 7 representa una segunda curva de crecimiento en la que los primeros valores corresponden a ejemplares de los colectores instalados el 2 y 3 de junio de 1972. Como en el caso anterior se han combinado las mediciones de los distintos colectores para determinar máximos y mínimos. Es importante recordar que las mediciones fueron al azar en diversos colectores y no siempre reflejan el máximo absoluto como puede constatarse en la recta vertical de septiembre. Las rectas a, b y c en esta curva representan a un lote de 19 ejemplares mantenidos en cajas de cultivo cuyo origen es impreciso, pero que medidos durante 3 meses parecen corroborar el notable aplanamiento de la curva de crecimiento al alcanzar los 70 a 75 mm de longitud. La disminución de crecimiento en este y en el anterior podría deberse, sin embargo, a otras causas, derivadas de su mantención en cajas de cultivo. 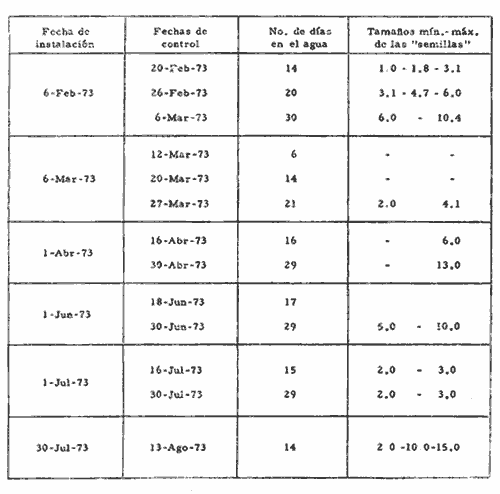 TABLA 5 FIJACIÓN Y CRECIMIENTO MÁXIMO-MÍNIMO DE C. CORTEZIENSIS EN COLECTORES DE TEJAS DE LOS ESTEROS DEL POZO Y DEL REY, SAN BLAS, NAY. (MEDIDAS BASADAS EN PROMEDIOS DE 5 O MÁS EJEMPLARES) Estos valores son comparables al crecimiento de C. virginica en aguas de Florida y de la costa de Texas y Alabama en el Golfo de México (Galtsoff, 1964; May. 1969). La misma figura incluye los valores máximos y mínimos de crecimiento obtenidos en dos muestras de 20 ejemplares marcados, provenientes de las localidades La Garita y Barra de Tiburón, respectivamente. Los valores d, e y f corresponden al aumento constatado en la primera localidad durante los meses de abril, mayo y junio de 1973 y los valores d1, e1 y f1 al aumento durante los mismos meses en la localidad Barra de Tiburón (Figs. 11 y 12). 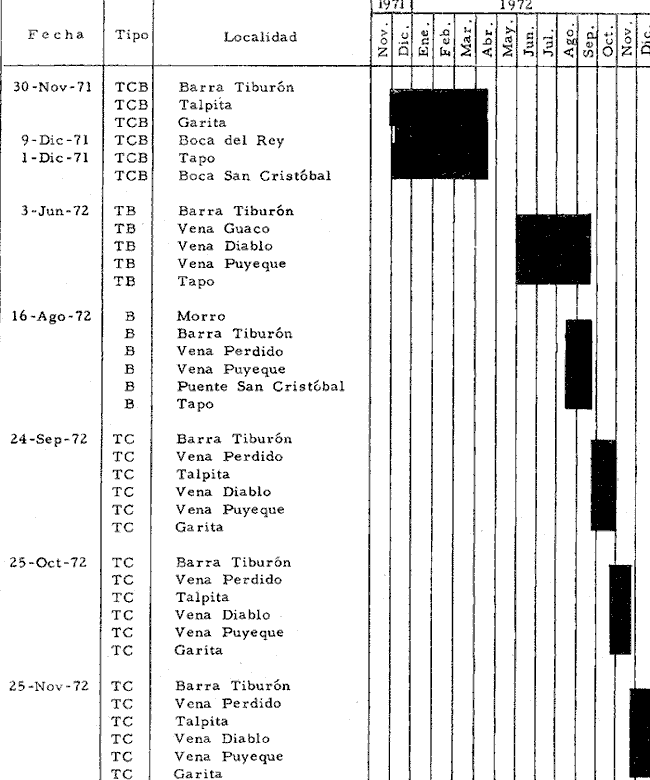 TABLA 6 LOCALIDA, TIPO Y FECHA DE INSTALACIÓN DE COLECTORES ENTRE 1971 Y 1972 EN SAN BLAS, NAY. Los últimos valores cuyos mínimos no fueron identificados corresponden a un crecimiento máximo en La Garita, en que la diferencia con respecto a los valores mínimos fueron considerablemente alterados por problemas de mortandad cercana al 50%, (Fig. 12). Aún así, plantea otra perspectiva de crecimiento, ya que demuestra que sobre 70 u 80 mm algunos ejemplares pueden alcanzar un incremento de tamaño mensual Igual a 10 mm. Se plantea así la posibilidad de que exista una curva más de crecimiento máximo representada en la figura 7 por una línea de trazos más finos. Este incremento es corroborado tanto en marcación individual de ejemplares, como ocurre en el caso de la misma última muestra representada en esta figura, o por muestreos al azar en poblaciones naturales (o seminaturales) discutidas más adelante. b) Medición mensual de tamaño en ejemplares provenientes de colectores y de raíces de mangleLa tabla 4 muestra un total de 9 controles de crecimiento cada uno con un número variable de ejemplares mantenidos en cajas ostrícolas de crecimiento. Los ejemplares de cada muestra fueron medidos semanalmente durante dos meses con objeto de constatar su crecimiento. En el tratamiento estadístico de estos controles debió descartarse la mayoría por problemas de mortalidad. La variación de tamaño experimentada con las muestras 1 y 3 procedentes de La Garita es representada en la figura 9, A, B. La variación de tamaño constatada en la muestra N° 9 ha sido representada en la figura 7, a, b, c. La representación de la variación mensual de los 50 ejemplares de la muestra 1 (tomados al azar de un total de 261, de tamaño pequeño) agrupados en clases de 10 mm (Fig. 9 A) demuestra el desplazamiento de las clases a un ritmo que refleja un crecimiento rápido mensual mínimo de 10 mm. No se constató mortalidad. La figura 9 B corresponde a una muestra total de 50 ejemplares de mayor tamaño, controlados también mensualmente, pero donde la mortalidad fue del 36%. El desplazamiento, de las clases de tamaño sobre todo en los extremos, es notorio en un ritmo equivalente al observado en la figura 9 A. Sin embargo, la mortalidad experimentada en el total es considerable ya al tercer mes, afectando especialmente a los ejemplares de mayor tamaño. Las causas son imprecisas. Conociendo de antemano la escasa representatividad de los controles con un número reducido de individuos respecto de la dinámica de las poblaciones del área en general, se programó un nuevo grupo de experiencias a partir de 4 poblaciones de 2 000 ejemplares cada una, recolectadas entre el 9 y el 13 de abril de raíces de mangle. Estas poblaciones fueron instaladas en cajas ostrícolas en 4 localidades diferentes, en las que se había observado previamente crecimiento óptimo: Barra de Tiburón y Estación 14 en el estero, del Rey y Vena del Diablo y La Garita en el estero del Pozo. El crecimiento de estas poblaciones "artificiales" se controló mensualmente en una muestra al azar de 200 ejemplares durante un periodo de 5 meses y los resultados se presentan en la figura 10. Se constata que la mortalidad era ya elevada al tercer mes de control (junio de 1973), por una combinación de factores entre los cuales la predación y especialmente la sedimentación de partículas sobre la malla de la caja ostrícola que impedía la circulación del agua, fueron las más importantes. La predación se efectuó por juveniles de "jaibas" (Callinectes spp.) que entraban a la caja en estados larvales planctónicos y se desarrollaban quedando aprisionadas en el interior de las cajas. De mayor importancia es sin embargo el hecho que durante todos los meses muestreados aparecen dos o tres nuevas clases a partir del muestreo original. Éste es el caso de la "población" de La Garita en donde aparecen ejemplares de las nuevas clases de 80 y 100 mm en el tercer mes y de la "población" de Barra de Tiburón en donde aparecen tres nuevas clases de 70, 80 y 90 mm a fines del primero, segundo y tercer mes respectivamente. El desplazamiento mensual de las clases restantes hacia longitudes mayores es también obvio prácticamente en todos los polígonos de frecuencia, corroborando que el crecimiento mensual por lo menos en parte de la población es de alrededor de 10 mm por mes, aun en las clases de mayor tamaño. Se constata también que la mayor mortalidad afecta principalmente a estas últimas. Mientras no se cambió a malla metálica de mayor tamaño, la mortalidad fue progresiva. Considerando que en definitiva la única manera de corroborar las velocidades de crecimiento sugeridas para ejemplares mayores de 30 o 40 mm. era mediante el control de crecimiento individual, se llevó a cabo una nueva serie de experiencias a partir de abril de 1973, durante tres meses consecutivos, que consideró a 20 ejemplares marcados en cajas de crecimiento ubicadas en las mismas cuatro últimas localidades nombradas: Barra de Tiburón, Estación 14, La Garita y Vena del Diablo. Los resultados de crecimiento individual se representan en las figuras 11 y 12. En las mismas se incluyen los promedios y el promedio de crecimiento para el total de ejemplares vivos en cada medición. Los números incluidos dentro de algunas de las columnas indican el mes en que murió el ejemplar respectivo. 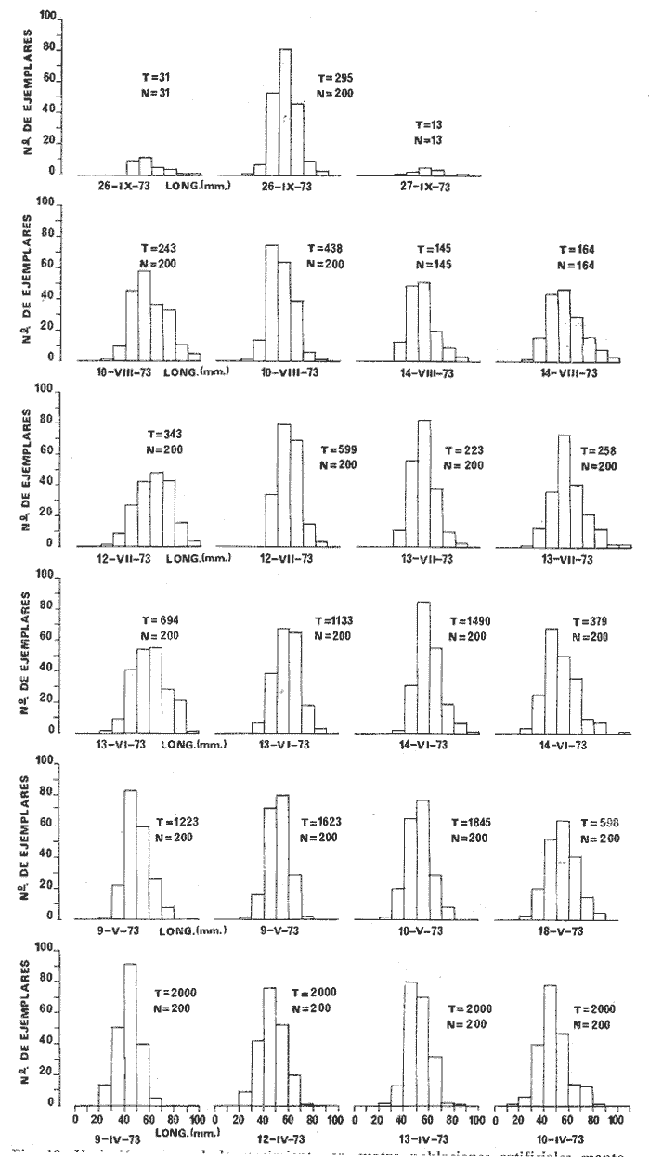 Fig. 10. Variación mensual de crecimiento en cuatro poblaciones artificiales mantenidas en cajas ostrícolas sumergidas en San Blas, Nay. 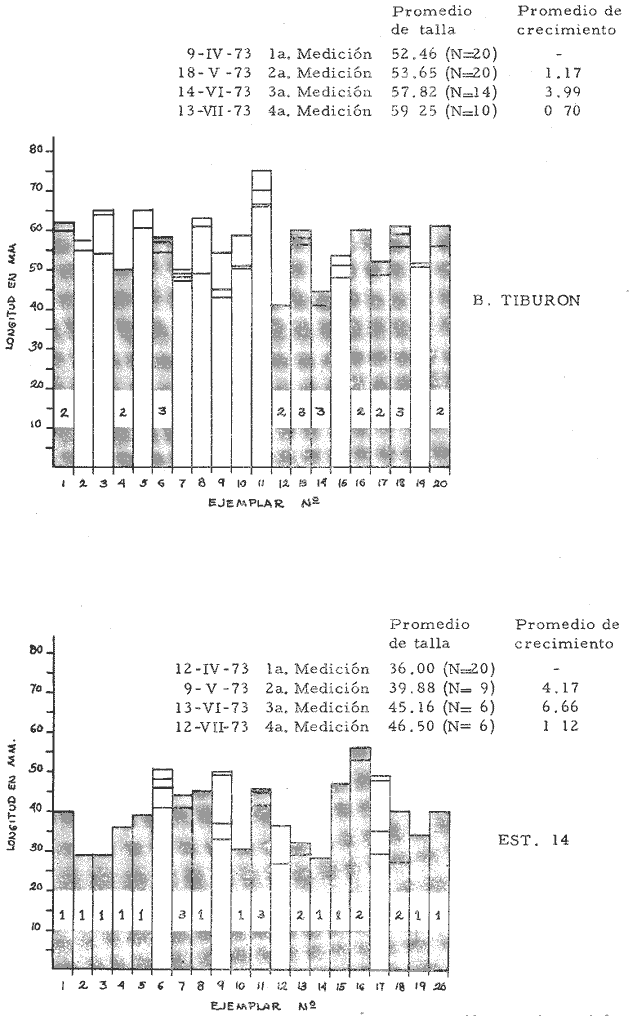 Fig. 11. Crecimiento individual mensual de dos muesras mantenidas en cajas ostrícolas sumeridas en dos localidades del estero del Rey, San Blas, Nay. 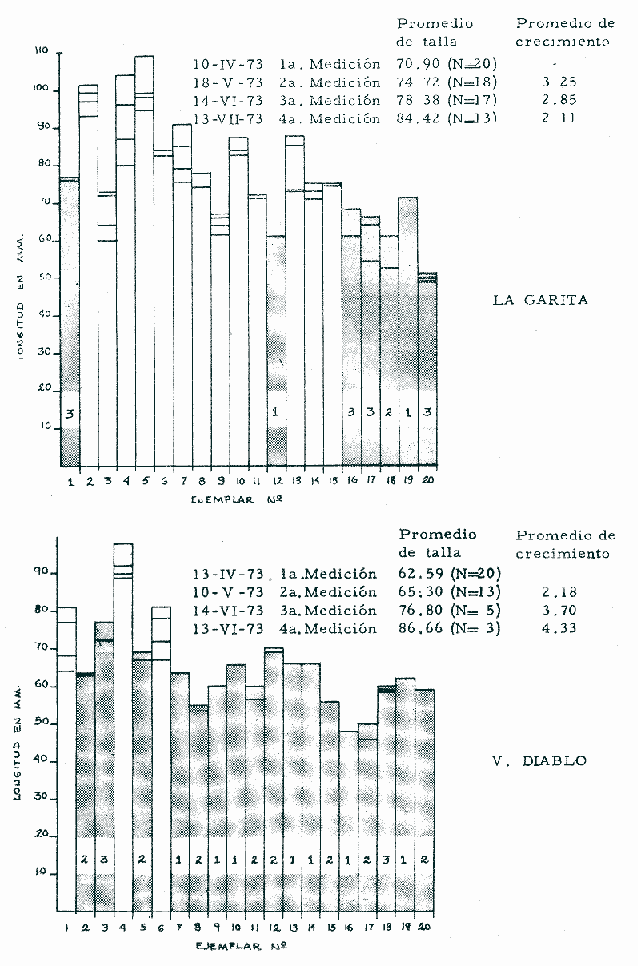 Fig. 12. Crecimiento individual en dos muesras mantenidas en cajas ostrícolas sumergidas en dos localidades del estero del Pozo, San Blas, Nay. Aceptando que en cada medición puede haber un error de 1 a 1.5 mm en el uso del vernier y que la mortalidad fue muy grande ya a fines del primer mes en la Estación 14 y Vena del Diablo, se constata: a) Que el crecimiento individual es muy variable en todas las muestras, habiendo ejemplares que en un momento dado crecen 1 o 2 mm y otros que crecen 10 o más mm. b) Que en consecuencia los promedios de crecimiento (Figs. 11 y 12) son también muy variables. c) Que, sin embargo, los ejemplares entre 25 y 45 mm de longitud inicial (Fig. 11, Est. 14) tienen tendencias a crecer más rápido que los de mayor longitud, con promedios de crecimiento de 4.17 a 6.66 mm durante el primero y segundo mes. d) Que hay también ejemplares con longitudes de más de 50 mm que pueden crecer igualmente 10 o más milímetros por mes, como se demuestra en las localidades de Barra de Tiburón, La Garita y Vena del Diablo (Figs. 11 y 12). En general, desconociéndose cuáles son las causas precisas que influyen en el crecimiento individual y general de cada muestra en las distintas localidades, no puede calcularse una curva general de crecimiento para todas las poblaciones de San Blas, ni menos una individual para cada localidad estudiada, sobre todo considerando que la mortalidad producida en algunas muestras, casi siempre por error humano no lo permite. En consecuencia, se ha preferido postular sólo curvas tentativas de crecimiento máximo (Fig. 7) susceptibles de precisarse en estudios posteriores. c) Medición semanal de tamaño en ejemplares de colectores renovados mensualmenteCon objeto de corroborar la curva de crecimiento del primer mes de vida obtenida de los colectores instalados en noviembre-diciembre de 1971 y estimar la densidad de fijación en los últimos meses del año 1972, se instalaron el 24 de septiembre, 25 de octubre y 25 de noviembre de este mismo año, los tipos de colectores ya mencionados (tabla 6) en cinco localidades distintas. La figura 8 representa los resultados obtenidos en las últimas tres semanas, después de su fijación (recuérdese que durante los primeros 7 a 10 días la "semilla" es tan pequeña que su medición en el terreno es imposible de realizar). En todos los casos se han combinado los tamaños máximos obtenidos, ya sea en el colector de tejas o de conchas, no existiendo diferencias apreciables en el crecimiento observado en uno u otro caso. Los límites de crecimiento constatados en Barra de Talpita, Vena El Perdido y Barra de Tiburón corresponden muy bien con la primera porción de las curvas postuladas (Fig. 7) alcanzando en un mes casi a los dos centímetros. Las gráficas siguientes indican valores inferiores que podrían reflejar condiciones de crecimiento no muy favorables o de fijaciones tardías con respecto a la instalación del colector. Es indudable que los valores sobre velocidad de crecimiento para cada mes dependerán del momento en que las larvas se fijen y nos parece que éste es el caso en los meses de noviembre y diciembre, ya que prescindiendo del periodo de la fijación en todas las localidades, la magnitud de crecimiento es muy similar. Es decir, en el momento dado (el de la instalación de colectores) la fijación dependerá de si hay o no larvas en primer lugar y luego de si las larvas están a punto de sufrir la metamorfosis. La importancia de otros factores tales como porcentaje de ejemplares en desove, distribución de larvas en la masa de agua por efecto de corrientes, etcétera, es en este momento imposible de considerar. Es posible que la predación sobre todo por los decápodos y peces que habitan en el área puedan en determinado momento alterar los índices de crecimiento, como se discute también más adelante. Aun aceptando todas las variables favorables o desfavorables posibles de imaginar debe concluirse que para los esteros estudiados puede garantizarse un aporte mensual continuo de juveniles en los meses de octubre, noviembre y diciembre que medirán entre 10 y 20 mm (1 a 2 cm) de longitud. De todo lo anterior puede concluirse que el crecimiento de C. corteziensis en el área estudiada ha demostrado ser rápido sobre todo en los primeros dos meses, alcanzando 70-75 mm de largo en, aproximadamente, 7 meses. Se ha constatado también que al menos para algunas poblaciones, la curva de crecimiento parece experimentar un aplanamiento notable después de los 70 a 75 mm de longitud. Se ignora si este aplanamiento es una tendencia general de las poblaciones, o si el crecimiento promedio de ellas puede continuar siendo relativamente rápido como se ha observado en diversos ejemplares marcados (Figs. 11 y 12) y en las curvas de crecimiento en muestras al azar (Fig. 7). La única información publicada hasta ahora sobre el crecimiento de C. corteziensis es la proporcionada por Sevilla (1959) para poblaciones de esta especie de Guaymas, Son. Según esta autora "ostiones" de colectores presentaron después de 10 meses una longitud promedio de 55.6 mm con máxima de 75 y mínima de 34 mm. La longitud promedio de los ejemplares de un año colectadas en los bancos fue de 52.5 mm. En la figura 7 hemos representado el crecimiento máximo conocido para los ejemplares de Guaymas, Son., comparados a los resultados obtenidos en San Blas, Nayarit. Ramírez y Sevilla (1965) en una discusión comparativa del crecimiento de ostras de diferentes especies en distintos países indican que en Guaymas, Son. C. corteziensis alcanzaría una talla promedio de 55 mm en 1 año, de 75 mm en 2 años y de 125 mm en tres años. La última cifra resulta extraña, ya que implicaría una velocidad de crecimiento proporcionalmente mayor durante el tercer año que durante el segundo. El crecimiento mínimo observado en C. corteziensis, por Sevilla (1959) en Guaymas, Son., parece relativo, ya que puede corresponder a individuos que se han fijado con algunas semanas si no meses de posterioridad respecto de los ejemplares de mayor tamaño. Sólo el control de tamaño en individuos marcados provenientes de una fijación que ha tenido lugar en un momento determinado, puede darnos la variación de crecimiento individual en una población. Mediciones de muestras al azar incluyen a clases de tamaño con fechas de fijación diferentes hasta en 4 o 5 meses dependiendo de la extensión del periodo de desove en la especie que se estudia. La velocidad de crecimiento indicada para C. corteziensis en San Blas, Nay. parece ser superior a la constatada en Guaymas, Son. correspondiendo a un principio fisiológico, ya conocido según el cual una especie crece más rápidamente en promedios de temperatura más elevados. En San Blas, Nay. la temperatura media mensual-anual varió a fines de 1971 entre 25 y 31°C y en 1972-1973 entre 23 y 32ºC (Figs. 1, 2, 3 y 6) indudablemente más elevada que en Guaymas, Son. (véanse gráficas núms. 2 y 4 en Sevilla, 1959). Depredación y otros factores ambientales negativosEntre los factores ambientales que afectan a las poblaciones de ostras, aparte de los factores abióticos ya discutidos los más importantes entre los que las afectan negativamente, incluyen: sedimentación, contaminación, enfermedades, comensales, competidores y depredadores. Se ha mencionado que la sedimentación en los periodos de mayor transporte de sedimento (periodo de lluvias; grandes mareas) no representa aparentemente un problema a las poblaciones naturales de C. corteziensis que habitan las raíces de mangle. Sí lo puede ser para las poblaciones que pudieran habitar los fondos duros, en aguas tranquilas, aunque durante todo el estudio no se constataron poblaciones de este tipo en ninguno de los esteros estudiados. Problemas de contaminación detectables en la supervivencia de las poblaciones del "ostión de placer" y de la fauna y flora locales no han sido observadas y de existir, deben ser mínimos. Tampoco se han observado comensales y la fauna de protozoos determinada en estudios preliminares correspondió a especies de los géneros Hexamita y Bodo y a la especie Halteria grandinella, presentes en infestaciones parciales del tracto digestivo y gónadas (determinaciones que agradecemos al Dr. E. López Ochoterena, Laboratorio de Protozoología, Fac. de Ciencias, UNAM). Entre las especies que representan mayores problemas competitivos, sobre todo al asentamiento de larvas, la más importante es el cirrípedo Balanus (M.) tintinnabulum. No se ha determinado la importancia temporal de hidrozoos o algas, ni tampoco la de organismos perforantes como algunas especies de esponjas, bivalvos y poliquetos. De gran importancia, sin embargo, es la depredación efectuada por jaibas del género Callinectes y en especial por un pez, el "botete" Sphaeroides annulatus (Jenyns, 1842) y el hombre. De gran interés fue el constatar a lo largo del estudio que la depredación era obvia sólo a partir de 1 1/2 a 2 cm, es decir que es selectiva. En tamaños menores la abundancia de C. corteúensis es considerable durante gran parte del periodo de reproducción. Sólo cuando el "ostión" alcanza cerca de los dos centímetros de tamaño, es cuando el "botete" empieza a depredar sobre las poblaciones, desprendiendo a los juveniles a mordidas que incluyen aun a trozos de la raíz de mangle. Es recién a ese tamaño que este depredador parece darse cuenta de la presencia del ostréido. Esto es quizás resultado de un acondicionamiento natural que ayuda a garantizar la supervivencia de las poblaciones como lo es también el caso del hombre que elige a ejemplares sólo de mayor tamaño, desdeñando a los juveniles. Si aparte de la competencia intraespecífica existe mortalidad importante por depredación en tamaños menores a los 2 cm es poco notoria y es probablemente "absorbida" por las poblaciones, dada la gran cantidad de juveniles existentes. AgradecimientosLos autores expresan sus agradecimientos a las autoridades de la Dirección de Acuacultura, Secretaría de Recursos Hidráulicos y a Agustín Ayala-Castañares, actual Coordinador de Ciencias de la Universidad Nacional Autónoma de México, por el financiamiento y las facilidades otorgadas para la realización y desarrollo de la investigación. Asimismo, hacen extensivos estos agradecimientos a las siguientes personas: Francisco Escobar de la Llata, Patricia Rogers, Armando Rodríguez Villamar y Daniel Méndez Quero por la ayuda de laboratorio y especialmente por su colaboración en los trabajos de campo. Finalmente apreciamos la ayuda secretarial de la señorita Susana Arellano. LITERATURACURRAY, J. R., F. J. EMMEL y P. J. CRAMPTON. Mem. Simp. Intern. Lagunas Costeras. Lagunas Costeras, un simposio. Holocene history of a strand plain, lagoonal coast, Nayarit, Mexico. In: Ayala-Castañares, A. y F. B. Phleger (Eds.) UNAM-UNESCO, 1969. 63-100. Nov. 28-30, 1967: GALTSOFF, P. S., The american oyster Crassostrea virginica Gmelin Fishery Bull. Fish Wildl. Serv.Wash., 1964. 1480. 64: KEEN, A. M., Sea shells of tropical West America. Stanford Univ. Press. 1971. 1064 p. KORRINGA, P., Q. Rev. Biol., Recent advances in oyster biology. 1952. 666-308 y 339-365. 27: Water temperature and breeding throughout the geographical range of Ostrea edulis. Année biol., 1957. 1-17. 33: LOOSANOFF, V. L., The American or eastern oyster. USDI, The American or eastern oyster. USDI 1965. 1-36. ,205: LOOSANOFF, V. L., H. C. DAVIS y P. E. CHANLEY, Malacologia, Dimensions and shapes of larvae of some marine bívalve mollusks. 1966. 351-435. 4 (2): MAY, E. B., Alabama Mar. Res. Bull Feasibility of off bottom oyster culture in Alabama.. 1969. 1-14. (3): OLSSON, A. A., Mollusks of the tropical eastern Pacific, partícularly from the southern half of the Panamic Pacific faunal province (Panama to Peru). Panamic-Pacific Pelecypoda. Paleontological Research Institution, Ithaca, N. Y. 1961. 574 p. RAMÌREZ, H. E. y M. L. SEVILLA, Las ostras de México. Instituto Nacional de Investigaciones Biolópesqueras, México, 1964. 1-100. 7: ROCHEBRUNE, A. T. de, Diagnoses de mollusques nouveaux, provenant du voyage de M. Diguet en Basse Californie. Bull Mus. Hist. nat., Paris, 1895. 239-243. 1: SEVILLA, M. L., Datos biológicos para el cultivo del ostión de Guaymas, Sonora. Secretaría de Industria y Comercio. Dirección General de Pesca, México, 1959. 87 p. STAFFORD, J., On the recognition of bivalve larvae in plankton collections. Contr. Cand. Biol. Fish. 1912. 221-242. 1906-1910: Rep. Comm. Fish. B. C. for the year ending The native oyster of British Columbia (Ostrea lurida Carpenter). 1914. 79-102. 31 Dec. 1913: STENZEL, H. B., Treatise on Invertebrate Paleontology. Oysters In: R. C. Moore (Ed.), Part N, 3 Mollusca 6 (Bivalvia) 1971. IV 272 p. STUARDO, J., A. MARTÍNEZ y F. ESCOBAR DE LA LLATA, Informe final. Contrato de Estudios No. EI71-11, Clave LL-3. Informe final de la Segunda Etapa de los estudios sobre la influencia de los cambios hidrológicos del Río Santiago en la biología del ostión y medidas para la rehabilitación de los bancos de dicha especie. Inst. Biol. Univ. Nal. Autón. México, Secretaría de Recursos Hidráulicos, México, 1973. 76 p. THOMSON, J. M., Aust. J. mar. Freshwat. Res., The genera of oysters and the Australian species. 1954. 132-168, láms. 1-11. 5 (1): WALNE, P. R., Observations on the fertility of the oyster (Ostrea edulis). J. mar. biol. Ass. U. K., n. s. 1964. 293-310. 44: LAMINAS
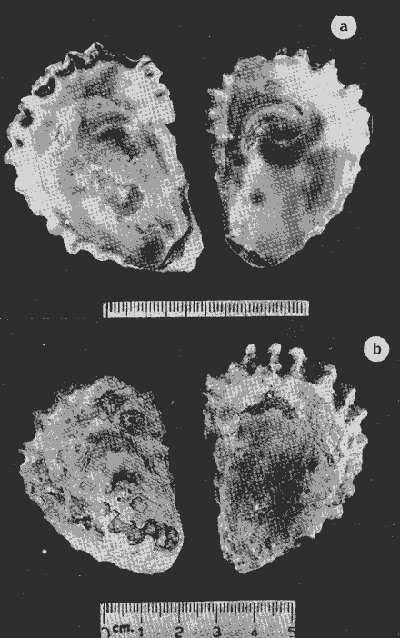 Lám. 1 Vista interna (a) y externa (b) de valvas de un ejemplar de Crassostreacorteziensis (Hertiein, 1951). 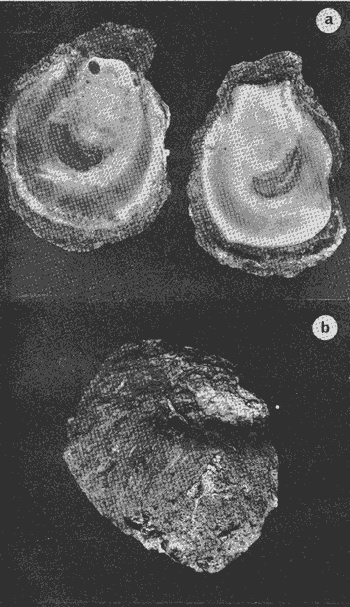 Lám. 2 Vista interna (a) y externa (b) de las valvas de un ejemplar de Crassostrea iridescens (Hanley, 1854). Ejemplres de ca. 200 mm de longitud. 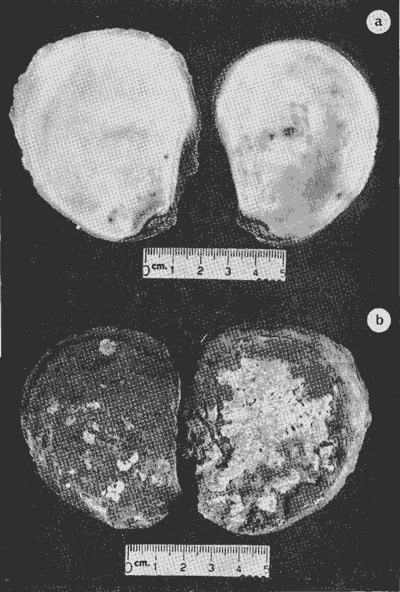 Lám. 3 Vista interna (a) y externa (b) de las valvas de un ejemplar de Cassostrea palmula (Carpenter, 1857).  Lám.4 a) Larvas "padiveliger" de C. corteziensis provenientes de muestras de placton de la Garita San Blas, Nay. (18-X-72) ca. 320 micras. b) Colector de tejas  Lám. 5 a) Corral experimental. b) Cajas utilizadas para control de cresimiento y engorda. Nótese la separación de muestras por separación de muestras por separaciones.
|